Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR in Macrobrachium Nipponense

 , ,
, ,
Abstract
:
1. Introduction
2. Results
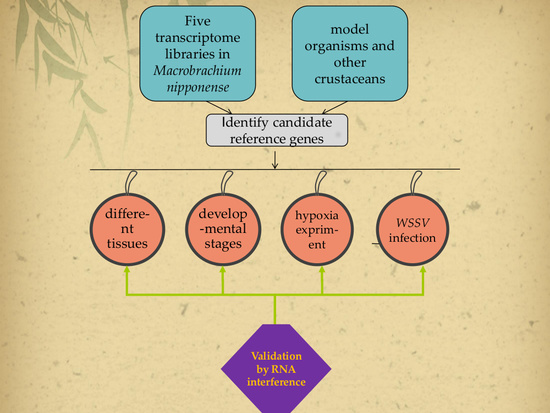
2.1. Selection of Target Internal Reference Genes
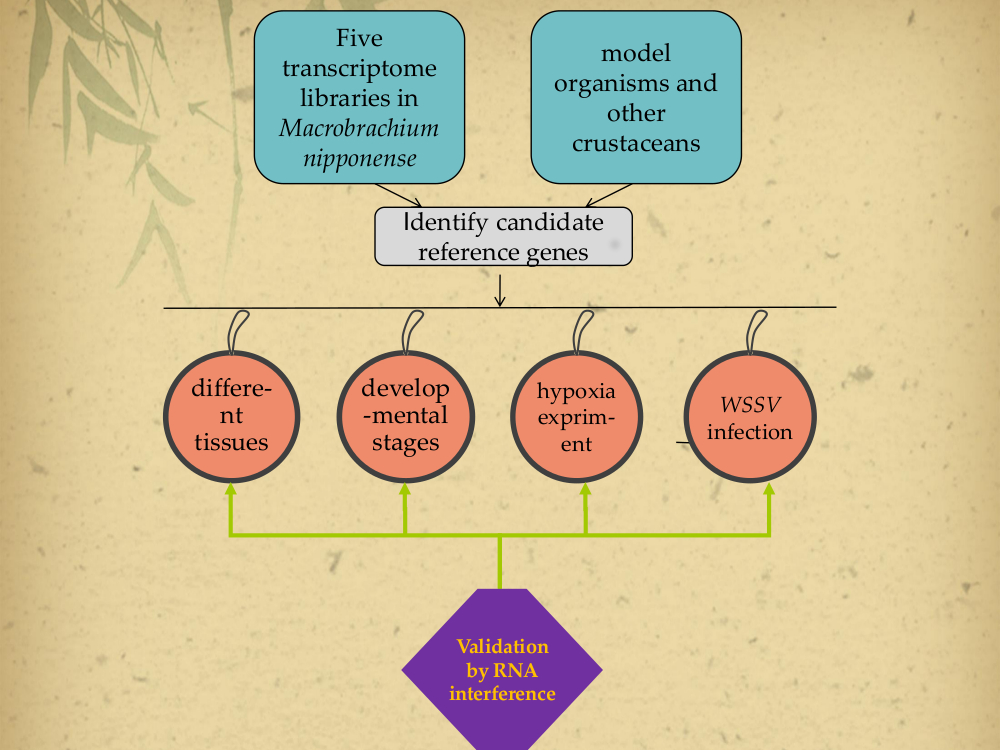
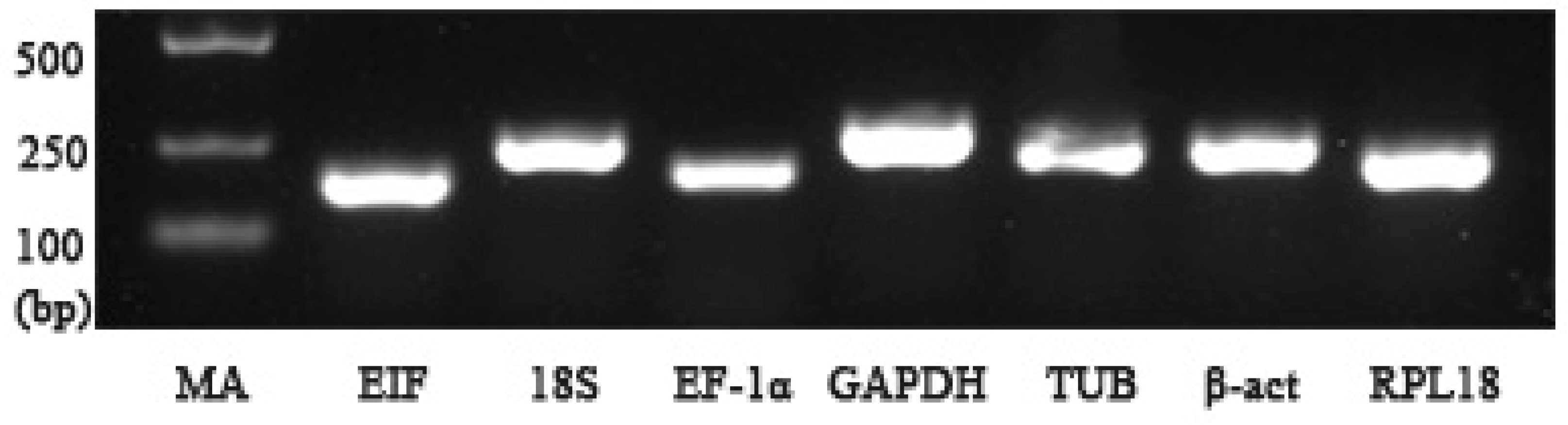
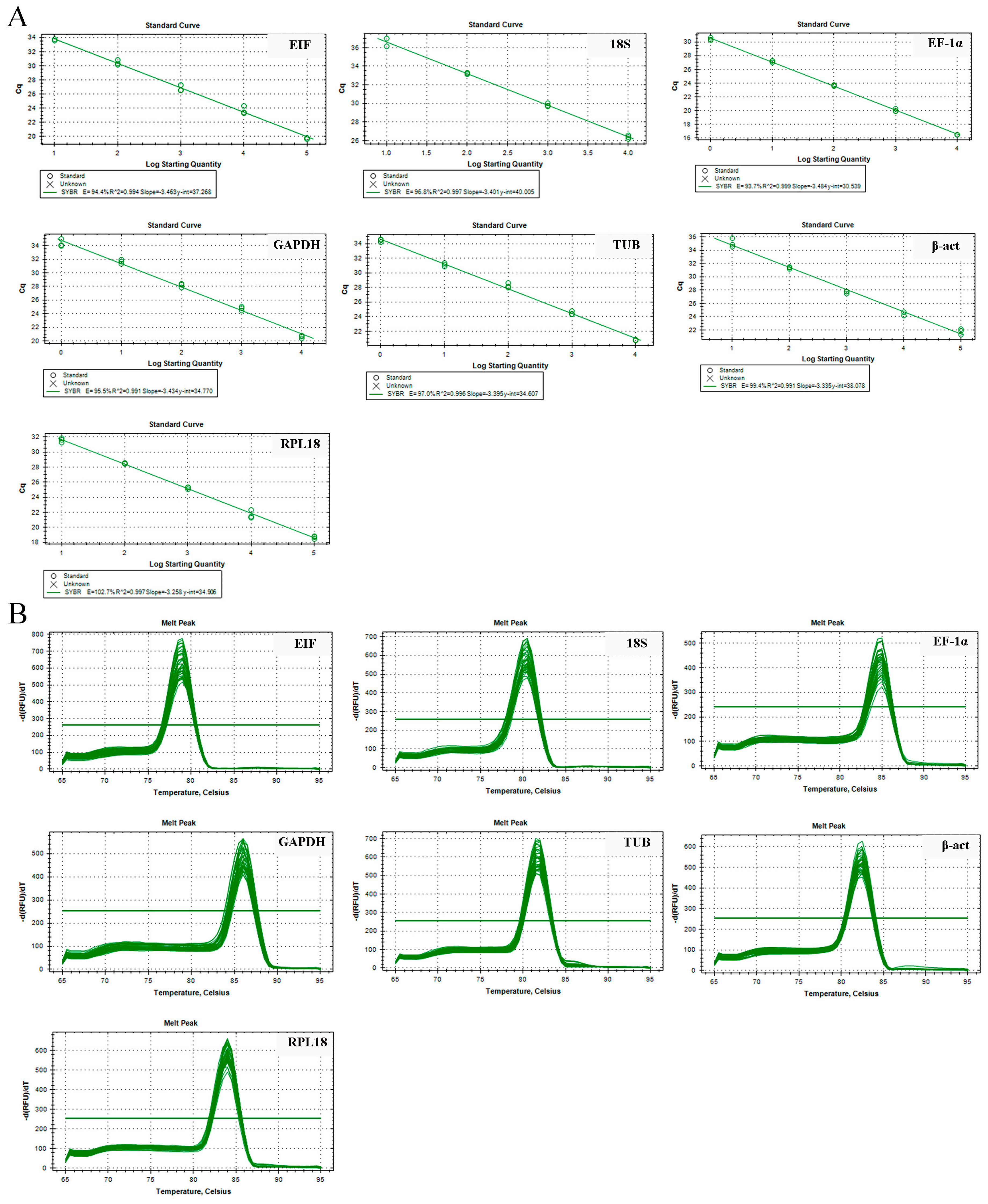
2.2. Primer Specificity and Amplification Efficiency for qPCR
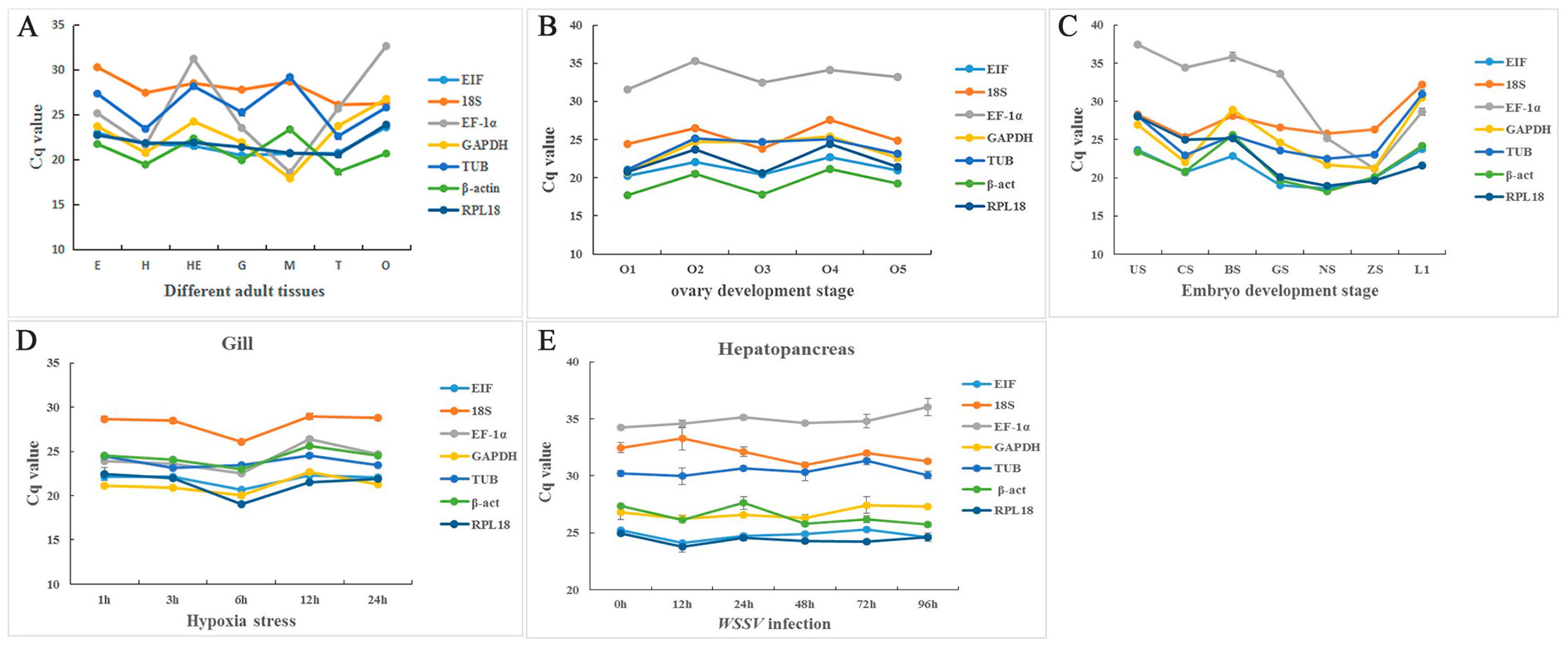
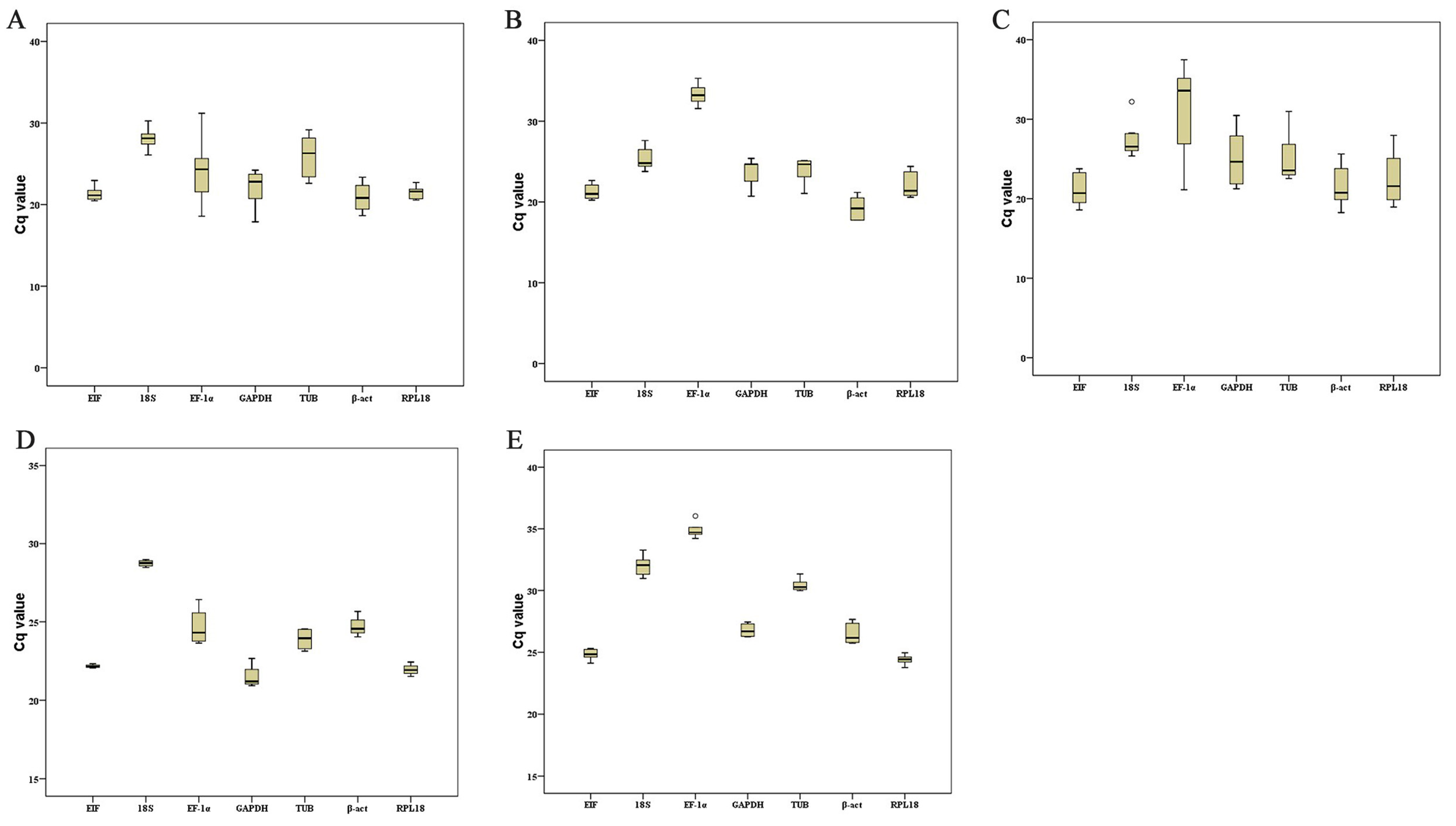
2.3. Distribution of Cycle Threshold (Cq) Values
2.4. Stability Analysis
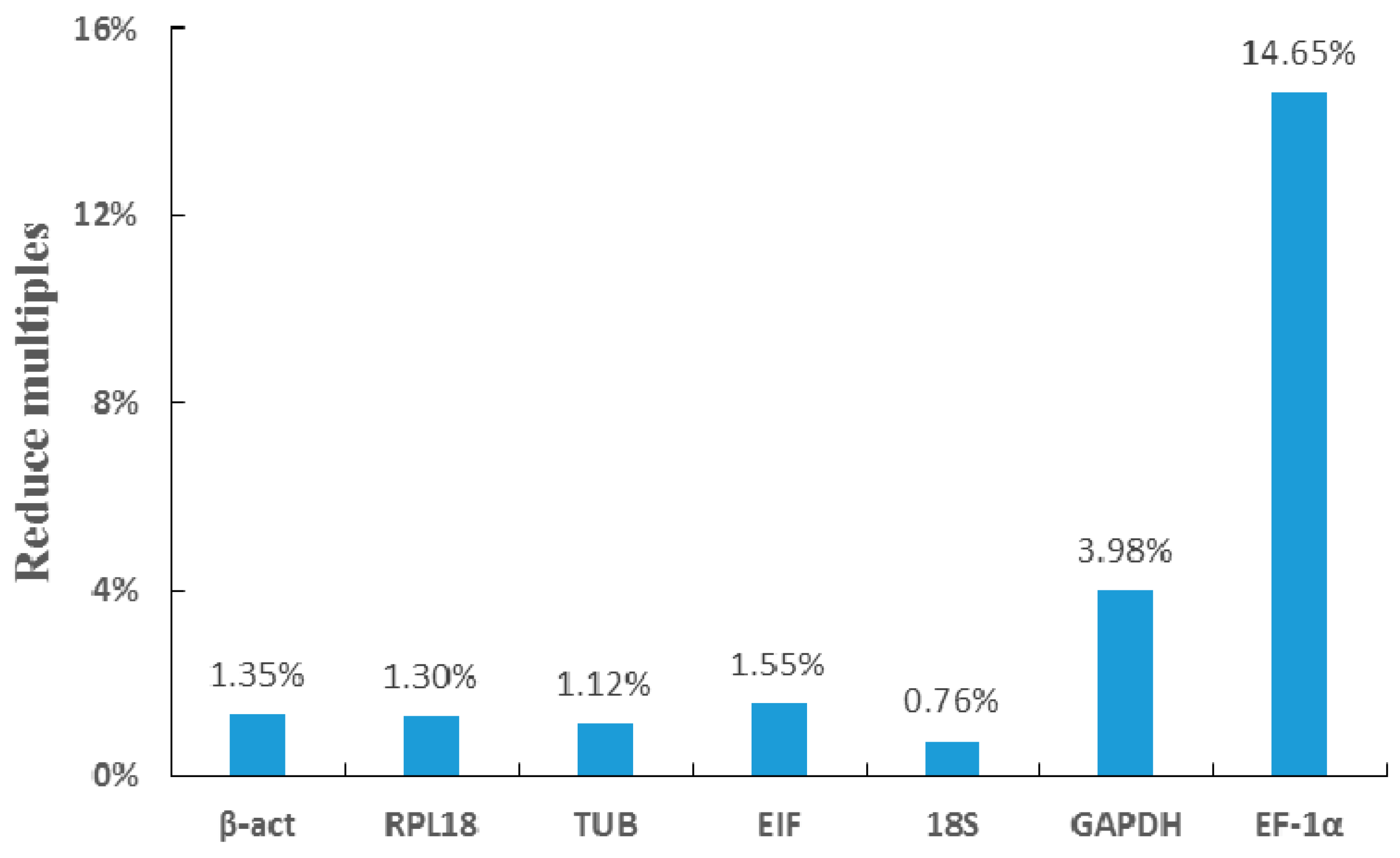
2.5. Validation of the Selected Reference Genes by RNAi
3. Discussion
4. Materials and Methods
4.1. Selection of Reference Genes and Primer Design
4.2. Amplification Efficiency and Primers Specificity of Reference Genes
4.3. Animals, Stress Experiments, and Tissue Collection
4.4. RNA Extraction and cDNA Synthesis
4.5. Quantitative Real-Time PCR (qPCR)
4.6. Methods for Analyzing Stability of Reference Genes
4.7. Validation of Reference Genes by SST Gene RNA Interference
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| qPCR | quantitative real-time reverse transcription PCR |
| ORF | open reading frame |
| RNAi | RNA interference |
| M. nipponense | Macrobrachium nipponense |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Gene Name | Primer Sequence (5′–3′) Forward/Reverse | Genbank Accession No. |
|---|---|---|---|
| EIF | eukaryotic translation initiation factor 5A | GTTGTATGCAGTCGGCCATATTT TGTCCTGAAGGTGGTGATAATGA | MH540106 |
| 18S | 18S ribosomal RNA | CTCCCCCTGAAGTGTATTATGGA GCGAATCTTTTTCAGTTGTTCCC | MH540107 |
| EF-1α | elongation factor 1-alpha | CCTGGTATGGTTGTCACTTTTGC CTCCTTCTTCGACACCTCTTTGA | MH540108 |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase | CCAAGGACATGAAGGTAGTCTCC TTCTGCATGTGCTTCAACAAGTC | MH540109 |
| TUB | α-tubulin | GGTCTGGAATTCAGTCAAGTCG AGTGCATCTCAGTTCATGTTGG | MH540110 |
| β-act | β-actin | CTCTTCTTCCCTGGAGAAGTCTTA ATCCACATCTGTTGGAAGGTAGA | MH540111 |
| RPL18 | ribosomal protein L18 | ACTTTTTGTACCCACAGCTTGAC GTGAAGGGCGAATCTTTGTTGTT | MH540112 |
| dsSST | slow-tonic S2 tropomyosin | TAATACGACTCACTATAGGGCGAGAGGTCTGAGGAACGAC TAATACGACTCACTATAGGGTAAGCTTCCTCACGCTGGTT |
