Endoplasmic Reticulum (ER) Stress and Unfolded Protein Response (UPR) in Mammalian Oocyte Maturation and Preimplantation Embryo Development

Abstract
:1. Introduction
2. Endoplasmic Reticulum Stress and Unfolded Protein Response Signaling Pathways
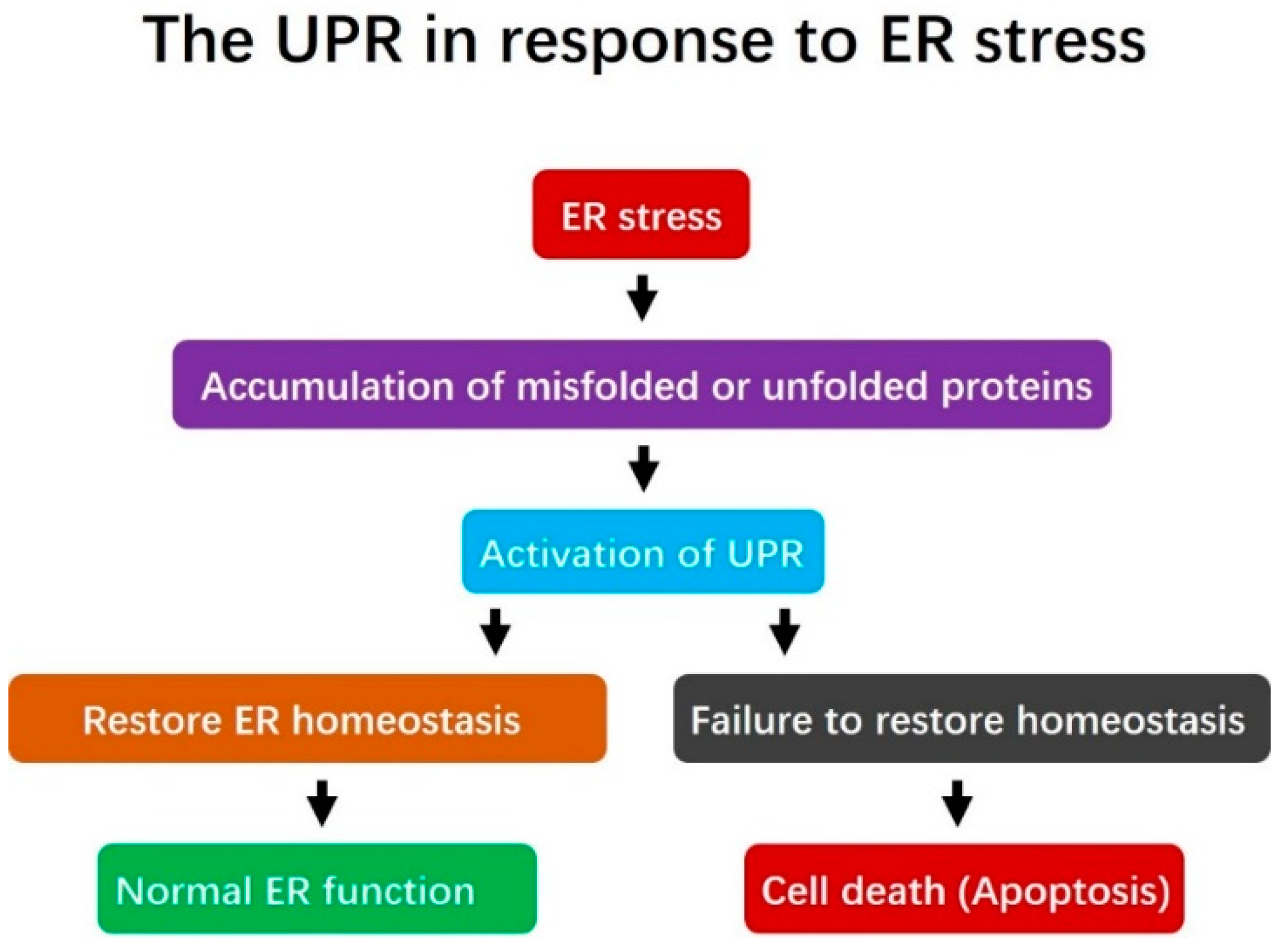
2.1. Endoplasmic Reticulum Stress and the Unfolded Protein Response
2.2. The Unfolded Protein Response Signaling Pathway
2.2.1. The PERK Signaling Pathway
2.2.2. The ATF6 Signaling Pathway
2.2.3. The IRE1 Signaling Pathway
3. Endoplasmic Reticulum Stress and the Unfolded Protein Response Are Intrinsic in Mammalian Oocytes and Preimplantation Embryos
3.1. Endoplasmic Reticulum Stress and the Unfolded Protein Response in Oocytes
3.2. Endoplasmic Reticulum Stress and the Unfolded Protein Response in Preimplantation Embryos
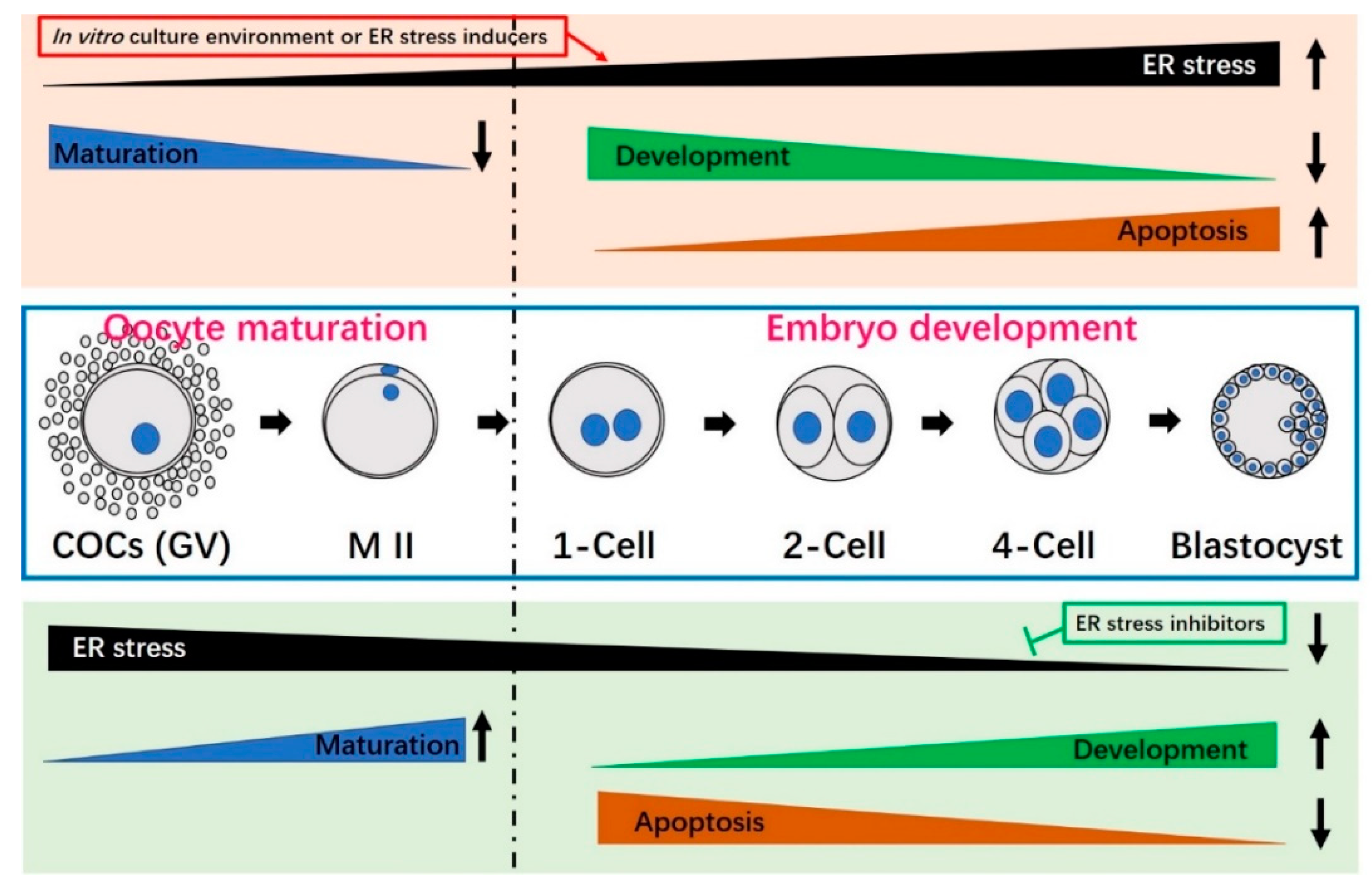
4. Activation and Induction of Endoplasmic Reticulum Stress in Oocytes and Preimplantation Embryos
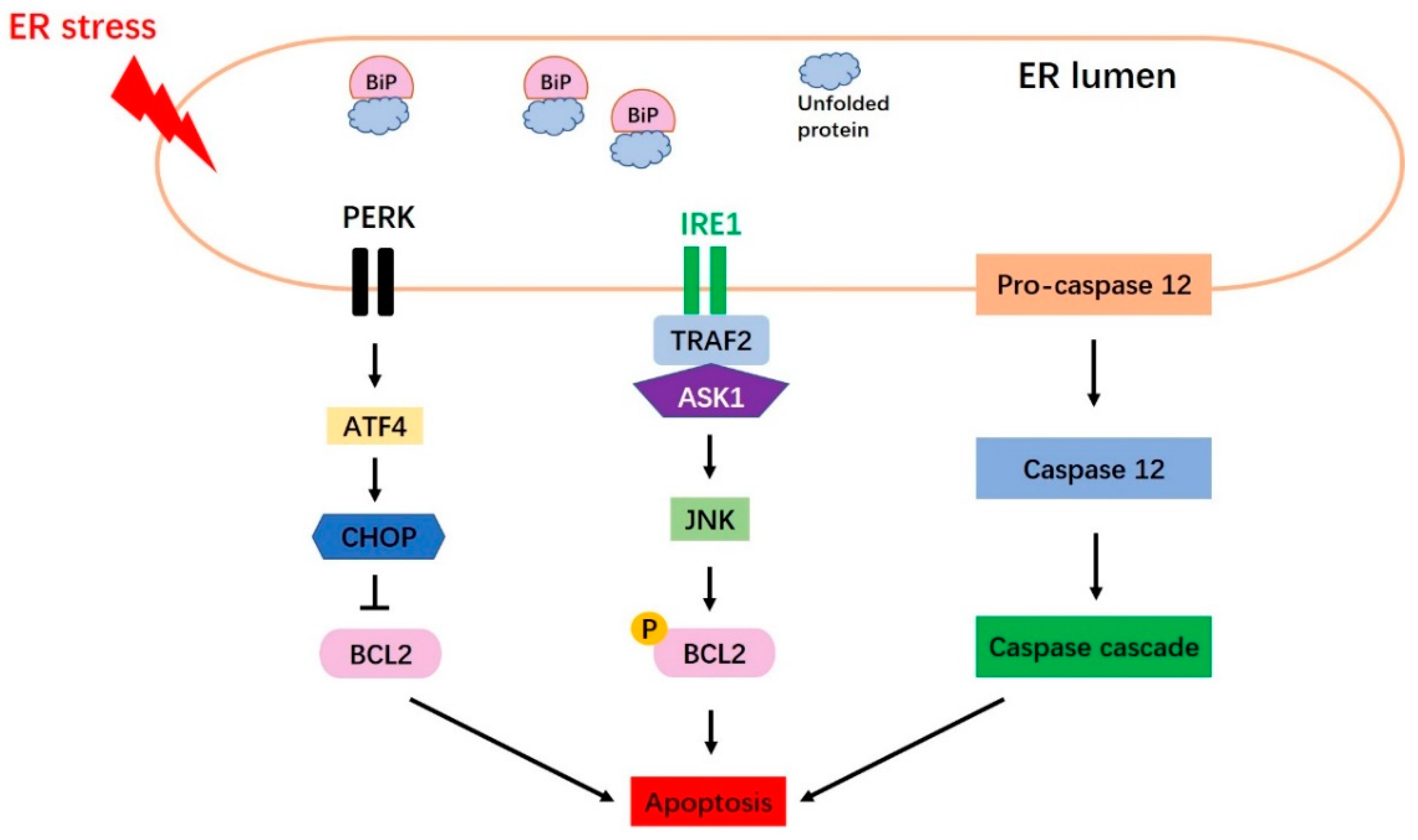
5. Endoplasmic Reticulum Stress Induces Apoptosis
5.1. Three Main Canonical Apoptosis Pathways Are Induced by Endoplasmic Reticulum Stress
5.1.1. The CHOP Pathway
5.1.2. The JNK Pathway
5.1.3. The Caspase 12 Pathway
5.2. Endoplasmic Reticulum Stress-Mediated Apoptosis in Oocytes and Preimplantation Embryos
6. Relief of Endoplasmic Reticulum Stress Reduces Apoptosis, Improves Oocyte Maturation and Enhances the Developmental Potential of Embryos
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASK1 | Apoptosis signal-regulating kinase 1 |
| ATF4 | Activating transcription factor 4 |
| ATF6 | Activating transcription factor 6 |
| BiP | Immunoglobulin-binding protein |
| bZip | Basic leucine zipper |
| CHOP | C/EBP homologous protein |
| COCs | Cumulus-oocyte complexes |
| DNA | Deoxyribonucleic acid |
| eIF2α | Eukaryotic translation initiation factor 2 alpha |
| ER | Endoplasmic reticulum |
| ERAD | ER-associated degradation |
| ERQC | Endoplasmic reticulum quality control |
| ERSE | ER stress response element |
| GADD153 | Growth arrest- and DNA damage-inducible gene 153 |
| GDP | Guanosine diphosphate |
| GRP78 | Glucose-regulated protein 78 |
| GSH | Glutathione |
| GTP | Guanosine triphosphate |
| GV | Germinal vesicle |
| ICM | Inner cell mass |
| IRE1 | Inositol-requiring enzyme 1 |
| IVC | In vitro culture |
| IVF | In vitro fertilization |
| IVM | In vitro maturation |
| IVP | In vitro-produced |
| JNK | Jun N-terminal kinase |
| M I | Metaphase I |
| M II | Metaphase II |
| mRNA | Messenger ribonucleic acid |
| PA | Parthenogenetic activation |
| PERK | Double-stranded RNA-activated protein kinase-like ER kinase |
| ROS | Reactive oxygen species |
| S1P | Site-1 protease |
| S2P | Site-2 protease |
| SCNT | Somatic cell nuclear transfer |
| TM | Tunicamycin |
| TRAF2 | TNF receptor-associated factor 2 |
| TUDCA | Tauroursodeoxycholic acid |
| UPR | Unfolded protein response |
| XBP1 | X-box binding protein-1 |
| XBP1-s | Spliced XBP1 |
| XBP1-u | Unspliced XBP1 |
References
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, R.J. Regulation of mRNA translation by protein folding in the endoplasmic reticulum. Trends Biochem. Sci. 2004, 29, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, J.Y.; Kim, J.W.; Yang, S.G.; Jung, J.M.; Kim, M.J.; Kang, M.J.; Cho, Y.H.; Wee, G.; Yang, H.Y.; et al. Melatonin improves the meiotic maturation of porcine oocytes by reducing endoplasmic reticulum stress during in vitro maturation. J. Pineal Res. 2018, 64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Diao, Y.F.; Oqani, R.K.; Han, R.X.; Jin, D.I. Effect of endoplasmic reticulum stress on porcine oocyte maturation and parthenogenetic embryonic development in vitro. Biol. Reprod. 2012, 86, 128. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, H.; Chiang, W.C.; Felden, J.; Nguyen, A.; Lin, J.H. ER stress and unfolded protein response in ocular health and disease. FEBS J. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Ghemrawi, R.; Battaglia-Hsu, S.F.; Arnold, C. Endoplasmic Reticulum Stress in Metabolic Disorders. Cells 2018, 7. [Google Scholar] [CrossRef]
