Physical Localization of a Locus from Agropyron cristatum Conferring Resistance to Stripe Rust in Common Wheat

 ,
,
Abstract
:1. Introduction
2. Result
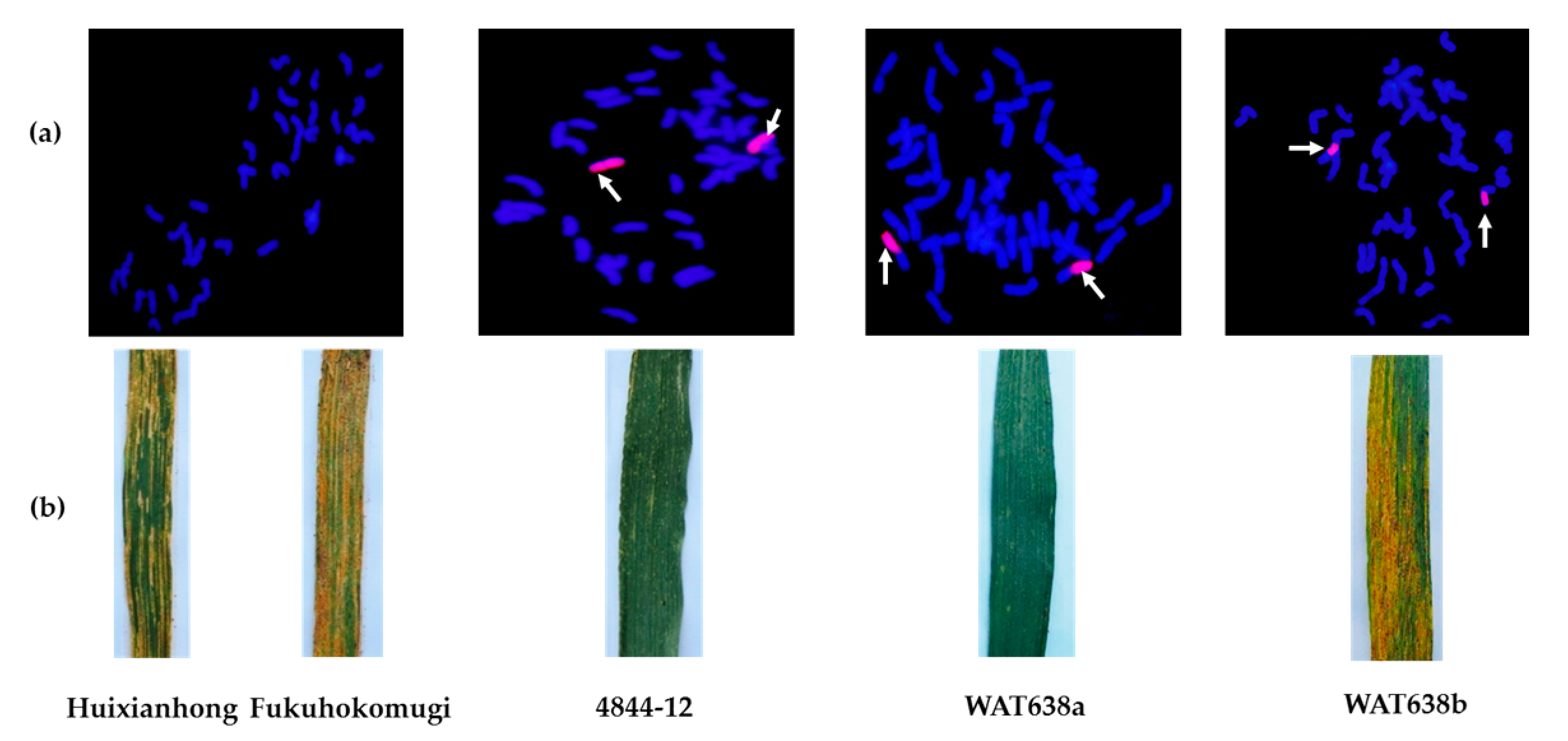
2.1. Evaluation of Stripe Rust Resistance of Wheat-A. cristatum 6P Disomic Addition Line 4844-12
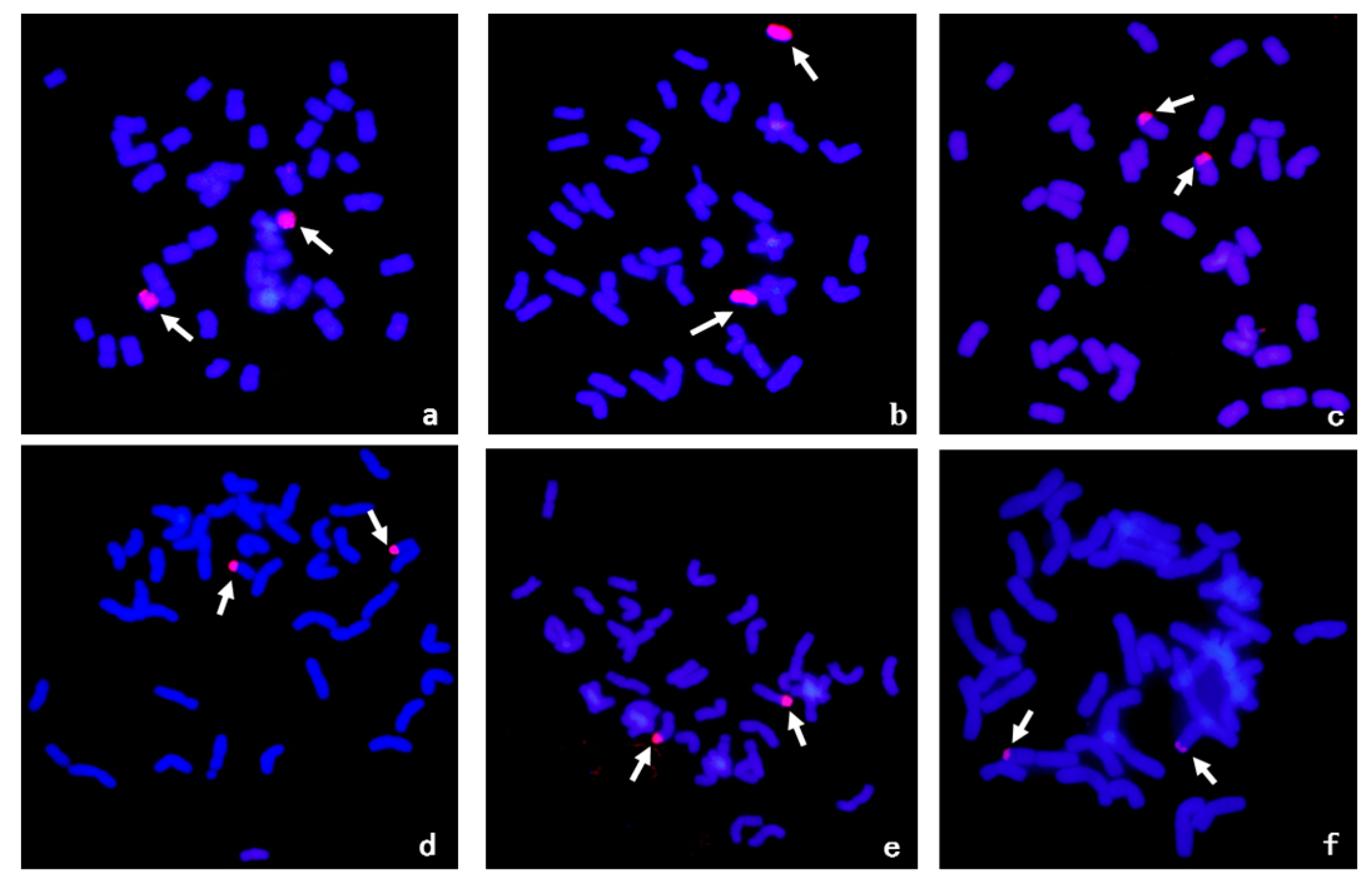
2.2. Molecular Cytogenetic Analysis and Chromosomal Arm Localization of the Locus Conferring Resistance to Stripe Rust
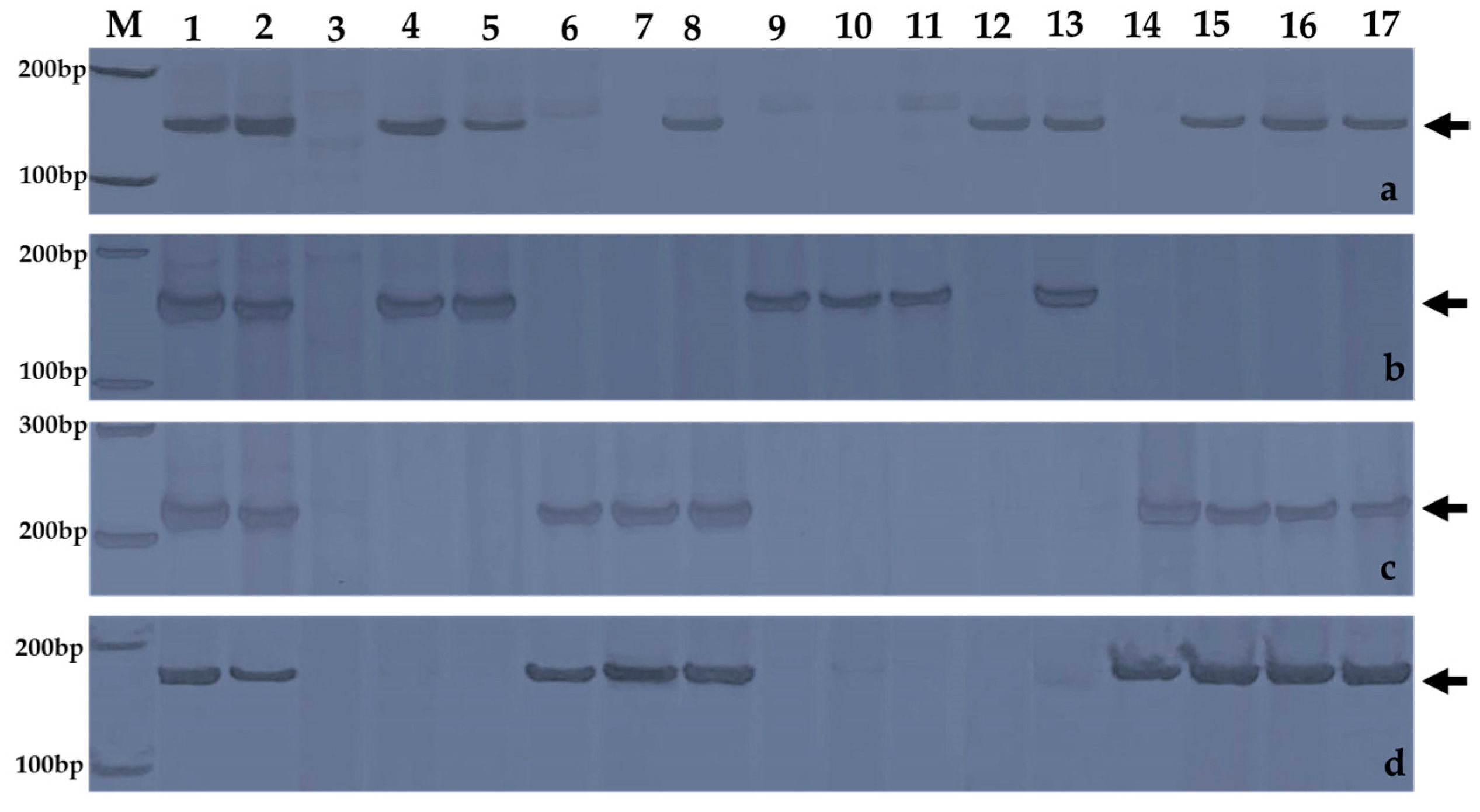
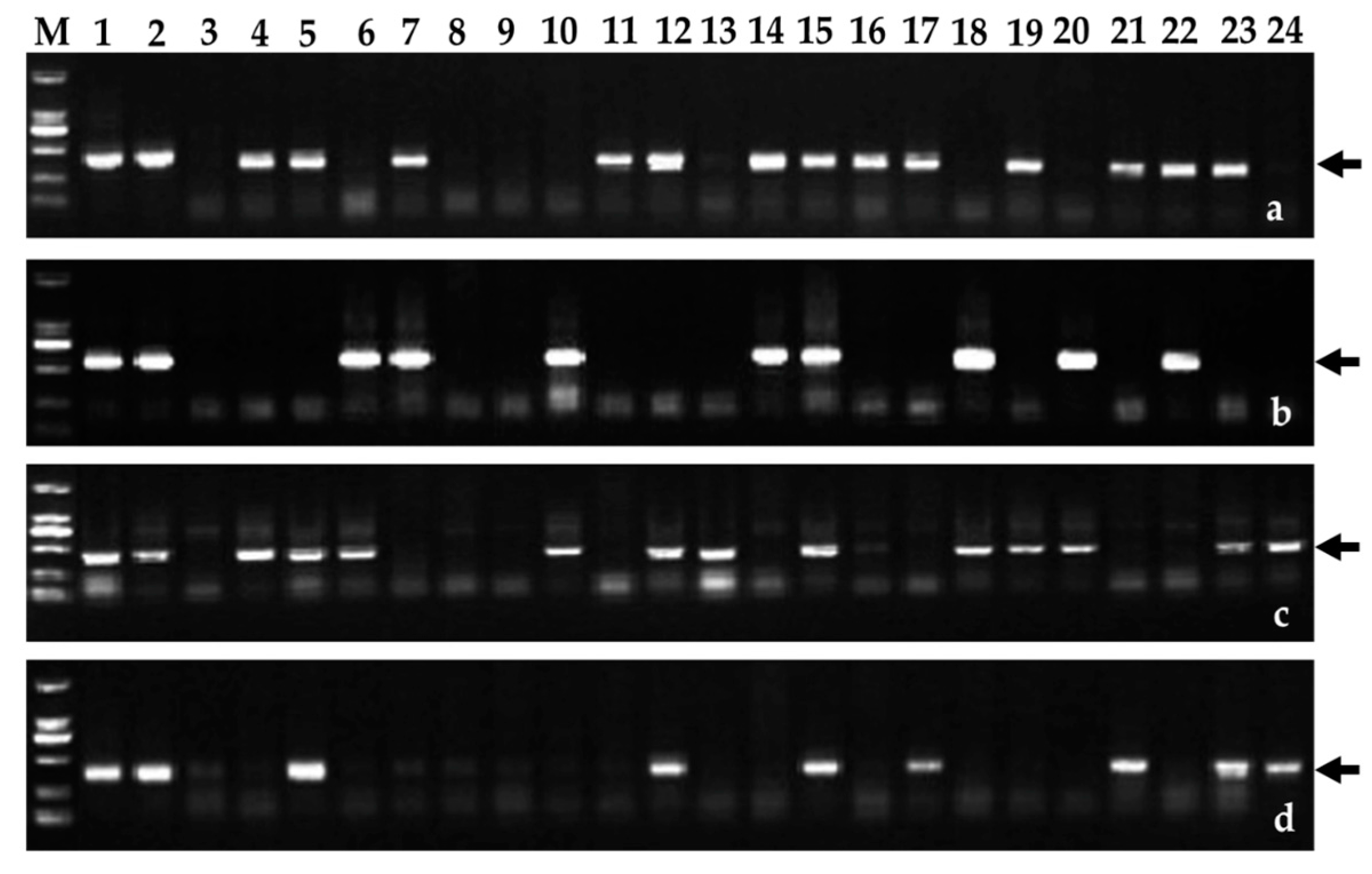
2.3. Chromosomal Segmental Localization of the Stripe Rust Resistance Locus
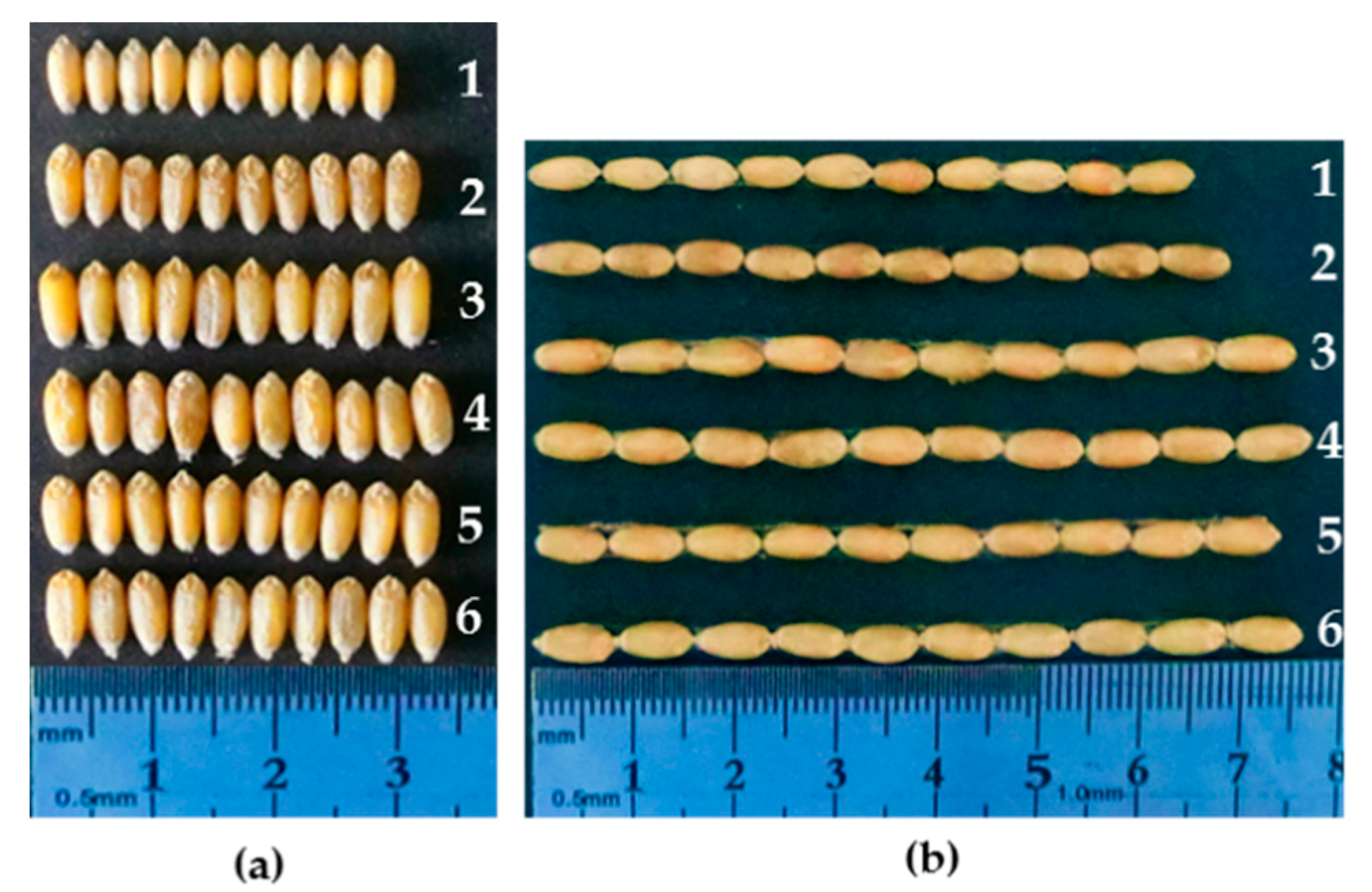
2.4. Evaluation of Agronomic Traits of the Stripe Rust-Resistant Translocation Lines
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Molecular Cytogenetic Analysis
4.3. Evaluation of Stripe Rust Resistance at Adult Stage
4.4. Statistical Analysis of Field Experiment
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GISH | Genomic in situ hybridization |
| STS | Sequence tagged sites |
| EST | Expressed sequence tag |
| DAPI | 4,6-diamino-2-phenyl indole |
| IT | Infection type |
| SAS | Statistical Analysis System |
References
- Line, R.F. Stripe rust of wheat and barley in North America: A retrospective historical review. Annu. Rev. Phytopathol. 2002, 40, 75–118. [Google Scholar] [CrossRef] [PubMed]
- Hovmøller, M.S.; Walter, S.; Justesen, A.F. Escalating threat of wheat rusts. Science 2010, 329, 369. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Hou, L.; Tang, M.S.; Wang, H.G.; Li, Q.; Jing, J.X. Genetic analysis and molecular mapping of a stripe rust resistance gene YrH9014 in wheat line H9014-14-4-6-1. J. Integr. Agric. 2013, 12, 638–645. [Google Scholar] [CrossRef]
