A Genome-Wide Association Study Reveals Candidate Genes Related to Salt Tolerance in Rice (Oryza sativa) at the Germination Stage

 ,
,
Abstract
:1. Introduction
2. Results
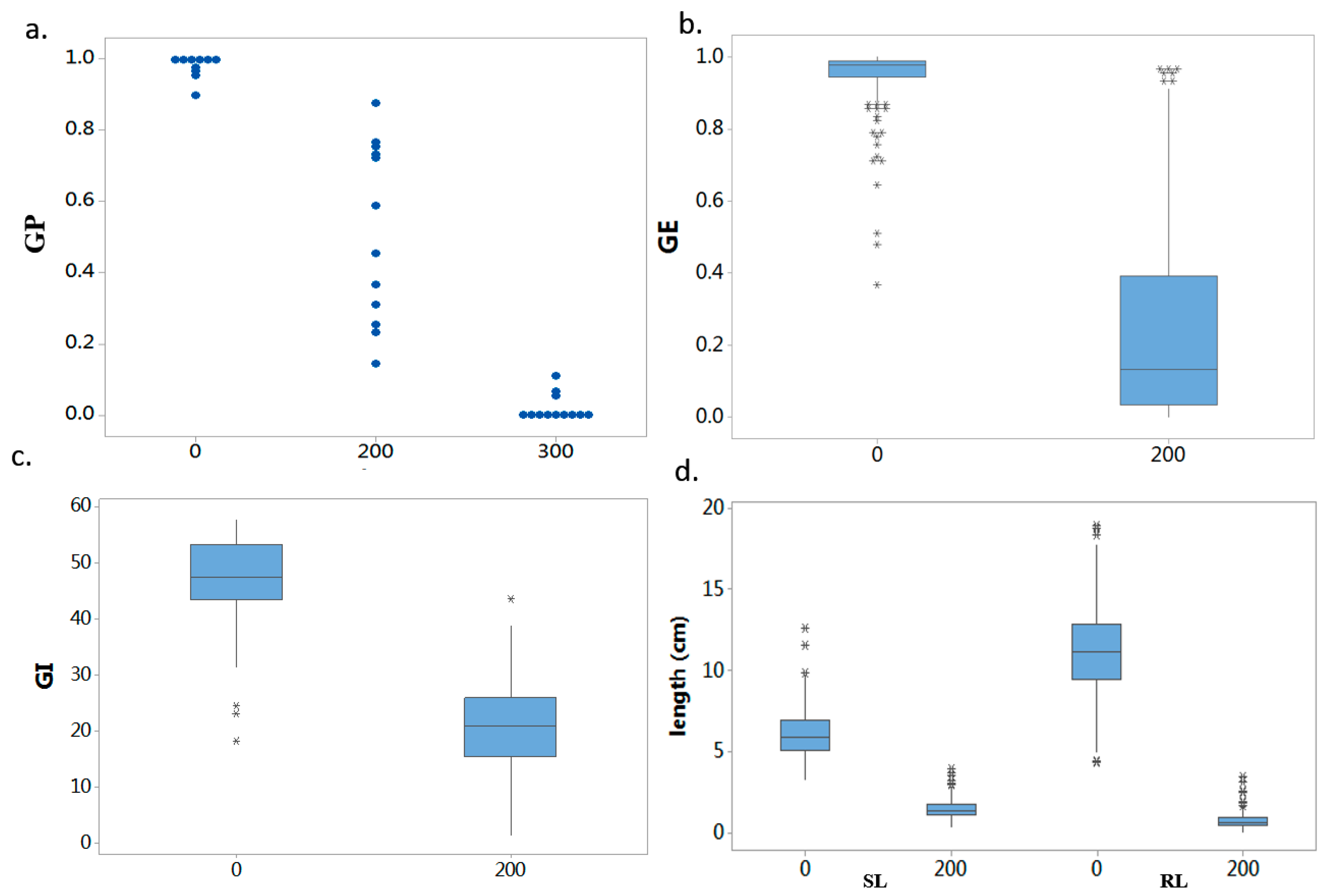
2.1. Phenotypic Screening and Evaluation
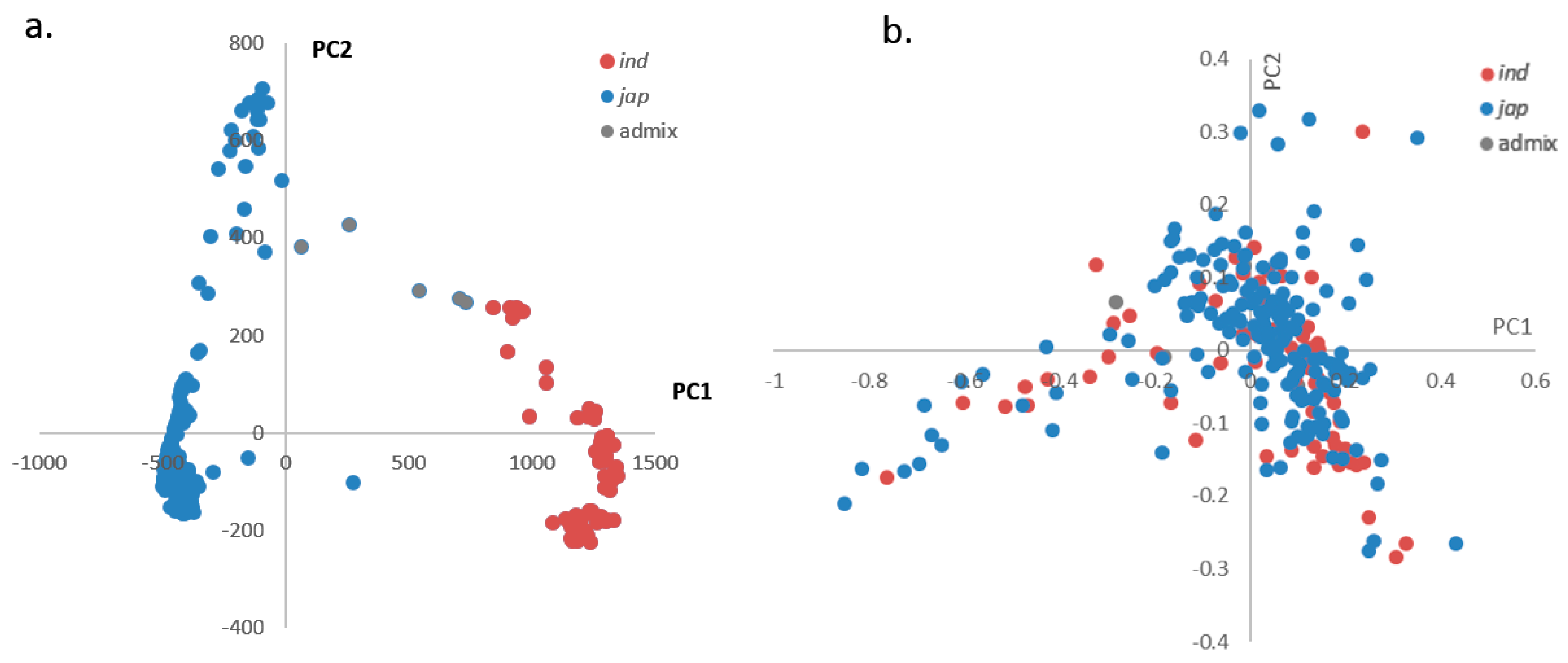
2.2. Principal Components Analysis (PCA)
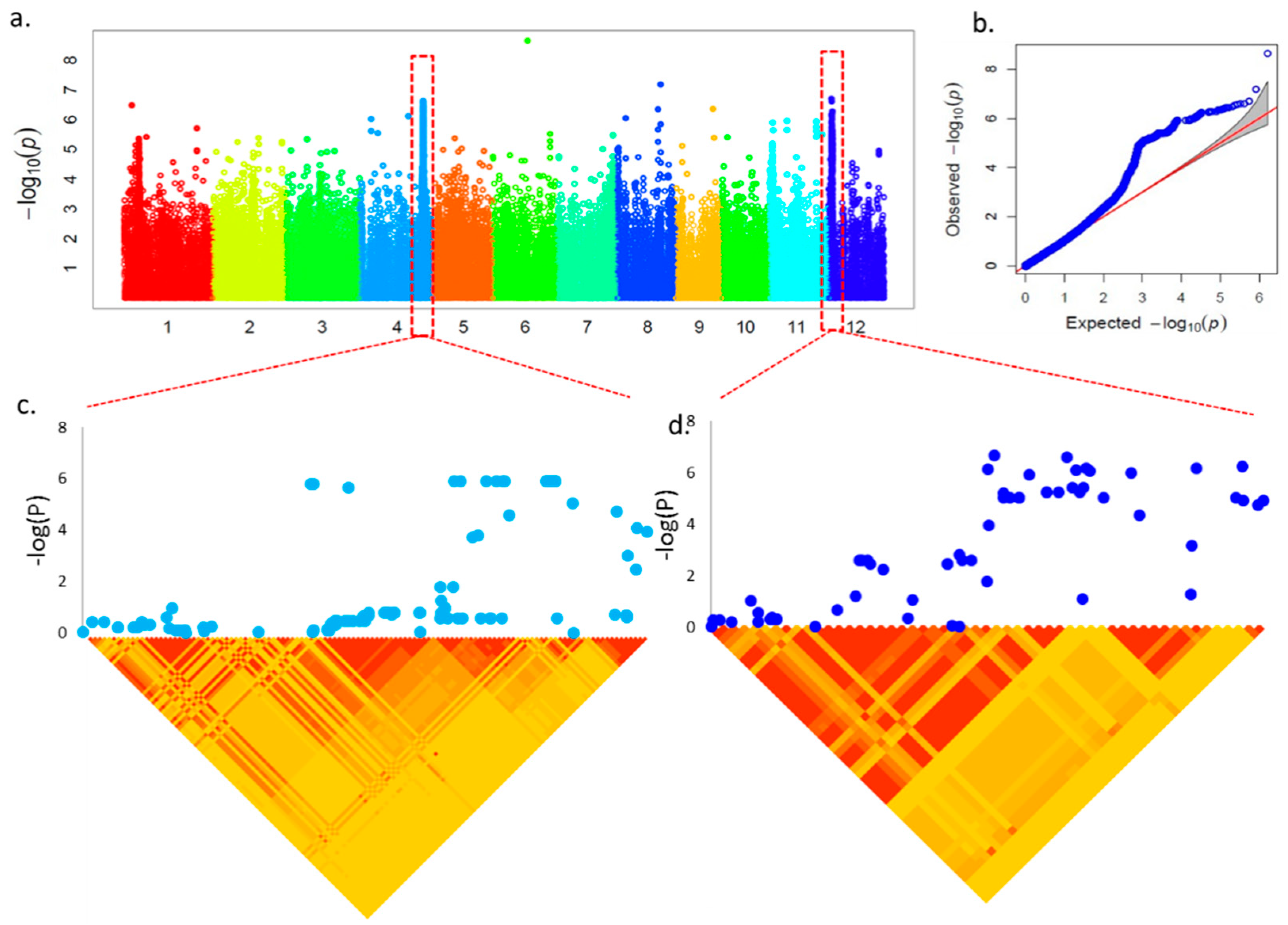
2.3. GWAS and Candidate Gene Identification
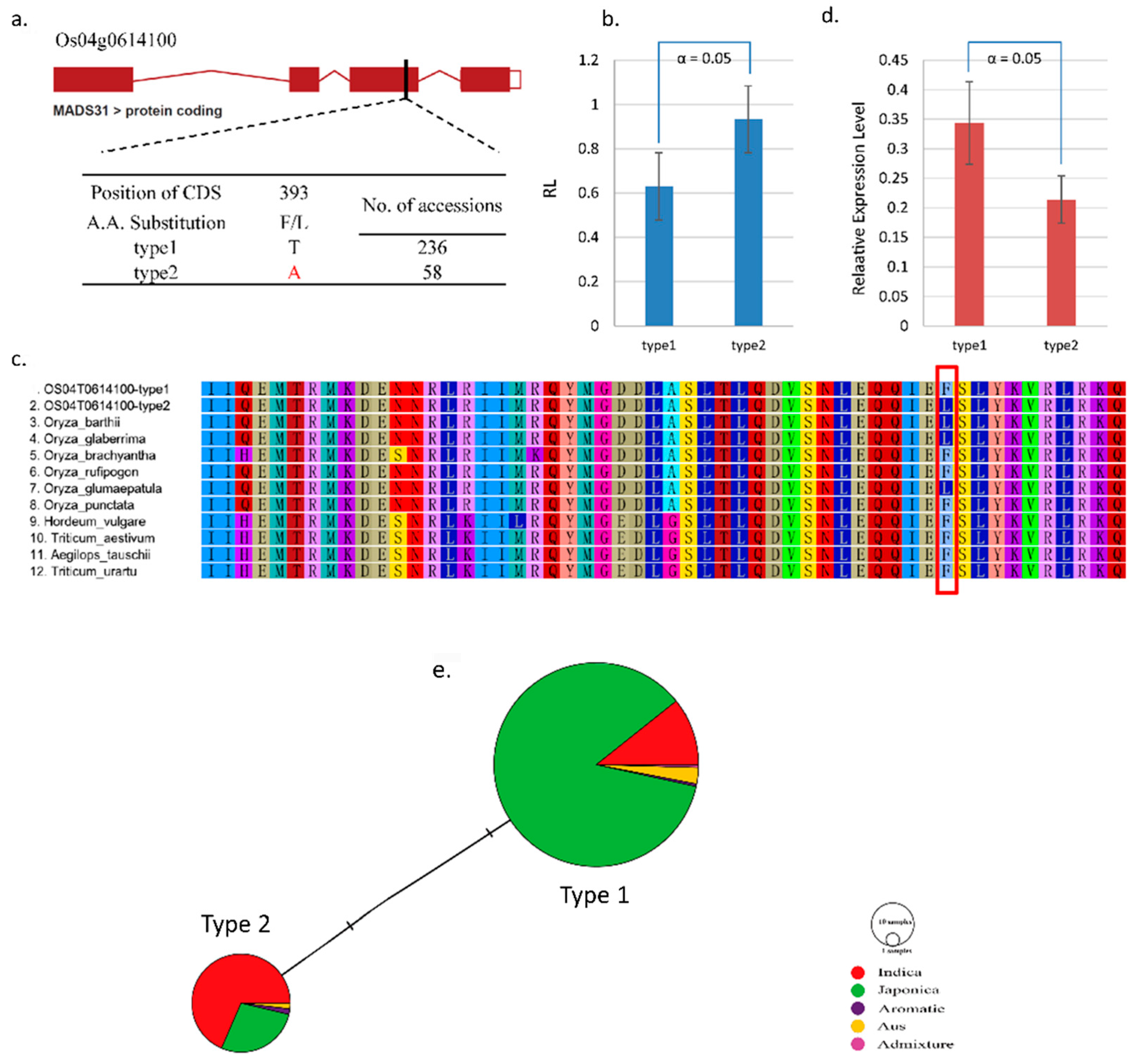
2.4. Natural Variations in Candidate Genes and Sequence Analysis
3. Discussion
3.1. Salt Tolerance at Rice Germination Stage
3.2. Salt Tolerance Is Not Strongly Correlated with Rice Subgroups
3.3. GWAS and Candidate Gene Identification
3.4. Novel Natural Variations of Candidate Genes
4. Materials and Methods
4.1. Materials
4.2. Whole Genome Re-Sequencing and Variation Detection
4.3. Evaluation of Salt Stress and Phenotyping
4.4. Principal Components and GWAS Analysis
4.5. Linkage Disequilibrium (LD) Block, Haplotype Analysis, and Expression Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GWAS | genome-wide association study |
| LD | linkage disequilibrium |
| NGS | next-generation sequencing |
| SNP | single-nucleotide polymorphism |
| INDEL | insertion and deletion |
| MAF | minor allele frequency |
| RL | root length |
| SL | shoot length |
| RS | root/shoot ratio |
| GP | Germination percentage |
| GE | Germination energy |
| GI | Germination index |
| CMLM | compressed mixed linear model |
| PCA | Principal component analysis |
References
- Sakadevan, K.; Nguyen, M.L. Extent, impact, and response to soil and water salinity in arid and semiarid regions. Adv. Agron. 2010, 109, 55. [Google Scholar]
- Mather, K.A.; Caicedo, A.L.; Polato, N.R.; Olsen, K.M.; McCouch, S.; Purugganan, M.D. The extent of linkage disequilibrium in rice (Oryza sativa L.). Genetics 2007, 177, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Z.; Cheng, J.; Lai, Y.; Wang, J.; Bao, Y.; Huang, J.; Zhang, H. QTL analysis of Na+ and K+ concentrations in roots and shoots under different levels of NaCl stress in rice (Oryza sativa L.). PLoS ONE 2012, 7, e51202. [Google Scholar] [CrossRef] [PubMed]
- Almansouri, M.; Kinet, J.M.; Lutts, S. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant and Soil 2001, 231, 243–254. [Google Scholar] [CrossRef]
- Rehman, S.; Harris, P.J.; Bourne, W.F.; Wilkin, J. The relationship between ions, vigour and salinity tolerance of acacia seeds. Plant Soil 2000, 220, 229–233. [Google Scholar] [CrossRef]
