
Maize-Pathogen Interactions: An Ongoing Combat from a Proteomics Perspective

Abstract
:1. Introduction
2. Maize-Pathogen Interactions from a Proteomics View
2.1. Maize vs. Aspergillus Flavus and Aflatoxins
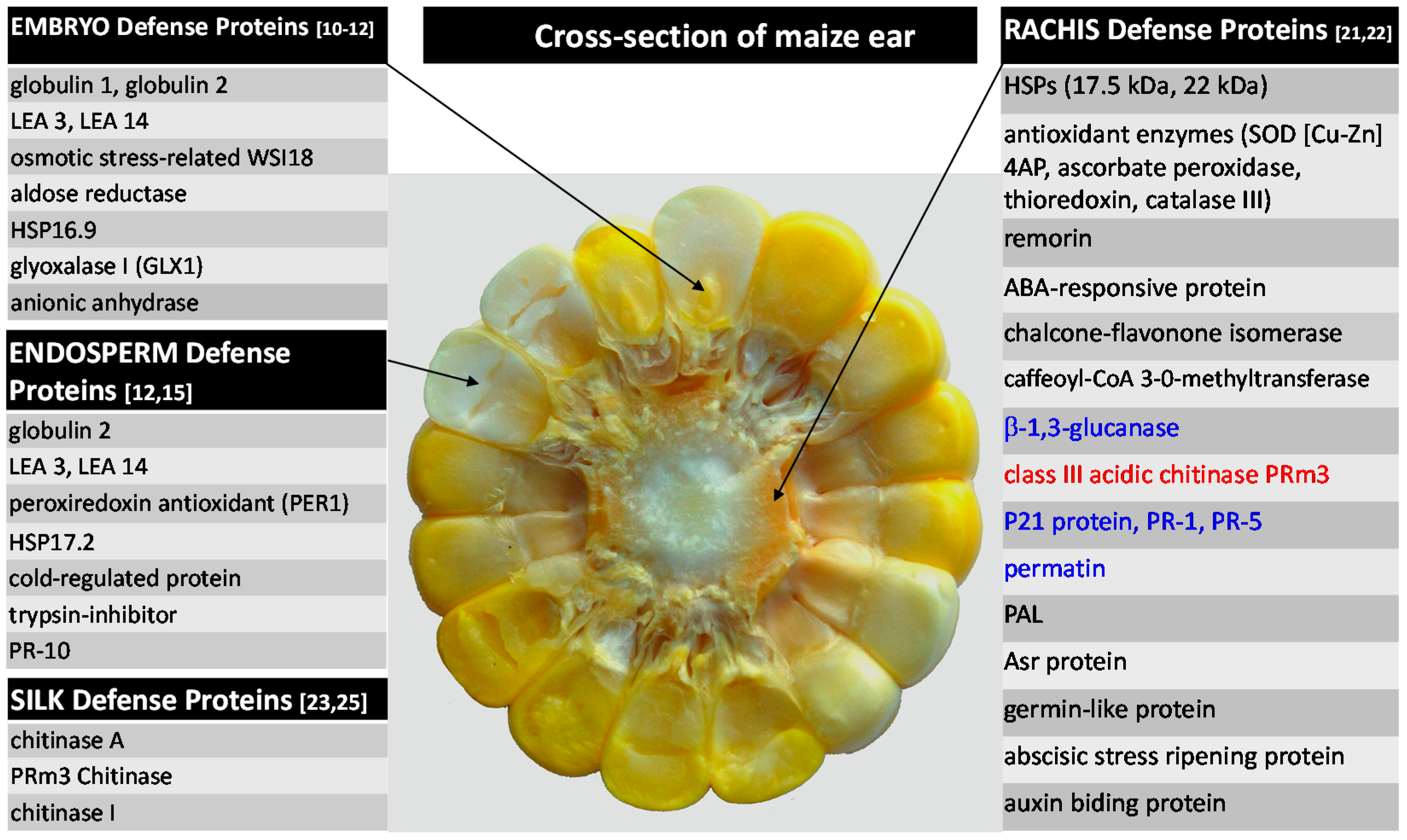
2.1.1. Kernel Resistance
2.1.2. Rachis Resistance
2.1.3. Silk Resistance
2.1.4. Summary of Maize Protein-Based Defenses against A. flavus

