Recognizing Depression from the Microbiota–Gut–Brain Axis

1
Key Laboratory of Mental Health, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China
2
Department of Psychology, University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(6), 1592; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061592
Submission received: 28 March 2018
/
Revised: 19 April 2018
/
Accepted: 2 May 2018
/
Published: 29 May 2018
(This article belongs to the Special Issue The (Microbiota)–Gut–Brain Axis: Hype or Revolution?)
Abstract
:Major depression is one of the leading causes of disability, morbidity, and mortality worldwide. The brain–gut axis functions are disturbed, revealed by a dysfunction of the brain, immune system, endocrine system, and gut. Traditional depression treatments all target the brain, with different drugs and/or psychotherapy. Unfortunately, most of the patients have never received any treatment. Studies indicate that gut microbiota could be a direct cause for the disorder. Abnormal microbiota and the microbiota–gut–brain dysfunction may cause mental disorders, while correcting these disturbance could alleviate depression. Nowadays, the gut microbiota modulation has become a hot topic in treatment research of mental disorders. Depression is closely related with the health condition of the brain–gut axis, and maintaining/restoring the normal condition of gut microbiota helps in the prevention/therapy of mental disorders.
{kind=link}
{kind=link}
1. Introduction
Major depressive disorder is one of the leading causes of disability, morbidity, and mortality worldwide. The disorder has already affected over 350,000,000 people, and every one in five people probably suffers from it in one stage of their lifespan [1,2,3]. More than 85% of patients with a first episode will relapse in the next 10 years, and most of the patients have suicidal thoughts, 15–20% of whom will die from suicide [4,5]. As shown in a Nature report in 2014, the years lived with disability (YDLs) brought by depression account for 10.3%, which is more than any other disease [6,7]. The latest report in Lancet in 2017 also showed that the disability-adjusted life years (DALYs) brought by depression had increased between 1990 and 2016 [8].
According to modern psychology and biology conceptions, major depression is not just a mental disorder but also a physiological disease. It has obvious biological foundations, such as brain changes including unbalanced neurotransmitters, impaired neurogenesis, neuroplasticity decline, and abnormal neuronal circuitry [9,10]. Large genome-wide association studies estimated the chances of heritability to be 37% to 48% for major depression [11,12]. However, the ever-increasing incidence of depression deviates from the Hardy–Weinberg Equilibrium, indicating that environmental factors play an important role in the disorder.
It is generally agreed upon that depression is induced by the cumulative effect of genetic information and environmental stresses [11]. Certain genes and psychological features might predispose some people to depression and stressful life events including early-life stress are important inducements of depression [1,13], but the latest research has indicated that gut microbiota probably plays a crucial part in the pathophysiology of depression [14,15,16,17,18].
2. The Pathophysiology of Major Depressive Disorder
The pathophysiology of major depression has been increasing clearly following the development of neuroscience and bioinformatics. It mainly involves four aspects, the dysfunction of the brain, the hypothalamus–pituitary–adrenal (HPA) axis, the immune system, and the gut–brain axis. The brain abnormalities are mainly reflected in the unbalanced neurotransmitters, the impaired neuroplasticity, and the abnormal neural circuitry [1,10]. The HPA axis dysfunction is mainly manifested as a maladjustment of negative feedback mechanisms [19,20]. The immune changes are mainly seen as chronic inflammation [21,22]. The gut–brain dysfunction mainly includes gastrointestinal disorders and gut microbiota abnormalities [1,11,23,24,25].
2.1. The Brain Dysfunction
Neurotransmitters play a crucial part in the brain and behavior. Depression is inseparable from neurotransmitter imbalance [26,27]. The monoaminergic neurotransmitter deficiency hypothesis posits that positive moods including happiness go hand in hand with monoamine neurotransmitters serotonin (5-HT), norepinephrine (NE), and/or dopamine (DA) and that symptoms of depression arise from insufficient levels of these neurotransmitters. Recovering these neurotransmitter levels will have antidepressive effects [26,28]. However, most of the selective serotonin reuptake inhibitors (SSRIs) work slowly and just bring alleviations for part of the patients, indicating that there are still other mechanisms involved in depression [9]. Subsequent research revealed that also signaling by other neurotransmitters probably changed in depression. For example, the glutamatergic system and acetylcholine system are hyperactive, while the gamma-aminobutyric acid (GABA) system is inhibited [27,29,30].
The prefrontal cortex, hippocampus, and amygdala play a vital role in the regulation of emotion, stress responses, self-control, motivation, and cognitive reaction, but in depressed patients the function of the prefrontal cortex and hippocampus are impaired, while the activity of the amygdala is increased [31]. The traditional brain-derived neurotrophic factor (BDNF) hypothesis posits that BDNF is an important regulator of neurogenesis and that depressive symptoms arise from the decrease in BDNF content and the following increase in neural apoptosis. Therefore, long-lasting antidepressant therapies increase the neurotrophic factors including BDNF, stimulate neurogenesis, reduce the hippocampus neuronal apoptosis, and improve the mood and cognition [1,20,32]. Later research found that depressed patients not only present impaired neurogenesis but also present disturbed neuralgias growth, reduced synaptic plasticity, impaired myelin function, and a decrease in total neuroplasticity [9,10,31]. The new neuroplasticity hypothesis posits that depressive symptoms arise from the impaired neuroplasticity, which can be induced by many risks factors, including neurotransmitter imbalance and insufficient BDNF. Antidepressant therapies focusing on neurotransmitter recovery and brain stimulation work through the increase of neuroplasticity and the decrease of neuronal apoptosis [2,33,34,35]. These theories put emphasis on the changes at the molecular and cellular level, while some other theories, including the neural circuit hypothesis point the changes in function. According to the neural circuit hypothesis, depression occurs as a result of aberrations in communication between specific neural structures of the brain, such as the DA neurons in the ventral tegmental area (VTA) and their projections and 5-HT neurons in the dorsal raphe nucleus and their projections. These abnormalities in neural circuits can be restored via therapies including deep brain stimulation [10,36].
2.2. The HPA Axis Dysfunction
The HPA axis is an important part of the stress response system, and dysfunction of the HPA axis is one of the most important mechanisms behind depression [11,21,37,38]. Both psychological and physiological stress activate the HPA axis and stimulate the release of corticotrophin-releasing factor (CRF) and vasopressin (AVP) by the hypothalamus. Both CRF and AVP induce the anterior pituitary gland to secrete adrenocorticotrophic hormone (ACTH), which enhances the release of adrenocortical hormones, including glucocorticoid (GC), and causes an increase in circulatory GC levels, which inhibits the secretion of CRF and AVP by the hypothalamus, forming a negative feedback circuit [37,39]. However, over half of the depressed patients present negative feedback dysfunction of the HPA axis, including a chronic increase in circulatory GC and ACTH, and some of the patients even suffer from hypercortisolemia [21,37]. One hypothesis posits that the glucocorticoid receptor (GR) plays an important role in the HPA axis function during depression; the excessive circulating GC reduces the sensitivity of GR, while antidepressant therapies increase the GR expression, enhance the GR function, and improve the negative feedback medicated by GR [37,38]. Later research found that HPA axis dysfunction also reduces BDNF expression [39], inhibits 5-HT synthesis [40], decreases Glu receptor expression [41], and even disturbs neuroplasticity and neural circuits [10,42].
2.3. Immune System Abnormalities
Inflammation is also an important pathological feature of depression. A subpopulation of depressed patients present immune dysregulation and chronic inflammation [21,43,44]. The cytokine hypothesis posits that in depression the proinflammatory cytokines, including IL-6 and TNF-α, increase in amount while the anti-inflammatory cytokines, including interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β), decrease, making the holistic immune response tend to inflammation. The excessive proinflammatory cytokines inhibit the negative feedback of the HPA axis, increase the permeability of the blood–brain barrier, reduce the synthesis of 5-HT, disturb the glutamatergic systems, and result in depression [40,43,45,46,47,48]. The early theories mainly focus on peripheral inflammation, but new theories, such as the neuroinflammation hypothesis and inflammasome hypothesis pay more attention to central inflammation [42,47]. The neuroinflammation hypothesis emphasizes the adverse effects on the central nervous system (CNS) exerted by excessive proinflammatory cytokines released by microglia, which can be induced by various factors such as psychological stress, disease, and infection [42]. The inflammasome hypothesis puts emphasis on the influences of neuroinflammation induced by the inflammasome [49,50]. Neuroglia cells play a vital part in the regulation of neuro immune and neuroplasticity. Although different theories focus on different aspects, all of them hypothesize that the neuroinflammation and neuroplasticity impairment induced by neuroglia dysfunction results in depression [42]. The anti-inflammatory effect of traditional antidepressant therapies is not obvious [44], and combining antiinflammation methods with antidepressive therapies will probably give better results [47].
