Genome-Wide Association Study Reveals Novel Genomic Regions for Grain Yield and Yield-Related Traits in Drought-Stressed Synthetic Hexaploid Wheat


Abstract
:1. Introduction
2. Results and Discussion
2.1. Weather Conditions
2.2. Phenotypic Variation for Yield and Yield-Related Traits
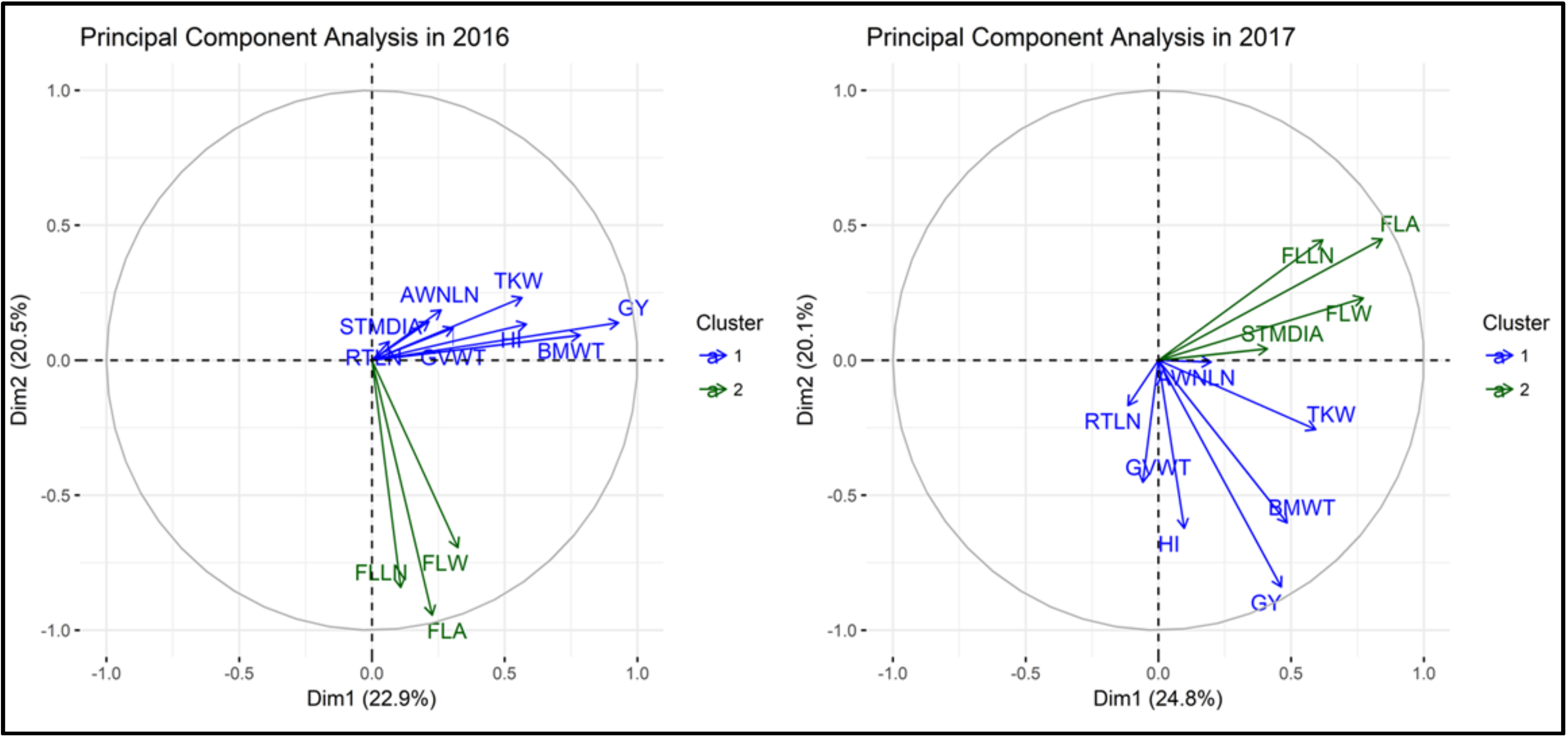
2.3. Principal Component Analysis and Phenotypic Correlation
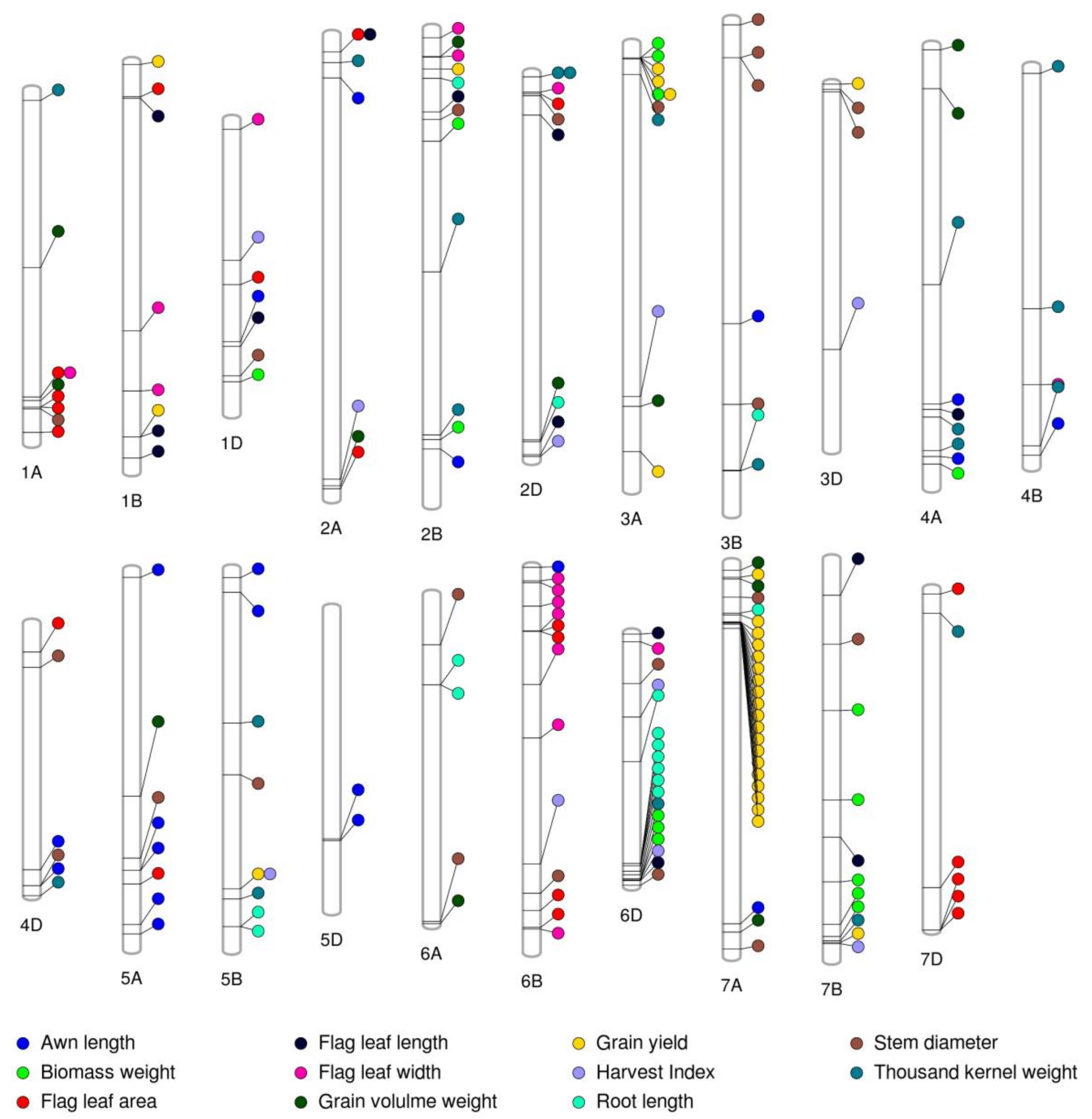
2.4. Population Structure and Genome-Wide Association Study
2.4.1. Grain Yield
2.4.2. Harvest Index
2.4.3. Biomass Weight
2.4.4. Thousand Kernel Weight
2.4.5. Grain Volume Weight
2.4.6. Awn Length
2.4.7. Flag Leaf Length
2.4.8. Flag Leaf Width
2.4.9. Flag Leaf Area
2.4.10. Stem Diameter
2.4.11. Root Length
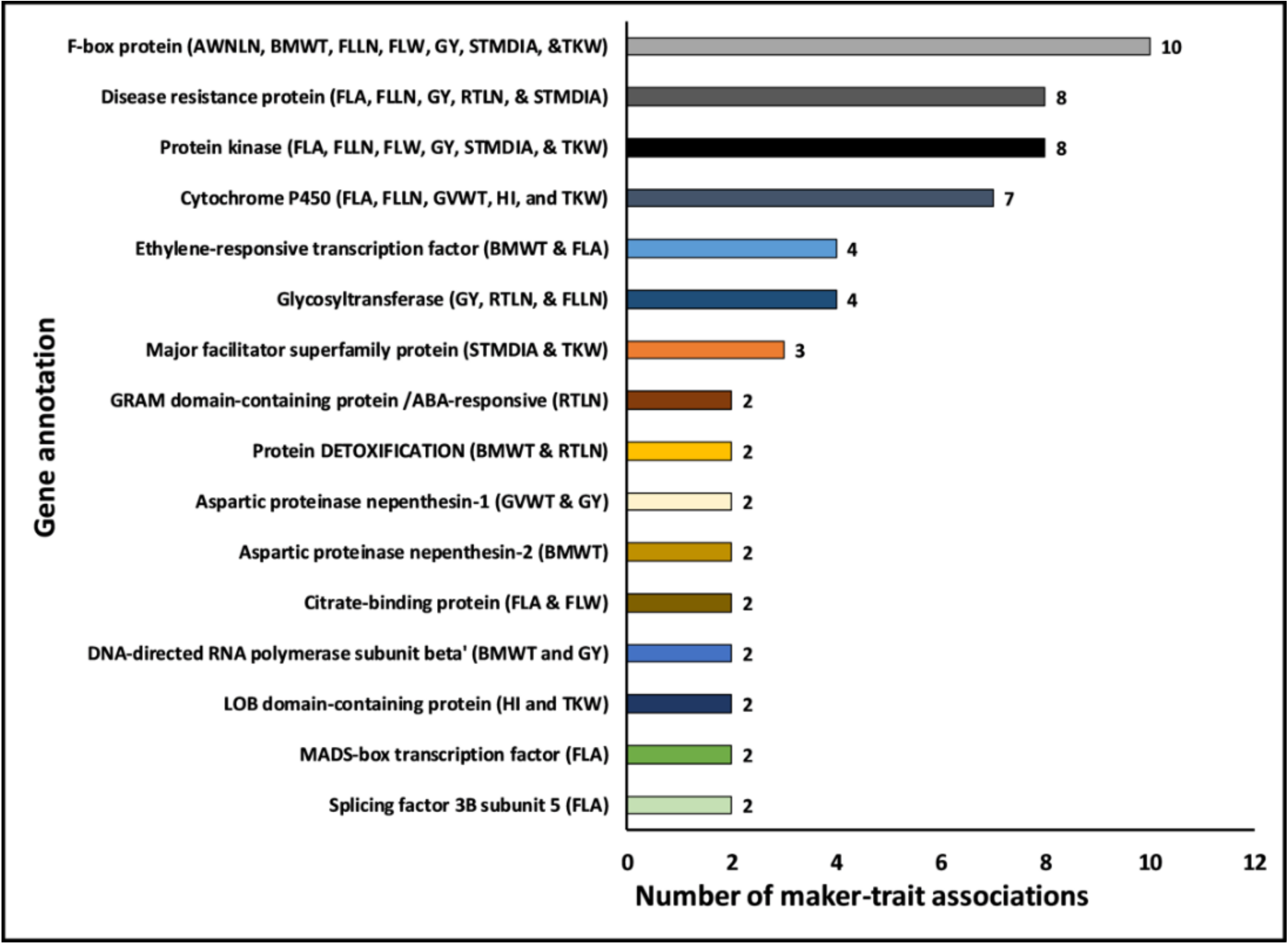
2.5. Potential Candidate Gene Annotations Affecting Yield and Yield-related Traits under Drought Stress
3. Materials and Methods
3.1. Site Description
3.2. Plant Materials and Experimental Design
3.3. Trait Measurements
3.4. Phenotypic Data Analysis
3.5. Genotyping and SNP Discovery
3.6. Population Structure and Genome-Wide Association Study Analysis
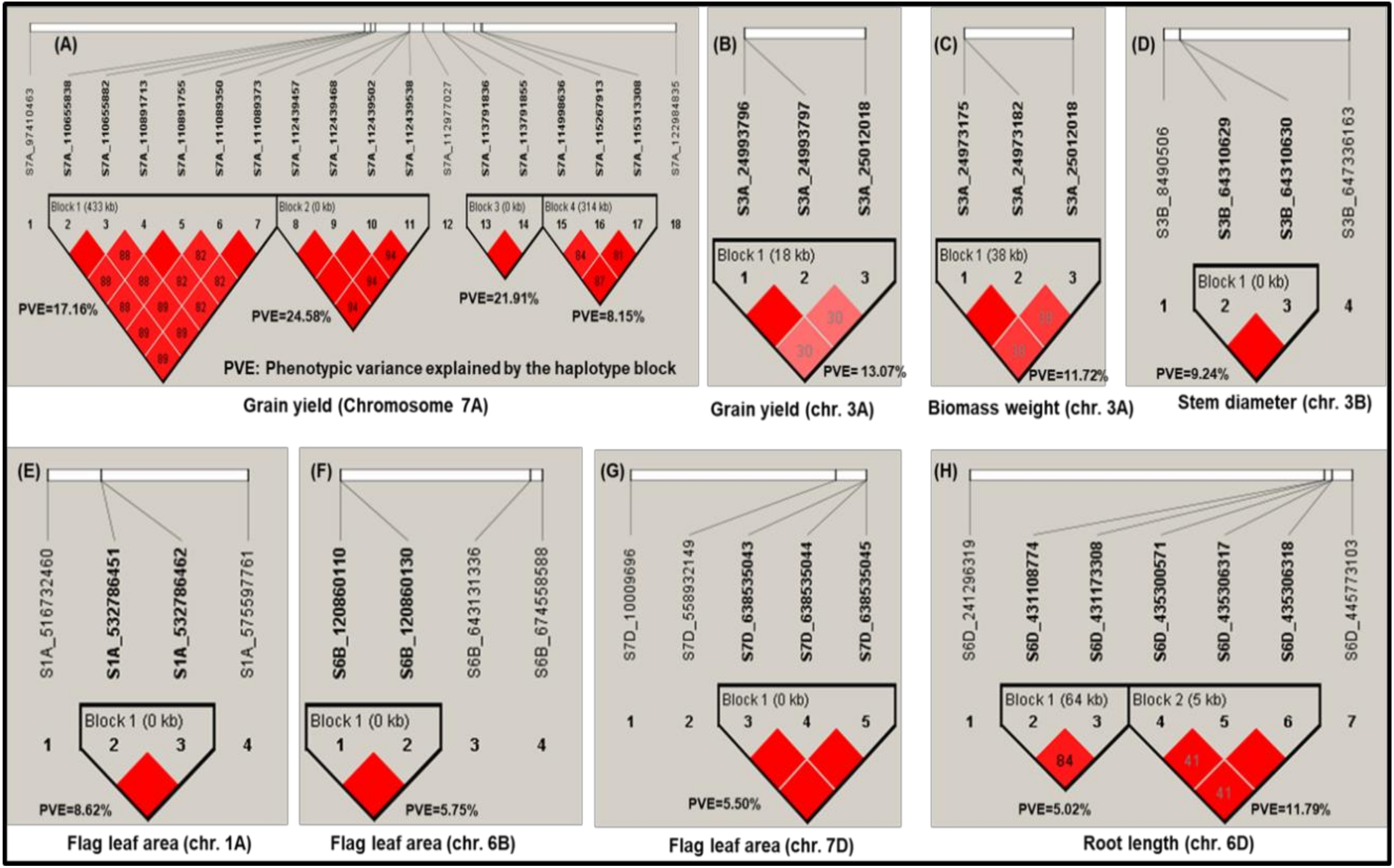
3.7. Haplotype Block Analysis
3.8. Putative Candidate Gene Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| AWNLN | awn length |
| BLUP | best linear unbiased predictor |
| BMWT | biomass weight |
| FarmCPU | fixed and random model circulating probability unification |
| FLA | flag leaf area |
| FLLN | flag leaf length |
| FLW | flag leaf width |
| GBS | genotyping-by-sequencing |
| GVWT | grain volume weight |
| GWAS | genome wide association study |
| GY | grain yield |
| HI | harvest index |
| IWGC | international wheat genome sequencing consortium |
| MAF | minor allele frequency |
| QTN | quantitative trait nucleotide |
| MLM | mixed linear model |
| MLMM | multilocus mixed model |
| MTA | marker trait association |
| QTL | quantitative trait loci |
| RTLN | root length |
| SHW | synthetic hexaploid wheat |
| SNP | single nucleotide polymorphism |
| STMDIA | stem diameter |
| TKW | thousand kernel weight |
References
- Kang, Y.; Khan, S.; Ma, X. Climate change impacts on crop yield, crop water productivity and food security—A review. Prog. Nat. Sci. 2009, 19, 1665–1674. [Google Scholar] [CrossRef]
- Becker, S.R.; Byrne, P.F.; Reid, S.D.; Bauerle, W.L.; McKay, J.K.; Haley, S.D. Root traits contributing to drought tolerance of synthetic hexaploid wheat in a greenhouse study. Euphytica 2016, 207, 213–224. [Google Scholar] [CrossRef]
- Gupta, P.; Balyan, H.; Gahlaut, V.; Gupta, P.K.; Balyan, H.S.; Gahlaut, V. QTL analysis for drought tolerance in wheat: Present status and future possibilities. Agronomy 2017, 7, 5. [Google Scholar] [CrossRef]
- Smith, A.B.; Matthews, J.L. Quantifying uncertainty and variable sensitivity within the US billion-dollar weather and climate disaster cost estimates. Nat. Hazards 2015, 77, 1829–1851. [Google Scholar] [CrossRef]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.L.; McIntyre, C.L.; Olivares-Villegas, J.-J.; Chapman, S.C. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Theor. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gummadov, N.; Keser, M.; Akin, B.; Cakmak, M.; Mert, Z.; Taner, S.; Ozturk, I.; Topal, A.; Yazar, S.; Morgounov, A. Genetic gains in wheat in Turkey: Winter wheat for irrigated conditions. Crop J. 2015, 3, 507–516. [Google Scholar] [CrossRef]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, T. Deepening the wheat gene pool. J. Crop Prod. 1997, 1, 145–168. [Google Scholar] [CrossRef]
- Dreisigacker, S.; Kishii, M.; Lage, J.; Warburton, M. Use of synthetic hexaploid wheat to increase diversity for CIMMYT bread wheat improvement. Aust. J. Agric Res. 2008, 59, 413–420. [Google Scholar] [CrossRef]