- Chen, X.M. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Sharma-Poudual, D.; Chen, X.M.; Wan, A.M.; Zhan, G.M.; Kang, Z.S.; Cao, S.Q.; Jin, S.L.; Morgounov, A.; Akin, B.; Mert, Z.; et al. Virulence characterization of international collections of the wheat stripe rust pathogen, Puccinia striiformis f. sp. tritici. Plant Dis. 2013, 97, 379–386. [Google Scholar] [CrossRef]
- Singh, N.K.; Shepherd, K.W.; McIntosh, R.A. Linkage mapping of genes for resistance to leaf, stem, and stripe rusts and X-secalins on the short arm of rye chromosome 1R. Theor. Appl. Genet. 1990, 80, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Friebe, B.; Jiang, J.; Raupp, W.J.; McIntosh, R.A.; Gill, B.S. Characterization of wheat-alien translocations conferring resistance to diseases and pests: Current status. Euphytica 1996, 91, 59–87. [Google Scholar] [CrossRef]
- Cao, A.; Xing, L.; Wang, X.; Yang, X.; Wang, W.; Sun, Y.; Qian, C.; Ni, J.; Chen, Y.; Liu, D. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 7727–7732. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Huang, J.; Hou, L.; Liu, P.; Jing, J.; Wang, B.; Kang, Z. Genetic and molecular mapping of stripe rust resistance gene in wheat-Psathyrostachys huashanica translocation line H9020-1-6-8-3. Plant Dis. 2012, 96, 1482–1487. [Google Scholar] [CrossRef]
- Dewey, D.R. The genomic system of classification as a guide to intergeneric hybridization with the perennial Triticeae. In Gene Manipulation in Plant Improvement; Gustafson, J.P., Ed.; Plenum Press: New York, NY, USA, 1984; pp. 209–279. [Google Scholar]
- Dong, Y.C.; Zhou, R.H.; Xu, S.J.; Li, L.H.; Cauderon, Y.; Wang, R.R.C. Desirable characteristics in perennial Triticeae collected in China for wheat improvement. Hereditas 1992, 116, 175–178. [Google Scholar] [CrossRef]
- Zhang, J.P.; Liu, W.H.; Han, H.M.; Song, L.Q.; Bai, L.; Gao, Z.H.; Zhang, Y.; Yang, X.; Li, X.; Gao, A.; et al. De novo transcriptome sequencing of Agropyron cristatum to identify available gene resources for the enhancement of wheat. Genomics 2015, 106, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.C.; Gill, B.S.; Uyemoto, J.K. High levels of resistance in Agropyron species to barley yellow dwarf and wheat streak mosaic viruses. J. Phytopathol. 1984, 110, 143–147. [Google Scholar] [CrossRef]