2.4. Gut Brain Dysfunction
The gut of mammals is also called gut brain because it has its own nervous system (enteric nervous system) and can make relatively independent responses to external signals [18,51]. The gut brain is a microbial organ, 90–95% of the total cells of which are microorganisms including bacteria, archaea, fungi, viruses, and some protozoa, and the metabolism, immune system, and signal transmission are all closely related with microbiota. Thus, the gut and gut microbiota can serve as a whole to respond to and influence other organs [52,53,54,55]. Depressed patients often have gut brain dysfunction, such as appetite disturbances, metabolic disturbances, functional gastrointestinal disorders, and gut microbiota abnormalities [15,23,25,56,57,58].
Major depression is not just a simple mental disorder or brain disease, but also a systemic disease. Patients often suffer from various disorders simultaneously, such as brain dysfunction and periphery dysfunction, such as HPA axis disturbances, immune dysregulation, and gut brain disturbances. These disturbances interplay with each other. For example, chronic stress reduces the 5-HT content in the brain; the synthesis and secretion of 5-HT are also influenced by various factors including the HPA axis, immune system, and gut brain, and in turn the 5-HT content affects the function of these organs [20,40,59]. As shown in Figure 1, taking both central and periphery abnormalities into account will facilitate the research of depression, and brain–gut axis dysfunction will explain the pathological basis better.
The brain-gut axis is the bidirectional message transformation pathway between brain and gut in mammals. It connects the brain and gut through several pathways including nerves, the HPA axis, and the immune system [14,18]. Factors such as psychological stress and disease impairing one or more pathways of the brain–gut axis probably induce brain–gut axis dysfunction and result in depression [25,60,61]. Following the development of gut microbiota, researchers not only focus on the top–down effects of the brain–gut axis (from brain to gut), but they also pay close attention to the down–top influences (from gut to brain) [23,25,62]. The functions of many a system, including metabolism, the immune system, the endocrine system, and the nervous system, are all closely related with the gut brain. Changes of the gut brain such as gut microbiota abnormalities influence the brain and behavior, and brain changes regulate the function and construction of the gut brain. Combining the brain and gut brain will probably become the new tendency of neuroscience, and targeting the gut microbiota will possibly be a promising area for the therapy of mental disorders and neurological diseases [18,63,64,65,66,67,68,69,70,71].
According to the gut microbiota hypothesis, the gut microbiota can influence the brain and behavior through the gut–brain axis, which is also called microbiota–gut–brain axis to emphasize the importance of the microbiota. It plays a crucial part in mental disorders [64,65]. Gut microbiota is a key component of the gut brain. It regulates the common functions and build-up of the gut brain [52,53,54,55], influences the development and maturation of the HPA axis [72,73,74,75], affects the development and function of the immune system [76,77,78], regulates the construction of the blood–brain barrier [79], influences the synthesis and recognition of neurotransmitters [59,72,80], affects neurogenesis [81], the development and function of neuralgias [82,83], and the formation of myelination [84], and impresses the development and function of the brain [66,78,85,86]. Thus, regulating gut microbiota cannot only improve gut brain dysfunction but also alleviate abnormalities in the immune system, HPA axis, and brain. All these results are in line with the gut microbiota hypothesis, which will probably be the promising direction of mental disorder therapy and prevention.
3. The Latest Research Progress of Depression: The Microbiota Hypothesis
The gut microbiota hypothesis posits that depression is closely related with gut microbiota, and microbiota–gut–brain axis dysfunction is the main pathological basis of depression. Gut microbiota abnormalities are a direct inducement and key risk factor hiding in environmental and genetic risk factors, and that microbiota regulation is the promising method for depression therapy and prevention. An increasing amount of research, exploring the gut brain in the last decades supports the hypothesis from different aspects [18,65,67,68,87,88,89,90,91,92].
3.1. Depressed Patients Have Different Gut Microbiota from Healthy Persons
Clinic studies have presented that the gut microbiota of depressed patients is significantly different from that of healthy controls. Some research found that both the microbiota diversity and richness declined in patients [57,87]. On the phylum level, the richness of Bacteroidetes and Proteobacteria increased while the richness of Firmicutes decreased; on the family level, the relative abundance of Prevotellaceae increased; on the genus level, the abundance of Prevotella increased, while the abundance of Faecalibacterium and Ruminococcus decreased [57,93]. The abundance of Lactobacillus and Bifidobacterium also declined [94]. Although all these studies have shown the gut microbiota abnormalities of depressed patients, the definite distinctions between that of patients and controls are still in debate [58,95,96], which is probably correlated with the differences of diagnostic criteria, grouping criteria, detection methods of fecal microbiota, etc.
Animal studies also presented the microbiota differences between depressive model animals and control animals. A variety of depression models have shown the phenomenon, including the bilateral olfactory bulbectomy model [97], maternal separation model [98], social disruption model [99], chronic variable stress model [100], and our chronic restraint stress model [101]. Furthermore, the microbiota of depressed animals have similarities with those of depressive patients; for example, the richness of Bacteroidetes increases while the richness of Firmicutes decreases and the abundance of Lactobacillus declines [97,98,100].
All of these investigations have suggested that depression is probably linked with certain gut microbiota phenotypes.
3.2. Depressive Symptoms Can Be Transmitted Following Fecal Microbiota Transplantation
The mental flu depression seems to have the same infectivity as flu to be transmitted from one subject to other, but the transmission medium, the gut microbiota of depressive patients, is more complex, and it is impossible to transfer under natural conditions.
After transplanting the fecal microbiota of depressed patients and healthy persons to germ-free mice, Zheng et al. found that the depressed recipient mice presented more depressive symptoms, and their microbiota were different from those of healthy recipients. These differences were similar to the differences between their respective human donors, indicating certain microbiota phenotypes can induce depressive symptoms through metabolism changes [95]. Kelly et al. studied another model, in which they transplanted the fecal microbiota of depressed patients to rats which were microbiota-depleted via antibiotic cocktail treatment. The recipient rats of patients presented obvious depressive symptoms, such as anhedonia, an increase in anxiety-like behavior, and tryptophan metabolism disturbances, all of which were similar to those of their microbiota providers [87]. Both the above studies showed that the psychological and physiological symptoms of depression can be transferred between different subjects, further indicating that psychological states are regulated by gut microbiota.
Offspring acquire similar microbiota from parents via longitudinal and horizontal gene transfer under natural conditions [102]. Our unpublished data also imply that the heredity of depression partly lies on the susceptible microbiota obtained from parents.
3.3. Gut Microbiota Disturbances Increase the Susceptibilities of Depression
Antibiotics damage the microbiota and increase the incidence of depression. Although antibiotics have played a vital role in human anti-infective therapy, they not only kill pathogens but also destroy beneficial microorganisms, induce microbiota–gut–brain axis dysfunction, and increase the incidence of various diseases, including mental disorders [103,104,105,106]. Large-scale human studies have revealed that the use of antibiotics in antiinfection therapy significantly increases the risk of mental disorders such as depression. The risk presents dose-dependent and time-dependent effects, which means the risk is positively correlated with the dose and time of antibiotic use; this elevated risk still exists 10 years after antibiotic use [107,108]. Study on infants also showed that infants who experienced antibiotic treatment during the first year of their lives had more possibilities to suffer from behavioral problems and depression, and the effects were obvious at the age of three [109]. Animal studies presented a similar phenomenon [104,110].
Stress disturbs the microbiota and increases the susceptibility of depression. Stressful life events are important inducements of depressive disorders, and they are often used in animal depression research. Chronic stress not only impacts the mind and the stress response system, but also disturbs the gut microbiota [111,112,113,114,115,116,117]. Our previous study also showed that chronic restraint stress disturbed the gut microbiota, inducing microbiota–gut–brain axis dysfunction including decreased hippocampus 5-HT content, reduced BDNF mRNA expression, increased plasma stress hormone levels, declined circulatory IL-10 levels, and abnormal gut microbiota, resulting in depression [101].
Diet is one of the most influential factors on the gut microbiota after weaning, and poor diets significantly perturb microbiota and increase the incidence of depression [102,118,119,120]. Many unhealthy diets, including the Western diet, the refined-food diet, and industrially processed food, which contains excessive saturated fat, sugar, and food additives, destroy normal gut microbiota and increase the susceptibility of depression [121,122,123]. The influences of poor diet are probably closely related with the dysfunction of the microbiota–gut–brain axis it induces [124,125].