| Experimental Conditions | Method | Rank | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| Different adult tissues | Comparative ∆Ct | RPL18 | EIF | β-act | 18S | TUB | GAPDH | EF-1α |
| BestKeeper | RPL18 | EIF | 18S | β-act | TUB | GAPDH | EF-1α | |
| NormFinder | RPL18 | EIF | β-act | 18S | GAPDH | TUB | EF-1α | |
| GeNorm | EIF/RPL18 | 18S | β-act | TUB | GAPDH | EF-1α | ||
| Recommended comprehensive ranking | RPL18 | EIF | 18S | β-act | TUB | GAPDH | EF-1α | |
| Different ovarian stages | Comparative ∆Ct | β-act | EIF | RPL18 | 18S | EF-1α | TUB | GAPDH |
| BestKeeper | EIF | EF-1α | β-act | 18S | TUB | RPL18 | GAPDH | |
| NormFinder | EIF | β-act | EF-1α | RPL18 | 18S | TUB | GAPDH | |
| GeNorm | 18S/RPL18 | β-act | EIF | EF-1α | TUB | GAPDH | ||
| Recommended comprehensive ranking | EIF | β-act | RPL18 | 18S | EF-1α | TUB | GAPDH | |
| Different embryo stages | Comparative ∆Ct | EIF | β-act | TUB | 18S | GAPDH | RPL18 | EF-1α |
| BestKeeper | 18S | EIF | β-act | TUB | RPL18 | GAPDH | EF-1α | |
| NormFinder | EIF | β-act | GAPDH | TUB | 18S | RPL18 | EF-1α | |
| GeNorm | 18S/TUB | EIF | β-act | GAPDH | RPL18 | EF-1α | ||
| Recommended comprehensive ranking | EIF | 18S | β-act | TUB | GAPDH | RPL18 | EF-1α | |
| Hypoxia stress | Comparative ∆Ct | β-act | GAPDH | EIF | 18S | EF-1α | TUB | RPL18 |
| BestKeeper | EIF | TUB | GAPDH | β-act | 18S | RPL18 | EF-1α | |
| NormFinder | β-act | GAPDH | EIF | 18S | TUB | EF-1α | RPL18 | |
| GeNorm | GAPDH/β-act | EIF | 18S | EF-1α | TUB | RPL18 | ||
| Recommended comprehensive ranking | β-act | GAPDH | EIF | 18S | TUB | EF-1α | RPL18 | |
| WSSV infection | Comparative ∆Ct | EIF | RPL18 | GAPDH | TUB | EF-1α | β-act | 18S |
| BestKeeper | RPL18 | EIF | TUB | GAPDH | EF-1α | 18S | β-act | |
| NormFinder | RPL18 | EIF | GAPDH | TUB | β-act | EF-1α | 18S | |
| GeNorm | EIF /TUB | GAPDH | RPL18 | EF-1α | β-act | 18S | ||
| Recommended comprehensive ranking | EIF | RPL18 | TUB | GAPDH | EF-1α | β-act | 18S | |
| Rank | ∆Ct | BestKeeper | NormFinder | GeNorm | |||||
|---|---|---|---|---|---|---|---|---|---|
| Genes | Average of Std dev | Genes | Std dev | Genes | Stability Index | Genes | Stability Value | ||
| Different adult tissues | 1 | RPL18 | 2.22 | RPL18 | 0.83 | RPL18 | 0.252 | EIF/RPL18 | 0.392 |
| 2 | EIF | 2.24 | EIF | 0.94 | EIF | 0.542 | |||
