Exercise Training Has Contrasting Effects in Myocardial Infarction and Pressure Overload Due to Divergent Endothelial Nitric Oxide Synthase Regulation


Abstract
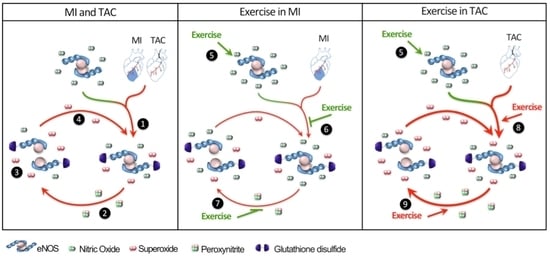
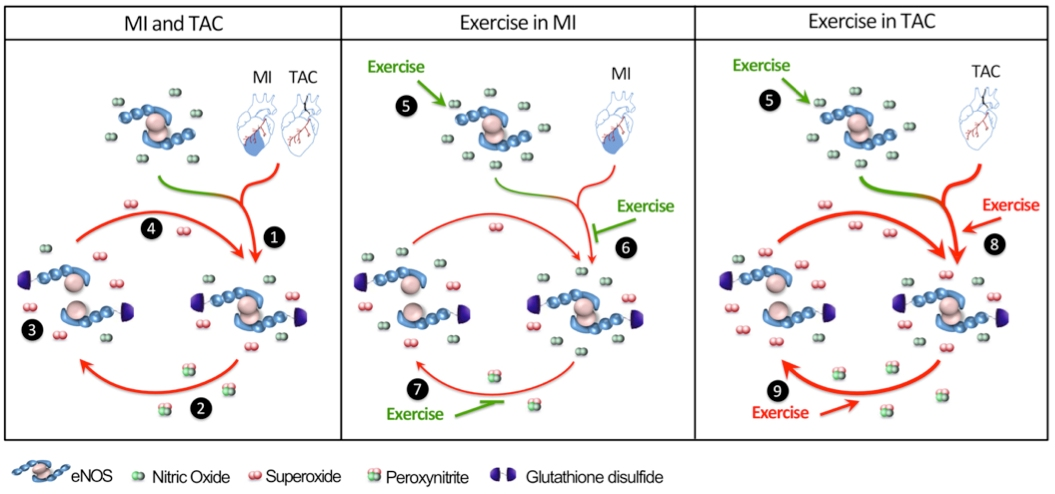
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Exercise and Survival
2.2. LV Remodeling and Dysfunction
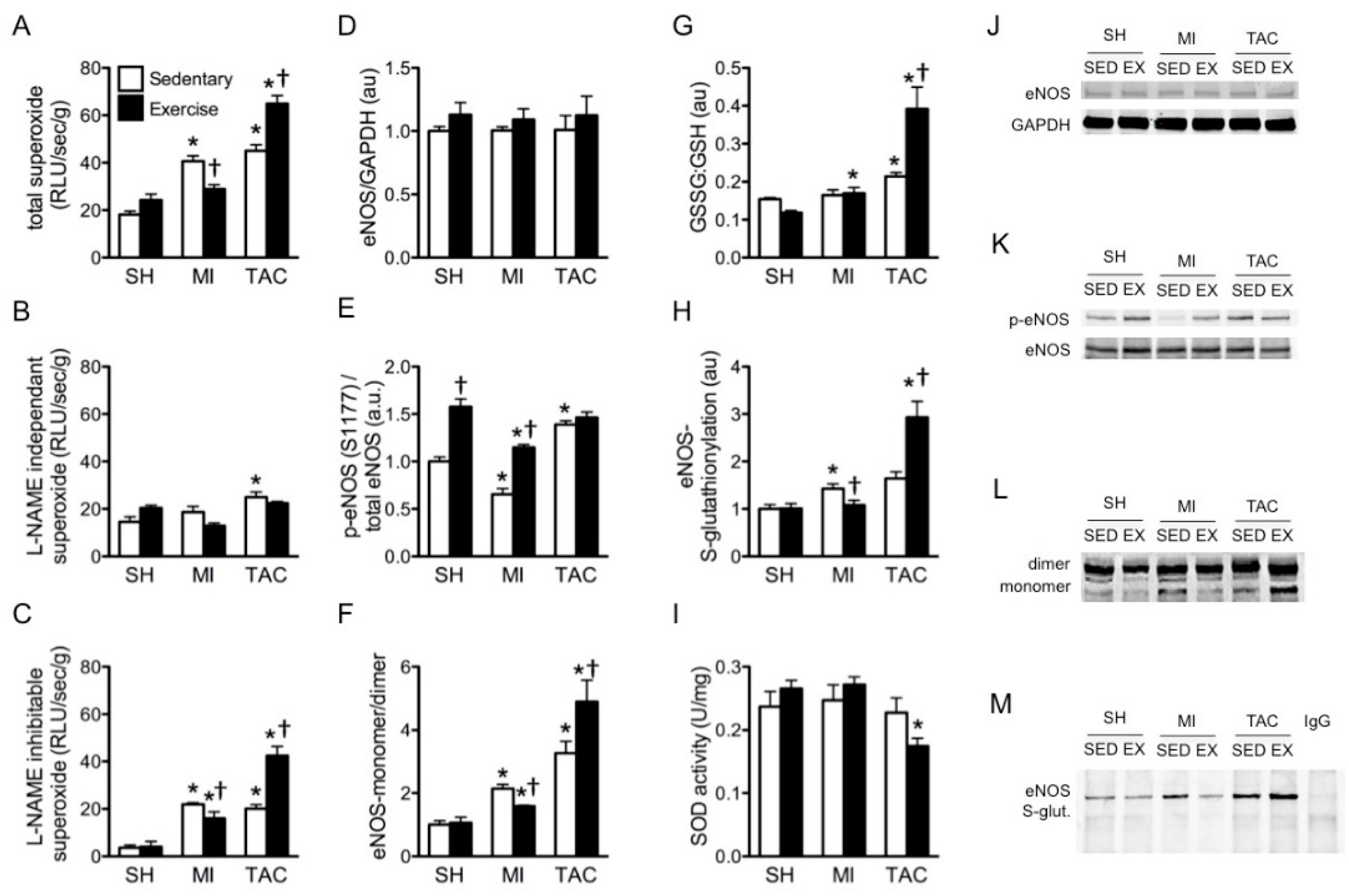
2.3. Superoxide Production
2.4. Total eNOS and Phosphorylated eNOS (p-eNOS) Protein Expression
2.5. eNOS Uncoupling and eNOS S-Glutathionylation
2.6. Superoxide Dismutase (SOD) Activity
3. Discussion
4. Materials and Methods
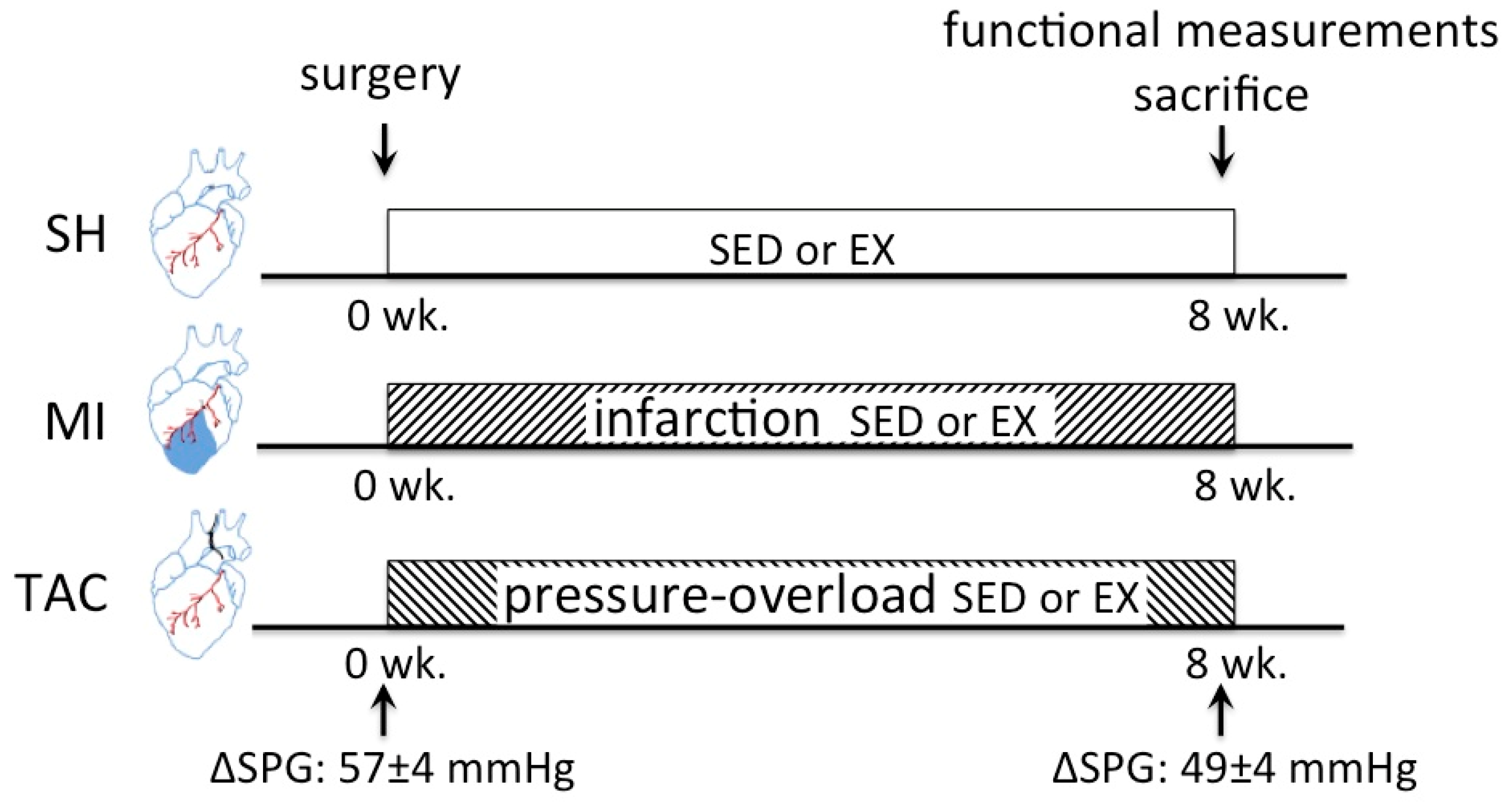
4.1. Animal Experiments
4.2. Cardiac Function and Geometry Measurements
4.3. Histology
4.4. Detection of Superoxide Production
4.5. Total and Phosphorylated eNOS Protein Analysis
4.6. Quantification of GSSG and GSH
4.7. eNOS S-Glutathionylation
4.8. eNOS Monomer–Dimer Ratio
4.9. SOD Activity
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; de Ferranti, S.; Despres, J.P.; Fullerton, H.J.; Howard, V.J.; et al. Heart disease and stroke statistics—2015 update: A report from the American Heart Association. Circulation 2015, 131, e29–e322. [Google Scholar] [CrossRef] [PubMed]
- Ambrosy, A.P.; Fonarow, G.C.; Butler, J.; Chioncel, O.; Greene, S.J.; Vaduganathan, M.; Nodari, S.; Lam, C.S.; Sato, N.; Shah, A.N.; et al. The global health and economic burden of hospitalizations for heart failure: Lessons learned from hospitalized heart failure registries. J. Am. Coll. Cardiol. 2014, 63, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.; Balady, G.J. The role of exercise training in heart failure. J. Am. Coll. Cardiol. 2011, 58, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, G.F.; Balady, G.; Blair, S.N.; Blumenthal, J.; Caspersen, C.; Chaitman, B.; Epstein, S.; Sivarajan Froelicher, E.S.; Froelicher, V.F.; Pina, I.L.; et al. Statement on exercise: Benefits and recommendations for physical activity programs for all Americans. A statement for health professionals by the Committee on Exercise and Cardiac Rehabilitation of the Council on Clinical Cardiology, American Heart Association. Circulation 1996, 94, 857–862. [Google Scholar] [PubMed]