3.4. Gut Microbiota Restoration Alleviates Depression
Gut microbiota are closely linked with host health and disease. Microbiota dysbiosis can induce various physiological and psychological diseases, and microbiota restoration brings improvement to these diseases [65,66,128,129,130]. There are four main effective methods to recover normal microbiota, which are probiotics, prebiotics, a healthy diet, and fecal microbiota transplantation (FMT) [120,131,132,133].
Probiotics are defined as “live microorganisms which when administered in adequate amounts confer a health benefit on the host” [134]. Their beneficial effects not only locate in gut, but reach the whole microbiota–gut–brain axis; researchers call these probiotics psychobiotics to emphasize their capabilities to improve behavior and mind [135]. Both clinic and animal research have shown that psychobiotics supplementation alleviates depression symptoms, even achieving similar effects to traditional antidepressant therapies. In double-blind, randomized, placebo-controlled studies, psychobiotics treatments alleviated the depressive and anxious symptoms of patients and improved cognition and metabolism [136,137,138]. Animal studies indicated that the antidepressant effects of psychobiotics are closely related with the regulation of the microbiota–gut–brain axis [99,101,139,140]. The psychobiotics that have been reported mostly belong to lactic acid bacteria, such as special strains of Lactobacillus casei [136,139], Lactobacillus helveticus [101], and Bifidobacterium bifidum [136,139].
Prebiotic is defined as “a substrate that is selectively utilized by host microorganisms conferring a health benefit” [141]. Prebiotics not only regulate gut microbiota but also improve behavior and cognition acting as psychobiotics. These effects are probably achieved through the functional improvement of the microbiota–gut–brain axis [131,142,143,144]. Prebiotics are a hot research topic; and the most popular kinds are fructose-oligosaccharide, galactooligosaccharide, omega-3 fatty acids, etc. [141].
Contrary to poor diets, healthy diets increase gut microbiota diversity and stability and improve health and wellbeing [16,131,145,146]. Healthy diets, including the Mediterranean diet, are rich in dietary fiber, unsaturated fatty acids, and fermented food such as yoghurt, cheese, and natto; they contain less refined carbohydrates, saturated fatty acids, sugar, and food additives. Healthy diets can stimulate the proliferation of beneficial microorganisms and improve behavior and cognition, probably through the microbiota–gut–brain axis [16,120,125,133,145]. These studies have greatly supported the ignored dietotherapy of depression [16].
FMT is the process of transplanting feces from a healthy donor to the receiver’s gut in order to recover the impaired gut microbiota. FMT has played important roles in the treatment of Clostridium difficile infection, inflammatory bowel disease, ulcerative colitis, etc. [147]. Some researchers have attempted to treat mental disorders including depression, anxiety, and autism.
3.5. The Mechanisms of Traditional Antidepressant Therapies are Probably Related with Microbiota
It is generally recognized that traditional antidepressant therapies alleviate mental and brain abnormalities and treat depression. But the latest research has found that traditional therapies not only regulate the brain but also influence gut microbiota; their antidepressant effects are probably partly related with the regulation of the microbiota–gut–brain axis [16,149,150].
Medical therapy regulates the gut microbiota. The first antidepressant, isoniazid, was originally used to treat Mycobacterium tuberculosis infections; the first-generation tricyclic antidepressants (TCAs) can inhibit the proliferation of many bacteria, including Escherichia coli, Yersinia, and Plasmodium; the currently common SSRI antidepressants can inhibit the proliferation of Gram-positive bacteria; even the recently developed antidepressant ketamine can inhibit the proliferation of Staphylococcus, Enterococcus, and Candida albicans. Common antibiotics, including ceftriaxone sodium and minocycline, present some antidepressant effects [149]. Citalopram showed antidepressant effects in our chronic restraint stress rat model [101]; it also changed the gut microbiota toward a new adaptation, different from the control. This indicates that the effects of antidepressants are probably correlated with gut microbiota. They also remind that considering gut microbiota possibly helps the therapy of treatment resistant depression.
Other therapies also changed the gut microbiota. Except the dietotherapy mentioned above, exercise therapy probably improves depression via the regulation of gut microbiota and the microbiota–gut–brain axis; a poor sedentary life style characterized by a lack of exercises increases the incidence of depression, while getting enough exercise alleviates depression [150,151].
3.6. New Therapies Integrating Gut Microbiota Regulation Present Promising Effects
Some researchers have tried to use new antidepressant therapies emphasizing gut microbiota regulation. In 2016, Schnorr and Bachner combined dietotherapy and psychotherapy to treat a panic attack patient; they also removed the foods that caused long-duration blood sugar spikes from and increased the food rich in probiotics in the diet. The integrated treatment alleviated the anxiety and insomnia symptoms of the patient, increased the beneficial bacterial abundances in feces, including Lactobacillus, reduced the harmful bacteria, including Clostridium, and changed the microbiota composition and diversity [152]. Bambling et al. conducted a pilot study in 2017, in which they used a new therapy combing probiotics, magnesium orotate, and SSRIs for the treatment resistant depression. The depressive symptoms were significantly improved after 8 weeks of intervention; the patients had relapsed after cessation of the test intervention while still on SSRI medication [153]. The above two studies indicate that better effects will be achieved if the gut microbiota is also taken into account in antidepressant therapies.
In conclusion, depressive patients had abnormal gut microbiota, which were probably induced by various factors, including antibiotic use, stress, poor diet, hereditary susceptibility, etc. This depressive gut microbiota phenotype is relatively stable and can be transferred from one subject to another in particular situations. Impairing the normal microbiota increases the incidence of depression, while microbiota recovery using many ways including psychobiotics, prebiotics, FMT, healthy diets, exercises, and medication has antidepressant effects.
4. Conclusions and Outlook
In the past 100 years, scientists have made great progress in depression research, and many antidepressant therapies have been developed, among which the most common is medical treatment [154]. However, currently available treatments are limited by low rates of efficacy, therapeutic time lag, and undesirable side effects [3,10,155]. For a variety of reasons, including stigma, most people with depression go undiagnosed or untreated (approximately 75% in the UK and over 92% in China). Even if the disorder is diagnosed, today’s medications will work well for only about half of those who seek help [3,6,156,157].
Negative cognitive style, unbalanced brain neurotransmitters, declined neuroplasticity, abnormal neural circuits, HPA axis dysfunction, chronic inflammation, and gut brain dysfunction are common symptoms of depression, several of which usually present at the same time, indicating that microbiota–gut–brain axis dysfunction is probably the main pathological mechanism of depression [69,70,71,158,159,160]. The focus of depression research has transferred from the mind to the brain, to other systems, to the brain-gut and gut-brain axis, and finally to the microbiota–gut–brain axis. Gut microbiota abnormalities can directly induce depression, gut microbiota can influence the behavior and mind via the microbiota–gut–brain axis, and microbiota–gut–brain axis dysfunction is the main pathophysiology of depression, according to the gut microbiota hypothesis. According to the hypothesis, gut microbiota regulation and the following microbiota–gut–brain axis improvement will alleviate and treat depression. Many microbiota recovery methods have been established, including supplementation with psychobiotics and/or prebiotics, diet regulation, and FMT.
For a long time, the antidepressant therapies usually targeted the brain abnormalities, while the dysfunction of other organs was ignored or supposed to alleviate following the improvement of brain function. Regulating the gut microbiota and improving the microbiota–gut–brain axis function will probably bring far-reaching influences to the therapy and prevention of depression. The therapy has higher flexibility and operability compared with traditional therapies, and it is easier for patients to accept and easier for high-risk individuals to prevent depression.
More than 100 years ago, Metchnikoff proposed that abnormal gut microbiota were probably the root of mental disorders including depression and anxiety, and supplementation with probiotics improved these disorders, but the theory has been neglected because of various limitations [161,162,163]. Logan and Katzman suggested that probiotics may be an adjuvant therapy for major depression in 2005 [56]. Scientists have started to pay attention to the role of the brain–gut axis in depression since 2009 [14,18,23,62,164,165]. Dinan et al. proposed the concept of “psychobiotics” to emphasize the potential of probiotics in the treatment of mental disorders [135]. Almost at the same time, some probiotics were proved to have antidepressant effects in animal depression research [101,140]. Since then, the clinical research using probiotics to treat depression has been increasing [136,137,138]. Recently, scientists have tried to treat treatment resistant depression using therapy integrating probiotic supplementation and SSRI medication [153]. It is expected that therapies targeting gut microbiota and the microbiota–gut–brain axis will play an important role in the treatment and prevention of depression in the near future.