- Ogbonnaya, F. C.; Abdalla, O.; Mujeeb-Kazi, A.; Kazi, A.G.; Xu, S.S.; Gosman, N.; Lagudah, E.S.; Bonnett, D.; Sorrells, M.E.; Tsujimoto, H. Synthetic hexaploids: Harnessing species of the primary gene pool for wheat improvement. Plant Breed. Rev. 2013, 37, 35–122. [Google Scholar] [CrossRef]
- Bhatta, M.; Morgounov, A.; Belamkar, V.; Poland, J.; Baenziger, P.S. Unlocking the novel genetic diversity and population structure of synthetic hexaploid wheat. BMC Genom. 2018, 19, 591. [Google Scholar] [CrossRef] [PubMed]
- Zegeye, H.; Rasheed, A.; Makdis, F.; Badebo, A.; Ogbonnaya, F.C. Genome-wide association mapping for seedling and adult plant resistance to stripe rust in synthetic hexaploid wheat. PLoS ONE 2014, 9, e105593. [Google Scholar] [CrossRef] [PubMed]
- Das, M.K.; Bai, G.; Mujeeb-Kazi, A.; Rajaram, S. Genetic diversity among synthetic hexaploid wheat accessions (Triticum aestivum) with resistance to several fungal diseases. Genet. Resour. Crop Evol. 2016, 63, 1285–1296. [Google Scholar] [CrossRef]
- Morgounov, A.; Abugalieva, A.; Akan, K.; Akın, B.; Baenziger, S.; Bhatta, M.; Dababat, A.A.; Demir, L.; Dutbayev, Y.; El Bouhssini, M.; et al. High-yielding winter synthetic hexaploid wheats resistant to multiple diseases and pests. Plant Genet. Resour. 2018, 16, 273–278. [Google Scholar] [CrossRef]
- Ogbonnaya, F.C.; Imtiaz, M.; Bariana, H.S.; McLean, M.; Shankar, M.M.; Hollaway, G.J.; Trethowan, R.M.; Lagudah, E.S.; Van Ginkel, M. Mining synthetic hexaploids for multiple disease resistance to improve bread wheat. Aust. J. Agric Res. 2008, 59, 421–431. [Google Scholar] [CrossRef]
- Jighly, A.; Alagu, M.; Makdis, F.; Singh, M.; Singh, S.; Emebiri, L.C.; Ogbonnaya, F.C. Genomic regions conferring resistance to multiple fungal pathogens in synthetic hexaploid wheat. Mol. Breed. 2016, 36, 1–19. [Google Scholar] [CrossRef]
- Sehgal, D.; Autrique, E.; Singh, R.; Ellis, M.; Singh, S.; Dreisigacker, S. Identification of genomic regions for grain yield and yield stability and their epistatic interactions. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, M.; Eskridge, K.M.; Rose, D.J.; Santra, D.K.; Baenziger, P.S.; Regassa, T. Seeding Rate, genotype, and topdressed nitrogen effects on yield and agronomic characteristics of winter wheat. Crop Sci. 2017, 57, 951–963. [Google Scholar] [CrossRef]
- Ogbonnaya, F.C.; Rasheed, A.; Okechukwu, E.C.; Jighly, A.; Makdis, F.; Wuletaw, T.; Hagras, A.; Uguru, M.I.; Agbo, C.U. Genome-wide association study for agronomic and physiological traits in spring wheat evaluated in a range of heat prone environments. Theor. Appl. Genet. 2017, 130, 1819–1835. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, S.; Lopes, M.; Dreisigacker, S.; Reynolds, M. Genetic analysis of multi-environmental spring wheat trials identifies genomic regions for locus-specific trade-offs for grain weight and grain number. Theor. Appl. Genet. 2018, 131, 985–998. [Google Scholar] [CrossRef] [PubMed]
- Murtas, G. A Nuclear Protease Required for Flowering-Time Regulation in Arabidopsis Reduces the Abundance of SMALL UBIQUITIN-RELATED MODIFIER Conjugates. Plant Cell 2003, 15, 2308–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, M.K.; Basu, D.; Datta, A.; Chakraborty, N.; Chakraborty, S. Dehydration-responsive nuclear proteome of rice (Oryza sativa L.) illustrates protein network, novel regulators of cellular adaptation, and evolutionary perspective. Mol. Cell. Proteomics 2009, 8, 1579–1598. [Google Scholar] [CrossRef] [PubMed]