| 3 | β-act | 2.52 | 18S | 1.10 | β-act | 1.767 | 18S | 1.354 | |
| 4 | 18S | 2.66 | β-act | 1.38 | 18S | 1.934 | β-act | 1.55 | |
| 5 | TUB | 2.91 | TUB | 1.93 | GAPDH | 2.128 | TUB | 1.705 | |
| 6 | GAPDH | 3.07 | GAPDH | 2.16 | TUB | 2.369 | GAPDH | 2.192 | |
| 7 | EF-1α | 4.65 | EF-1α | 3.74 | EF-1α | 4.462 | EF-1α | 2.895 | |
| Different ovarian stages | 1 | β-act | 0.85 | EIF | 0.87 | EIF | 0.439 | 18S/RPL18 | 0.319 |
| 2 | EIF | 0.88 | EF-1α | 1.11 | β-act | 0.506 | |||
| 3 | RPL18 | 0.91 | β-act | 1.25 | EF-1α | 0.609 | β-act | 0.388 | |
| 4 | 18S | 0.95 | 18S | 1.3 | RPL18 | 0.637 | EIF | 0.513 | |
| 5 | EF-1α | 0.99 | TUB | 1.38 | 18S | 0.749 | EF-1α | 0.653 | |
| 6 | TUB | 1.2 | RPL18 | 1.5 | TUB | 1.032 | TUB | 0.892 | |
| 7 | GAPDH | 1.33 | GAPDH | 1.57 | GAPDH | 1.218 | GAPDH | 1.016 | |
| Different embryo stages | 1 | EIF | 2.37 | 18S | 1.89 | EIF | 0.642 | 18S/TUB | 1.165 |
| 2 | β-act | 2.47 | EIF | 1.73 | β-act | 0.786 | |||
| 3 | TUB | 2.74 | β-act | 5.07 | GAPDH | 1.775 | EIF | 1.44 | |
| 4 | 18S | 2.77 | TUB | 3.11 | TUB | 1.906 | β-act | 1.578 | |
| 5 | GAPDH | 2.84 | RPL18 | 2.55 | 18S | 2.059 | GAPDH | 1.724 | |
| 6 | RPL18 | 3.36 | GAPDH | 2.3 | RPL18 | 2.295 | RPL18 | 2.24 | |
| 7 | EF-1α | 5.35 | EF-1α | 2.92 | EF-1α | 5.137 | EF-1α | 3.129 | |
| Hypoxia stress | 1 | β-act | 0.6 | EIF | 0.49 | β-act | 0.092 | GAPDH/β-act | 0.185 |
| 2 | GAPDH | 0.66 | TUB | 0.57 | GAPDH | 0.316 | |||
| 3 | EIF | 0.7 | GAPDH | 0.61 | EIF | 0.323 | EIF | 0.434 | |
| 4 | 18S | 0.74 | β-act | 0.67 | 18S | 0.45 | 18S | 0.532 | |
| 5 | EF-1α | 0.95 | 18S | 0.85 | TUB | 0.822 | EF-1α | 0.631 | |
| 6 | TUB | 0.97 | RPL18 | 0.93 | EF-1α | 0.828 | TUB | 0.72 | |
| 7 | RPL18 | 1.01 | EF-1α | 1.06 | RPL18 | 0.912 | RPL18 | 0.803 | |
| WSSV infection | 1 | EIF | 0.66 | RPL18 | 0.31 | RPL18 | 0.297 | EIF/TUB | 0.397 |
| 2 | RPL18 | 0.67 | EIF | 0.32 | EIF | 0.334 | |||
| 3 | GAPDH | 0.69 | TUB | 0.38 | GAPDH | 0.391 | GAPDH | 0.454 | |
| 4 | TUB | 0.72 | GAPDH | 0.41 | TUB | 0.424 | RPL18 | 0.489 | |
| 5 | EF-1α | 0.91 | EF-1α | 0.46 | β-act | 0.737 | EF-1α | 0.593 | |
| 6 | β-act | 0.92 | 18S | 0.6 | EF-1α | 0.748 | β-act | 0.703 | |
| 7 | 18S | 1.06 | β-act | 0.68 | 18S | 0.929 | 18S | 0.805 | |
References