Acknowledgments
The study was granted by NS Bio Japan. We also thank LetPub (www.letpub.com) for providing linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aan het Rot, M.; Mathew, S.J.; Charney, D.S. Neurobiological Mechanisms in Major Depressive Disorder. Can. Med. Assoc. J. 2009, 180, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, D.M.; Wohleb, E.S.; Duman, R.S. Emerging Treatment Mechanisms for Depression: Focus on Glutamate and Synaptic Plasticity. Drug Discov. Today 2016, 21, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Ledford, H. Medical Research: If Depression Were Cancer. Nature 2014, 515, 182–184. [Google Scholar] [CrossRef] [PubMed]
- Sim, K.; Lau, W.K.; Sim, J.; Sum, M.Y.; Baldessarini, R.J. Prevention of Relapse and Recurrence in Adults with Major Depressive Disorder: Systematic Review and Meta-Analyses of Controlled Trials. Int. J. Neuropsychopharmacol. 2015, 19. [Google Scholar] [CrossRef] [PubMed]
- Miret, M.; Ayuso-Mateos, J.L.; Sanchez-Moreno, J.; Vieta, E. Depressive Disorders and Suicide: Epidemiology, Risk Factors, and Burden. Neurosci. Biobehav. Rev. 2013, 37, 2372–2374. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Torres, C.D. Mental Health: A World of Depression. Nature 2014, 515, 181. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, A.J.; Charlson, F.J.; Norman, R.E.; Patten, S.B.; Freedman, G.; Murray, C.J.; Vos, T.; Whiteford, H.A. Burden of Depressive Disorders by Country, Sex, Age, and Year: Findings from the Global Burden of Disease Study 2010. PLoS Med. 2013, 10, E1001547. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 DALYs and HALE Collaborators. Global, Regional, and National Disability-Adjusted Life-Years (Dalys) for 333 Diseases and Injuries and Healthy Life Expectancy (Hale) for 195 Countries and Territories, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1260–1344. [Google Scholar] [CrossRef]
- Liu, B.; Liu, J.; Wang, M.; Zhang, Y.; Li, L. From Serotonin to Neuroplasticity: Evolvement of Theories for Major Depressive Disorder. Front. Cell. Neurosci. 2017, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, D.; Liu, H.; Han, M.H. Neuronal Correlates of Depression. Cell. Mol. Life Sci. 2015, 72, 4825–4848. [Google Scholar] [CrossRef] [PubMed]
- Belmaker, R.H.; Agam, G. Major Depression Disorder. N. Engl. J. Med. 2008, 358, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Corfield, E.C.; Yang, Y.; Martin, N.G.; Nyholt, D.R. A Continuum of Genetic Liability for Minor and Major Depression. Transl. Psychiatry 2017, 7, E1131. [Google Scholar] [CrossRef] [PubMed]
- Bukh, J.D.; Bock, C.; Vinberg, M.; Werge, T.; Gether, U.; Vedel Kessing, L. Interaction between Genetic Polymorphisms and Stressful Life Events in First Episode Depression. J. Affect. Disord. 2009, 119, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, P.; Sudo, N.; Dinan, T.; Taylor, V.H.; Bienenstock, J. Mood and Gut Feelings. Brain Behav. Immun. 2010, 24, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Evrensel, A.; Ceylan, M.E. The Gut-Brain Axis: The Missing Link in Depression. Clin. Psychopharmacol. Neurosci. 2015, 13, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Clarke, G.; Berk, M.; Jacka, F.N. The Gut Microbiome and Diet in Psychiatry: Focus on Depression. Curr. Opin. Psychiatry 2015, 28, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kundu, P.; Blacher, E.; Elinav, E.; Pettersson, S. Our Gut Microbiome: The Evolving Inner Self. Cell 2017, 171, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wang, T.; Hu, X.; Li, W.; Jin, F.; Wang, L. Microorganism and Behavior and Psychiatric Disorders. Adv. Psychol. Sci. 2012, 20, 75–97. (In Chinese) [Google Scholar] [CrossRef]
- Leonard, B.E. The HPA and Immune Axes in Stress: The Involvement of the Serotonergic System. Eur. Psychiatry 2005, 20, 302–306. [Google Scholar] [CrossRef]
- Mahar, I.; Bambico, F.R.; Mechawar, N.; Nobrega, J.N. Stress, Serotonin, and Hippocampal Neurogenesis in Relation to Depression and Antidepressant Effects. Neurosci. Biobehav. Rev. 2014, 38, 173–192. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.W.; Lowry, C.A. The Hygiene Hypothesis and Psychiatric Disorders. Trends Immunol. 2008, 29, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Lima-Ojeda, J.M.; Rupprecht, R.; Baghai, T.C. “I Am I and My Bacterial Circumstances”: Linking Gut Microbiome, Neurodevelopment, and Depression. Front. Psychiatry 2017, 8, 153. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M.; Bercik, P. The Relationship Between Intestinal Microbiota and the Central Nervous System in Normal Gastrointestinal Function and Disease. Gastroenterology 2009, 136, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- O’mahony, S.M.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Irritable Bowel Syndrome and Stress-Related Psychiatric Co-Morbidities: Focus on Early Life Stress. Handb. Exp. Pharmacol. 2017, 239, 219–246. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmsen, I. Brain-Gut Axis as an Example of the Bio-Psycho-Social Model. Gut 2000, 47, 5–7. [Google Scholar] [CrossRef]
- Hamon, M.; Blier, P. Monoamine Neurocircuitry in Depression and Strategies for New Treatments. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Lener, M.S.; Niciu, M.J.; Ballard, E.D.; Park, M.; Park, L.T.; Nugent, A.C.; Zarate, C.A., Jr. Glutamate and Gamma-Aminobutyric Acid Systems in the Pathophysiology of Major Depression and Antidepressant Response to Ketamine. Biol. Psychiatry 2017, 81, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Rm, H. History and Evolution of the Monoamine Hypothesis of Depression. J. Clin. Psychiatry 2000, 61 (Suppl. 6), 4–6. [Google Scholar]
- Pytka, K.; Dziubina, A.; Mlyniec, K.; Dziedziczak, A.; Zmudzka, E.; Furgala, A.; Olczyk, A.; Sapa, J.; Filipek, B. The Role of Glutamatergic, Gaba-Ergic, and Cholinergic Receptors in Depression and Antidepressant-Like Effect. Pharmacol. Rep. 2016, 68, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Murrough, J.W.; Abdallah, C.G.; Mathew, S.J. Targeting Glutamate Signalling in Depression: Progress and Prospects. Nat. Rev. Drug Discov. 2017, 16, 472–486. [Google Scholar] [CrossRef] [PubMed]
- Serafini, G. Neuroplasticity and Major Depression, the Role of Modern Antidepressant Drugs. World J. Psychiatry 2012, 2, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Sahay, A.; Hen, R. Adult Hippocampal Neurogenesis in Depression. Nat. Neurosci. 2007, 10, 1110–1115. [Google Scholar] [CrossRef] [PubMed]
- Kraus, C.; Castren, E.; Kasper, S.; Lanzenberger, R. Serotonin and Neuroplasticity—Links between Molecular, Functional and Structural Pathophysiology in Depression. Neurosci. Biobehav. Rev. 2017, 77, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.D.; Correia, J.S.; Patricio, P.; Mateus-Pinheiro, A.; Machado-Santos, A.R.; Loureiro-Campos, E.; Morais, M.; Bessa, J.M.; Sousa, N.; Pinto, L. Adult Hippocampal Neuroplasticity Triggers Susceptibility to Recurrent Depression. Transl. Psychiatry 2017, 7, E1058. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Lane, H.Y.; Lin, C.H. New Treatment Strategies of Depression: Based on Mechanisms Related to Neuroplasticity. Neural Plast. 2017, 2017, 4605971. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.M. Precision Psychiatry: A Neural Circuit Taxonomy for Depression and Anxiety. Lancet Psychiatry 2016, 3, 472–480. [Google Scholar] [CrossRef]