2.2. Maize vs. Fusarium Pathogens
2.2.1. F. graminearum and Gibberella Ear Rot
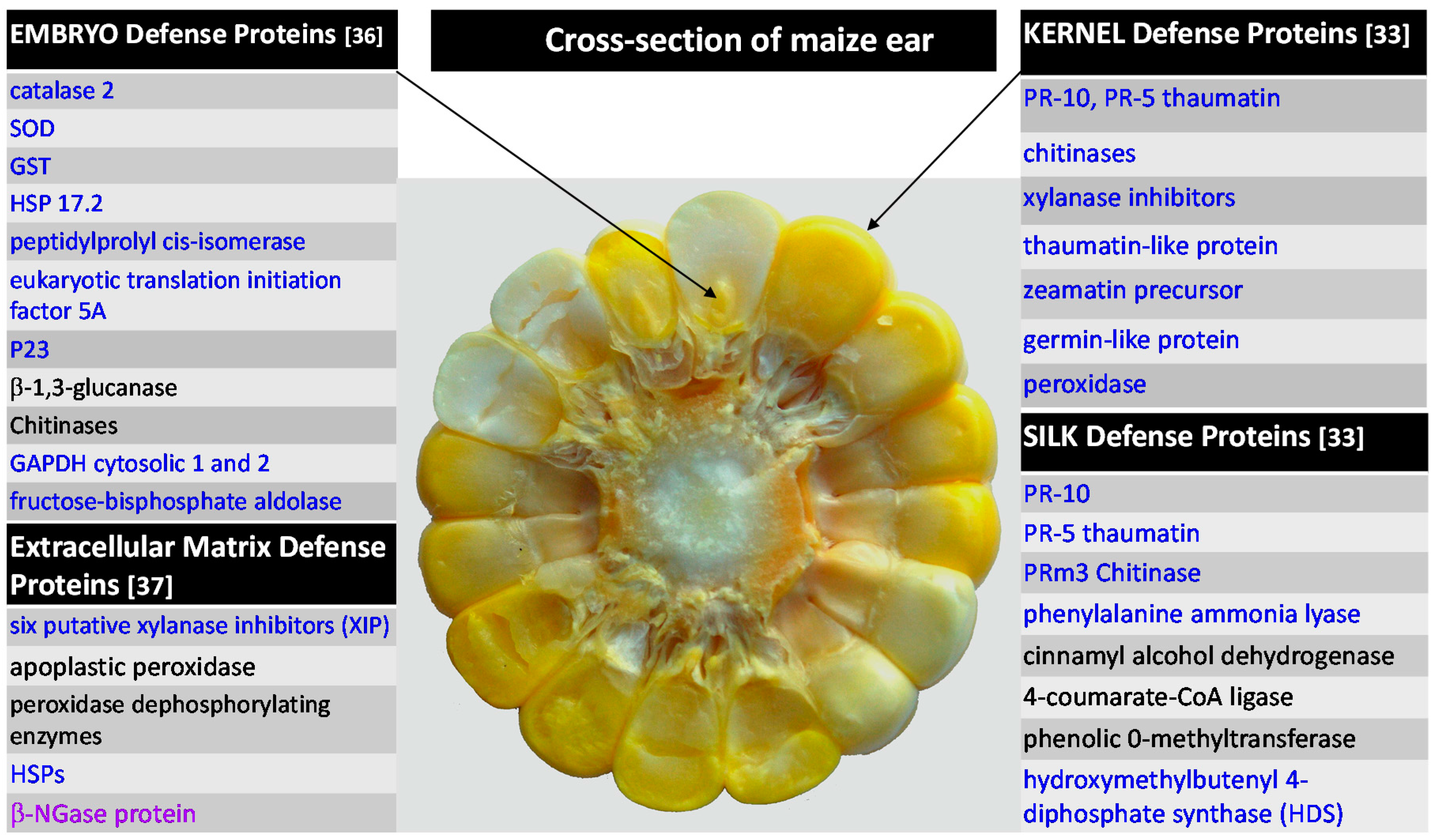
2.2.2. F. verticillioides and Fusarium Ear Rot
2.2.3. F. verticillioides and Extracellular Matrix
2.2.4. Summary of Maize Protein-Based Defenses against Fusarium spp.

2.3. Maize vs. Curvularia lunata
2.4. Maize vs.Viral Pathogens
2.4.1. Rice Black-Streaked Dwarf Virus
2.4.2. Sugarcane Mosaic Virus

{kind=link}
{kind=link}
| Pathogen | Maize Genotype | Induced Proteins | Repressed Proteins | Reference |
|---|---|---|---|---|
| Black-Streaked Dwarf Virus | Ye478 Susceptible | Peroxidase 39, APx2-cytosolic ascorbate peroxidase, catalase 3, oxygenase, caffeic acid 3-O-methyltransferase, cinnamyl alcohol degydrogenase, UDP-glucose pyrophosphorylase, GAPDH, ADP-glucose phosphorylase, lipoxygenases, GTP-binding proteins, enzymes of starch synthesis, hotosynthesis, carbon fixation and assimilation | Enzymes of glycolysis, TCA cycle, glycogenesis, and pentose phosphate metabolism, particularly transketolase | [58] |
| Sugarcane Mosaic Virus | Siyi Resistant | Enzymes of glycolysis and gluconeogenesis, remorin, cysteine synthase, glutamate dehydrogenase, calcium-dependent protein kinase, bZIP transcription factor ABI5, serine/threonine-protein kinase, nucleolar RNA helicase 2, 30S ribosomal protein S5 | [56,57] | |
| Sugarcane Mosaic Virus | Mo17 Susceptible | Enzymes of glycolysis and gluconeogenesis, remorin, cysteine synthase, ATP synthase CF1 α subunit | [56,57] | |
| Sugarcane Mosaic Virus | Siyi and Mo17 | Stress/defense-related and signal transduction/transcription-associated proteins,aconitase, transketolase,nucleoside diphosphate kinase, abscisic stress ripening protein, chaperonin, T-complex protein, ferredoxin-NADP reductase, histidine triad nucleotide binding protein, β-d-glucosidase precursor, electron transporter protein | Photosynthesis-associated proteins, thioredoxin, glutathione S-transferase, RuBisCo large subunit, RuBisCo subunit binding protein α subunit | [56,57] |
3. Commonly Responsive Proteins in Maize-Pathogen Interaction
3.1. Pathogenesis–Related Proteins
| Protein Group | Elicited by Pathogen | Tissue | Proteins | References |
|---|---|---|---|---|
| Pathogen related proteins | Af, Fv, Fg | Embryo, silk, rachis, kernels | Chitinase, glucanase, trypsin and amylase inhibitor, peroxidase | [14,22,23,33,36] |