- Bustin, S.A. Quantification of mRNA using real-time reverse transcription PCR (RTPCR): Trends and problems. J. Mol. Endocrinol. 2002, 29, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Nolan, T. Pitfalls of quantitative real-time reverse-transcription polymerase chain reaction. J. Biomol. Tech. 2004, 15, 155–166. [Google Scholar] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Thellin, O.; Zorzi, W.; Lakaye, B.; De Borman, B.; Coumans, B.; Hennen, G.; Grisar, T.; Igout, A.; Heinen, E. Housekeeping genes as internal standards: Use and limits. J. Biotechnol. 1999, 75, 291–295. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; An, S.; Li, Z.; Wu, F.; Yang, Q.; Liu, Y.; Cao, J.; Zhang, H.; Zhang, Q.; Liu, X. Identification and validation of reference genes for normalization of gene expression analysis using qRT-PCR in Helicoverpa armigera (Lepidoptera: Noctuidae). Gene 2015, 555, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Qian, Z.; Lu, W.; Ding, H.; Yu, H.; Wang, H.; Li, J. Identification and characterization of reference genes for normalizing expression data from red swamp crawfish Procambarus clarkii. Int. J. Mol. Sci. 2015, 16, 21591–21605. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Wang, K.; Chen, D.; Wang, J.; He, Y.; Long, B.; Yang, L.; Yang, Q.; Geng, Y.; Huang, X.; et al. Evaluation and selection of appropriate reference genes for real-time quantitative PCR analysis of gene expression in Nile tilapia (Oreochromis niloticus) during vaccination and infection. Int. J. Mol. Sci. 2015, 16, 9998–10015. [Google Scholar] [CrossRef] [PubMed]
- Subramoniam, T. Sexual Biology and Reproduction in Crustaceans, 1st ed.; Academic Press: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Dhar, A.K.; Bowers, R.M.; Licon, K.S.; Veazey, G.; Read, B. Validation of reference genes for quantitative measurement of immune gene expression in shrimp. Mol. Immunol. 2009, 46, 1688–1695. [Google Scholar] [CrossRef] [PubMed]
- Leelatanawit, R.; Klanchui, A.; Uawisetwathana, U.; Karoonuthaisiri, N. Validation of reference genes for real-time PCR of reproductive system in the black tiger shrimp. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cottin, D.; Shillito, B.; Chertemps, T.; Thatje, S.; Léger, N.; Ravaux, J. Comparison of heat-shock responses between the hydrothermal vent shrimp Rimicaris exoculata and the related coastal shrimp Palaemonetes varians. J. Exp. Marine Biol. Ecol. 2010, 393, 9–16. [Google Scholar] [CrossRef]