- Xu, S.; Hu, B.; He, Z.; Ma, F.; Feng, J.; Shen, W.; Yang, J. Enhancement of salinity tolerance during rice seed germination by presoaking with hemoglobin. Int. J. Mol. Sci. 2011, 12, 2488–2501. [Google Scholar] [CrossRef] [PubMed]
- Khodarahmpour, Z.; Ifar, M.; Motamedi, M. Effects of NaCl salinity on maize (Zea mays L.) at germination and early seedling stage. Afr. J. Biotechnol. 2014, 11, 298–304. [Google Scholar] [CrossRef]
- Mutlu, F.; Bozcuk, S. Salinity-induced changes of free and bound polyamine levels in sunflower (Helianthus annuus L.) roots differing in salt tolerance. Pak. J. Bot. 2007, 39, 1097–1102. [Google Scholar]
- Ulfat, M.; Athar, H.U.R.; Ashraf, M.; Akram, N.A.; Jamil, A. Appraisal of physiological and biochemical selection criteria for evaluation of salt tolerance in canola (Brassica napus L.). Pak. J. Bot. 2007, 39, 1593–1608. [Google Scholar]
- Khan, M.A.; Weber, D.J. Ecophysiology of High Salinity Tolerant Plants; Springer Science & Business Media: New York, NY, USA, 2006. [Google Scholar]
- Koyro, H.W. Ultrastructural Effects of Salinity in Higher Plants. In Salinity, Environment-Plants-Molecules; Springer: New York, NY, USA, 2002; ISBN 978-1-4020-0492-6. [Google Scholar]
- Gomes-Filho, E.; Lima, C.R.F.M.; Costa, J.H.; da Silva, A.C.M.; Lima, M.d.G.S.; de Lacerda, C.F.; Prisco, J.T. Cowpea ribonuclease, properties and effect of NaCl-salinity on its activation during seed germination and seedling establishment. Plant Cell Rep. 2008, 27, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Rizvi, Y. Effect of salinity, temperature, and growth regulators on the germination and early seedling growth of Atriplex griffithii var. stocksii. Can. J. Bot. 1994, 72, 475–479. [Google Scholar] [CrossRef]
- Yupsanis, T.; Moustakas, M.; Eleftheriou, P.; Damianidou, K. Protein phosphorylation-dephosphorylation in alfalfa seeds germinating under salt stress. J. Plant Physiol. 1994, 143, 234–240. [Google Scholar] [CrossRef]
- Othman, Y.; Al-Karaki, G.; Al-Tawaha, A.; Al-Horani, A. Variation in germination and ion uptake in barley genotypes under salinity conditions. World J. Agric. Sci. 2006, 2, 11–15. [Google Scholar]
- Wahid, A.; Rasul, E.; Rao, A.U.R. Germination of Seeds and Propagules under Salt Stress; Handbook of Plant and Crop Stress: Boca Raton, FL, USA, 1999; Volume 2, pp. 153–167. [Google Scholar]
- Foolad, M.; Hyman, J.; Lin, G. Relationships between cold-and salt-tolerance during seed germination in tomato, Analysis of response and correlated response to selection. Plant Breed. 1999, 118, 49–52. [Google Scholar] [CrossRef]