- Wang, R.R.C. Agropyron and Psathyrostachys. In Wild Crop Relatives: Genomic and Breeding Resources, Cereals; Kole, C., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2011; pp. 77–108. [Google Scholar]
- Ochoa, V.; Madrid, E.; Said, M.; Rubiales, D.; Cabrera, A. Molecular and cytogenetic characterization of a common wheat-Agropyron cristatum chromosome translocation conferring resistance to leaf rust. Euphytica 2015, 201, 89–95. [Google Scholar] [CrossRef]
- Li, L.H.; Dong, Y.C.; Zhou, R.H.; Li, X.Q.; Li, P. Cytogenetics and self-fertility of hybrids between Triticum aestivum L. and Agropyron cristatum (L.) Gaertn. Acta Genet. Sin. 1995, 22, 109–114. [Google Scholar]
- Li, L.H.; Li, X.Q.; Li, P.; Dong, Y.C.; Zhao, G.S. Establishment of wheat-Agropyron cristatum alien addition lines. I. Cytology of F3, F2BC1, BC4, and BC3F1 progenies. Acta Genet. Sin. 1997, 24, 154–159. [Google Scholar]
- Limin, A.E.; Fowler, D.B. An interspecific hybrid and amphiploid produced from Triticum aestivum crosses with Agropyron cristatum and Agropyron desertorum. Genome 1990, 33, 581–584. [Google Scholar] [CrossRef]
- Wu, J.; Yang, X.M.; Wang, H.; Li, H.J.; Li, L.H.; Li, X.Q.; Liu, W.H. The introgression of chromosome 6P specifying for increased numbers of florets and kernels from Agropyron cristatum into wheat. Theor. Appl. Genet. 2006, 114, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.J.; Lu, Y.Q.; Li, H.H.; Pan, C.L.; Guo, Y.; Zhang, J.P.; Yang, X.M.; Li, X.Q.; Liu, W.H.; Li, L.H. Transferring Desirable Genes from Agropyron cristatum 7P Chromosome into Common Wheat. PLoS ONE 2016, 11, e0159577. [Google Scholar] [CrossRef] [PubMed]