- Schroder, M.; Kaufman, R.J. The mammalian unfolded protein response. Annu. Rev. Biochem. 2005, 74, 739–789. [Google Scholar] [CrossRef]
- Guzel, E.; Arlier, S.; Guzeloglu-Kayisli, O.; Tabak, M.S.; Ekiz, T.; Semerci, N.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Endoplasmic Reticulum Stress and Homeostasis in Reproductive Physiology and Pathology. Int. J. Mol. Sci. 2017, 18, 792. [Google Scholar] [CrossRef]
- Zeng, F.; Schultz, R.M. RNA transcript profiling during zygotic gene activation in the preimplantation mouse embryo. Dev. Biol. 2005, 283, 40–57. [Google Scholar] [CrossRef] [Green Version]
- Latham, K.E. Stress signaling in mammalian oocytes and embryos: A basis for intervention and improvement of outcomes. Cell Tissue Res. 2016, 363, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.B.; Choi, S.A.; Sim, B.W.; Kim, J.S.; Mun, S.E.; Jeong, P.S.; Yang, H.J.; Lee, Y.; Park, Y.H.; Song, B.S.; et al. Developmental competence of bovine early embryos depends on the coupled response between oxidative and endoplasmic reticulum stress. Biol. Reprod. 2014, 90. [Google Scholar] [CrossRef]
- Lin, T.; Lee, J.E.; Oqani, R.K.; Kim, S.Y.; Cho, E.S.; Jeong, Y.D.; Baek, J.J.; Jin, D.I. Tauroursodeoxycholic acid improves pre-implantation development of porcine SCNT embryo by endoplasmic reticulum stress inhibition. Reprod. Biol. 2016, 16, 269–278. [Google Scholar] [CrossRef]
- Sharma, A.; Agrawal, H.; Mullani, N.; Sandhu, A.; Singh, M.K.; Chauhan, M.S.; Singla, S.K.; Palta, P.; Manik, R.S. Supplementation of tauroursodeoxycholic acid during IVC did not enhance in vitro development and quality of buffalo IVF embryos but combated endoplasmic reticulum stress. Theriogenology 2015, 84, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Song, B.S.; Yoon, S.B.; Sim, B.W.; Kim, Y.H.; Cha, J.J.; Choi, S.A.; Jeong, K.J.; Kim, J.S.; Huh, J.W.; Lee, S.R.; et al. Valproic acid enhances early development of bovine somatic cell nuclear transfer embryos by alleviating endoplasmic reticulum stress. Reprod. Fertil. Dev. 2014, 26, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Diao, Y.F.; Kim, H.R.; Jin, D.I. Inhibition of endoplasmic reticulum stress improves mouse embryo development. PLoS ONE 2012, 7, e40433. [Google Scholar] [CrossRef]
- Michalak, M.; Gye, M.C. Endoplasmic reticulum stress in periimplantation embryos. Clin. Exp. Reprod. Med. 2015, 42, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, I.; Shah, S.Z.; Jin, Y.; Li, Z.S.; Ullah, O.; Fang, N.Z. Reactive oxygen species-mediated unfolded protein response pathways in preimplantation embryos. J. Vet. Sci. 2017, 18, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.H.; Zhang, K.Z.; Kaufman, R.J. The unfolded protein response—A stress signaling pathway of the endoplasmic reticulum. J. Chem. Neuroanat. 2004, 28, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Oslowski, C.M.; Urano, F. Measuring ER stress and the unfolded protein response using mammalian tissue culture system. Methods Enzymol. 2011, 490, 71–92. [Google Scholar]
- Harding, H.P.; Zhang, Y.H.; Bertolotti, A.; Zeng, H.Q.; Ron, D. Perk is essential for translational regulation and cell survival during the unfolded protein response. Mol. Cell 2000, 5, 897–904. [Google Scholar] [CrossRef]