- Wang, Z.F.; Wang, J.F.; Bao, Y.M.; Wu, Y.Y.; Zhang, H.S. Quantitative trait loci controlling rice seed germination under salt stress. Euphytica 2011, 178, 297–307. [Google Scholar] [CrossRef]
- Csanádi, G.; Vollmann, J.; Stift, G.; Lelley, T. Seed quality QTLs identified in a molecular map of early maturing soybean. Theor. Appl. Genet. 2001, 103, 912–919. [Google Scholar] [CrossRef]
- Bai, C.; Liang, Y.; Hawkesford, M.J. Identification of QTLs associated with seedling root traits and their correlation with plant height in wheat. J. Exp. Bot. 2013, 64, 1745–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRose-Wilson, L.; Gaut, B.S. Mapping salinity tolerance during Arabidopsis thaliana germination and seedling growth. PLoS ONE 2011, 6, e22832. [Google Scholar] [CrossRef] [PubMed]
- Basnet, R.K.; Duwal, A.; Tiwari, D.N.; Xiao, D.; Monakhos, S.; Bucher, J.; Visser, R.G.F.; Groot, S.P.C.; Bonnema, G.; Maliepaard, C. Quantitative trait locus analysis of seed germination and seedling vigor in Brassica rapa reveals QTL hotspots and epistatic interactions. Front. Plant Sci. 2015, 6, 1032. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.M.L.; Tran, T.N.; Nguyen, T.K.T.; Williams, B.; Wurm, P.; Bellairs, S.; Mundree, S. Improvement of salinity stress tolerance in rice, challenges and opportunities. Agronomy 2016, 6, 54. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, J.; Bao, Y.; Wang, F.; Zhang, H. Quantitative trait loci analysis for rice seed vigor during the germination stage. Zhejiang Univ. Sci. B (Biomed. & Biotechnol.) 2010, 11, 958–964. [Google Scholar] [Green Version]
- Cheng, J.; He, Y.; Yang, B.; Lai, Y.; Wang, Z.; Zhang, H. Association mapping of seed germination and seedling growth at three conditions in indica rice (Oryza sativa L.). Euphytica 2015, 206, 103–115. [Google Scholar] [CrossRef]
- Abe, A.; Takagi, H.; Fujibe, T.; Aya, K.; Kojima, M.; Sakakibara, H. OsGA20ox1.; a candidate gene for a major QTL controlling seedling vigor in rice. Theor. Appl. Genet. 2012, 125, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Liu, B.; Zhao, H.; Wang, J.; Liu, H.; Sun, J.; Xing, J.; Zou, D. Identification of QTLs for salt tolerance at the germination and early seedling stage using linkage and association analysis in japonica rice. Chin. J. Rice Sci. 2014, 28, 358–366. [Google Scholar]