- O’Connor, C.M.; Whellan, D.J.; Lee, K.L.; Keteyian, S.J.; Cooper, L.S.; Ellis, S.J.; Leifer, E.S.; Kraus, W.E.; Kitzman, D.W.; Blumenthal, J.A.; et al. Efficacy and safety of exercise training in patients with chronic heart failure: HF-ACTION randomized controlled trial. JAMA 2009, 301, 1439–1450. [Google Scholar] [CrossRef] [PubMed]
- Davies, E.J.; Moxham, T.; Rees, K.; Singh, S.; Coats, A.J.; Ebrahim, S.; Lough, F.; Taylor, R.S. Exercise training for systolic heart failure: Cochrane systematic review and meta-analysis. Eur. J. Heart Fail. 2010, 12, 706–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gielen, S.; Laughlin, M.H.; O’Conner, C.; Duncker, D.J. Exercise training in patients with heart disease: Review of beneficial effects and clinical recommendations. Prog. Cardiovasc. Dis. 2015, 57, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Farah, C.; Kleindienst, A.; Bolea, G.; Meyer, G.; Gayrard, S.; Geny, B.; Obert, P.; Cazorla, O.; Tanguy, S.; Reboul, C. Exercise-induced cardioprotection: A role for eNOS uncoupling and NO metabolites. Basic Res. Cardiol. 2013, 108, 389. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Smuder, A.J.; Kavazis, A.N.; Quindry, J.C. Mechanisms of exercise-induced cardioprotection. Physiology 2014, 29, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Mann, N.; Rosenzweig, A. Can exercise teach us how to treat heart disease? Circulation 2012, 126, 2625–2635. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, M.H.; Davis, M.J.; Secher, N.H.; van Lieshout, J.J.; Arce-Esquivel, A.A.; Simmons, G.H.; Bender, S.B.; Padilla, J.; Bache, R.J.; Merkus, D.; et al. Peripheral circulation. Compr. Physiol. 2012, 2, 321–447. [Google Scholar] [PubMed]
- Balligand, J.L.; Feron, O.; Dessy, C. eNOS activation by physical forces: From short-term regulation of contraction to chronic remodeling of cardiovascular tissues. Physiol. Rev. 2009, 89, 481–534. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L. Modulation of skeletal muscle antioxidant defense by exercise: Role of redox signaling. Free Radic. Biol. Med. 2008, 44, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Linke, A.; Adams, V.; Schulze, P.C.; Erbs, S.; Gielen, S.; Fiehn, E.; Mobius-Winkler, S.; Schubert, A.; Schuler, G.; Hambrecht, R. Antioxidative effects of exercise training in patients with chronic heart failure: Increase in radical scavenger enzyme activity in skeletal muscle. Circulation 2005, 111, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Bito, V.; de Waard, M.C.; Biesmans, L.; Lenaerts, I.; Ozdemir, S.; van Deel, E.; Abdel-Mottaleb, Y.; Driesen, R.; Holemans, P.; Duncker, D.J.; et al. Early exercise training after myocardial infarction prevents contractile but not electrical remodelling or hypertrophy. Cardiovasc. Res. 2010, 86, 72–81. [Google Scholar] [CrossRef] [PubMed]
- De Waard, M.C.; van der Velden, J.; Bito, V.; Ozdemir, S.; Biesmans, L.; Boontje, N.M.; Dekkers, D.H.; Schoonderwoerd, K.; Schuurbiers, H.C.; de Crom, R.; et al. Early exercise training normalizes myofilament function and attenuates left ventricular pump dysfunction in mice with a large myocardial infarction. Circ. Res. 2007, 100, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Duncker, D.J.; van Deel, E.D.; de Waard, M.C.; de Boer, M.; Merkus, D.; van der Velden, J. Exercise training in adverse cardiac remodeling. Pflugers Arch. 2014, 466, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- Rolim, N.; Skardal, K.; Hoydal, M.; Sousa, M.M.; Malmo, V.; Kaurstad, G.; Ingul, C.B.; Hansen, H.E.; Alves, M.N.; Thuen, M.; et al. Aerobic interval training reduces inducible ventricular arrhythmias in diabetic mice after myocardial infarction. Basic. Res. Cardiol. 2015, 110, 44. [Google Scholar] [CrossRef] [PubMed]
- De Waard, M.C.; van Haperen, R.; Soullie, T.; Tempel, D.; de Crom, R.; Duncker, D.J. Beneficial effects of exercise training after myocardial infarction require full eNOS expression. J. Mol. Cell. Cardiol. 2010, 48, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Haykowsky, M.; Scott, J.; Esch, B.; Schopflocher, D.; Myers, J.; Paterson, I.; Warburton, D.; Jones, L.; Clark, A.M. A meta-analysis of the effects of exercise training on left ventricular remodeling following myocardial infarction: Start early and go longer for greatest exercise benefits on remodeling. Trials 2011, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Bonow, R.O.; Cheitlin, M.D.; Crawford, M.H.; Douglas, P.S. Task Force 3: Valvular heart disease. J. Am. Coll. Cardiol. 2005, 45, 1334–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rome, J.J. Exercise restriction to prevent sudden death in congenital aortic stenosis: Whom are we treating? J. Am. Coll. Cardiol. 2010, 56, 1947–1948. [Google Scholar] [CrossRef] [PubMed]