| Detoxifying enzymes | Af, Fv, Fg, Cl, RBSDV, SCMV | Leaves, foliage, embryo, silk, rachis, kernels | SOD, catalase, PER, thioredoxin, glutaredoxin, glutathione reductase, GST, dehydroascorbate reductase | [12,22,33,36,46,56,57,68] |
| Proteins involved in secondary metabolism | Af, Fv, Fg, RBSDV, SCMV | Rachis, leaves, ear | PAL, caffeoyl-CoA 3-O-methyltransferase, chalcone-flavonone isomerase, cinnamyl alcohol dehydrogenase, hydroxymethyl-butenyl 4-diphosphate synthase | [22,56,58] |
| Proteins involved in energy producing pathways | Af, Fv, RBSDV, SCMV | Rachis, leaves | GAPDH, ADP-glucose pyro-phosphorylase, UDP-glucose pyrophosphorylase, glucose phosphate isomerase, fructose-bisphosphate aldolase, transketolase | [22,33,36,37,47,56,57,58] |
| Proteins involved in protein synthesis, folding and stabilization | Af, Fv, Fg | Embryo, rachis, kernel | elF-5A/initiation of translation; HSPs, chaperonins, peptidylprolyl cis-trans isomerase, cyclophilin | [22,33,36,37,46,47,56] |
3.2. Detoxifying Enzymes
3.3. Proteins Involved in Secondary Metabolism
3.4. Proteins Involved in Energy-Producing Carbohydrate Metabolic Pathways
3.5. Proteins Involved in Protein Synthesis, Folding and Stabilization
4. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Castegnaro, M.; McGregor, D. Carcinogenic risk assessment of mycotoxins. Rev. Med. Vet. 1998, 149, 671–678. [Google Scholar]
- McKean, C.; Tang, L.; Billam, M.; Tang, M.; Theodorakis, C.W.; Kendall, R.J.; Wang, J.S. Comparative acute and combinative toxicity of aflatoxin B1 and T-2 toxin in animals and immortalized human cell lines. J. Appl. Toxicol. 2006, 26, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Diener, U.L.; Davis, N.D. Aflatoxin formation by Aspergillus flavus. In Aflatoxin: Scientific Background, Control and Implications; Goldblatt, L.A., Ed.; Academic Press: New York, NY, USA, 1969; pp. 360–391. [Google Scholar]
- Payne, G.A. Aflatoxins in maize. Crit. Rev. Plant. Sci. 1992, 10, 423–440. [Google Scholar] [CrossRef]
- Schmale, D.G.; Munkvold, G.P. Mycotoxins in Crops: A Threat to Human and Domestic Animal Health. Available online: http://www.apsnet.org/edcenter/intropp/topics/Mycotoxins/Pages/default.aspx (accessed on 11 November 2015).