- Priyadarshi, H.; Das, R.; Kumar, A.P.; Babu-Gireesh, P.; Javed, H.; Krishna, G.; Marappan, M.; Chaudhari, A. Characterization and evaluation of selected house-keeping genes for quantitative RT-PCR in Macrobrachium rosenbergii morphotypes. Fish. Technol. 2015, 52, 177–183. [Google Scholar]
- Jaramillo, M.L.; Ammar, D.; Quispe, R.L.; Guzman, F.; Margis, R.; Nazari, E.M.; Müller, Y.M. Identification and evaluation of reference genes for expression studies by RT-qPCR during embryonic development of the emerging model organism, Macrobrachium olfersii. Gene 2017, 598, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Fu, H.; Sun, S.; Jiang, S.; Xiong, Y.; Gong, Y.; Qiao, H.; Zhang, W.; Wu, Y. Integrated analysis of microRNA and mRNA expression profiles during the sex-differentiation sensitive period in oriental river prawn, Macrobrachium nipponense. Sci. Rep. 2017, 7, 12011. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Xuan, F.; Fu, H.; Ge, X.; Zhu, J.; Qiao, H.; Jin, S.; Zhang, Y. Comparative proteomic study of the response to hypoxia in the muscle of Oriental River prawn (Macrobrachium nipponense). J. Proteom. 2016, 138, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Y.; Jiang, S.F.; Xiong, Y.W.; Fu, H.T.; Qiao, H.; Sun, S.M.; Gong, Y.S.; Jin, S.B. Molecular cloning and expression analysis of female sterile homeotic gene (fsh) in the oriental river prawn Macrobrachium nipponense. Genet. Mol. Res. 2015, 14, 4318–4330. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Fu, H.; Xiong, Y.; Jiang, S.; Zhang, W.; Sun, S.; Jin, S.; Gong, Y.; Wang, Y.; Shan, D.; et al. Molecular insights into reproduction regulation of female Oriental River prawns Macrobrachium nipponense through comparative transcriptomic analysis. Sci. Rep. 2017, 7, 12161. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Fu, H.; Sun, S.; Qiao, H.; Zhang, W.; Jiang, S.; Xiong, Y.; Jin, S.; Gong, Y.; Wu, Y. Dietary cholesterol-induced transcriptome differences in the intestine, hepatopancreas, and muscle of Oriental River prawn Macrobrachium nipponense. Comp. Biochem. Physicol. Part D Genom. Proteom. 