- Van Deel, E.D.; de Boer, M.; Kuster, D.W.; Boontje, N.M.; Holemans, P.; Sipido, K.R.; van der Velden, J.; Duncker, D.J. Exercise training does not improve cardiac function in compensated or decompensated left ventricular hypertrophy induced by aortic stenosis. J. Mol. Cell. Cardiol. 2011, 50, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.A.; Wang, T.Y.; Varadharaj, S.; Reyes, L.A.; Hemann, C.; Talukder, M.A.; Chen, Y.R.; Druhan, L.J.; Zweier, J.L. S-glutathionylation uncouples eNOS and regulates its cellular and vascular function. Nature 2010, 468, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Gielen, S.; Schuler, G.; Adams, V. Cardiovascular effects of exercise training: Molecular mechanisms. Circulation 2010, 122, 1221–1238. [Google Scholar] [CrossRef] [PubMed]
- Van den Borne, S.W.; van de Schans, V.A.; Strzelecka, A.E.; Vervoort-Peters, H.T.; Lijnen, P.M.; Cleutjens, J.P.; Smits, J.F.; Daemen, M.J.; Janssen, B.J.; Blankesteijn, W.M. Mouse strain determines the outcome of wound healing after myocardial infarction. Cardiovasc. Res. 2009, 84, 273–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Waard, M.C.; Duncker, D.J. Prior exercise improves survival, infarct healing, and left ventricular function after myocardial infarction. J. Appl. Physiol. 2009, 107, 928–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Jia, Z.; Yang, L.; Zhu, M.; Zhang, J.; Liu, J.; Wu, P.; Tian, W.; Li, J.; Qi, Z.; et al. Exercise Protects against Chronic beta-Adrenergic Remodeling of the Heart by Activation of Endothelial Nitric Oxide Synthase. PLoS ONE 2014, 9, e96892. [Google Scholar]
- Jones, S.P.; Greer, J.J.; van Haperen, R.; Duncker, D.J.; de Crom, R.; Lefer, D.J. Endothelial nitric oxide synthase overexpression attenuates congestive heart failure in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 4891–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Waard, M.C.; van der Velden, J.; Boontje, N.M.; Dekkers, D.H.; van Haperen, R.; Kuster, D.W.; Lamers, J.M.; de Crom, R.; Duncker, D.J. Detrimental effect of combined exercise training and eNOS overexpression on cardiac function after myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1513–H1523. [Google Scholar] [CrossRef] [PubMed]