- Neal, G.E. Participation of animal biotransformation in mycotoxin toxicity. Rev. Med. Vet. 1998, 149, 555–560. [Google Scholar]
- Chen, Z.Y.; Brown, R.L.; Cleveland, T.E. Evidence for an association in corn between stress tolerance and resistance to Aspergillus flavus infection and aflatoxin contamination. Afr. J. Biotechnol. 2004, 3, 693–699. [Google Scholar]
- Chen, Z.Y.; Brown, R.L.; Cleveland, T.E.; Damann, K.E.; Russin, J.S. Comparison of constitutive and inducible maize kernel proteins of genotypes resistant or susceptible to aflatoxin production. J. Food Prot. 2001, 64, 1785–1792. [Google Scholar] [PubMed]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of unique or elevated levels of kernel proteins in aflatoxin-resistant maize genotypes through proteome analysis. Phytopathology 2002, 92, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of a maize kernel stress-related protein and its effect on aflatoxin accumulation. Phytopathology 2004, 94, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of maize kernel endosperm proteins associated with resistance to aflatoxin contamination by Aspergillus flavus. Phytopathology 2007, 97, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Brown, R.L.; Lax, A.R.; Guo, B.Z.; Cleveland, T.E.; Russin, J.S. Resistance to Aspergillus flavus in corn kernels is associated with a 14-kDa protein. Phytopathology 1998, 88, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Brown, R.L.; Rajasekaran, K.; Damann, K.E.; Cleveland, T.E. Identification of a maize kernel pathogenesis-related protein and evidence for its involvement in resistance to Aspergillus flavus infection and aflatoxin production. Phytopathology 2006, 96, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Brown, R.L.; Russin, J.S.; Lax, A.R.; Cleveland, T.E. A corn trypsin inhibitor with antifungal activity inhibits Aspergillus flavus α-amylase. Phytopathology 1999, 89, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.Z.; Chen, Z.Y.; Brown, R.L.; Lax, A.R.; Cleveland, T.E.; Russin, J.S.; Mehta, A.D.; Selitrennikoff, C.P.; Widstrom, N.W. Germination induces accumulation of specific proteins and antifungal activities in corn kernels. Phytopathology 1997, 87, 1174–1178. [Google Scholar] [CrossRef] [PubMed]
- Lozovaya, V.V.; Waranyuwat, A.; Widholm, J.M. B-1,3-glucanase and resistance to Aspergillus flavus infection in maize. Crop Sci. 1998, 38, 1255–1260. [Google Scholar] [CrossRef]
- Hong-Bo, S.; Zong-Suo, L.; Ming-An, S. Lea proteins in higher plants: Structure, function, gene expression and regulation. Colloids Surf. 2005, 45, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, Y. Response of Resistant and Susceptible Maize Genotypes to Inoculation with Transformed Aspergillus flavus Isolates. Ph.D. Thesis, Mississippi State University, Mississippi State, MS, USA, 1999. [Google Scholar]
- Magbanua, Z.V.; Williams, W.P.; Luthe, D.S. The maize rachis affects Aspergillus flavus spread during ear development. Maydica 2013, 58, 182–188. [Google Scholar]
- Pechanova, O.; Pechan, T.; Ozkan, S.; McCarthy, F.M.; Williams, W.P.; Luthe, D.S. Proteome profile of the developing maize (Zea mays L.) rachis. Proteomics 2010, 10, 3051–3055. [Google Scholar] [CrossRef] [PubMed]
- Pechanova, O.; Pechan, T.; Williams, W.P.; Luthe, D.S. Proteomic analysis of the maize rachis: Potential roles of constitutive and induced proteins in resistance to Aspergillus flavus infection and aflatoxin accumulation. Proteomics 2011, 11, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Peethambaran, B.; Hawkins, L.; Windham, G.L.; Williams, W.P.; Luthe, D.S. Anti-fungal activity of maize silk proteins and role of chitinase in Aspergillus flavus resistance. Toxin Rev. 2010, 29, 27–39. [Google Scholar] [CrossRef]
- Marsh, S.F.; Payne, G.A. Preharvest infection of corn silks and kernels by Aspergillus flavus. Phytopathology 1984, 74, 1284–1289. [Google Scholar] [CrossRef]
- Neucere, J.N. Inhibition of Aspergillus flavus growth by silk extracts of resistant and susceptible corn. J. Agric. Food Chem. 1996, 44, 1982–1983. [Google Scholar] [CrossRef]
- Naidoo, G.; Forbes, A.M.; White, C.P.; Rocheford, T.R. Resistance to Aspergillus flavus ear rot and aflatoxin accumulation in maize f1 hybrids. Crop Sci. 2002, 42, 360–364. [Google Scholar] [CrossRef]
- Paul, C.; Naidoo, G.; Forbes, A.; Mikkilineni, V.; White, D.; Rocheford, T. Quantitative trait loci for low aflatoxin production in two related maize populations. TAG Theor. Appl. Genet. 2003, 107, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.D.; White, D.G. Inheritance of resistance to Aspergillus ear rot and aflatoxin production of corn from ci2. Plant Dis. 2001, 85, 322–327. [Google Scholar] [CrossRef]
- Pechanova, O.; Takac, T.; Samaj, J.; Pechan, T. Maize proteomics: An insight into the biology of an important cereal crop. Proteomics 2013, 13, 637–662. [Google Scholar] [CrossRef] [PubMed]
- Fandohan, P.; Hell, K.; Marasas, W.F.O.; Wingfield, M.J. Infection of maize by Fusarium species and contamination with fumonisin in Africa. Afr. J. Biotechnol. 2003, 2, 570–579. [Google Scholar]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Woloshuk, C.; Wise, K. Diseases of Corn: Gibberella Ear Rot. Available online: http//www.extension.purdue.edu/extmedia/BP/BP-77-W.pdf (accessed on 3 September 2015).