- Huang, X.; Wei, X.; Sang, T.; Zhao, Q.; Feng, Q.; Zhao, Y.; Li, C.; Zhu, C.; Lu, T.; Zhang, Z.; et al. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat. Genet. 2010, 42, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Tung, C.W.; Eizenga, G.C.; Wright, M.H.; Ali, M.L.; Price, A.H.; Norton, G.J.; Islam, M.R.; Reynolds, A.; Mezey, J.; et al. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat. Commun. 2011, 2, 467. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Park, E.J.; Chung, J.W.; Park, Y.J.; Chung, I.M.; Ahn, J.K.; Kim, G.H. Association analysis of the amino acid contents in rice. J. Integr. Plant Biol. 2009, 51, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Na, Y.W.; Kwon, S.W.; Park, Y.J. Association analysis of seed longevity in rice under conventional and high-temperature germination conditions. Plant Syst. Evol. 2014, 300, 389–402. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, Y.; Wei, X.; Li, C.; Wang, A.; Zhao, Q.; Li, W.; Guo, Y.; Deng, L.; Zhu, C.; et al. Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm. Nat. Genet. 2012, 44, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Feng, F.; Wei, H.; Mei, H.; Xu, K.; Chen, S.; Li, T.; Liang, X.; Liu, H.; Luo, L. Genome-wide association study for plant height and grain yield in rice under contrasting moisture regimes. Front. Plant Sci. 2016, 7, 1801. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Singh, A.; Mithra, S.V.; Krishnamurthy, S.L.; Parida, S.K.; Jain, S.; Tiwari, K.K.; Kumar, P.; Rao, A.R.; Sharma, S.K.; et al. Genome wide association mapping of salinity tolerance in rice (Oryza sativa). DNA Res. 2015, 22, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhao, W.; He, Q.; Kim, T.S.; Park, Y.J. Genome-wide association study and gene set analysis for understanding candidate genes involved in salt tolerance at the rice seedling stage. Mol. Genet. Genomics 2017, 292, 1391–1403. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Gao, L.; Wu, Z.; Zhang, X.; Wang, M.; Zhang, C.; Zhang, F.; Zhou, Y.; Li, Z. Genome-wide association study of salt tolerance at the seed germination stage in rice. BMC Plant Biol. 2017, 17, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveed, S.A.; Zhang, F.; Zhang, J.; Zheng, T.Q.; Meng, L.J.; Pang, Y.L.; Xu, J.L.; Li, Z.K. Identification of QTN and candidate genes for salinity tolerance at the germination and seedling stages in rice by genome-wide association analyses. Sci. Rep. 2018, 8, 6505. [Google Scholar] [CrossRef] [PubMed]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT, genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Arora, R.; Lama, T.; Nijhawan, A.; Khurana, J.P.; Tyagi, A.K.; Kapoor, S. Genome-wide identification, organization and phylogenetic analysis of Dicer-like, argonaute and RNA-dependent RNA polymerase gene families and their expression analysis during reproductive development and stress in rice. BMC Genom. 