- Harding, H.P.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.; et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol. Cell 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Han, J.; Back, S.H.; Hur, J.; Lin, Y.H.; Gildersleeve, R.; Shan, J.; Yuan, C.L.; Krokowski, D.; Wang, S.; Hatzoglou, M.; et al. ER-stress-induced transcriptional regulation increases protein synthesis leading to cell death. Nat. Cell Biol. 2013, 15, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, H.; Okada, T.; Haze, K.; Yanagi, H.; Yura, T.; Negishi, M.; Mori, K. Endoplasmic reticulum stress-induced formation of transcription factor complex ERSF including NF-Y (CBF) and activating transcription factors 6alpha and 6beta that activates the mammalian unfolded protein response. Mol. Cell. Biol. 2001, 21, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Abraham, T.; Pin, C.L.; Watson, A.J. Embryo collection induces transient activation of XBP1 arm of the ER stress response while embryo vitrification does not. Mol. Hum. Reprod. 2012, 18, 229–242. [Google Scholar] [CrossRef]
- Basseri, S.; Austin, R.C. Endoplasmic reticulum stress and lipid metabolism: Mechanisms and therapeutic potential. Biochem. Res. Int. 2012, 2012, 841362. [Google Scholar] [CrossRef]
- Lee, A.H.; Iwakoshi, N.N.; Glimcher, L.H. XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Mol. Cell. Biol. 2003, 23, 7448–7459. [Google Scholar] [CrossRef]
- Wu, L.L.; Russell, D.L.; Norman, R.J.; Robker, R.L. Endoplasmic reticulum (ER) stress in cumulus-oocyte complexes impairs pentraxin-3 secretion, mitochondrial membrane potential (DeltaPsi m), and embryo development. Mol. Endocrinol. 2012, 26, 562–573. [Google Scholar] [CrossRef]
- Yang, X.; Wu, L.L.; Chura, L.R.; Liang, X.Y.; Lane, M.; Norman, R.J.; Robker, R.L. Exposure to lipid-rich follicular fluid is associated with endoplasmic reticulum stress and impaired oocyte maturation in cumulus-oocyte complexes. Fertil. Steril. 2012, 97, 1438–1443. [Google Scholar] [CrossRef]
- Basar, M.; Bozkurt, I.; Guzeloglu-Kayisli, O.; Sozen, B.; Tekmen, I.; Schatz, F.; Arici, A.; Lockwood, C.J.; Kayisli, U.A. Unfolded protein response prevents blastocyst formation during preimplantation embryo development in vitro. Fertil. Steril. 2014, 102, 1777–1784. [Google Scholar] [CrossRef] [PubMed]
- Lian, I.A.; Loset, M.; Mundal, S.B.; Fenstad, M.H.; Johnson, M.P.; Eide, I.P.; Bjorge, L.; Freed, K.A.; Moses, E.K.; Austgulen, R. Increased endoplasmic reticulum stress in decidual tissue from pregnancies complicated by fetal growth restriction with and without pre-eclampsia. Placenta 2011, 32, 823–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.X.; He, W.H.; Yin, L.J.; Lv, P.P.; Zhang, Y.; Sheng, J.Z.; Leung, P.C.; Huang, H.F. Sustained endoplasmic reticulum stress as a cofactor of oxidative stress in decidual cells from patients with early pregnancy loss. J. Clin. Endocrinol. Metab. 2011, 96, E493–E497. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Mao, C.; Lee, B.; Lee, A.S. GRP78/BiP is required for cell proliferation and protecting the inner cell mass from apoptosis during early mouse embryonic development. Mol. Cell. Biol. 2006, 26, 5688–5697. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Scheuner, D.; Chen, J.J.; Kaufman, R.J.; Ron, D. Ppp1r15 gene knockout reveals an essential role for translation initiation factor 2 alpha (eIF2alpha) dephosphorylation in mammalian development. Proc. Natl. Acad. Sci. USA 2009, 106, 1832–1837. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, Y.K.; Lee, A.S. Expression of the glucose-regulated proteins (GRP94 and GRP78) in differentiated and undifferentiated mouse embryonic cells and the use of the GRP78 promoter as an expression system in embryonic cells. Differentiation 1990, 42, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Liu, H.X.; Zhong-Shu, L.; Dong-Xue, M.; Xu, L.; Shah, S.Z.A.; Ullah, O.; Nan-Zhu, F. Reduced glutathione alleviates tunicamycin-induced endoplasmic reticulum stress in mouse preimplantation embryos. J. Reprod. Dev. 2018, 64, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicks, N.; Bohrer, R.C.; Gutierrez, K.; Michalak, M.; Agellon, L.B.; Bordignon, V. Relief of endoplasmic reticulum stress enhances DNA damage repair and improves development of pre-implantation embryos. PLoS ONE 2017, 12, e0187717. [Google Scholar] [CrossRef] [PubMed]