- Mohammadi, M.; Anoop, V.; Gleddie, S.; Harris, L.J. Proteomic profiling of two maize inbreds during early gibberella ear rot infection. Proteomics 2011, 11, 3675–3684. [Google Scholar] [CrossRef] [PubMed]
- Marasas, W.F. Fumonisins: Their implications for human and animal health. Nat. Toxins 1995, 3, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Stockmann-Juvala, H.; Savolainen, K. A review of the toxic effects and mechanisms of action of fumonisin B1. Hum. Exp. Toxicol. 2008, 27, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Carrascal, M.; Coca, M.; Abian, J.; San Segundo, B. The defense response of germinating maize embryos against fungal infection: A proteomics approach. Proteomics 2004, 4, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Chivasa, S.; Simon, W.J.; Yu, X.L.; Yalpani, N.; Slabas, A.R. Pathogen elicitor-induced changes in the maize extracellular matrix proteome. Proteomics 2005, 5, 4894–4904. [Google Scholar] [CrossRef] [PubMed]
- Juge, N.; Payan, F.; Williamson, G. XIP-I, a xylanase inhibitor protein from wheat: A novel protein function. Biochim. Biophys. Acta 2004, 1696, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.M. Mutant of bacillus subtilis lacking Exo-β-N-acetylglucosaminidase activity. J. Bacteriol. 1974, 117, 909–910. [Google Scholar] [PubMed]
- Robertson-Hoyt, L.A.; Betran, J.; Payne, G.A.; White, D.G.; Isakeit, T.; Maragos, C.M.; Molnar, T.L.; Holland, J.B. Relationships among resistances to Fusarium and Aspergillus ear rots and contamination by fumonisin and aflatoxin in maize. Phytopathology 2007, 97, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Maschietto, V.; de Leonardis, S.; Battilani, P.; Paciolla, C.; Marocco, A. Defense responses to mycotoxin-producing fungi Fusarium proliferatum, F. subglutinans, and Aspergillus flavus in kernels of susceptible and resistant maize genotypes. Mol. Plant Microbe Interact. 2015, 28, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.A.; Nemchenko, A.; Park, Y.S.; Borrego, E.; Huang, P.C.; Schmelz, E.A.; Kunze, S.; Feussner, I.; Yalpani, N.; Meeley, R.; et al. The novel monocot-specific 9-lipoxygenase ZmLOX12 is required to mount an effective jasmonate-mediated defense against Fusarium verticillioides in maize. Mol. Plant Microbe Interact. 2014, 27, 1263–1276. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Brodhagen, M.; Isakeit, T.; Brown, S.H.; Gobel, C.; Betran, J.; Feussner, I.; Keller, N.P.; Kolomiets, M.V. Inactivation of the lipoxygenase ZmLOX3 increases susceptibility of maize to Aspergillus spp. Mol. Plant Microbe Interact. 2009, 22, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.F.; Paradela, F.; Soave, J.; Sugimori, M.H. Leaf spot caused in maize (Zea mays L.) by Culvularia lunata (wakker) boedijn. Summa Phytopathol. 1979, 5, 181–184. [Google Scholar]
- Liu, T.; Liu, L.X.; Huang, X.L.; Jiang, X.; Zhou, P.H. A new furanoid toxin produced by Culvularia lunata, the causal agent of maize Curvularia leaf spot. Can. J. Plant Pathol. 2009, 31, 22–27. [Google Scholar] [CrossRef]
- Huang, X.; Liu, L.; Chen, J.; Zhai, Y. Comparative proteomic analysis of the response in resistant and susceptible maize inbred lines to infection by Culvularia lunata. Prog. Nat. Sci. 2009, 19, 845–850. [Google Scholar] [CrossRef]
- Huang, X.; Liu, L.; Zhai, Y.; Liu, T.; Chen, C. Proteomic comparison of four maize inbred lines with different levels of resistance to Culvularia lunata (wakker) boed infection. Prog. Nat. Sci. 2009, 19, 353–358. [Google Scholar] [CrossRef]
- Bernier, F.; Berna, A. Germins and germin-like proteins: Plant do-all proteins. But what do they do exactly? Plant Physiol. Biochem. 2001, 39, 545–554. [Google Scholar] [CrossRef]