- Yokosho, K.; Yamaji, N.; Fujii-Kashino, M.; Ma, J.F. Functional Analysis of a MATE gene OsFRDL2 revealed its involvement in Al-induced secretion of citrate, but a lower contribution to Al tolerance in rice. Plant Cell Physiol. 2016, 57, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, F.; Yan, X.; Zhang, X.; Dong, Z.; Cui, D.; Chen, F. Genome-wide association study for 13 agronomic traits reveals distribution of superior alleles in bread wheat from the Yellow and Huai Valley of China. Plant Biotechnol. J. 2017, 15, 953–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanke, C.D.; Ling, J.; Plieske, J.; Kollers, S.; Ebmeyer, E.; Korzun, V.; Argillier, O.; Stiewe, G.; Hinze, M.; Neumann, K.; et al. Whole genome association mapping of plant height in winter wheat (Triticum aestivum L.). PLoS ONE 2014, 9, e113287. [Google Scholar] [CrossRef] [PubMed]
- Golabadi, M.; Arzani, A.; Mirmohammadi Maibody, S.A.M.; Sayed Tabatabaei, B.E.; Mohammadi, S.A. Identification of microsatellite markers linked with yield components under drought stress at terminal growth stages in durum wheat. Euphytica 2011, 177, 207–221. [Google Scholar] [CrossRef]
- Kumar, N.; Kulwal, P.L.; Balyan, H.S.; Gupta, P.K. QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat. Mol. Breed. 2007, 19, 163–177. [Google Scholar] [CrossRef]
- Sukumaran, S.; Dreisigacker, S.; Lopes, M.; Chavez, P.; Reynolds, M.P. Genome-wide association study for grain yield and related traits in an elite spring wheat population grown in temperate irrigated environments. Theor. Appl. Genet. 2015, 128, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Lozada, D.N.; Mason, R.E.; Babar, M.A.; Carver, B.F.; Guedira, G.B.; Merrill, K.; Arguello, M.N.; Acuna, A.; Vieira, L.; Holder, A.; et al. Association mapping reveals loci associated with multiple traits that affect grain yield and adaptation in soft winter wheat. Euphytica 2017, 213. [Google Scholar] [CrossRef]
- Neumann, K.; Kobiljski, B.; Denčić, S.; Varshney, R.K.; Börner, A. Genome-wide association mapping: A case study in bread wheat (Triticum aestivum L.). Mol. Breed. 2011, 27, 37–58. [Google Scholar] [CrossRef]
- Edae, E.A.; Byrne, P.F.; Haley, S.D.; Lopes, M.S.; Reynolds, M.P. Genome-wide association mapping of yield and yield components of spring wheat under contrasting moisture regimes. Theor. Appl. Genet. 2014, 127, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, M.; Regassa, T.; Rose, D.J.; Baenziger, P.S.; Eskridge, K.M.; Santra, D.K.; Poudel, R. Genotype, environment, seeding rate, and top-dressed nitrogen effects on end-use quality of modern Nebraska winter wheat. J. Sci. Food Agric. 2017, 97, 5311–5318. [Google Scholar] [CrossRef] [PubMed]
- Bordes, J.; Goudemand, E.; Duchalais, L.; Chevarin, L.; Oury, F.X.; Heumez, E.; Lapierre, A.; Perretant, M.R.; Rolland, B.; Beghin, D.; et al. Genome-wide association mapping of three important traits using bread wheat elite breeding populations. Mol. Breed. 2014, 33, 755–768. [Google Scholar] [CrossRef]
- Hoffstetter, A.; Cabrera, A.; Sneller, C. Identifying quantitative trait loci for economic traits in an elite soft red winter wheat population. Crop Sci. 2016, 56, 547. [Google Scholar] [CrossRef]
- Wang, S.-X.; Zhu, Y.-L.; Zhang, D.-X.; Shao, H.; Liu, P.; Hu, J.-B.; Zhang, H.; Zhang, H.-P.; Chang, C.; Lu, J.; et al. Genome-wide association study for grain yield and related traits in elite wheat varieties and advanced lines using SNP markers. PLOS ONE 2017, 12, e0188662. [Google Scholar] [CrossRef] [PubMed]
- Catala, R.; Ouyang, J.; Abreu, I.A.; Hu, Y.; Seo, H.; Zhang, X.; Chua, N.-H. The Arabidopsis E3 SUMO Ligase SIZ1 Regulates Plant Growth and Drought Responses. Plant Cell 2007, 19, 2952–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.J.; Kim, W.-Y.; Park, H.C.; Lee, S.Y.; Bohnert, H.J.; Yun, D.-J. SUMO and SUMOylation in plants. Mol. Cells 2011, 32, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.L.; Baenziger, P.S.; Ajlouni, Z.A.; Campbell, B.T.; Gill, K.S.; Eskridge, K.M.; Mujeeb-Kazi, A.; Dweikat, I. Mapping QTL for agronomic traits on wheat chromosome 3a and a comparison of recombinant inbred chromosome line populations. Crop Sci. 2011, 51, 553–566. [Google Scholar] [CrossRef]