- Song, B.S.; Kim, J.S.; Yoon, S.B.; Lee, K.S.; Koo, D.B.; Lee, D.S.; Choo, Y.K.; Huh, J.W.; Lee, S.R.; Kim, S.U.; et al. Inactivated Sendai-virus-mediated fusion improves early development of cloned bovine embryos by avoiding endoplasmic-reticulum-stress-associated apoptosis. Reprod. Fertil. Dev. 2011, 23, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Liu, X.J.; Li, J.T.; Zhang, L.; Fu, Y.; Zhang, Y.J.; Chen, R.X.; Wei, X.Q.; Wang, R.; Wang, Y.; et al. Endoplasmic reticulum stress inhibition is a valid therapeutic strategy in vitrifying oocytes. Cryobiology 2015, 70, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qu, P.; Ma, X.; Qiao, F.; Ma, Y.; Qing, S.; Zhang, Y.; Wang, Y.; Cui, W. Tauroursodeoxycholic acid (TUDCA) alleviates endoplasmic reticulum stress of nuclear donor cells under serum starvation. PLoS ONE 2018, 13, e0196785. [Google Scholar] [CrossRef] [PubMed]
- Barrera, N.; Dos Santos Neto, P.C.; Cuadro, F.; Bosolasco, D.; Mulet, A.P.; Crispo, M.; Menchaca, A. Impact of delipidated estrous sheep serum supplementation on in vitro maturation, cryotolerance and endoplasmic reticulum stress gene expression of sheep oocytes. PLoS ONE 2018, 13, e0198742. [Google Scholar] [CrossRef] [PubMed]
- Latham, K.E. Endoplasmic reticulum stress signaling in mammalian oocytes and embryos: Life in balance. Int. Rev. Cell Mol. Biol. 2015, 316, 227–265. [Google Scholar] [PubMed]
- Lane, M.; Gardner, D.K. Understanding cellular disruptions during early embryo development that perturb viability and fetal development. Reprod. Fertil. Dev. 2005, 17, 371–378. [Google Scholar] [CrossRef]
- Anifandis, G.; Messini, C.I.; Dafopoulos, K.; Messinis, I.E. Genes and Conditions Controlling Mammalian Pre- and Post-implantation Embryo Development. Curr. Genom. 2015, 16, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.T.; Koo, O.J.; Kwon, D.K.; Park, H.J.; Jang, G.; Kang, S.K.; Lee, B.C. Effects of melatonin on in vitro maturation of porcine oocyte and expression of melatonin receptor RNA in cumulus and granulosa cells. J. Pineal Res. 2009, 46, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Nasr-Esfahani, M.H.; Aitken, J.R.; Johnson, M.H. Hydrogen peroxide levels in mouse oocytes and early cleavage stage embryos developed in vitro or in vivo. Development 1990, 109, 501–507. [Google Scholar]
- Takahashi, M.; Keicho, K.; Takahashi, H.; Ogawa, H.; Schultz, R.M.; Okano, A. Effect of oxidative stress on development and DNA damage in in-vitro cultured bovine embryos by comet assay. Theriogenology 2000, 54, 137–145. [Google Scholar] [CrossRef]
- Simopoulou, M.; Sfakianoudis, K.; Rapani, A.; Giannelou, P.; Anifandis, G.; Bolaris, S.; Pantou, A.; Lambropoulou, M.; Pappas, A.; Deligeoroglou, E.; et al. Considerations Regarding Embryo Culture Conditions: From Media to Epigenetics. In Vivo 2018, 32, 451–460. [Google Scholar] [Green Version]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef]
- Xu, X.; Liu, T.; Zhang, A.; Huo, X.; Luo, Q.; Chen, Z.; Yu, L.; Li, Q.; Liu, L.; Lun, Z.R.; et al. Reactive oxygen species-triggered trophoblast apoptosis is initiated by endoplasmic reticulum stress via activation of caspase-12, CHOP, and the JNK pathway in Toxoplasma gondii infection in mice. Infect. Immun. 2012, 80, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Landau, G.; Kodali, V.K.; Malhotra, J.D.; Kaufman, R.J. Detection of oxidative damage in response to protein misfolding in the endoplasmic reticulum. Methods Enzymol. 2013, 526, 231–250. [Google Scholar] [PubMed]