- Li, H.H.; Jiang, B.; Wang, J.C.; Lu, Y.Q.; Zhang, J.P.; Pan, C.L.; Yang, X.M.; Li, X.Q.; Liu, W.H.; Li, L.H. Mapping of novel powdery mildew resistance gene(s) from Agropyron cristatum chromosome 2P. Theor. Appl. Genet. 2017, 130, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Copete, A.; Cabrera, A. Chromosomal location of genes for resistance to powdery mildew in Agropyron cristatum and mapping of conserved orthologous set molecular markers. Euphytica 2017, 213, 189. [Google Scholar] [CrossRef]
- Qi, L.L.; Wang, S.L.; Chen, P.D.; Liu, D.J.; Gill, B.S. Identification and physical mapping of three Haynaldia villosa chromosome-6V deletion lines. Theor. Appl. Genet. 1998, 97, 1042–1046. [Google Scholar] [CrossRef]
- Ashida, T.; Nasuda, S.; Sato, K.; Endo, T.R. Dissection of barley chromosome 5H in common wheat. Genes Genet. Syst. 2007, 82, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Song, L.Q.; Lu, Y.Q.; Zhang, J.P.; Pan, C.L.; Yang, X.M.; Li, X.Q.; Liu, W.; Li, L. Physical mapping of Agropyron cristatum chromosome 6P using deletion lines in common wheat background. Theor. Appl. Genet. 2016, 129, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Wang, X.G.; Liu, W.H.; Li, C.Y.; Zhang, J.P.; Gao, A.N.; Wang, Y.; Yang, X.; Li, L. Production and identification of wheat-Agropyron cristatum 6P translocation lines. Planta 2010, 232, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Song, L.Q.; Jiang, L.L.; Han, H.M.; Gao, A.N.; Yang, X.M.; Li, L.H.; Liu, W.H. Efficient induction of wheat-Agropyron cristatum 6P translocation lines and GISH detection. PLoS ONE 2013, 8, e69501. [Google Scholar] [CrossRef] [PubMed]
- Song, L.Q.; Lu, Y.Q.; Zhang, J.P.; Pan, C.L.; Yang, X.M.; Li, X.Q.; Liu, W.H.; Li, L.H. Cytological and molecular analysis of wheat-Agropyron cristatum translocation lines with 6P chromosome fragments conferring superior agronomic traits in common wheat. Genome 2016, 59, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Han, H.M.; Liu, W.H.; Lu, Y.Q.; Zhang, J.P.; Yang, X.M.; Li, X.; Hu, Z.; Li, L. Isolation and application of P genome-specific DNA sequences of Agropyron Gaertn. in Triticeae. Planta 2017, 245, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Friebe, B.; Gill, B.S. Recent advances in alien gene transfer in wheat. Euphytica 1994, 73, 199–212. [Google Scholar] [CrossRef]
- Zhang, J.P.; Liu, W.H.; Lu, Y.Q.; Liu, Q.; Yang, X.; Li, X.; Li, L. A resource of large-scale molecular markers for monitoring Agropyron cristatum chromatin introgression in wheat background based on transcriptome sequences. Sci. Rep. 2017, 7, 11942. [Google Scholar] [CrossRef] [PubMed]
- Wan, A.M.; Zhao, Z.H.; Chen, X.M.; He, Z.; Jin, S.; Jia, Q.; Yao, G.; Yang, J.; Wang, B.; Li, G.; et al. Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis. 2004, 88, 896–904. [Google Scholar] [CrossRef]
- Wan, A.M.; Chen, X.M.; He, Z.H. Wheat stripe rust in China. Aust. J. Agric. Res. 2007, 58, 605–619. [Google Scholar] [CrossRef]
- Chen, W.Q.; Wu, L.R.; Liu, T.G.; Xu, S.C.; Jin, S.L.; Peng, Y.L.; Wang, B.T. Race dynamics diversity, and virulence evolution in Puccinia striiformis f. sp. tritici, the causal agent of wheat stripe rust in China from 2003 to 2007. Plant Dis. 2009, 93, 1093–1101. [Google Scholar] [CrossRef]
- Liu, J.; Chang, Z.J.; Zhang, X.J.; Yang, Z.; Li, X.; Jia, J.; Zhan, H.; Guo, H.; Wang, J. Putative Thinopyrum intermedium-derived stripe rust resistance gene Yr50 maps on wheat chromosome arm 4BL. Theor. Appl. Genet. 2013, 126, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Jauhar, P.P.; Peterson, T.S.; Xu, S.S. Cytogenetic and molecular characterization of a durum alien disomic addition line with enhanced tolerance to Fusarium head blight. Genome 2009, 52, 467–483. [Google Scholar] [CrossRef] [PubMed]