- Lechner, E.; Achard, P.; Vansiri, A.; Potuschak, T.; Genschik, P. F-box proteins everywhere. Curr. Opin. Plant Biol. 2006, 9, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-A.; Yeom, S.-I. Plant NB-LRR proteins: Tightly regulated sensors in a complex manner. Brief. Funct. Genomics 2015, 14, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Ain, Q.; Rasheed, A.; Anwar, A.; Mahmood, T.; Imtiaz, M.; Mahmood, T.; Xia, X.; He, Z.; Quraishi, U.M. Genome-wide association for grain yield under rainfed conditions in historical wheat cultivars from Pakistan. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- He, G.-H.; Xu, J.-Y.; Wang, Y.-X.; Liu, J.-M.; Li, P.-S.; Chen, M.; Ma, Y.-Z.; Xu, Z.-S. Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis. BMC Plant Biol. 2016, 16, 116. [Google Scholar] [CrossRef] [PubMed]
- Ning, P.; Liu, C.; Kang, J.; Lv, J. Genome-wide analysis of WRKY transcription factors in wheat (Triticum aestivum L.) and differential expression under water deficit condition. PeerJ 2017, 5, e3232. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, M.; Undan, J. R.; Takagi, H.; Abe, A.; Yoshida, K.; Undan, J. Q.; Natsume, S.; Uemura, A.; Saitoh, H.; Matsumura, H.; Urasaki, N.; Yokota, T.; Terauchi, R. A cytochrome P450, OsDSS1, is involved in growth and drought stress responses in rice (Oryza sativa L.). Plant Mol. Biol. 2015, 88, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fujita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray: Expression profiling under abiotic stresses. Plant J. 2002, 31, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Kushiro, T.; Okamoto, M.; Nakabayashi, K.; Yamagishi, K.; Kitamura, S.; Asami, T.; Hirai, N.; Koshiba, T.; Kamiya, Y.; Nambara, E. The Arabidopsis cytochrome P450 CYP707A encodes ABA 8′-hydroxylases: key enzymes in ABA catabolism. EMBO J. 2004, 23, 1647–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Su, P.; Wei, Z.; Nevo, E.; Kong, L. Genome-wide identification, classification, evolutionary analysis and gene expression patterns of the protein kinase gene family in wheat and Aegilops tauschii. Plant Mol. Biol. 2017, 95, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Maruyama, K.; Sakuma, Y.; Meshi, T.; Iwabuchi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis Cys2/His2-Type zinc-finger proteins function as transcription repressors under drought, cold, and high-salinity stress conditions. Plant Physiol. 2004, 136, 2734–2746. [Google Scholar] [CrossRef] [PubMed]
- Ciftci-Yilmaz, S.; Morsy, M.R.; Song, L.; Coutu, A.; Krizek, B.A.; Lewis, M.W.; Warren, D.; Cushman, J.; Connolly, E.L.; Mittler, R. The EAR-motif of the Cys2/His2-type zinc finger protein Zat7 plays a key role in the defense response of Arabidopsis to salinity stress. J. Biol. Chem. 2007, 282, 9260–9268. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Sequence and expression analysis of the C3HC4-type RING finger gene family in rice. Gene 2009, 444, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Cabral, A.L.; Jordan, M.C.; Larson, G.; Somers, D.J.; Humphreys, D.G.; McCartney, C.A. Relationship between QTL for grain shape, grain weight, test weight, milling yield, and plant height in the spring wheat cross RL4452/‘AC Domain’. PLoS ONE 2018, 13, e0190681. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, X.; Jiao, Y.; Qin, Y.; Liu, X.; He, K.; Chen, C.; Ma, L.; Wang, J.; Xiong, L.; et al. Global genome expression analysis of rice in response to drought and high-salinity stresses in shoot, flag leaf, and panicle. Plant Mol. Biol. 2007, 63, 591–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sourdille, P.; Cadalen, T.; Gay, G.; Gill, B.; Bernard, M. Molecular and physical mapping of genes affecting awning in wheat. Plant Breed. 2002, 121, 320–324. [Google Scholar] [CrossRef] [Green Version]