- Dorner, A.J.; Wasley, L.C.; Raney, P.; Haugejorden, S.; Green, M.; Kaufman, R.J. The stress response in Chinese hamster ovary cells. Regulation of ERp72 and protein disulfide isomerase expression and secretion. J. Biol. Chem. 1990, 265, 22029–22034. [Google Scholar] [PubMed]
- Zhang, J.Y.; Lee, K.S.; Kim, J.S.; Song, B.S.; Jin, D.I.; Koo, D.B.; Yu, K. Functional characterization of the ER stress induced X-box-binding protein-1 (Xbp-1) in the porcine system. BMC Mol. Biol. 2011, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Song, B.S.; Lee, K.S.; Kim, D.H.; Kim, S.U.; Choo, Y.K.; Chang, K.T.; Koo, D.B. Tauroursodeoxycholic acid enhances the pre-implantation embryo development by reducing apoptosis in pigs. Reprod. Domest. Anim. 2012, 47, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, M.; Sparman, M.; Sritanaudomchai, H.; Ma, H.; Clepper, L.; Woodward, J.; Li, Y.; Ramsey, C.; Kolotushkina, O.; Mitalipov, S. Mitochondrial gene replacement in primate offspring and embryonic stem cells. Nature 2009, 461, 367–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.; Diao, Y.F.; Kang, J.W.; Lee, J.E.; Kim, D.K.; Jin, D.I. Tauroursodeoxycholic acid improves the implantation and live-birth rates of mouse embryos. Reprod. Biol. 2015, 15, 101–105. [Google Scholar] [CrossRef]
- Lin, T.; Zhang, J.Y.; Diao, Y.F.; Kang, J.W.; Jin, D.I. Effects of sorbitol on porcine oocyte maturation and embryo development in vitro. Zygote 2015, 23, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Im, G.S.; Yang, B.S.; Lai, L.; Liu, Z.; Hao, Y.; Prather, R.S. Fragmentation and development of preimplantation porcine embryos derived by parthenogenetic activation and nuclear transfer. Mol. Reprod. Dev. 2005, 71, 159–165. [Google Scholar] [CrossRef]
- Lin, T.; Diao, Y.F.; Choi, H.S.; Oqani, R.K.; Kang, J.W.; Lee, J.E.; Jin, D.I. Procedure used for denuding pig oocytes influences oocyte damage, and development of in vitro and nuclear transfer embryos. Anim. Reprod. Sci. 2015, 152, 65–76. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, F.; Puscheck, E.E.; Rappolee, D.A. Pipetting causes shear stress and elevation of phosphorylated stress-activated protein kinase/jun kinase in preimplantation embryos. Mol. Reprod. Dev. 2007, 74, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Van der Kallen, C.J.; van Greevenbroek, M.M.; Stehouwer, C.D.; Schalkwijk, C.G. Endoplasmic reticulum stress-induced apoptosis in the development of diabetes: Is there a role for adipose tissue and liver? Apoptosis 2009, 14, 1424–1434. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Yu, Y.; Qiao, J. Dual role for the unfolded protein response in the ovary: Adaption and apoptosis. Protein Cell 2017, 8, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.J. Signal transduction by the JNK group of MAP kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef]
- Szegezdi, E.; Fitzgerald, U.; Samali, A. Caspase-12 and ER-stress-mediated apoptosis—The story so far. Ann. N. Y. Acad. Sci. 2003, 1010, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Yuan, J. Cross-talk between two cysteine protease families. Activation of caspase-12 by calpain in apoptosis. J. Cell Biol. 2000, 150, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.V.; Hermel, E.; Castro-Obregon, S.; del Rio, G.; Ellerby, L.M.; Ellerby, H.M.; Bredesen, D.E. Coupling endoplasmic reticulum stress to the cell death program. Mechanism of caspase activation. J. Biol. Chem. 2001, 276, 33869–33874. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Lee, J.E.; Oqani, R.K.; Kim, S.Y.; Cho, E.S.; Jeong, Y.D.; Baek, J.J.; Jin, D.I. Delayed blastocyst formation or an extra day culture increases apoptosis in pig blastocysts. Anim. Reprod. Sci. 2017, 185, 128–139. [Google Scholar] [CrossRef]