- Manosalva, P.M.; Davidson, R.M.; Liu, B.; Zhu, X.; Hulbert, S.H.; Leung, H.; Leach, J.E. A germin-like protein gene family functions as a complex quantitative trait locus conferring broad-spectrum disease resistance in rice. Plant Physiol. 2009, 149, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G.; Baumlein, H.; Mock, H.P.; Himmelbach, A.; Schweizer, P. The multigene family encoding germin-like proteins of barley. Regulation and function in basal host resistance. Plant Physiol. 2006, 142, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.B.; Thordal-Christensen, H.; Zimmermann, G.; Gjetting, T.; Lyngkjaer, M.F.; Dudler, R.; Schweizer, P. The germinlike protein GLP4 exhibits superoxide dismutase activity and is an important component of quantitative resistance in wheat and barley. Mol. Plant Microbe Interact. 2004, 17, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Lane, B.G. Oxalate, germins, and higher-plant pathogens. IUBMB Life 2002, 53, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Vallelian-Bindschedler, L.; Mosinger, E.; Metraux, J.P.; Schweizer, P. Structure, expression and localization of a germin-like protein in barley (Hordeum vulgare L.) that is insolubilized in stressed leaves. Plant Mol. Biol. 1998, 37, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Thomas, V.; Bennett, M.H.; Mansfield, J.; Grant, M. Modifications to the Arabidopsis defense proteome occur prior to significant transcriptional change in response to inoculation with pseudomonas syringae. Plant Physiol. 2006, 142, 1603–1620. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, K.; Hanagata, N.; Dubinsky, Z.; Baba, S.; Karube, I. Molecular characterization of cDNA encoding oxygen evolving enhancer protein 1 increased by salt treatment in the mangrove Bruguiera gymnorrhiza. Plant Cell Physiol. 2000, 41, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Han, Z.; Wang, S.; Wang, X.; Sun, A.; Zu, X.; Chen, Y. Comparative proteomic analysis of the plant-virus interaction in resistant and susceptible ecotypes of maize infected with sugarcane mosaic virus. J. Proteom. 2013, 89, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, S.; Chen, X.; Wang, X.; Wu, L.; Zu, X.; Chen, Y. Proteomic and phytohormone analysis of the response of maize (Zea mays L.) seedlings to sugarcane mosaic virus. PLoS ONE 2013, 8, e70295. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xu, C.; Zhang, J. Proteome profile of maize (Zea mays L.) leaf tissue at the flowering stage after long-term adjustment to rice black-streaked dwarf virus infection. Gene 2011, 485, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Von Rad, U.; Huttl, R.; Lottspeich, F.; Gierl, A.; Frey, M. Two glucosyltransferases are involved in detoxification of benzoxazinoids in maize. Plant J. 2001, 28, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Block, A.; Schmelz, E.; O’Donnell, P.J.; Jones, J.B.; Klee, H.J. Systemic acquired tolerance to virulent bacterial pathogens in tomato. Plant Physiol. 2005, 138, 1481–1490. [Google Scholar] [CrossRef] [PubMed]
- DebRoy, S.; Thilmony, R.; Kwack, Y.B.; Nomura, K.; He, S.Y. A family of conserved bacterial effectors inhibits salicylic acid-mediated basal immunity and promotes disease necrosis in plants. Proc. Natl. Acad. Sci. USA 2004, 101, 9927–9932. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K.; Melotto, M.; He, S.Y. Suppression of host defense in compatible plant-pseudomonas syringae interactions. Curr. Opin. Plant Biol. 2005, 8, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.J. Defense-related proteins in higher plants. Annu. Rev. Biochem. 1992, 59, 873–907. [Google Scholar] [CrossRef] [PubMed]
- Kasprzewska, A. Plant chitinases—Regulation and function. Cell. Mol. Biol. 2003, 8, 809–824. [Google Scholar]
- Leah, R.; Tommerup, H.; Svendsen, I.; Mundy, J. Biochemical and molecular characterization of three barley seed proteins with antifungal properties. J. Biol. Chem. 1991, 266, 1564–1573. [Google Scholar]
- Schlumbaum, A.; Mauch, F.; Vogeli, U.; Boller, T. Plant chitinases are potent inhibitors of fungal growth. Nature 1986, 324, 365–367. [Google Scholar] [CrossRef]