- Gerechter-Amitai, Z.; Van Silfhout, C.; Grama, A.; Kleitman, F. Yr15-a new gene for resistance to Puccinia striiformis in Triticum dicoccoides sel. G-25. Euphytica 1989, 43, 187–190. [Google Scholar] [CrossRef]
- Dadkhodaie, N.; Karaoglou, H.; Wellings, C.; Park, R. Mapping genes Lr53 and Yr35 on the short arm of chromosome 6B of common wheat with microsatellite markers and studies of their association with Lr36. Theor. Appl. Genet. 2011, 122, 479–487. [Google Scholar] [CrossRef] [PubMed]
- McFadden, E.S. A successful transfer of emmer characters to vulgare wheat. J. Am. Soc. Agron. 1930, 22, 1020–1034. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to imageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.R.; Gill, B.S. The deletion stocks of common wheat. J. Hered. 1996, 87, 295–307. [Google Scholar] [CrossRef]
- Han, F.P.; Lamb, J.C.; Birchler, J.A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Schwarzacher, T.; Jouve, N. Identification of different chromatin classes in wheat using in situ hybridization with simple sequence repeat oligonucleotides. Theor. Appl. Genet. 2000, 101, 711–717. [Google Scholar] [CrossRef]
- Roland, F.L.; Abdul, Q. Collecting and Evaluating Rust Samples for Virulence. In Virulence, Aggressiveness, Evolution, and Distribution of Races of Puccinia striiformis (the Cause of Stripe Rust of Wheat) in North America, 1968–87; U.S. Department of Agriculture: Washington, DC, USA, 1992; pp. 4–9. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
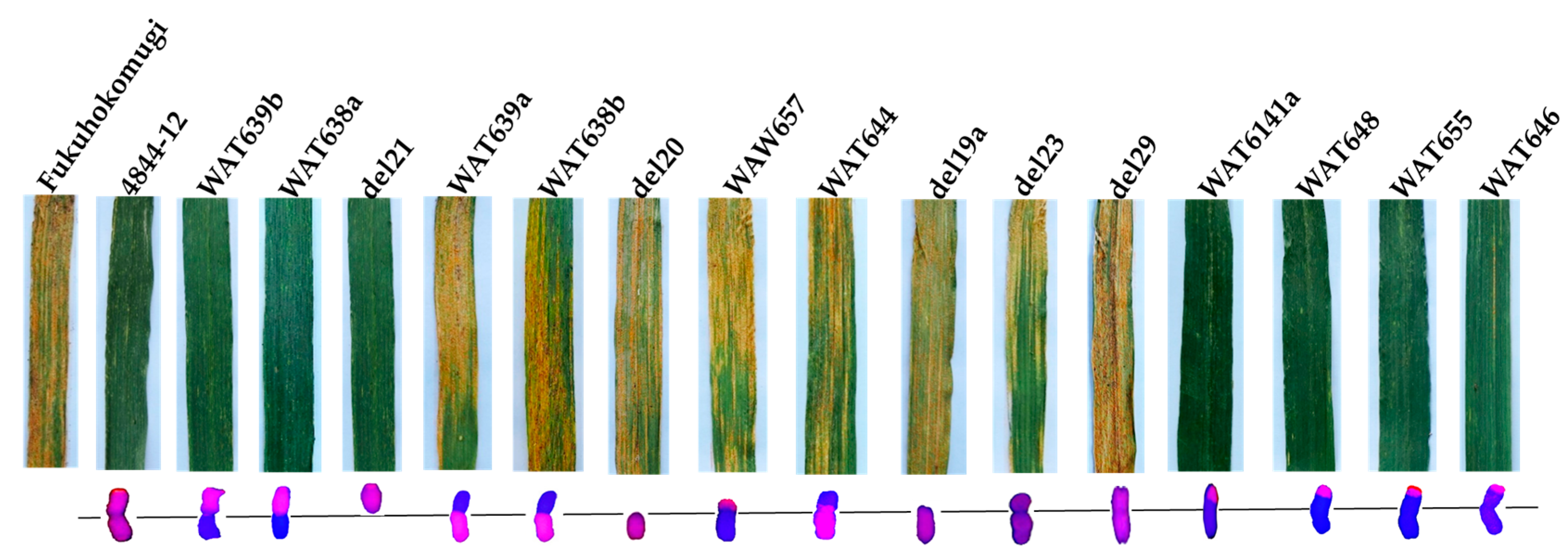
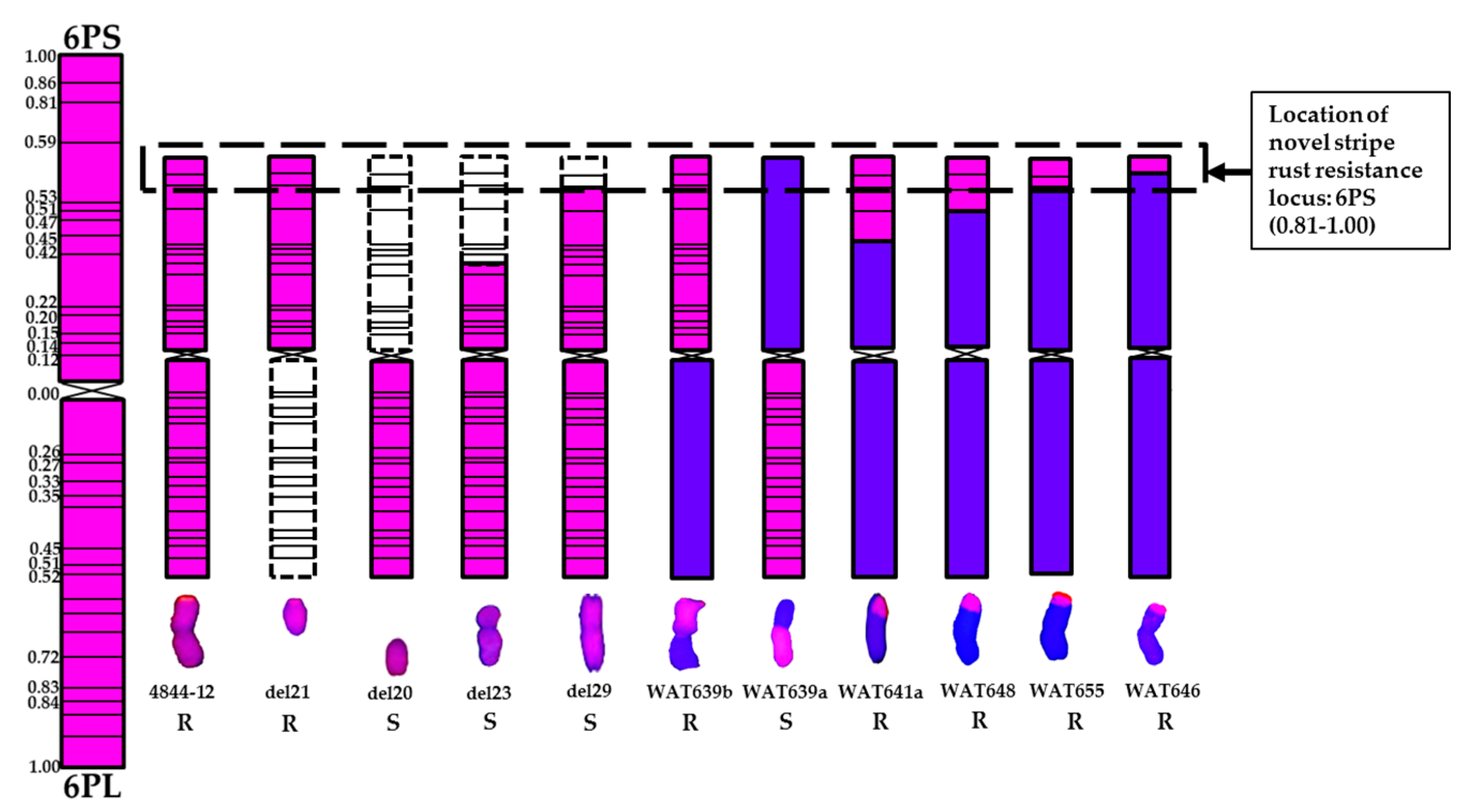
| Materials | 6P Segment Size | Stripe Rust Response | No. Detected of Each Replicate | |
|---|---|---|---|---|
| Xinxiang | Yangling | |||
| WAT639b | 6PS arm | R | 40 | 40 |
| WAT638a | 6PS arm | R | 40 | 40 |
| del21 | 6PS arm | R | 40 | 30 |
| WAT639a | 6PL arm | S | 40 | 40 |
| WAT638b | 6PL arm | S | 40 | 40 |
| del20 | 6PL arm | S | 40 | 30 |
| WAT657 | 6PS (0.00–0.15) | S | 40 | 40 |
| WAT644 | 6PS (0.00–0.20) + 6PL | S | 40 | 40 |