- Echeverry-Solarte, M.; Kumar, A.; Kianian, S.; Mantovani, E.E.; McClean, P.E.; Deckard, E.L.; Elias, E.; Simsek, S.; Alamri, M.S.; Hegstad, J.; et al. Genome-wide mapping of spike-related and agronomic traits in a common wheat population derived from a supernumerary spikelet parent and an elite parent. Plant Genome 2015, 8, 0. [Google Scholar] [CrossRef]
- Li, H.Y.; Wang, T.Y.; Shi, Y.S.; Fu, J.J.; Song, Y.C.; Wang, G.Y.; Li, Y. Isolation and characterization of induced genes under drought stress at the flowering stage in maize (Zea mays): Full Length Research Paper. DNA Seq 2007, 18, 445–460. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Assmann, S.M. The Arabidopsis putative g protein–coupled receptor GCR1 interacts with the G protein α subunit GPA1 and regulates abscisic acid signaling. Plant Cell 2004, 16, 1616–1632. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Chen, Y.; Fu, L.; Zhou, S.; Chen, J.; Zhao, X.; Zhang, D.; Ouyang, S.; Wang, Z.; Li, D.; et al. QTL mapping of flag leaf traits in common wheat using an integrated high-density SSR and SNP genetic linkage map. Euphytica 2016, 208, 337–351. [Google Scholar] [CrossRef]
- Yang, D.; Liu, Y.; Cheng, H.; Chang, L.; Chen, J.; Chai, S.; Li, M. Genetic dissection of flag leaf morphology in wheat (Triticum aestivum L.) under diverse water regimes. BMC Genet 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Cui, F.; Zhao, C.; Zhang, W.; Yang, L.; Zhao, X.; Han, J.; Su, Q.; Ji, J.; Zhao, Z.; et al. QTLs for flag leaf size and their influence on yield-related traits in wheat (Triticum aestivum L.). Mol. Breed. 2015, 35, 24. [Google Scholar] [CrossRef]
- Liu, K.; Xu, H.; Liu, G.; Guan, P.; Zhou, X.; Peng, H.; Yao, Y.; Ni, Z.; Sun, Q.; Du, J. QTL mapping of flag leaf-related traits in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2018, 131, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Hussain, W.; Baenziger, P.S.; Belamkar, V.; Guttieri, M.J.; Venegas, J.P.; Easterly, A.; Sallam, A.; Poland, J. Genotyping-by-sequencing derived high-density linkage map and its application to QTL mapping of flag leaf traits in bread wheat. Sci. Rep. 2017, 7, 16394. [Google Scholar] [CrossRef] [PubMed]
- Ros, B.; Thümmler, F.; Wenzel, G. Analysis of differentially expressed genes in a susceptible and moderately resistant potato cultivar upon Phytophthora infestans infection. Mol. Plant Pathol. 2004, 5, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cai, M.; Zhang, X.; Li, Y.; Zhang, J.; Zhao, H.; Kong, F.; Zheng, Y.; Qiu, F. Genome-Wide identification, evolution and expression analysis of mTERF gene family in maize. PLoS ONE 2014, 9, e94126. [Google Scholar] [CrossRef] [PubMed]
- Cotsaftis, O.; Plett, D.; Johnson, A.A.T.; Walia, H.; Wilson, C.; Ismail, A.M.; Close, T.J.; Tester, M.; Baumann, U. Root-specific transcript profiling of contrasting rice genotypes in response to salinity stress. Mol. Plant 2011, 4, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Hai, L.; Guo, H.; Xiao, S.; Jiang, G.; Zhang, X.; Yan, C.; Xin, Z.; Jia, J. Quantitative trait loci (QTL) of stem strength and related traits in a doubled-haploid population of wheat (Triticum aestivum L.). Euphytica 2005, 141, 1–9. [Google Scholar] [CrossRef]