- Barden, N. Implication of the Hypothalamic-Pituitary-Adrenal Axis in the Physiopathology of Depression. J. Psychiatry Neurosci. 2004, 29, 185–193. [Google Scholar] [PubMed]
- Juruena, M.F.; Cleare, A.J.; Pariante, C.M. The Hypothalamic Pituitary Adrenal Axis, Glucocorticoid Receptor Function and Relevance to Depression. Rev. Bras. Psiquiatr. 2004, 26, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Kunugi, H.; Hori, H.; Adachi, N.; Numakawa, T. Interface between Hypothalamic-Pituitary-Adrenal Axis and Brain-Derived Neurotrophic Factor in Depression. Psychiatry Clin. Neurosci. 2010, 64, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Leonard, B.E.; Myint, A.M.; Kubera, M.; Verkerk, R. The New ‘5-Ht’ Hypothesis of Depression: Cell-Mediated Immune Activation Induces Indoleamine 2,3-Dioxygenase, Which Leads to Lower Plasma Tryptophan and an Increased Synthesis of Detrimental Tryptophan Catabolites (Trycats), Both of Which Contribute to the Onset of Depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 702–721. [Google Scholar] [CrossRef] [PubMed]
- Nasca, C.; Bigio, B.; Zelli, D.; Nicoletti, F.; Mcewen, B.S. Mind the Gap: Glucocorticoids Modulate Hippocampal Glutamate Tone Underlying Individual Differences in Stress Susceptibility. Mol. Psychiatry 2015, 20, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Singhal, G.; Baune, B.T. Microglia: An Interface between the Loss of Neuroplasticity and Depression. Front. Cell. Neurosci. 2017, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Schiepers, O.J.; Wichers, M.C.; Maes, M. Cytokines and Major Depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, D.; Dhabhar, F.S.; James, S.J.; Hough, C.M.; Jain, F.A.; Bersani, F.S.; Reus, V.I.; Verhoeven, J.E.; Epel, E.S.; Mahan, L.; et al. Oxidative Stress, Inflammation and Treatment Response in Major Depression. Psychoneuroendocrinology 2017, 76, 197–205. [Google Scholar] [CrossRef] [PubMed]
- O’brien, S.M.; Scott, L.V.; Dinan, T.G. Cytokines: Abnormalities in Major Depression and Implications for Pharmacological Treatment. Hum. Psychopharmacol. 2004, 19, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Wichers, M.; Maes, M. The Psychoneuroimmuno-Pathophysiology of Cytokine-Induced Depression in Humans. Int. J. Neuropsychopharmacol. 2002, 5, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.E. Inflammation and Depression: A Causal or Coincidental Link to the Pathophysiology? Acta Neuropsychiatr. 2018, 30, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Haroon, E.; Miller, A.H. Inflammation Effects on Brain Glutamate in Depression: Mechanistic Considerations and Treatment Implications. Curr. Top. Behav. Neurosci. 2017, 31, 173–198. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.E.; Hinz, R.; Conen, S.; Gregory, C.J.; Matthews, J.C.; Anton-Rodriguez, J.M.; Gerhard, A.; Talbot, P.S. Elevated Translocator Protein in Anterior Cingulate in Major Depression and a Role for Inflammation in Suicidal Thinking: A Positron Emission Tomography Study. Biol. Psychiatry 2018, 83, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Franklin, T.C.; Wohleb, E.S.; Zhang, Y.; Fogaca, M.; Hare, B.; Duman, R.S. Persistent Increase in Microglial Rage Contributes to Chronic Stress-Induced Priming of Depressive-Like Behavior. Biol. Psychiatry 2018, 83, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Gershon, M.D. The Bowel and Beyond: The Enteric Nervous System in Neurological Disorders. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 517–528. [Google Scholar] [CrossRef] [PubMed]
- O’hara, A.M.; Shanahan, F. The Gut Flora as a Forgotten Organ. Embo Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Lyte, M. The Microbial Organ in the Gut as a Driver of Homeostasis And Disease. Med. Hypotheses 2010, 74, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Avetisyan, M.; Schill, E.M.; Heuckeroth, R.O. Building a Second Brain in the Bowel. J. Clin. Investig. 2015, 125, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Knight, R.; Callewaert, C.; Marotz, C.; Hyde, E.R.; Debelius, J.W.; Mcdonald, D.; Sogin, M.L. The Microbiome and Human Biology. Annu. Rev. Genomics Hum. Genet. 2017, 18, 65–86. [Google Scholar] [CrossRef] [PubMed]
- Logan, A.C.; Katzman, M. Major Depressive Disorder: Probiotics May Be an Adjuvant Therapy. Med. Hypotheses 2005, 64, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered Fecal Microbiota Composition in Patients with Major Depressive Disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linlokken, A.; Wilson, R.; Rudi, K. Correlation between the Human Fecal Microbiota and Depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- O’mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, Tryptophan Metabolism and the Brain-Gut-Microbiome Axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef] [PubMed]
- O’mahony, S.M.; Hyland, N.P.; Dinan, T.G.; Cryan, J.F. Maternal Separation as a Model of Brain-Gut Axis Dysfunction. Psychopharmacology 2011, 214, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.V.; Clarke, G.; Dinan, T.G. The Brain-Gut Axis: A Target for Treating Stress-Related Disorders. Mod. Trends Pharmacopsychiatry 2013, 28, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, K.A.; Foster, J.A. Effects of Gut Microbiota on the Brain: Implications for Psychiatry. J. Psychiatry Neurosci. 2009, 34, 230–231. [Google Scholar] [PubMed]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/Brain Axis and the Microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain-Gut-Microbiota Axis: Challenges for Translation in Psychiatry. Ann. Epidemiol. 2016, 26, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Rieder, R.; Wisniewski, P.J.; Alderman, B.L.; Campbell, S.C. Microbes and Mental Health: A Review. Brain Behav. Immun. 2017, 66, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.J.; Murphy, A.B.; Cryan, J.F.; Ross, P.R.; Dinan, T.G.; Stanton, C. Microbiome in Brain Function and Mental Health. Trends Food Sci. Technol. 2016, 57, 289–301. [Google Scholar] [CrossRef]
- Yarandi, S.S.; Peterson, D.A.; Treisman, G.J.; Moran, T.H.; Pasricha, P.J. Modulatory Effects of Gut Microbiota on the Central Nervous System: How Gut Could Play a Role in Neuropsychiatric Health and Diseases. J. Neurogastroenterol. Motil. 2016, 22, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Fond, G.; Boukouaci, W.; Chevalier, G.; Regnault, A.; Eberl, G.; Hamdani, N.; Dickerson, F.; Macgregor, A.; Boyer, L.; Dargel, A.; et al. The “Psychomicrobiotic”: Targeting Microbiota in Major Psychiatric Disorders: A Systematic Review. Pathol.-Biol. 2015, 63, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.A. Brain, Meet Gut. Nature 2015, 526, 312–314. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut Microbes and the Brain: Paradigm Shift in Neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Lyte, M.; Meyer, E.; Cryan, J.F. Gut Microbiota and Brain Function: An Evolving Field in Neuroscience. Int. J. Neuropsychopharmacol. 2016, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N. Microbiome, HPA Axis and Production of Endocrine Hormones in the Gut. Adv. Exp. Med. Biol. 2014, 817, 177–194. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G.; Jury, J.; Macqueen, G.; Sherman, P.M.; Perdue, M.H. Probiotic Treatment of Rat Pups Normalises Corticosterone Release and Ameliorates Colonic Dysfunction Induced by Maternal Separation. Gut 2007, 56, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Eutamene, H.; Bueno, L. Role of Probiotics in Correcting Abnormalities of Colonic Flora Induced by Stress. Gut 2007, 56, 1495–1497. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N.; Chida, Y.; Kubo, C. Postnatal Microbial Colonization Programs the Hypothalamic-Pituitary-Adrenal System for Stress Response in Mice. J. Psychosom. Res. 2005, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The Microbiota in Adaptive Immune Homeostasis and Disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Suez, J.; Elinav, E. The Interplay between the Innate Immune System and the Microbiota. Curr. Opin. Immunol. 2014, 26, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.; Yim, Y.S.; Ha, S.; Atarashi, K.; Tan, T.G.; Longman, R.S.; Honda, K.; Littman, D.R.; Choi, G.B.; et al. Maternal Gut Bacteria Promote Neurodevelopmental Abnormalities in Mouse Offspring. Nature 2017, 549, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Bien-Ly, N.; Watts, R.J. The Blood-Brain Barrier’s Gut Check. Sci. Transl. Med. 2014, 6, 263fs46. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus Strain Regulates Emotional Behavior and Central Gaba Receptor Expression in a Mouse via the Vagus Nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed]
- Ogbonnaya, E.S.; Clarke, G.; Shanahan, F.; Dinan, T.G.; Cryan, J.F.; O’leary, O.F. Adult Hippocampal Neurogenesis Is Regulated by the Microbiome. Biol. Psychiatry 2015, 78, E7–E9. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Ruiz, A.; Mosley, M.; George, A.J.; Mussaji, L.F.; Fullerton, E.F.; Ruszkowski, E.M.; Jacobs, A.J.; Gewirtz, A.T.; Chassaing, B.; Forger, N.G. The Microbiota Influences Cell Death and Microglial Colonization in the Perinatal Mouse Brain. Brain Behav. Immun. 2018, 67, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Hrabe De Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Hoban, A.E.; Stilling, R.M.; Ryan, F.J.; Shanahan, F.; Dinan, T.G.; Claesson, M.J.; Clarke, G.; Cryan, J.F. Regulation of Prefrontal Cortex Myelination by the Microbiota. Transl. Psychiatry 2016, 6, E774. [Google Scholar] [CrossRef] [PubMed]
- Borre, Y.E.; O’keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and Neurodevelopmental Windows: Implications for Brain Disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Bjorkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal Gut Microbiota Modulates Brain Development and Behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Borre, Y.; O’brien, C.; Patterson, E.; El Aidy, S.; Deane, J.; Kennedy, P.J.; Beers, S.; Scott, K.; Moloney, G.; et al. Transferring the Blues: Depression-Associated Gut Microbiota Induces Neurobehavioural Changes in the Rat. J. Psychiatr. Res. 2016, 82, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Luna, R.A.; Foster, J.A. Gut Brain Axis: Diet Microbiota Interactions and Implications for Modulation of Anxiety and Depression. Curr. Opin. Biotechnol. 2015, 32, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, R.; Stone, T.W. The Gut-Brain Axis, BDNF, NMDA and CNS Disorders. Neurochem. Res. 2016, 41, 2819–2835. [Google Scholar] [CrossRef] [PubMed]
- Farmer, A.D.; Randall, H.A.; Aziz, Q. It’s a Gut Feeling: How the Gut Microbiota Affects the State of Mind. J. Physiol. 2014, 592, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine Pathway Metabolism and the Microbiota-Gut-Brain Axis. Neuropharmacology 2017, 112, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-Altering Microorganisms: The Impact of the Gut Microbiota on Brain and Behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Wang, X.; Wang, Z.; Zhang, J.; Jiang, R.; Wang, X.; Wang, K.; Liu, Z.; Xia, Z.; et al. Similar Fecal Microbiota Signatures in Patients with Diarrhea-Predominant Irritable Bowel Syndrome and Patients with Depression. Clin. Gastroenterol. Hepatol. 2016, 14, 1602–1611.E5. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, E.; Tsuji, H.; Asahara, T.; Takahashi, T.; Teraishi, T.; Yoshida, S.; Ota, M.; Koga, N.; Hattori, K.; Kunugi, H. Possible Association of Bifidobacterium and Lactobacillus in the Gut Microbiota of Patients with Major Depressive Disorder. J. Affect. Disord. 2016, 202, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut Microbiome Remodeling Induces Depressive-Like Behaviors through a Pathway Mediated by the Host’s Metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Ding, B.; Feng, C.; Yin, S.; Zhang, T.; Qi, X.; Lv, H.; Guo, X.; Dong, K.; Zhu, Y.; et al. Prevotella and Klebsiella Proportions in Fecal Microbial Communities Are Potential Characteristic Parameters for Patients with Major Depressive Disorder. J. Affect. Disord. 2017, 207, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Park, A.J.; Collins, J.; Blennerhassett, P.A.; Ghia, J.E.; Verdu, E.F.; Bercik, P.; Collins, S.M. Altered Colonic Function and Microbiota Profile in a Mouse Model of Chronic Depression. Neurogastroenterol. Motil. 2013, 25, 733-e575. [Google Scholar] [CrossRef] [PubMed]
- O’mahony, S.M.; Marchesi, J.R.; Scully, P.; Codling, C.; Ceolho, A.M.; Quigley, E.M.; Cryan, J.F.; Dinan, T.G. Early Life Stress Alters Behavior, Immunity, and Microbiota in Rats: Implications for Irritable Bowel Syndrome and Psychiatric Illnesses. Biol. Psychiatry 2009, 65, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Bharwani, A.; Mian, M.F.; Surette, M.G.; Bienenstock, J.; Forsythe, P. Oral Treatment with Lactobacillus Rhamnosus Attenuates Behavioural Deficits and Immune Changes in Chronic Social Stress. BMC Med. 2017, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Jia, H.; Zhou, C.; Yang, Y.; Zhao, Y.; Yang, M.; Zou, Z. Variations in Gut Microbiota and Fecal Metabolic Phenotype Associated with Depression by 16s rRNA Gene Sequencing and Lc/Ms-Based Metabolomics. J. Pharm. Biomed. Anal. 2017, 138, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wang, T.; Hu, X.; Luo, J.; Li, W.; Wu, X.; Duan, Y.; Jin, F. Administration of Lactobacillus Helveticus Ns8 Improves Behavioral, Cognitive, and Biochemical Aberrations Caused by Chronic Restraint Stress. Neuroscience 2015, 310, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human Gut Microbiome Viewed across Age and Geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, T.; Liang, S.; Li, W.; Wu, X.; Jin, F. Antibiotic-Induced Imbalances in Gut Microbiota Aggravates Cholesterol Accumulation and Liver Injuries in Rats Fed a High-Cholesterol Diet. Appl. Microbiol. Biotechnol. 2015, 99, 9111–9122. [Google Scholar] [CrossRef] [PubMed]