2008, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Ricachenevsky, F.K.; Sperotto, R.A.; Menguer, P.K.; Sperb, E.R.; Lopes, K.L.; Fett, J.P. ZINC-INDUCED FACILITATOR-LIKE family in plants, lineage-specific expansion in monocotyledons and conserved genomic and expression features among rice (Oryza sativa) paralogs. BMC Plant Biol. 2011, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Nakayama, H.; Shinmyo, A.; Yoshida, K. Expression of OsHAK genes encoding potassium ion transporters in rice. Plant Biotechnol. 2008, 25, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, L.; Wang, Y.; Zhang, J.; Liang, Y.; Xu, D. A computational systems biology study for understanding salt tolerance mechanism in rice. PLoS ONE 2013, 8, e64929. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Agarwal, P.; Ray, S.; Singh, A.K.; Singh, V.P.; Tyagi, A.K.; Kapoor, S. MADS-box gene family in rice, genome-wide identification, organization and expression profiling during reproductive development and stress. BMC Genom. 2007, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Shannon, M.; Grieve, C. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica 2002, 127, 235–245. [Google Scholar] [CrossRef]
- Ferdose, J.; Kawasaki, M.; Taniguchi, M.; Miyake, H. Differential sensitivity of rice cultivars to salinity and its relation to ion accumulation and root tip structure. Plant Prod. Sci. 2009, 12, 453–461. [Google Scholar] [CrossRef]
- Lee, K.S.; Choi, W.Y.; Ko, J.C.; Kim, T.S.; Gregorio, G.B. Salinity tolerance of japonica and indica rice (Oryza sativa L.) at the seedling stage. Planta. 2003, 216, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Platten, J.D.; Egdane, J.A.; Ismail, A.M. Salinity tolerance.; Na+ exclusion and allele mining of HKT1;5 in Oryza sativa and O. glaberrima, many sources.; many genes.; one mechanism? BMC Plant Biol. 2013, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Leon, T.B.; Steven, L.; Prasanta, K.S. Molecular dissection of seedling salinity tolerance in rice (Oryza sativa L.) using a high-density GBS-based SNP linkage map. Rice 2016, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.X.; Yanagihara, S.; Zhuang, J.Y.; Senboku, T.; Zheng, K.L.; Yashima, S. Identification of QTLs for salt tolerance in rice via molecular markers. Chin. J. Rice Sci. 1998, 12, 72–78. [Google Scholar]