- Heineke, J.; Molkentin, J.D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nat. Rev. Mol. Cell. Biol. 2006, 7, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.A.; Rush, J.W. Exercise training improves aortic endothelium-dependent vasorelaxation and determinants of nitric oxide bioavailability in spontaneously hypertensive rats. J. Appl. Physiol. 2004, 96, 2088–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, T.; Nakamuta, J.S.; Magalhaes, F.C.; Roque, F.R.; Lavini-Ramos, C.; Schettert, I.T.; Coelho, V.; Krieger, J.E.; Oliveira, E.M. Exercise training restores the endothelial progenitor cells number and function in hypertension: Implications for angiogenesis. J. Hypertens. 2012, 30, 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- Duncker, D.J.; Ishibashi, Y.; Bache, R.J. Effect of treadmill exercise on transmural distribution of blood flow in hypertrophied left ventricle. Am. J. Physiol. 1998, 275, H1274–H1282. [Google Scholar] [CrossRef] [PubMed]
- Haykowsky, M.J.; Liang, Y.; Pechter, D.; Jones, L.W.; McAlister, F.A.; Clark, A.M. A meta-analysis of the effect of exercise training on left ventricular remodeling in heart failure patients: The benefit depends on the type of training performed. J. Am. Coll. Cardiol. 2007, 49, 2329–2336. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.; Nagayama, T.; Koitabashi, N.; Zhang, M.; Zhou, L.; Bedja, D.; Gabrielson, K.L.; Molkentin, J.D.; Kass, D.A.; Takimoto, E. Phosphodiesterase 5 inhibition blocks pressure overload-induced cardiac hypertrophy independent of the calcineurin pathway. Cardiovasc. Res. 2009, 81, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Haskell, W.L.; Lee, I.M.; Pate, R.R.; Powell, K.E.; Blair, S.N.; Franklin, B.A.; Macera, C.A.; Heath, G.W.; Thompson, P.D.; Bauman, A.; et al. Physical activity and public health: Updated recommendation for adults from the American College of Sports Medicine and the American Heart Association. Circulation 2007, 116, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Steg, P.G.; James, S.K.; Atar, D.; Badano, L.P.; Blomstrom-Lundqvist, C.; Borger, M.A.; Di Mario, C.; Dickstein, K.; Ducrocq, G.; Fernandez-Aviles, F.; et al. ESC Guidelines for the management of acute myocardial infarction in patients presenting with ST-segment elevation: The Task Force on the management of ST-segment elevation acute myocardial infarction of the European Society of Cardiology (ESC). Eur. Heart J. 2012, 33, 2569–2619. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Dipilato, A.E.; Chong, E.C.; Gauvreau, K.; McElhinney, D.B.; Colan, S.D.; Lock, J.E. Sudden unexpected death after balloon valvuloplasty for congenital aortic stenosis. J. Am. Coll. Cardiol. 2010, 56, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Van Deel, E.; Ridwan, Y.; van Vliet, J.N.; Belenkov, S.; Essers, J. In Vivo Quantitative Assessment of Myocardial Structure, Function, Perfusion and Viability Using Cardiac Micro-computed Tomography. J. Vis. Exp. 2016, 53603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.M.; Guzik, T.J.; Zhang, Y.H.; Zhang, M.H.; Kattach, H.; Ratnatunga, C.; Pillai, R.; Channon, K.M.; Casadei, B. A myocardial Nox2 containing NAD(P)H oxidase contributes to oxidative stress in human atrial fibrillation. Circ. Res. 2005, 97, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Moens, A.L.; Champion, H.C.; Claeys, M.J.; Tavazzi, B.; Kaminski, P.M.; Wolin, M.S.; Borgonjon, D.J.; Van Nassauw, L.; Haile, A.; Zviman, M.; et al. High-dose folic acid pretreatment blunts cardiac dysfunction during ischemia coupled to maintenance of high-energy phosphates and reduces postreperfusion injury. Circulation 2008, 117, 1810–1819. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Deel, E.D.; Octavia, Y.; De Waard, M.C.; De Boer, M.; Duncker, D.J. Exercise Training Has Contrasting Effects in Myocardial Infarction and Pressure Overload Due to Divergent Endothelial Nitric Oxide Synthase Regulation. Int. J. Mol. Sci. 2018, 19, 1968. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071968
Van Deel ED, Octavia Y, De Waard MC, De Boer M, Duncker DJ. Exercise Training Has Contrasting Effects in Myocardial Infarction and Pressure Overload Due to Divergent Endothelial Nitric Oxide Synthase Regulation. International Journal of Molecular Sciences. 2018; 19(7):1968. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071968
Chicago/Turabian StyleVan Deel, Elza D., Yanti Octavia, Monique C. De Waard, Martine De Boer, and Dirk J. Duncker. 2018. "Exercise Training Has Contrasting Effects in Myocardial Infarction and Pressure Overload Due to Divergent Endothelial Nitric Oxide Synthase Regulation" International Journal of Molecular Sciences 19, no. 7: 1968. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071968