- Magbanua, Z.V.; de Moraes, C.M.; Brooks, T.D.; Williams, W.P.; Luthe, D.S. Is catalase activity one of the factors associated with maize resistance to Aspergillus flavus? Mol. Plant Microbe Interact. 2007, 20, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Vigers, A.; Roberts, W.; Selitrennikof, C. A new family of plant antifungal proteins. Mol. Plant Microbe Interact. 1991, 4, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.; Valdes-Rodrigues, S.; Blanco-Labra, A. A possible function for thaumatin and TMV-induced protein suggested by homology to a maize inhibitor. Nature 1987, 327, 432–434. [Google Scholar] [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Almagro, L.; Gomez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barcelo, A.; Pedreno, M.A. Class III peroxidases in plant defence reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Bolwell, G.P.; Wojtaszek, P. Mechanisms for the generation of reactive oxygen species in plant defence—A broad perspective. Physiol. Mol. Plant Pathol. 1997, 51, 347–366. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A large family of class III plant peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol. 2001, 42, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Kurama, E.E.; Fenille, R.C.; Rosa, V.E., Jr.; Rosa, D.D.; Ulian, E.C. Mining the enzymes involved in the detoxification of reactive oxygen species (ROS) in sugarcane. Mol. Plant Pathol. 2002, 3, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Scandalios, J.G. Two structurally similar maize cytosolic superoxide dismutase genes, Sod4 and Sod4A, respond differentially to abscisic acid and high osmoticum. Plant Physiol. 1998, 117, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J.; Jacob, S.; Oelze, M.L.; Laxa, M.; Tognetti, V.; de Miranda, S.M.; Baier, M.; Finkemeier, I. The function of peroxiredoxins in plant organelle redox metabolism. J. Exp. Bot. 2006, 57, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- Gutsche, N.; Thurow, C.; Zachgo, S.; Gatz, C. Plant-specific CC-type glutaredoxins: Functions in developmental processes and stress responses. Biol. Chem. 2015, 396, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Chugh, V.; Kaur, N.; Grewal, M.S.; Gupta, A.K. Differential antioxidative response of tolerant and sensitive maize (Zea mays L.) genotypes to drought stress at reproductive stage. Indian J. Biochem. Biophys. 2013, 50, 150–158. [Google Scholar] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant glutathione transferases. Genome Biol. 2002, 3. REVIEWS3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sari-Gorla, M.; Ferrario, S.; Rossini, L.; Frova, C.; Villa, M. Developmental expression of glutathione S-transferase in maize and its possible connection with herbicide tolerance. Euphytica 1993, 67, 221–230. [Google Scholar] [CrossRef]
- Timmerman, K.P. Molecular characterization of corn glutathione S-transferase isozymes involved in herbicide detoxication. Physiol. Plant 1989, 77, 465–471. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Chemical diversity and defence metabolism: How plants cope with pathogens and ozone pollution. Int. J. Mol. Sci. 2009, 10, 3371–3399. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.A.; Huffaker, A.; Kaplan, F.; Sims, J.; Ziemann, S.; Doehlemann, G.; Ji, L.; Schmitz, R.J.; Kolomiets, M.V.; Alborn, H.T.; et al. Maize death acids, 9-lipoxygenase-derived cyclopente(a)nones, display activity as cytotoxic phytoalexins and transcriptional mediators. Proc. Natl. Acad. Sci. USA 2015, 112, 11407–11412. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Campbell, B.C.; Mahoney, N.E.; Chan, K.L.; Molyneux, R.J. Identification of phenolics for control of Aspergillus flavus using saccharomyces cerevisae in a model target-gene bioassay. J. Agric. Food Chem. 2004, 52, 7814–7821. [Google Scholar] [CrossRef] [PubMed]
- Beekrum, S.; Govinden, R.; Padayachee, T.; Odhav, B. Naturally occurring phenols: A detoxification strategy for fumonism B1. Food Addit. Contam. 2003, 20, 490–493. [Google Scholar] [CrossRef] [PubMed]