2017, 23, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Fu, H.; Jin, S.; Wu, Y.; Jiang, S.; Gong, Y.; Xiong, Y. Constructing and random sequencing analysis of normalized cDNA library of testis tissue from oriental river prawn (Macrobrachium nipponense). Comp. Biochem. Physicol. Part D Genom. Proteom. 2012, 7, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Fu, H.; Zhou, Q.; Sun, S.; Jiang, S.; Xiong, Y.; Gong, Y.; Qiao, H.; Zhang, W. Transcriptome analysis of androgenic gland for discovery of novel genes from the oriental river prawn, Macrobrachium nipponense, using Illumina Hiseq 2000. PLoS ONE 2013, 8, e76840. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Altman, N. Points of significance: Visualizing samples with box plots. Nat. Methods 2014, 11, 119–120. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Qiao, H.; Li, F.; Fu, H.; Sun, S.; Zhang, W.; Jin, S.; Gong, Y.; Jiang, S.; Xiong, Y. Molecular characterization and developmental expression of vitellogenin in the oriental river prawn Macrobrachium nipponense and the effects of RNA interference and eyestalk ablation on ovarian maturation. Gene 2015, 562, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Qiao, H.; Fu, H.; Sun, S.; Zhang, W.; Jin, S.; Jiang, S.; Gong, Y.; Xiong, Y.; Wu, Y.; et al. Identification and characterization of opsin gene and its role in ovarian maturation in the oriental river prawn Macrobrachium nipponense. Comp. Biochem. Phys. Part B Biochem. Mol. Biol. 2018, 218, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Subramoniam, T. Mechanisms and control of vitellogenesis in crustaceans. Fish. Sci. 2011, 77, 1–21. [Google Scholar] [CrossRef]
- Eissa, N.; Hussein, H.; Wang, H.; Rabbi, M.F.; Bernstein, N.; Ghia, J.-E. Stability of reference genes for messenger RNA quantification by real-time PCR in mouse dextran sodium sulfate experimental colitis. PLoS ONE 2016, 11, e0156289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Han, X.; Chen, S.; Zheng, L.; He, X.; Liu, M.; Qiao, G.; Wang, Y.; Zhuo, R. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Salix matsudana under different abiotic stresses. Sci. Rep. 2017, 7, 40290. [Google Scholar] [CrossRef] [PubMed]