- Frohlich, E.E.; Farzi, A.; Mayerhofer, R.; Reichmann, F.; Jacan, A.; Wagner, B.; Zinser, E.; Bordag, N.; Magnes, C.; Frohlich, E.; et al. Cognitive Impairment by Antibiotic-Induced Gut Dysbiosis: Analysis of Gut Microbiota-Brain Communication. Brain Behav. Immun. 2016, 56, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Bercik, P.; Collins, S.M. The Effects of Inflammation, Infection and Antibiotics on the Microbiota-Gut-Brain Axis. Adv. Exp. Med. Biol. 2014, 817, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hu, X.; Liang, S.; Li, W.; Wu, X.; Wang, L.; Jin, F. Lactobacillus Fermentum Ns9 Restores the Antibiotic Induced Physiological and Psychological Abnormalities in Rats. Benef. Microbes 2015, 6, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Lurie, I.; Yang, Y.X.; Haynes, K.; Mamtani, R.; Boursi, B. Antibiotic Exposure and the Risk for Depression, Anxiety, or Psychosis: A Nested Case-Control Study. J. Clin. Psychiatry 2015, 76, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Kohler, O.; Petersen, L.; Mors, O.; Mortensen, P.B.; Yolken, R.H.; Gasse, C.; Benros, M.E. Infections and Exposure to Anti-Infective Agents and the Risk of Severe Mental Disorders: A Nationwide Study. Acta Psychiatr. Scand. 2017, 135, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Slykerman, R.F.; Thompson, J.; Waldie, K.E.; Murphy, R.; Wall, C.; Mitchell, E.A. Antibiotics in the First Year of Life and Subsequent Neurocognitive Outcomes. Acta Paediatr. 2017, 106, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Turco, F.; Iannotta, M.; De Gregorio, D.; Palumbo, I.; Sarnelli, G.; Furiano, A.; Napolitano, F.; Boccella, S.; Luongo, L.; et al. Antibiotic-Induced Microbiota Perturbation Causes Gut Endocannabinoidome Changes, Hippocampal Neuroglial Reorganization and Depression in Mice. Brain Behav. Immun. 2018, 67, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Holdeman, L.V.; Good, I.J.; Moore, W.E. Human Fecal Flora Variation in Bacterial Composition within Individuals and a Possible Effect of Emotional Stress. Appl. Environ. Microbiol. 1976, 31, 359–375. [Google Scholar] [PubMed]
- Galley, J.D.; Nelson, M.C.; Yu, Z.T.; Dowd, S.E.; Walter, J.; Kumar, P.S.; Lyte, M.; Bailey, M.T. Exposure to a Social Stressor Disrupts the Community Structure of the Colonic Mucosa-Associated Microbiota. BMC Microbiol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Coe, C.L. Maternal Separation Disrupts the Integrity of the Intestinal Microflora in Infant Rhesus Monkeys. Dev. Psychobiol. 1999, 35, 146–155. [Google Scholar] [CrossRef]
- Gur, T.L.; Worly, B.L.; Bailey, M.T. Stress and the Commensal Microbiota: Importance in Parturition and Infant Neurodevelopment. Front. Psychiatry 2015, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. Regulation of the Stress Response by the Gut Microbiota: Implications for Psychoneuroendocrinology. Psychoneuroendocrinology 2012, 37, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Collins, S.M.; Bercik, P.; Verdu, E.F. The Microbiota-Gut-Brain Axis in Gastrointestinal Disorders: Stressed Bugs, Stressed Brain or Both? J. Physiol. 2014, 592, 2989–2997. [Google Scholar] [CrossRef] [PubMed]
- Marin, I.A.; Goertz, J.E.; Ren, T.; Rich, S.S.; Onengut-Gumuscu, S.; Farber, E.; Wu, M.; Overall, C.C.; Kipnis, J.; Gaultier, A. Microbiota Alteration Is Associated with the Development of Stress-Induced Despair Behavior. Sci. Rep. 2017, 7, 43859. [Google Scholar] [CrossRef] [PubMed]
- Frei, R.; Lauener, R.P.; Crameri, R.; O’mahony, L. Microbiota and Dietary Interactions—An Update to the Hygiene Hypothesis? Allergy 2012, 67, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-Induced Extinctions in the Gut Microbiota Compound over Generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, K.V.; Sherwin, E.; Schellekens, H.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Feeding the Microbiota-Gut-Brain Axis: Diet, Microbiome, and Neuropsychiatry. Transl. Res. 2017, 179, 223–244. [Google Scholar] [CrossRef] [PubMed]
- Slyepchenko, A.; Maes, M.; Jacka, F.N.; Köhler, C.A.; Barichello, T.; Mcintyre, R.S.; Berk, M.; Grande, I.; Foster, J.A.; Vieta, E.; et al. Gut Microbiota, Bacterial Translocation, and Interactions with Diet: Pathophysiological Links between Major Depressive Disorder and Non-Communicable Medical Comorbidities. Psychother. Psychosom. 2017, 86, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Owen, L.; Corfe, B. The Role of Diet and Nutrition on Mental Health and Wellbeing. Proc. Nutr. Soc. 2017, 76, 425–426. [Google Scholar] [CrossRef] [PubMed]
- Noble, E.E.; Hsu, T.M.; Kanoski, S.E. Gut to Brain Dysbiosis: Mechanisms Linking Western Diet Consumption, The Microbiome, and Cognitive Impairment. Front. Behav. Neurosci. 2017, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Pyndt Jørgensen, B.; Hansen, J.T.; Krych, L.; Larsen, C.; Klein, A.B.; Nielsen, D.S.; Josefsen, K.; Hansen, A.K.; Sørensen, D.B. A Possible Link between Food and Mood: Dietary Impact on Gut Microbiota and Behavior in Balb/C Mice. PLoS ONE 2014, 9, E103398. [Google Scholar] [CrossRef]
- Oriach, C.S.; Robertson, R.C.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Food for Thought: The Role of Nutrition in the Microbiota-Gut–Brain Axis. Clin. Nutr. Exp. 2016, 6, 25–38. [Google Scholar] [CrossRef]
- Ng, K.M.; Ferreyra, J.A.; Higginbottom, S.K.; Lynch, J.B.; Kashyap, P.C.; Gopinath, S.; Naidu, N.; Choudhury, B.; Weimer, B.C.; Monack, D.M.; et al. Microbiota-Liberated Host Sugars Facilitate Post-Antibiotic Expansion of Enteric Pathogens. Nature 2013, 502, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Roca-Saavedra, P.; Mendez-Vilabrille, V.; Miranda, J.M.; Nebot, C.; Cardelle-Cobas, A.; Franco, C.M.; Cepeda, A. Food Additives, Contaminants and Other Minor Components: Effects on Human Gut Microbiota—A Review. J. Physiol. Biochem. 2017, 71, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain-Gut-Microbe Communication in Health and Disease. Front. Physiol. 2011, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, F. Linking Lifestyle with Microbiota and Risk of Chronic Inflammatory Disorders. In The Hygiene Hypothesis and Darwinian Medicine; Rook, G.A.W., Ed.; Birkhäuser: Basel, Switzerland, 2009; pp. 93–102. [Google Scholar]
- Liu, X.; Cao, S.; Zhang, X. Modulation of Gut Microbiota-Brain Axis by Probiotics, Prebiotics, and Diet. J. Agric. Food Chem. 2015, 63, 7885–7895. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, G.; Ianiro, G.; Bibbo, S.; Gasbarrini, A. Gut Microbiota Modulation: Probiotics, Antibiotics or Fecal Microbiota Transplantation? Intern. Emerg. Med. 2014, 9, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Marques, T.M.; Cryan, J.F.; Shanahan, F.; Fitzgerald, G.F.; Ross, R.P.; Dinan, T.G.; Stanton, C. Gut Microbiota Modulation and Implications for Host Health: Dietary Strategies to Influence the Gut–Brain Axis. Innov. Food Sci. Emerg. Technol. 2014, 22, 239–247. [Google Scholar] [CrossRef]
- FAO; WHO. Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A Novel Class of Psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Akkasheh, G.; Kashani-Poor, Z.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akbari, H.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z.; Esmaillzadeh, A. Clinical and Metabolic Response to Probiotic Administration in Patients with Major Depressive Disorder: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrition 2016, 32, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Wallace, C.J.K.; Milev, R. The Effects of Probiotics on Depressive Symptoms in Humans: A Systematic Review. Ann. Gen. Psychiatry 2017, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- Pirbaglou, M.; Katz, J.; De Souza, R.J.; Stearns, J.C.; Motamed, M.; Ritvo, P. Probiotic Supplementation Can Positively Affect Anxiety and Depressive Symptoms: A Systematic Review of Randomized Controlled Trials. Nutr. Res. 2016, 36, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Abildgaard, A.; Elfving, B.; Hokland, M.; Wegener, G.; Lund, S. Probiotic Treatment Reduces Depressive-Like Behaviour in Rats Independently of Diet. Psychoneuroendocrinology 2017, 79, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Kiely, B.; Cryan, J.F.; Dinan, T.G. Effects of the Probiotic Bifidobacterium Infantis in the Maternal Separation Model of Depression. Neuroscience 2010, 170, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (Isapp) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Cowen, P.J.; Harmer, C.J.; Tzortzis, G.; Errington, S.; Burnet, P.W.J. Prebiotic Intake Reduces the Waking Cortisol Response and Alters Emotional Bias in Healthy Volunteers. Psychopharmacology 2015, 232, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Robertson, R.C.; Seira Oriach, C.; Murphy, K.; Moloney, G.M.; Cryan, J.F.; Dinan, T.G.; Paul Ross, R.; Stanton, C. Omega-3 Polyunsaturated Fatty Acids Critically Regulate Behaviour and Gut Microbiota Development in Adolescence and Adulthood. Brain Behav. Immun. 