| del19a | 6PS (0.00–0.15) + 6PL | S | 40 | 30 |
| del23 | 6PS (0.00–0.45) + 6PL | S | 40 | 30 |
| del29 | 6PS (0.00–0.81) + 6PL | S | 40 | 30 |
| WAT641a | 6PS (0.53–1.00) | R | 40 | 40 |
| WAT648 | 6PS (0.59–1.00) | R | 40 | 40 |
| WAT655 | 6PS (0.81–1.00) | R | 40 | 40 |
| WAT646 | 6PS (0.86–1.00) | R | 40 | 40 |
| Materials | Type | 2015–2016 (BC2F2) | Total | 2016–2017 (BC3F2) | Total | ||
|---|---|---|---|---|---|---|---|
| 6P+ | 6P− | 6P− | 6P− | ||||
| Huixianhong | S (30) | 30 | S (40) | 40 | |||
| Fukuhokomugi | S (30) | 30 | S (40) | 40 | |||
| 4844-12 | R (30) | S (0) | 30 | R (40) | S (0) | 40 | |
| WAT639b * | 6PS·7AL | R (22) | S (32) | 54 | R (97) | S (52) | 149 |
| WAT639a | 7AS·6PL | S (41) | S (29) | 70 | S (135) | S (54) | 189 |
| Materials | Type | Spike Length (cm) | Spikelet Number per Spike | Kernel Number per Spikelet | Grain Number per Spike | Thousand-Grain Weight (g) |
|---|---|---|---|---|---|---|
| 4844-12 | 10.35 ± 0.69 | 23.36 ± 2.27 | 4.48 ± 0.51 | 72.96 ± 5.79 | 37.74 ± 0.46 | |
| Fukuhokomugi | 10.24 ± 0.84 | 18.76 ± 1.42 | 4.20 ± 0.41 | 55.08 ± 5.85 | 32.98 ± 1.40 | |
| WAT638a | 6PS·6AL | 10.60 ± 1.16 | 20.79 ± 2.08 * | 3.76 ± 0.57 | 55.71 ± 4.48 | 40.84 ± 3.82 ** |
| WAT639b | 6PS·7AL | 10.08 ± 0.98 | 19.48 ± 1.58 | 4.22 ± 0.68 | 57.00 ± 3.62 | 35.16 ± 3.47 * |
| WAT641a | 7A-6PS | 9.50 ± 1.15 | 17.50 ± 1.91 | 4.25 ± 0.50 | 48.50 ± 7.93 | 33.08 ± 3.52 |
| WAT648 | 5DS·5DL-6PS | 10.05 ± 0.99 | 20.44 ± 1.65 * | 3.77 ± 0.94 | 56.22 ± 5.40 | 46.92 ± 4.14 ** |
| WAT655 | 6DS·6DL-6PS | 11.26 ± 0.86 * | 21.28 ± 1.50 * | 4.07 ± 0.45 | 64.38 ± 6.65 ** | 38.38 ± 4.84 ** |
| WAT646 | 1BL·1BS-6PS | 11.52 ± 0.94 * | 21.07 ± 1.59 * | 4.07 ± 0.47 | 58.42 ± 6.15 * | 44.25 ± 4.65 ** |
| Materials | Zygosity | Progeny | Type | 6P Segment Size |
|---|---|---|---|---|
| del21 | Homozygous | M5 | 6PS telosomic | 6PS arm |
| del20 | Homozygous | M5 | 6PL telosomic | 6PL arm |