- Teste, M.A.; Duquenne, M.; François, J.M.; Parrou, J.L. Validation of reference genes for quantitative expression analysis by real-time RT-PCR in Saccharomyces cerevisiae. BMC Mol. Biol. 2009, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Maksup, S.; Supaibulwatana, K.; Selvaraj, G. High-quality reference genes for quantifying the transcriptional responses of Oryza sativa L. (ssp. indica and japonica) to abiotic stress conditions. Chin. Sci. Bull. 2013, 58, 1919–1930. [Google Scholar]
- Xu, H.; Li, C.; Zeng, Q.; Agrawal, I.; Zhu, X.; Gong, Z. Genome-wide identification of suitable zebrafish Danio rerio reference genes for normalization of gene expression data by RT-qPCR. J. Fish Biol. 2016, 88, 2095–2110. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Zhang, Z.; He, X.; Liang, G. Validation of reference genes in Solenopsis invicta in different developmental stages, castes and tissues. PLoS ONE 2013, 8, e57718. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.M.; Chahad-Ehlers, S.; Lima, A.L.; Taniguti, C.H.; Sobrinho, I., Jr.; Torres, F.R.; de Brito, R.A. Reference genes for accessing differential expression among developmental stages and analysis of differential expression of OBP genes in Anastrepha obliqua. Sci. Rep. 2016, 6, 17480. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Jiang, S.; Xiong, Y.; Qiao, H.; Sun, S.; Zhang, W.; Li, F.; Gong, Y.; Fu, H. Molecular cloning of two tropomyosin family genes and expression analysis during development in oriental river prawn, Macrobrachium nipponense. Gene 2014, 546, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Kubista, M.; Andrade, J.M.; Bengtsson, M.; Forootan, A.; Jonák, J.; Lind, K.; Sindelka, R.; Sjöback, R.; Sjögreen, B.; Strömbom, L.; et al. The real-time polymerase chain reaction. Mol. Asp. Med. 2006, 27, 95–125. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Xiong, Y.; Zhang, W.; Fu, H.; Jiang, S.; Sun, S.; Bai, H.; Jin, S.; Gong, Y. Characterization, expression, and function analysis of gonad-inhibiting hormone in Oriental River prawn, Macrobrachium nipponense and its induced expression by temperature. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 185, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.G.; Liu, H.; Xu, J.N.; Cai, S.L. Study on site of vitellogenin synthesis in the freshwater prawn Macrobrachium nipponese. Biotechnol. Bull. 2006, 438–444. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-SWJT2006S1096.htm (accessed on 1 August 2018). (In Chinese).