- Cai, H.W.; Morishima, H. QTL clusters reflect character associations in wild and cultivated rice. Theor. Appl. Genet. 2002, 104, 1217–1228. [Google Scholar] [PubMed]
- Courtois, B.; Audebert, A.; Dardou, A.; Roques, S.; Ghneim-Herrera, T.; Droc, G.; Frouin, J.; Rouan, L.; Goze, E.; Kilian, A.; et al. Genome-wide association mapping of root traits in a japonica rice panel. PLoS ONE 2013, 8, e78037. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; DeClerck, G.; Tian, F.; Spooner, W.; McCouch, S.; Buckler, E. PICARA.; an analytical pipeline providing probabilistic inference about a priori candidates genes underlying genome-wide association QTL in plants. PLoS ONE 2012, 7, e46596. [Google Scholar] [CrossRef] [PubMed]
- Sur, I.; Tuupanen, S.; Whitington, T.; Aaltonen, L.A.; Taipale, J. Lessons from functional analysis of genome-wide association studies. Cancer Res. 2013, 73, 4180–4184. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, X.; Ge, S.; Jensen, J.D.; Hu, F.; Li, X.; Dong, Y.; Gutenkunst, R.N.; Fang, L.; Huang, L.; et al. Resequencing 50 accessions of cultivated and wild rice yields markers for identifying agronomically important genes. Nat. Biotechnol. 2012, 30, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Reddy, I.N.B.L.; Kim, B.K.; Yoon, I.S.; Kim, K.H.; Kwon, T.R. Salt tolerance in rice, focus on mechanisms and approaches. Rice Sci. 2017, 24, 123–144. [Google Scholar] [CrossRef]
- Yang, Y.; Zhong, J.; Ouyang, Y.D.; Yao, J. The integrative expression and co-expression analysis of the AGO gene family in rice. Gene 2013, 528, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Inahashi, H.; Shelley, I.J.; Yamauchi, T.; Nishiuchi, S.; Takahashi-Nosaka, M.; Matsunami, M.; Ogawa, A.; Noda, Y.; Inukai, Y. OsPIN2, which encodes a member of the auxin efflux carrier proteins, is involved in root elongation growth and lateral root formation patterns via the regulation of auxin distribution in rice. Physiol. Plantarum 2018. [Google Scholar] [CrossRef] [PubMed]
- Rebecca, M.D.; Patrick, A.R.; Patricia, M.M.; Jan, E.L. Germins, a diverse protein family important for crop improvement. Plant Sci. 2009, 177, 499–510. [Google Scholar]
- Kothari, K.S.; Dansana, P.K.; Giri, J.; Tyagi, A.K. Rice stress associated protein 1 (OsSAP1) interacts with aminotransferase (OsAMTR1) and pathogenesis-related 1a protein (OsSCP) and regulates abiotic stress responses. Front. Plant Sci. 2016, 7, 1057. [Google Scholar] [CrossRef] [PubMed]