- Shumayla; Sharma, S.; Kumar, R.; Mendu, V.; Singh, K.; Upadhyay, S.K. Genomic dissection and expression profiling revealed functional divergence in Triticum aestivum leucine rich repeat receptor like kinases (TaLRRKs). Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Landjeva, S.; Neumann, K.; Lohwasser, U.; Börner, A. Molecular mapping of genomic regions associated with wheat seedling growth under osmotic stress. Biol. Plant. 2008, 52, 259–266. [Google Scholar] [CrossRef]
- Ren, Y.; He, X.; Liu, D.; Li, J.; Zhao, X.; Li, B.; Tong, Y.; Zhang, A.; Li, Z. Major quantitative trait loci for seminal root morphology of wheat seedlings. Mol. Breed. 2012. [Google Scholar]
- Bai, C.; Liang, Y.; Hawkesford, M.J. Identification of QTLs associated with seedling root traits and their correlation with plant height in wheat. J. Exp. Bot. 2013, 64, 1745–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, J.A.; Wingen, L.U.; Griffiths, M.; Pound, M.P.; Gaju, O.; Foulkes, M.J.; Le Gouis, J.; Griffiths, S.; Bennett, M.J.; King, J.; et al. Phenotyping pipeline reveals major seedling root growth QTL in hexaploid wheat. J. Exp. Bot. 2015, 66, 2283–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canè, M.A.; Maccaferri, M.; Nazemi, G.; Salvi, S.; Francia, R.; Colalongo, C.; Tuberosa, R. Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance. Mol. Breed. 2014, 34, 1629–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccaferri, M.; El-Feki, W.; Nazemi, G.; Salvi, S.; Canè, M.A.; Colalongo, M.C.; Stefanelli, S.; Tuberosa, R. Prioritizing quantitative trait loci for root system architecture in tetraploid wheat. J. Exp. Bot. 2016, 67, 1161–1178. [Google Scholar] [CrossRef] [PubMed]
- Oono, Y.; Yazawa, T.; Kanamori, H.; Sasaki, H.; Mori, S.; Handa, H.; Matsumoto, T. Genome-wide transcriptome analysis of cadmium stress in rice. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, J.; Yamaji, N.; Wang, H.; Mitani, N.; Murata, Y.; Sato, K.; Katsuhara, M.; Takeda, K.; Ma, J.F. An aluminum-activated citrate transporter in barley. Plant Cell Physiol. 2007, 48, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.R.; Raman, H.; Gupta, S.; Horst, W.J.; Delhaize, E. A second mechanism for aluminum resistance in wheat relies on the constitutive efflux of citrate from roots. Plant Physiol. 2009, 149, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Baron, K.N.; Schroeder, D.F.; Stasolla, C. GEm-related 5 (GER5), an ABA and stress-responsive GRAM domain protein regulating seed development and inflorescence architecture. Plant Sci. 2014, 223, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, N.; Yao, C.; Meng, S.; Song, C. Functional analysis of the ABA-responsive protein family in ABA and stress signal transduction in Arabidopsis. Chin. Sci. Bull. 2013, 58, 3721–3730. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Singh, N.; Kaur, S.; Bains, N.S.; Uauy, C.; Poland, J.; Chhuneja, P. Genome-wide association study of grain architecture in wild wheat Aegilops tauschii. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, S.; Xiang, W.; Bean, S.R.; Pedersen, J.F.; Kresovich, S.; Tuinstra, M.R.; Tesso, T.T.; Hamblin, M.T.; Yu, J. Association mapping for grain quality in a diverse sorghum collection. Plant Genome J. 2012, 5, 126. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Giri, J.; Kapoor, S.; Tyagi, A.K.; Pandey, G.K. Protein phosphatase complement in rice: Genome-wide identification and transcriptional analysis under abiotic stress conditions and reproductive development. BMC Genom. 2010, 11, 435. [Google Scholar] [CrossRef] [PubMed]
- Evaluation of deficit irrigation for efficient sheep production from permanent sown pastures in a dry continental climate. Agric. Water Manag. 2013, 119, 135–143. [CrossRef]
- SAS 9.4 Product Documentation. Available online: https://support.sas.com/documentation/94/ (accessed on 16 August 2018).