- Hao, Y.H.; Lai, L.X.; Mao, J.D.; Im, G.S.; Bonk, A.; Prather, R.S. Apoptosis and in vitro development of preimplantation porcine embryos derived in vitro or by nuclear transfer. Biol. Reprod. 2003, 69, 501–507. [Google Scholar] [CrossRef]
- Kim, E.; Hyun, S.H. Apoptosis in Porcine Pluripotent Cells: From ICM to iPSCs. Int. J. Mol. Sci. 2016, 17, 1533. [Google Scholar] [CrossRef]
- Miller, S.D.; Greene, C.M.; McLean, C.; Lawless, M.W.; Taggart, C.C.; O’Neill, S.J.; McElvaney, N.G. Tauroursodeoxycholic acid inhibits apoptosis induced by Z alpha-1 antitrypsin via inhibition of Bad. Hepatology 2007, 46, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Khaoustov, V.I.; Chung, C.C.; Sohn, J.; Krishnan, B.; Lewis, D.E.; Yoffe, B. Effect of tauroursodeoxycholic acid on endoplasmic reticulum stress-induced caspase-12 activation. Hepatology 2002, 36, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Seyhun, E.; Malo, A.; Schafer, C.; Moskaluk, C.A.; Hoffmann, R.T.; Goke, B.; Kubisch, C.H. Tauroursodeoxycholic acid reduces endoplasmic reticulum stress, acinar cell damage, and systemic inflammation in acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G773–G782. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Melatonin and the pineal gland: Influence on mammalian seasonal and circadian physiology. Rev. Reprod. 1998, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Barrett, P.; Bolborea, M. Molecular pathways involved in seasonal body weight and reproductive responses governed by melatonin. J. Pineal Res. 2012, 52, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, R.A.; Reiter, R.J. Pineal gland: Influence on gonads of male hamsters. Science 1965, 148, 1609–1611. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y. Melatonin as a free radical scavenger in the ovarian follicle. Endocr. J. 2013, 60, 1–13. [Google Scholar] [CrossRef]
- Zarazaga, L.A.; Celi, I.; Guzman, J.L.; Malpaux, B. Melatonin concentrations in the two jugular veins, and relationship with the seasonal reproductive activity in goats. Theriogenology 2010, 74, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Hardeland, R. Antioxidative protection by melatonin: Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005, 27, 119–130. [Google Scholar] [CrossRef]
- Cruz, M.H.; Leal, C.L.; da Cruz, J.F.; Tan, D.X.; Reiter, R.J. Role of melatonin on production and preservation of gametes and embryos: A brief review. Anim. Reprod. Sci. 2014, 145, 150–160. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; He, C.; Zhu, K.; Xu, Z.; Ma, T.; Tao, J.; Liu, G. Melatonin protects porcine oocyte in vitro maturation from heat stress. J. Pineal Res. 2015, 59, 365–375. [Google Scholar] [CrossRef]
- Zhao, X.M.; Hao, H.S.; Du, W.H.; Zhao, S.J.; Wang, H.Y.; Wang, N.; Wang, D.; Liu, Y.; Qin, T.; Zhu, H.B. Melatonin inhibits apoptosis and improves the developmental potential of vitrified bovine oocytes. J. Pineal Res. 2016, 60, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Lee, J.E.; Kang, J.W.; Kim, S.Y.; Jin, D.I. The influence and role of melatonin on in vitro oocyte maturation and embryonic development in pig and cattle. Korean J. Agric. Sci. 2017, 44, 309–317. [Google Scholar]