- Konishi, S.; Izawa, T.; Lin, SY.; Ebana, K.; Fukuta, Y.; Sasaki, T.; Yano, M. An SNP caused loss of seed shattering during rice domestication. Science 2006, 312, 1392–1396. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Zhang, Z.; Chai, X.; Wang, H.; Wang, Y. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Chung, H.K.; Cho, G.T.; Ma, K.H.; Chandrabalan, D.; Gwag, J.G.; Kim, T.S.; Cho, E.G.; Park, Y.J. PowerCore, a program applying the advanced M strategy with a heuristic search for establishing core sets. Bioinformatics 2007, 23, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Cho, G.T.; Ma, K.H.; Chung, J.W.; Gwag, J.G.; Park, Y.J. Development of an allele-mining set in rice using a heuristic algorithm and SSR genotype data with least redundancy for the post-genomic era. Mol. Breed. 2010, 26, 639–651. [Google Scholar] [CrossRef]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; He, Q.; Kim, K.W.; Yoon, M.Y.; Ra, W.H.; Li, F.P.; Tong, W.; Yu, J.; Oo, W.H.; Choi, B.; et al. Genome-wide resequencing of KRICE_CORE reveals their potential for future breeding.; as well as functional and evolutionary studies in the post-genomic era. BMC Genom. 2016, 17, 408. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL, software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious basic, an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Posada, D.; Crandall, K.A. TCS, a computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. PopART, Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Salinity Level (NaCl/mM) | Mean ± SD a | Range | Median | IQR b |
|---|---|---|---|---|---|
| GP | 0 | 0.97 ± 0.06 | 0.47–1.00 | 0.99 | 0.97–1.00 |
| 200 | 0.87 ± 0.18 | 0.14–1.00 | 0.94 | 0.83–0.98 | |
| GE | 0 | 0.95 ± 0.08 | 0.37–1.00 | 0.98 | 0.94–0.99 |
| 200 | 0.25 ± 0.29 | 0–0.97 | 0.13 | 0.03–0.39 | |
| GI | 0 | 47.66 ± 6.71 | 18.33–57.87 | 47.55 | 43.40–53.44 |
| 200 | 20.66 ± 8.53 | 1.51–43.72 | 21.05 | 15.48–26.03 | |
| SL | 0 | 6.07 ± 1.38 | 3.25–12.62 | 5.82 | 5.08–6.90 |
| 200 | 1.45 ± 0.63 | 0.38–3.87 | 1.29 | 1.03–1.76 | |
| RL | 0 | 11.28 ± 2.89 | 4.28–18.89 | 11.11 | 9.46–12.81 |
| 200 | 0.69 ± 0.49 | 0.013–3.44 | 0.55 | 0.39–0.87 |
| Trait | GP | GE | GI | SL | RL | |
|---|---|---|---|---|---|---|
| Control | GP | |||||
| GE | 0.946 *** | |||||
| GI | 0.624 *** | 0.710 *** | ||||
| SL | −0.001 ns | 0.037 ns | 0.168 ** | |||
| RL | 0.073 ns | 0.087 ns | 0.110 ns | 0.254 *** | ||
| 200 mM | GP | |||||
| GE | 0.394 *** | |||||
| GI | 0.758 *** | 0.849 *** | ||||
| SL | 0.439 *** | 0.738 *** | 0.725 *** | |||
| RL | 0.357 *** | 0.618 *** | 0.594 *** | 0.712 *** |
| Chr_Pos a | Trait | p-Value | FDR b | Gene ID | Description |
|---|---|---|---|---|---|