2017, 59, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Mika, A.; Day, H.E.; Martinez, A.; Rumian, N.L.; Greenwood, B.N.; Chichlowski, M.; Berg, B.M.; Fleshner, M. Early Life Diets with Prebiotics and Bioactive Milk Fractions Attenuate the Impact of Stress on Learned Helplessness Behaviours and Alter Gene Expression within Neural Circuits Important for Stress Resistance. Eur. J. Neurosci. 2017, 45, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.; Dias, G.P.; Thuret, S. Effects of Diet on Brain Plasticity in Animal and Human Studies: Mind the Gap. Neural Plast. 2014, 2014, 563160. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.L.; Greenway, F.L. A Healthy Gastrointestinal Microbiome Is Dependent on Dietary Diversity. Mol. Metab. 2016, 5, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Evrensel, A.; Ceylan, M.E. Fecal Microbiota Transplantation and Its Usage in Neuropsychiatric Disorders. Clin. Psychopharmacol. Neurosci. 2016, 14, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Kali, A. Psychobiotics: An Emerging Probiotic in Psychiatric Practice. Biomed. J. 2016, 39, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Macedo, D.; Filho, A.; Soares De Sousa, C.N.; Quevedo, J.; Barichello, T.; Junior, H.V.N.; Freitas De Lucena, D. Antidepressants, Antimicrobials or Both? Gut Microbiota Dysbiosis in Depression and Possible Implications of the Antimicrobial Effects of Antidepressant Drugs for Antidepressant Effectiveness. J. Affect. Disord. 2017, 208, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.F.; Ferreira Rocha, N.B.; Paes, F.; Arias-Carrión, O.; Machado, S.; de Sá Filho, A.S. Neural Mechanisms of Exercise—Effects on Gut Miccrobiota and Depression. CNS Neurol. Disord. Drug Targets 2015, 14, 1312–1314. [Google Scholar] [CrossRef] [PubMed]
- Hallgren, M.; Herring, M.P.; Owen, N.; Dunstan, D.; Ekblom, O.; Helgadottir, B.; Nakitanda, O.A.; Forsell, Y. Exercise, Physical Activity, And Sedentary Behavior in the Treatment of Depression: Broadening the Scientific Perspectives and Clinical Opportunities. Front. Psychiatry 2016, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Schnorr, S.L.; Bachner, H.A. Integrative Therapies in Anxiety Treatment with Special Emphasis on the Gut Microbiome. Yale J. Biol. Med. 2016, 89, 397–422. [Google Scholar] [PubMed]
- Bambling, M.; Edwards, S.C.; Hall, S.; Vitetta, L. A Combination of Probiotics and Magnesium Orotate Attenuate Depression in a Small Ssri Resistant Cohort: An Intestinal Anti-Inflammatory Response Is Suggested. Inflammopharmacology 2017, 25, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Bain, L.; Stroud, C. Enabling Discovery, Development, And Translation Of Treatments For Cognitive Dysfunction In Depression: Workshop Summary; National Academies Press: Washington, DC, USA, 2015. [Google Scholar] [CrossRef]
- Garay, R.P.; Zarate, C.A., Jr.; Charpeaud, T.; Citrome, L.; Correll, C.U.; Hameg, A.; Llorca, P.M. Investigational Drugs in Recent Clinical Trials for Treatment-Resistant Depression. Expert Rev. Neurother. 2017, 17, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Hyman, S. Mental Health: Depression Needs Large Human-Genetics Studies. Nature 2014, 515, 189–191. [Google Scholar] [CrossRef] [PubMed]
- The Economist Intelligence Unit. Provision for Supporting People with Mental Illness: A Comparison of 15 Asia Pacific Countries; The Economist Intelligence Unit: London, UK, 2016. [Google Scholar]
- Dinan, T.G.; Stilling, R.M.; Stanton, C.; Cryan, J.F. Collective Unconscious: How Gut Microbes Shape Human Behavior. J. Psychiatr. Res. 2015, 63, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Parashar, A.; Udayabanu, M. Gut Microbiota Regulates Key Modulators of Social Behavior. Eur. Neuropsychopharmacol. 2016, 26, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Yang, Y.; Zhu, W. Gut Microbiota: The Brain Peacekeeper. Front. Microbiol. 2016, 7, 345. [Google Scholar] [CrossRef] [PubMed]
- Bested, A.C.; Logan, A.C.; Selhub, E.M. Intestinal Microbiota, Probiotics and Mental Health: From Metchnikoff to Modern Advances: Part III—Convergence toward Clinical Trials. Gut Pathog. 2013, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bested, A.C.; Logan, A.C.; Selhub, E.M. Intestinal Microbiota, Probiotics and Mental Health: From Metchnikoff to Modern Advances: Part I—Autointoxication Revisited. Gut Pathog. 2013, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bested, A.C.; Logan, A.C.; Selhub, E.M. Intestinal Microbiota, Probiotics and Mental Health: From Metchnikoff to Modern Advances: Part II—Contemporary Contextual Research. Gut Pathog. 2013, 5, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.M.; Surette, M.; Bercik, P. The Interplay between the Intestinal Microbiota and the Brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Bercik, P.; Collins, S.M.; Verdu, E.F. Microbes and the Gut-Brain Axis. Neurogastroenterol. Motil. 2012, 24, 405–413. [Google Scholar] [CrossRef] [PubMed]
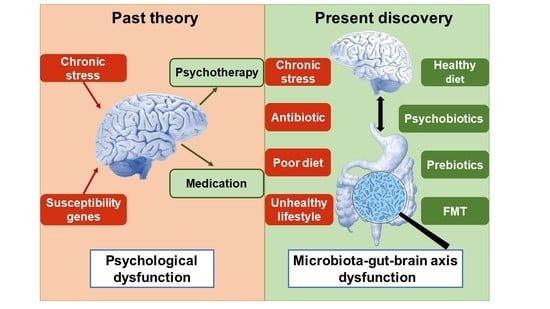
Figure 1.
The main pathophysiology and therapy targets of depression. Microbiota–gut–brain axis dysfunction is the main pathophysiology and potential treatment target of major depression. It includes brain dysfunction and gut brain dysfunction. The light red frames in the top and bottom are risk factors of depression, while the light green frames are therapies of depression. EC, enteroendocrine cell; ENS, enteric nervous system; MC, mast cell.
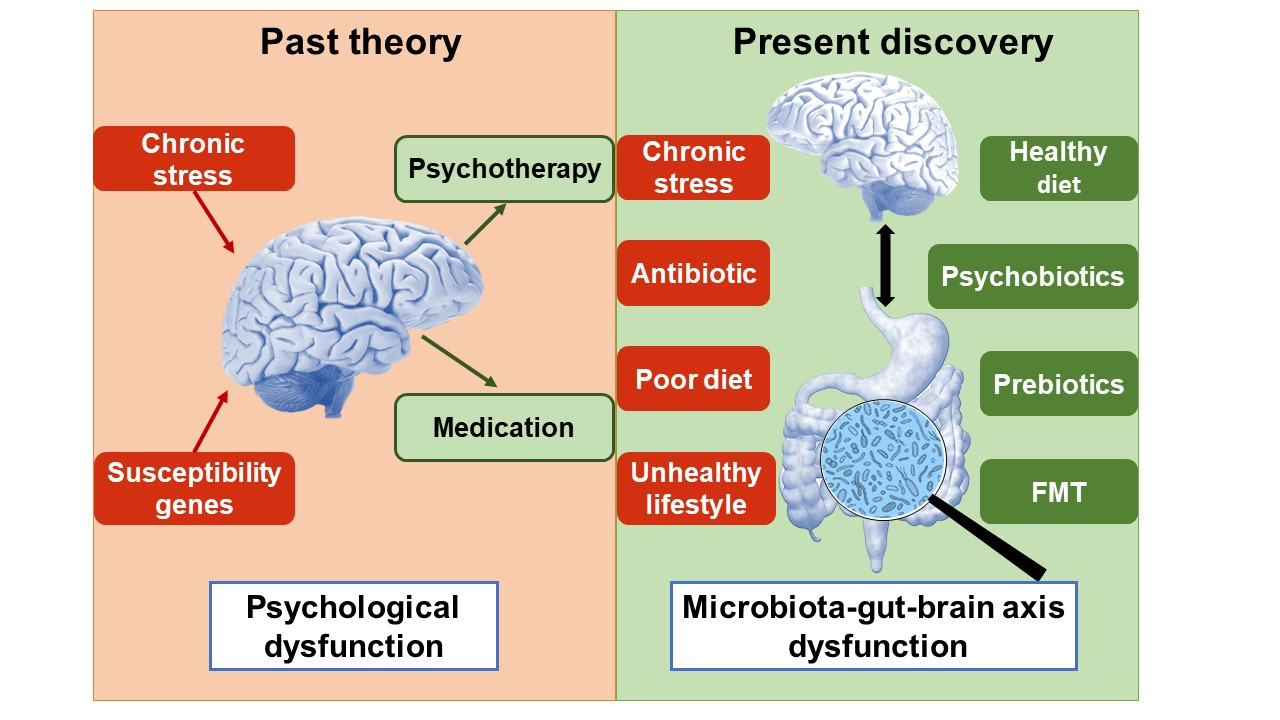
Figure 1.
The main pathophysiology and therapy targets of depression. Microbiota–gut–brain axis dysfunction is the main pathophysiology and potential treatment target of major depression. It includes brain dysfunction and gut brain dysfunction. The light red frames in the top and bottom are risk factors of depression, while the light green frames are therapies of depression. EC, enteroendocrine cell; ENS, enteric nervous system; MC, mast cell.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liang, S.; Wu, X.; Hu, X.; Wang, T.; Jin, F. Recognizing Depression from the Microbiota–Gut–Brain Axis. Int. J. Mol. Sci. 2018, 19, 1592. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061592
AMA Style
Liang S, Wu X, Hu X, Wang T, Jin F. Recognizing Depression from the Microbiota–Gut–Brain Axis. International Journal of Molecular Sciences. 2018; 19(6):1592. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061592
Chicago/Turabian StyleLiang, Shan, Xiaoli Wu, Xu Hu, Tao Wang, and Feng Jin. 2018. "Recognizing Depression from the Microbiota–Gut–Brain Axis" International Journal of Molecular Sciences 19, no. 6: 1592. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061592
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.