- Lin, T.; Lee, J.E.; Kang, J.W.; Oqani, R.K.; Cho, E.S.; Kim, S.B.; Il Jin, D. Melatonin supplementation during prolonged in vitro maturation improves the quality and development of poor-quality porcine oocytes via anti-oxidative and anti-apoptotic effects. Mol. Reprod. Dev. 2018, 85, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Sun, G.; Ma, T.; Zhong, F.; Lei, Y.; Li, X.; Wei, W. Melatonin reverses tunicamycin-induced endoplasmic reticulum stress in human hepatocellular carcinoma cells and improves cytotoxic response to doxorubicin by increasing CHOP and decreasing survivin. J. Pineal Res. 2013, 55, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Quan, X.J.; Wang, J.Y.; Liang, C.L.; Zheng, H.D.; Zhang, L. Melatonin inhibits tunicamycin-induced endoplasmic reticulum stress and insulin resistance in skeletal muscle cells. Biochem. Biophys. Res. Commun. 2015, 463, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Boyce, M.; Bryant, K.F.; Jousse, C.; Long, K.; Harding, H.P.; Scheuner, D.; Kaufman, R.J.; Ma, D.; Coen, D.M.; Ron, D.; et al. A selective inhibitor of eIF2alpha dephosphorylation protects cells from ER stress. Science 2005, 307, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Bavister, B. The role of animal studies in supporting human assisted reproductive technology. Reprod. Fertil. Dev. 2004, 16, 719–728. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Stage (Derivation) | ER Stress Markers | Refs | |
|---|---|---|---|---|
| Genes | Proteins | |||
| Mouse | Oocytes | ATF4, ATF6 and BiP/GRP78 | [30] | |
| Mouse | Oocytes | XBP1, ATF4, ATF6 and HSPA5 | [29] | |
| Mouse | Oocytes (vitrified) | XBP1 | [40] | |
| Mouse | Oocytes/blastocysts | XBP1-u and XBP1-s | XBP1 | [16] |
| Mouse | Blastocysts | ATF4, ATF6, BiP, CHOP and PERK | [37] | |
| Mouse | Blastocysts | XBP1, BIP, IRE1, ASK1, ATF4, ATF6, CHOP and PERK | [26] | |
| Mouse | Blastocysts | XBP1 | BiP | [31] |
| Pig | Oocytes | BiP, ATF4, XBP1-u, XBP1-s and CHOP | ATF4, ATF6, BiP/GRP78, P90ATF6, P50ATF6 and CHOP | [3] |
| Pig | Oocytes and 1-cell to blastocyst-stage embryos (PA) | XBP1-u and XBP1-s | XBP1 | [4] |
| Pig | Embryos (day 5)/blastocysts (PA) | XBP1 and GRP78 | [38] | |
| Pig | Blastocysts (SCNT) | XBP1-u and XBP1-s | XBP1 | [13] |
| Bovine | Blastocysts (IVF) | XBP1-u, XBP1-s, IRE1, ATF4, ATF6 and CHOP | [12] | |
| Bovine | Blastocysts (IVF and SCNT) | XBP1-u, XBP1-s and CHOP | [15] | |
| Bovine | Blastocysts (IVF and SCNT) | XBP1, BiP/GRP78, EDEM, ATF4, ATF6, CHOP and IRE1 | [39] | |
| Bovine | Blastocysts (SCNT) | BiP, CHOP and IRE1 | [41] | |
| Buffalo | Blastocysts (IVF) | GRP78 and GRP94 | [14] | |
| Sheep | Oocytes | ATF4, ATF6, GRP78 and CHOP | [42] | |
| Inducer | Treatment | Species | Results | Refs |
|---|---|---|---|---|
| TM | 0.1–1 μM during IVC | Pig | Reduces porcine SCNT-derived blastocyst formation rate, decreases total cell and ICM cell numbers, increases XBP1-s mRNA expression and apoptosis | [13] |
| TM | 0.5 μM during IVC | Mouse | Reduces blastocyst formation rate, increases ROS and apoptosis | [37] |
| TM | 0.5 μM during IVC | Mouse | Prevents blastocyst formation, increases the levels of cleaved caspase 3 protein and the mRNA for XBP1-s | [31] |
| TM | 1 μM in freezing medium | Mouse | Increases XBP1 and caspase 12 protein expression levels | [40] |
| TM | 1–2 μM during IVC | Buffalo | Decreases blastocyst formation rate and cell numbers, increases apoptosis | [14] |
| TM | 2 μM during IVC | Pig | Induces active XBP1 in nuclei of 4-cell stage PA-derived embryos, reduces cleavage rate, blastocyst formation rate, and cell numbers in blastocysts | [4] |
| TM | 1–5 μM during IVC | Pig | Reduces porcine IVF-derived embryo development by reducing the rates of cleavage and blastocyst formation | [55] |
| TM | 1–5 μM during IVM | Pig | Reduces oocyte maturation in pig by increasing the expression levels of BiP, ATF4 and ATF6 | [3] |
| TM | 1–10 μM during IVC | Mouse | Reduces blastocyst formation rate, increases apoptotic index | [16] |
| Sorbitol | 10–75 mM during IVC | Mouse | Reduces blastocyst formation rate, increases apoptotic index | [16] |
| Oxidative stress | Embryos cultured under 20% O2 | Bovine | Reduces bovine IVF-derived blastocyst formation rate; increases XBP1-s, ATF4, ATF6, CHOP and IRE1 gene expression levels and the apoptotic index | [12] |