- Curir, P.; Dolci, M.; Dolci, P.; Lanzotti, V.; de Cooman, L. Fungitoxic phenols from carnation (Dianthus caryophyllus) effective against Fusarium oxysporum, f. sp. dianthi. Phytochem. Anal. 2003, 14, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.; Sinha, A.K.; Roitsch, T. Plant physiology meets phytopathology: Plant primary metabolism and plant-pathogen interactions. J. Exp. Bot. 2007, 58, 4019–4026. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D. Primary metabolism and plant defense—Fuel for the fire. Mol. Plant Microbe Interact. 2009, 22, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K.S. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C. Disease induction by plant viruses. Adv. Virus Res. 1987, 33, 205–255. [Google Scholar] [PubMed]
- Hancock, J.T.; Henson, D.; Nyirenda, M.; Desikan, R.; Harrison, J.; Lewis, M.; Hughes, J.; Neill, S.J. Proteomic identification of glyceraldehyde 3-phosphate dehydrogenase as an inhibitory target of hydrogen peroxide in arabidopsis. Plant Physiol. Biochem. 2005, 43, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.; Fung, N.; Liu, J.; Drakakaki, G.; Coaker, G. Beyond glycolysis: GAPDHs are multi-functional enzymes involved in regulation of ROS, autophagy, and plant immune responses. PLoS Genet. 2015, 11, e1005199. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.C.; Sheen, J. Sugar sensing in higher plants. Trends Plant Sci. 1997, 2, 208–214. [Google Scholar] [CrossRef]
- Tsukaya, H.; Ohshima, T.; Naito, S.; Chino, M.; Komeda, Y. Sugar-dependent expression of the CHS-A gene for chalcone synthase from petunia in transgenic Arabidopsis. Plant Physiol. 1991, 97, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.T.; Lampi, Y.; Wang, T.W.; Liu, Z.; Thompson, J.E. Eukaryotic translation initiation factor 5A is involved in pathogen-induced cell death and development of disease symptoms in Arabidopsis. Plant Physiol. 2008, 148, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, L.; Heads, R.; Green, C.J. Cyclophilins and their possible role in the stress response. Int. J. Exp. Pathol. 1999, 80, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, J.P.; Hartl, F.U. Molecular chaperone functions of heat shock proteins. Annu. Rev. Biochem. 1993, 62, 349–384. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; van Montagu, M.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. Biochim. Biophys. Acta 2002, 1577, 1–9. [Google Scholar] [CrossRef]
- Kim, S.K.; You, Y.N.; Park, J.C.; Joung, Y.; Kim, B.G.; Ahn, J.C.; Cho, H.S. The rice thylakoid lumenal cyclophilin OsCYP20-2 confers enhanced environmental stress tolerance in tobacco and Arabidopsis. Plant Cell Rep. 2012, 31, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, K.; Priyanka, B.; Reddy, V.D.; Rao, K.V. Isolation and characterization of a pigeonpea cyclophilin (CcCYP) gene, and its over-expression in Arabidopsis confers multiple abiotic stress tolerance. Plant Cell Environ. 2010, 33, 1324–1338. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.D.; Singh, P. Effect of water stress on expression of a 20 kD cyclophilin-like protein in drought susceptible and tolerant cultivars of sorghum. J. Plant Biochem. Biotechnol. 2003, 12, 77–80. [Google Scholar] [CrossRef]
- Trivedi, D.K.; Ansari, M.W.; Tuteja, N. Multiple abiotic stress responsive rice cyclophilin: (OsCYP-25) mediates a wide range of cellular responses. Commun. Integr. Biol. 2013, 6, e25260. [Google Scholar] [CrossRef] [PubMed]
- Wisser, R.J.; Balint-Kurti, P.J.; Nelson, R.J. The genetic architecture of disease resistance in maize: A synthesis of published studies. Phytopathology 2006, 96, 120–129. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pechanova, O.; Pechan, T. Maize-Pathogen Interactions: An Ongoing Combat from a Proteomics Perspective. Int. J. Mol. Sci. 2015, 16, 28429-28448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226106
Pechanova O, Pechan T. Maize-Pathogen Interactions: An Ongoing Combat from a Proteomics Perspective. International Journal of Molecular Sciences. 2015; 16(12):28429-28448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226106
Chicago/Turabian StylePechanova, Olga, and Tibor Pechan. 2015. "Maize-Pathogen Interactions: An Ongoing Combat from a Proteomics Perspective" International Journal of Molecular Sciences 16, no. 12: 28429-28448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms161226106