- Sun, S.M.; Xuan, F.J.; Ge, X.P.; Fu, H.T.; Zhu, J.; Zhang, S.Y. Identification of differentially expressed genes in hepatopancreas of oriental river prawn, Macrobrachium nipponense exposed to environmental hypoxia. Gene 2014, 534, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Fu, H.; Sun, S.; Qiao, H.; Zhang, W.; Jin, S.; Jiang, S.; Xiong, Y.; Gong, Y. Experimental inoculation of oriental river prawn Macrobrachium nipponense with white spot syndrome virus (WSSV). Dis. Aquat. Organ. 2017, 126, 125–134. [Google Scholar] [CrossRef] [PubMed]







| Gene | Primer Sequence (5′–3′) Forward/Reverse | Length (bp) | PCR Efficiency (%) | Correlation Coefficient (R2) |
|---|---|---|---|---|
| EIF | CATGGATGTACCTGTGGTGAAAC CTGTCAGCAGAAGGTCCTCATTA | 179 | 94.4 | 0.994 |
| 18S | GTTGGATGTTGCTGTTGAGAGAG CTGGGCATCATTCTCTGGGTAAA | 250 | 96.8 | 0.997 |
| EF-1α | CAAGGATCTGAAACGTGGCTTC GTACGTCTGTCGATCTTGGTCAG | 198 | 93.7 | 0.999 |
| GAPDH | GTCGGTAAGGTCATTCCAGAGC CGAAAGTTTTGCTGAGCTGGAT | 274 | 95.5 | 0.991 |
| TUB | AGGAATGGAAAATCAGGAAGCCC GTTTGTCGATCTGGAACCCTCT | 224 | 97.0 | 0.996 |
| β-act | CACGAGACCACCTACAATTCCA ATAGAGAAGCCAAGATAGAACCGC | 226 | 99.4 | 0.991 |
| RPL18 | CTTTTTGTACCCACAGCTTGACC CACTTTTGATGTATTGGCCCGTC | 202 | 102.7 | 0.997 |
| Experimental Conditions | Statistics | Reference Gene | ||||||
|---|---|---|---|---|---|---|---|---|
| EIF | 18S | EF-1α | GAPDH | TUB | β-act | RPL18 | ||
| Hypoxia stress | Group | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| Mean | 21.86 | 28.19 | 24.20 | 21.19 | 23.80 | 24.35 | 21.33 | |
| Min Cq | 20.64 | 26.08 | 22.50 | 20.02 | 23.13 | 23.01 | 19.03 | |
| Max Cq | 22.33 | 28.98 | 26.42 | 22.66 | 24.56 | 25.66 | 22.43 | |
| WSSV infection | Group | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
| Mean | 24.82 | 32.02 | 34.89 | 26.78 | 30.44 | 26.47 | 24.40 | |
| Min Cq | 24.12 | 30.97 | 34.22 | 26.26 | 29.98 | 25.72 | 23.76 | |
| Max Cq | 25.30 | 33.28 | 36.04 | 27.44 | 31.34 | 27.64 | 24.96 | |
| Different adult tissues | Group | 7 | 7 | 7 | 7 | 7 | 7 | 7 |
| Mean | 21.65 | 27.81 | 25.05 | 22.55 | 25.86 | 20.82 | 21.83 | |
| Min Cq | 20.47 | 26.09 | 18.57 | 17.90 | 22.61 | 18.64 | 20.56 | |
| Max Cq | 23.59 | 30.26 | 32.62 | 26.74 | 29.17 | 23.36 | 23.88 | |
| Different ovarian stages | Group | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| Mean | 21.29 | 25.42 | 33.34 | 23.61 | 23.81 | 19.28 | 22.18 | |
| Min Cq | 20.24 | 23.77 | 31.56 | 20.71 | 21.06 | 17.75 | 20.59 | |
| Max Cq | 22.66 | 27.62 | 35.32 | 25.40 | 25.11 | 21.19 | 24.40 | |
| Different embryo stages | Group | 7 | 7 | 7 | 7 | 7 | 7 | 7 |
| Mean | 21.23 | 27.51 | 30.90 | 25.13 | 25.25 | 21.71 | 22.63 | |
| Min Cq | 18.58 | 25.37 | 21.12 | 21.25 | 22.53 | 18.26 | 18.95 | |
| Max Cq | 23.77 | 32.10 | 37.48 | 30.47 | 30.98 | 25.62 | 27.97 | |
| Experimental Conditions | Statistics | Reference Gene | ||||||
|---|---|---|---|---|---|---|---|---|
| EIF | 18S | EF-1α | GAPDH | TUB | β-act | RPL18 | ||
| RNAi (Androgenic glands) | Group | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Mean | 21.29 | 27.62 | 24.51 | 22.14 | 23.46 | 19.11 | 20.58 | |
| Min Cq | 21.07 | 27.33 | 22.63 | 21.34 | 23.40 | 18.99 | 20.55 | |
| Max Cq | 21.51 | 27.91 | 26.38 | 23.14 | 23.51 | 19.23 | 20.62 | |
| Experimental Conditions | Method | Rank | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| RNAi (Androgenic glands) | Comparative ∆Ct | β-act | RPL18 | EIF | TUB | 18S | GAPDH | EF-1α |
| BestKeeper | RPL18 | TUB | β-act | EIF | 18S | GAPDH | EF-1α | |
| NormFinder | β-act | EIF | RPL18 | TUB | GAPDH | 18S | EF-1α | |
| GeNorm | β-act/RPL18 | TUB | EIF | 18S | GAPDH | EF-1α | ||
| Recommended comprehensive ranking | β-act | RPL18 | TUB | EIF | 18S | GAPDH | EF-1α | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Fu, H.; Qiao, H.; Sun, S.; Zhang, W.; Jin, S.; Jiang, S.; Gong, Y.; Xiong, Y.; Wu, Y. Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR in Macrobrachium Nipponense. Int. J. Mol. Sci. 2018, 19, 2258. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082258
Hu Y, Fu H, Qiao H, Sun S, Zhang W, Jin S, Jiang S, Gong Y, Xiong Y, Wu Y. Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR in Macrobrachium Nipponense. International Journal of Molecular Sciences. 2018; 19(8):2258. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082258
Chicago/Turabian StyleHu, Yuning, Hongtuo Fu, Hui Qiao, Shengming Sun, Wenyi Zhang, Shubo Jin, Sufei Jiang, Yongsheng Gong, Yiwei Xiong, and Yan Wu. 2018. "Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR in Macrobrachium Nipponense" International Journal of Molecular Sciences 19, no. 8: 2258. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082258