| del19a | Homozygous | M5 | 6PS terminal deletion | 6PS (0.00–0.15) + 6PL |
| del23 | Homozygous | M5 | 6PS terminal deletion | 6PS (0.00–0.45) + 6PL |
| del29 | Homozygous | M5 | 6PS terminal deletion | 6PS (0.00–0.81) + 6PL |
| WAT639b | Homozygous | BC2F3 | 6PS·7AL | 6PS arm |
| Heterozygous | BC2F2, BC3F2 | |||
| WAT638a | Homozygous | BC2F3 | 6PS·6AL | 6PS arm |
| WAT639a | Homozygous | BC2F3 | 7AS·6PL | 6PL arm |
| Heterozygous | BC2F2, BC3F2 | |||
| WAT638b | Homozygous | BC2F3 | 6AS·6PL | 6PL arm |
| WAT657 | Homozygous | BC2F3 | 6AS·6PS | 6PS (0.00–0.15) |
| WAT641a | Homozygous | BC2F3 | 7A-6PS | 6PS (0.53–1.00) |
| WAT648 | Homozygous | BC2F3 | 5DS·5DL-6PS | 6PS (0.59–1.00) |
| WAT655 | Homozygous | BC2F3 | 6DS·6DL-6PS | 6PS (0.81–1.00) |
| WAT646 | Homozygous | BC2F3 | 1BL·1BS-6PS | 6PS (0.86–1.00) |
| WAT644 | Homozygous | BC2F3 | 6PL·6PS-A | 6PS (0.00–0.20) + 6PL |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Song, L.; Han, H.; Zhou, S.; Zhang, J.; Yang, X.; Li, X.; Liu, W.; Li, L. Physical Localization of a Locus from Agropyron cristatum Conferring Resistance to Stripe Rust in Common Wheat. Int. J. Mol. Sci. 2017, 18, 2403. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112403
Zhang Z, Song L, Han H, Zhou S, Zhang J, Yang X, Li X, Liu W, Li L. Physical Localization of a Locus from Agropyron cristatum Conferring Resistance to Stripe Rust in Common Wheat. International Journal of Molecular Sciences. 2017; 18(11):2403. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112403
Chicago/Turabian StyleZhang, Zhi, Liqiang Song, Haiming Han, Shenghui Zhou, Jinpeng Zhang, Xinming Yang, Xiuquan Li, Weihua Liu, and Lihui Li. 2017. "Physical Localization of a Locus from Agropyron cristatum Conferring Resistance to Stripe Rust in Common Wheat" International Journal of Molecular Sciences 18, no. 11: 2403. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18112403