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. 2017. Available online: http://www.sthda.com/english/rpkgs/factoextra/ (accessed on 16 August 2018).
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, 7191. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ersoz, E.; Lai, C.-Q.; Todhunter, R.J.; Tiwari, H.K.; Gore, M.A.; Bradbury, P.J.; Yu, J.; Arnett, D.K.; Ordovas, J.M.; et al. Mixed linear model approach adapted for genome-wide association studies. Nature Genetics 2010, 42, 355–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff. Fly (Austin) 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Konya, 2015–2016 | Konya, 2016–2017 | Turkey, 1991–2015 | Konya, 2015–2016 | Konya, 2016–2017 | Turkey, 1991–2015 |
|---|---|---|---|---|---|---|
| Temperature (temp) (°C) a | Temp (°C) | Temp (°C) b | Rainfall (mm) c | Rainfall (mm) | Rainfall (mm) d | |
| September | 22.8 | 17.1 | 19.0 | 35.8 | 11.2 | 23.1 |
| October | 15.3 | 13.2 | 13.7 | 34.4 | 0.0 | 48.3 |
| November | 7.5 | 4.9 | 7.0 | 5.8 | 16.6 | 58.0 |
| December | −0.1 | 0.5 | 2.1 | 8.0 | 26.8 | 73.0 |
| January | 1.6 | 0.2 | 0.1 | 37.0 | 9.0 | 65.6 |
| February | 4.9 | 3.4 | 1.3 | 0.4 | 69.2 | 60.0 |
| March | 8.5 | 8.2 | 5.3 | 37.8 | 31.0 | 61.6 |
| April | 15.2 | 12.7 | 10.4 | 9.6 | 33.2 | 62.7 |
| May | 18.4 | 16.7 | 15.2 | 38.4 | 41.2 | 54.6 |
| June | 23.7 | 24.4 | 19.5 | 15.0 | 4.8 | 34.7 |
| July | 26.6 | 27.7 | 22.9 | 0.2 | 0.0 | 15.1 |
| Total/average | 13.1 | 11.7 | 10.6 | 222.4 | 243.0 | 435.1 |
| Trait | 2016 | 2017 | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean | Range | CV | H2 | Mean | Range | CV | H2 | |
| Grain yield (g⋅m−2) | 259 | 200–341 | 9.7 | 0.32 | 290 | 241–392 | 9.9 | 0.56 |
| Harvest index | 0.4 | 0.24–0.66 | 10.9 | 0.63 | 0.34 | 0.27–0.41 | 6.3 | 0.64 |
| Biomass weight (g⋅m−2) | 671 | 537–827 | 9.1 | 0.39 | 865 | 684–1098 | 8.9 | 0.63 |
| Thousand kernel weight (g) | 32.1 | 24–42 | 10.5 | 0.75 | 41 | 33–50 | 8 | 0.90 |
| Grain volume weight (Kg⋅hL−1) | 65.6 | 52–77 | 7.2 | 0.91 | 74 | 68–77 | 2.3 | 0.76 |
| Awn length (cm) | 6 | 2.3–8.6 | 24.3 | 0.61 | 5.6 | 0.5–8.0 | 28.3 | 0.95 |
| Flag leaf length (cm) | 22.4 | 21.8–22.8 | 0.8 | 0.91 | 12 | 9.9–16.4 | 7.6 | 0.53 |
| Flag leaf width (cm) | 1 | 0.96–1.13 | 2.8 | 0.67 | 1 | 0.9–1.3 | 6.1 | 0.49 |
| Flag leaf area (cm2) | 18.9 | 17.6–19.7 | 2.2 | 0.85 | 10.1 | 7.7–14 | 11.6 | 0.52 |
| Stem diameter (mm) | 2.9 | 2.4–3.5 | 6.9 | 0.57 | 2.9 | 2.5–4.0 | 7.4 | 0.63 |
| Root length (cm) | 393 | 392–395 | 0.20 | 0.6 | 192.2 | 72–375 | 20 | 0.31 |
| Trait | SNP a | −log10 (p) | Alleles | SNP Effect | PVE (%) | Gene ID | Annotation |
|---|---|---|---|---|---|---|---|
| GY | S3A_686179591 | 4.08 | A/G | −14.28 | 10.7 | TraesCS3A01G445100 | F-box family protein |
| GY | S7A_112977027 | 5.24 | A/T | −19.17 | 12.8 | TraesCS7A01G158200.1 | Sentrin-specific protease |
| HI | S3A_593313534 | 13.56 | T/C | 0.08 | 16.0 | TraesCS3A01G343700 | WRKY transcription factor |
| HI | S6D_157451060 | 4.01 | A/G | −0.03 | 6.2 | TraesCS6D01G170900.1 | Cytochrome P450, putative |
| HI | S6D_462272376 | 12.01 | G/A | 0.02 | 14.5 | TraesCS6D01G382600.1 | LOB-domain protein-like |
| BMWT | S1D_441309135 | 4.82 | C/G | −105.34 | 14.4 | TraesCS1D01G357500.1 | Protein DETOXIFICATION |
| BMWT | S7B_450630784 | 4.06 | A/G | −25.88 | 10.7 | TraesCS7B01G242600.1 | F-box family protein |
| TKW | S2A_47781717 | 4.52 | G/A | 0.93 | 4.2 | TraesCS2A01G093500 | F-box family protein |
| TKW | S4A_625466381 | 4.12 | T/G | 1.21 | 15.3 | TraesCS4A01G347600 | Protein kinase family protein |
| TKW | S4D_509427923 | 4.91 | C/G | −1.72 | 10.1 | TraesCS4D01G364700 | Cytochrome P450 family protein |
| TKW | S6D_452410667 | 8.16 | A/G | −1.54 | 17.7 | TraesCS6D01G360800 | Protein kinase family protein |
| AWNLN | S4D_461573496 | 5.71 | T/C | 0.32 | 9.0 | TraesCS4D01G290700.1 | 60S ribosomal protein L18a |
| AWNLN | S5A_562540562 | 11.67 | C/T | −1.71 | 11.3 | TraesCS5A01G361300.1 | Guanine nucleotide exchange family protein |
| FLLN | S1B_667135914 | 4.38 | C/T | −0.16 | 20.8 | TraesCS1B01G447400 | Disease resistance protein RPM1 |
| FLW | S6D_16376439 | 4.85 | C/T | −0.02 | 13.3 | TraesCS6D01G040100.1 | Mitochondrial transcription termination factor-like |
| FLA | S1D_278097355 | 4.74 | G/C | 0.21 | 11.5 | TraesCS1D01G197200.1 | P-loop containing nucleoside triphosphate hydrolases superfamily protein |