| chr02_19605493 | R/S | 1.72 × 10−7 | 0.00779 | Os02g0532500 | Germin family protein, Germin-like protein 2-4 |
| Os02g0532900 | Glycoside hydrolase family 17 protein | ||||
| Os02g0533300 | Carbonic anhydrase, CAH1-like domain, containing protein | ||||
| Os02g0533800 | Similar to ATPase inhibitor | ||||
| chr04_31168058 | RL | 2.42 × 10−7 | 0.00859 | Os04g0612900 | Vacuolar ATPase assembly integral membrane protein VMA21-like domain-containing protein |
| Os04g0613900 | Similar to Potassium transporter 18, OsHAK11 | ||||
| Os04g0614000 | Similar to Peroxisomal 2,4-dienoyl-CoA reductase | ||||
| Os04g0614100 | MADS-box domain-containing protein, OsMADS31 | ||||
| Os04g0614600 | Similar to Viroid RNA-binding protein, aminotransferase | ||||
| Os04g0614500 | Pyridoxal phosphate-dependent transferase, major region, subdomain 1 domain-containing protein | ||||
| Os04g0615100 | Similar to Lecithine cholesterol acyltransferase-like protein | ||||
| Os04g0615700 | Protein argonaute 2, OsAGO2 | ||||
| chr12_1628276 | RL, R/S, R-R/S | 2.02 × 10−7 | 0.00859 | Os12g0133100 | Major facilitator superfamily protein, OsZIFL12 |
| Os12g0133300 | zinc-induced facilitator-like 13, OsZIFL13 | ||||
| Os12g0133400 | 4′-phosphopantetheinyl transferase domain-containing protein | ||||
| Os12g0133700 | Stress-activated protein kinase pathway-regulating phosphatase 1 | ||||
| Os12g0133800 | Similar to Auxin efflux carrier protein, OsPIN1d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Zhao, W.; Tong, W.; He, Q.; Yoon, M.-Y.; Li, F.-P.; Choi, B.; Heo, E.-B.; Kim, K.-W.; Park, Y.-J. A Genome-Wide Association Study Reveals Candidate Genes Related to Salt Tolerance in Rice (Oryza sativa) at the Germination Stage. Int. J. Mol. Sci. 2018, 19, 3145. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103145
Yu J, Zhao W, Tong W, He Q, Yoon M-Y, Li F-P, Choi B, Heo E-B, Kim K-W, Park Y-J. A Genome-Wide Association Study Reveals Candidate Genes Related to Salt Tolerance in Rice (Oryza sativa) at the Germination Stage. International Journal of Molecular Sciences. 2018; 19(10):3145. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103145
Chicago/Turabian StyleYu, Jie, Weiguo Zhao, Wei Tong, Qiang He, Min-Young Yoon, Feng-Peng Li, Buung Choi, Eun-Beom Heo, Kyu-Won Kim, and Yong-Jin Park. 2018. "A Genome-Wide Association Study Reveals Candidate Genes Related to Salt Tolerance in Rice (Oryza sativa) at the Germination Stage" International Journal of Molecular Sciences 19, no. 10: 3145. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103145