| Shear stress | Embryo collection | Mouse | Triggers transient activation of the XBP1 arm of ER stress | [26] |
| Electro-fusion | SCNT embryos produced by electrofusion | Bovine | Reduces SCNT-derived embryonic development by increasing XBP1-s, CHOP, GRP78 and EDEM gene expression levels and the apoptotic index, while reducing the blastocyst formation rate and the total and ICM cell numbers | [39] |
| Inhibitor | Treatment * | Species | Results | Refs |
|---|---|---|---|---|
| TUDCA | 50 μM during IVC | Mouse | Improves embryo development and increases the implantation and live birth rates of transferred mouse embryos | [57] |
| TUDCA | 50 μM during IVC | Mouse | Improves the blastocyst formation rate and reduces apoptosis | [16] |
| TUDCA | 50 μM in the freezing medium | Mouse | Enhances the viability and embryonic developmental capacity of vitrified-warmed mouse oocytes by reducing ER stress | [40] |
| TUDCA | 50 μM during IVC | Bovine | Reduces ER stress and ROS levels; improves bovine embryo development | [12] |
| TUDCA | 50 μM during IVC | Bovine | Improves bovine SCNT embryo development by increasing cell numbers and reducing ER stress and apoptosis | [39] |
| TUDCA | 50 μM during IVC | Buffalo | Attenuates apoptosis and ER stress in buffalo IVF-derived embryos | [14] |
| TUDCA | 50 μM during IVM, 50 μM during IVC | Pig | Enhances porcine oocyte maturation and PA-derived embryo developmental potential by preventing ER stress | [4] |
| TUDCA | 50 μM during IVC | Pig | Enhances DNA damage repair and improves porcine preimplantation embryo development by reducing ER stress | [38] |
| TUDCA | 100 μM during IVC | Pig | Improves porcine SCNT embryonic development by attenuating ER stress and reducing apoptosis | [57] |
| TUDCA | 100 μM (treatment of donor cells) | bovine | Improves the development of bovine SCNT-derived embryos by reducing ER stress | [41] |
| TUDCA | 200 μM during IVC | Pig | Improves the development of porcine IVF-derived embryos by modulating ER stress-induced apoptosis | [55] |
| TUDCA | 200 μM during IVM | Pig | Improves the quality and maturation of porcine oocytes | [3] |
| Melatonin | 0.1 μM during IVM | Pig | Improves cumulus cell expansion and oocyte maturation by combating ER stress | [3] |
| Valproic acid | 3 mM during IVC | Bovine | Improves the development of bovine SCNT-derived embryos by reducing ER stress and apoptosis | [15] |
| GSH | 1 mM during IVC | Mouse | Increases the mouse blastocyst formation rate and alleviates ER stress | [37] |
| GSH | 1 mM during IVC | Bovine | Reduces ROS levels and increases the blastocyst formation rate | [12] |
| Salubrinal | 100 nM during IVM | Mouse | Improves pentraxin-3 secretion, mitochondrial membrane potential, and embryonic development by reducing ER stress | [29] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, T.; Lee, J.E.; Kang, J.W.; Shin, H.Y.; Lee, J.B.; Jin, D.I. Endoplasmic Reticulum (ER) Stress and Unfolded Protein Response (UPR) in Mammalian Oocyte Maturation and Preimplantation Embryo Development. Int. J. Mol. Sci. 2019, 20, 409. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020409
Lin T, Lee JE, Kang JW, Shin HY, Lee JB, Jin DI. Endoplasmic Reticulum (ER) Stress and Unfolded Protein Response (UPR) in Mammalian Oocyte Maturation and Preimplantation Embryo Development. International Journal of Molecular Sciences. 2019; 20(2):409. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020409
Chicago/Turabian StyleLin, Tao, Jae Eun Lee, Jung Won Kang, Hyeon Yeong Shin, Ju Bin Lee, and Dong Il Jin. 2019. "Endoplasmic Reticulum (ER) Stress and Unfolded Protein Response (UPR) in Mammalian Oocyte Maturation and Preimplantation Embryo Development" International Journal of Molecular Sciences 20, no. 2: 409. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020409