| FLA | S6B_120860110 | 4.01 | G/A | 0.17 | 9.3 | TraesCS6B01G125800 | Cytochrome P450 family protein, expressed |
| FLA | S6B_120860130 | 4.01 | A/T | −0.17 | 9.3 | TraesCS6B01G125900 | Cytochrome P450 family protein, expressed |
| STMDIA | S1D_431523575 | 6.58 | A/G | −0.06 | 10.3 | TraesCS1D01G341500 | Disease resistance protein (NBS-LRR class) family |
| STMDIA | S3D_10133372 | 9.83 | G/T | −0.11 | 8.6 | TraesCS3D01G028500.1 | Leucine-rich repeat receptor-like protein kinase family protein |
| STMDIA | S6A_94238211 | 6.9 | T/G | 0.06 | 7.5 | TraesCS6A01G122200.1 | Protein kinase, putative |
| RTLN | S5B_669373985 | 4.62 | T/C | 0.27 | 6.9 | TraesCS5B01G502200 | GRAM domain-containing protein/ABA-responsive |
| RTLN | S5B_669374027 | 4.62 | T/C | 0.27 | 6.9 | TraesCS5B01G502200 | GRAM domain-containing protein/ABA-responsive |
| RTLN | S6D_431108774 | 4.01 | A/G | −0.27 | 5.8 | TraesCS6D01G332800.1 | Protein DETOXIFICATION |
| RTLN | S7A_94404310 | 4.01 | G/A | 0.52 | 7.5 | TraesCS7A01G143200.2 | Phosphatase 2C family protein |
| Trait | SNP a | −log10 (p) | Alleles | SNP Effect | PVE (%) | Gene-ID | Annotation |
|---|---|---|---|---|---|---|---|
| GY | S3A_25012018 | 4.81 | A/G | −20.02 | 12.7 | TraesCS3A01G047300 | F-box-domain-containing protein |
| GY | S3D_1203058 | 4.12 | T/G | 14.32 | 12.8 | TraesCS3D01G002700 | Disease resistance protein RPM1 |
| BMWT | S3A_25012018 | 6.08 | A/G | −59.44 | 14.4 | TraesCS3A01G047300 | F-box-domain-containing protein |
| TKW | S4B_11905230 | 8.94 | C/G | −1.11 | 3.9 | TraesCS4B01G016200.1 | LOB domain-containing protein, putative |
| TKW | S4B_637722874 | 5.17 | T/C | 0.86 | 2.0 | TraesCS4B01G344200.1 | Zinc finger (C3HC4-type RING finger) family protein |
| GVWT | S1A_522189599 | 4.11 | A/G | −0.55 | 2.5 | TraesCS1A01G334800 | Cytochrome P450 |
| GVWT | S4A_73454791 | 5.64 | C/T | −0.63 | 5.5 | TraesCS4A01G074200.2 | Microtubule associated protein family protein, putative, expressed |
| AWNLN | S5B_43896804 | 7.31 | C/T | −1.13 | 6.0 | TraesCS5B01G038700 | F-box family protein |
| AWNLN | S6B_643657 | 10.55 | C/T | −1.04 | 5.7 | TraesCS6B01G001000 | F-box family protein |
| FLLN | S1B_631203243 | 5.26 | A/G | −0.21 | 9.8 | TraesCS1B01G400600.1 | Rp1-like protein |
| FLLN | S2B_140752747 | 4.15 | G/C | 0.29 | 1.6 | TraesCS2B01G167500.1 | Cytochrome P450, putative |
| FLLN | S2D_642055122 | 4.12 | T/C | 0.25 | 5.9 | TraesCS2D01G579800 | protein kinase family protein |
| FLLN | S4A_612662321 | 5.3 | C/T | −0.23 | 5.3 | TraesCS4A01G325200 | F-box family protein |
| FLLN | S6D_463762312 | 5.72 | G/A | 0.25 | 6.0 | TraesCS6D01G386300 | Cytochrome P450, putative |
| FLW | S1A_516732460 | 6.99 | A/G | −0.03 | 9.5 | TraesCS1A01G326700.1 | Citrate-binding protein |
| FLW | S6B_26200560 | 7.13 | C/A | 0.03 | 12.3 | TraesCS6B01G042800 | F-box family protein |
| FLA | S1A_516732460 | 6.9 | A/G | −0.42 | 8.0 | TraesCS1A01G326700.1 | Citrate-binding protein |
| FLA | S2A_764065400 | 4.18 | G/T | −0.19 | 3.8 | TraesCS2A01G563200 | NBS-LRR resistance-like protein |
| STMDIA | S6B_610963076 | 5.7 | T/G | 0.06 | 7.9 | TraesCS6B01G346900-TraesCS6B01G347000 | NBS-LRR disease resistance protein and F-box protein-like |
| RTLN | S2D_620326979 | 4.22 | T/C | 192.21 | 9.9 | TraesCS2D01G541000.1 | Disease resistance protein RPM1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatta, M.; Morgounov, A.; Belamkar, V.; Baenziger, P.S. Genome-Wide Association Study Reveals Novel Genomic Regions for Grain Yield and Yield-Related Traits in Drought-Stressed Synthetic Hexaploid Wheat. Int. J. Mol. Sci. 2018, 19, 3011. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103011
Bhatta M, Morgounov A, Belamkar V, Baenziger PS. Genome-Wide Association Study Reveals Novel Genomic Regions for Grain Yield and Yield-Related Traits in Drought-Stressed Synthetic Hexaploid Wheat. International Journal of Molecular Sciences. 2018; 19(10):3011. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103011
Chicago/Turabian StyleBhatta, Madhav, Alexey Morgounov, Vikas Belamkar, and P. Stephen Baenziger. 2018. "Genome-Wide Association Study Reveals Novel Genomic Regions for Grain Yield and Yield-Related Traits in Drought-Stressed Synthetic Hexaploid Wheat" International Journal of Molecular Sciences 19, no. 10: 3011. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103011