Comparative Transcriptome Analysis of the Pinewood Nematode Bursaphelenchus xylophilus Reveals the Molecular Mechanism Underlying Its Defense Response to Host-Derived α-pinene

 ,
,
Abstract
:
1. Introduction
2. Results
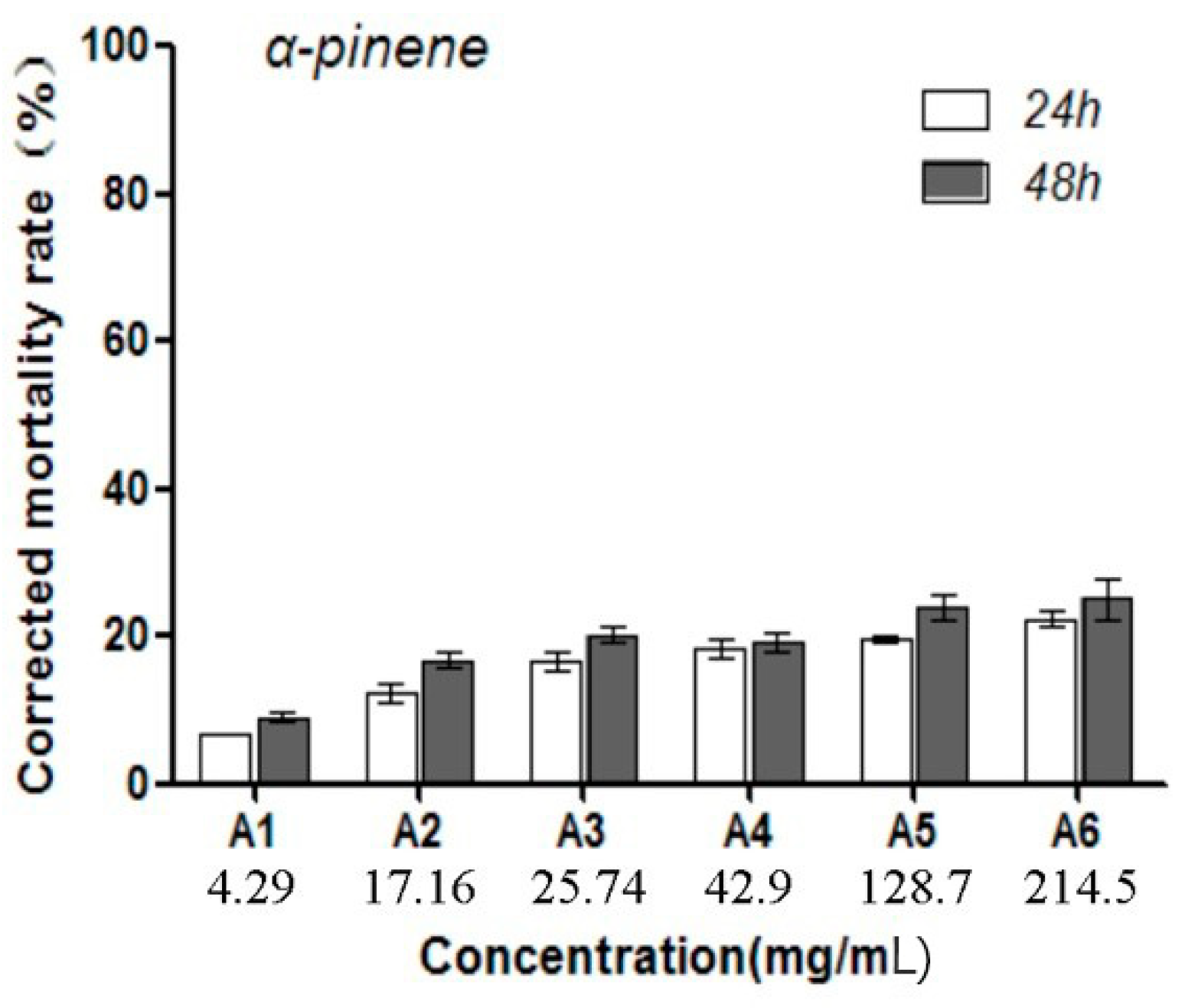
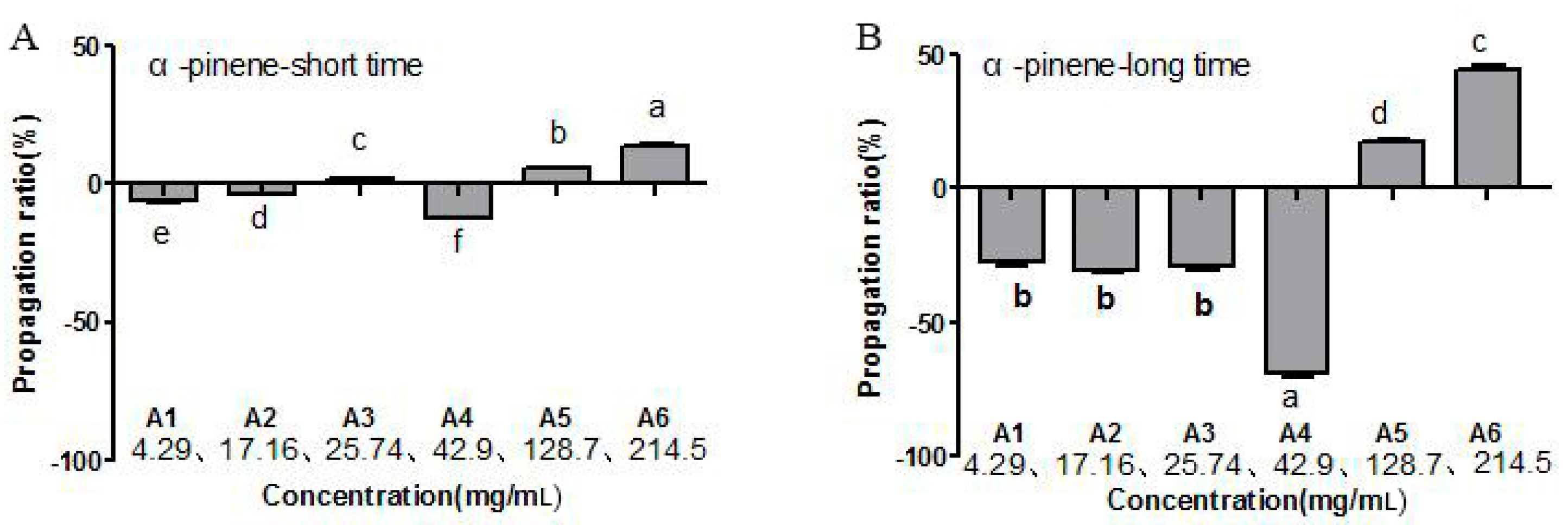
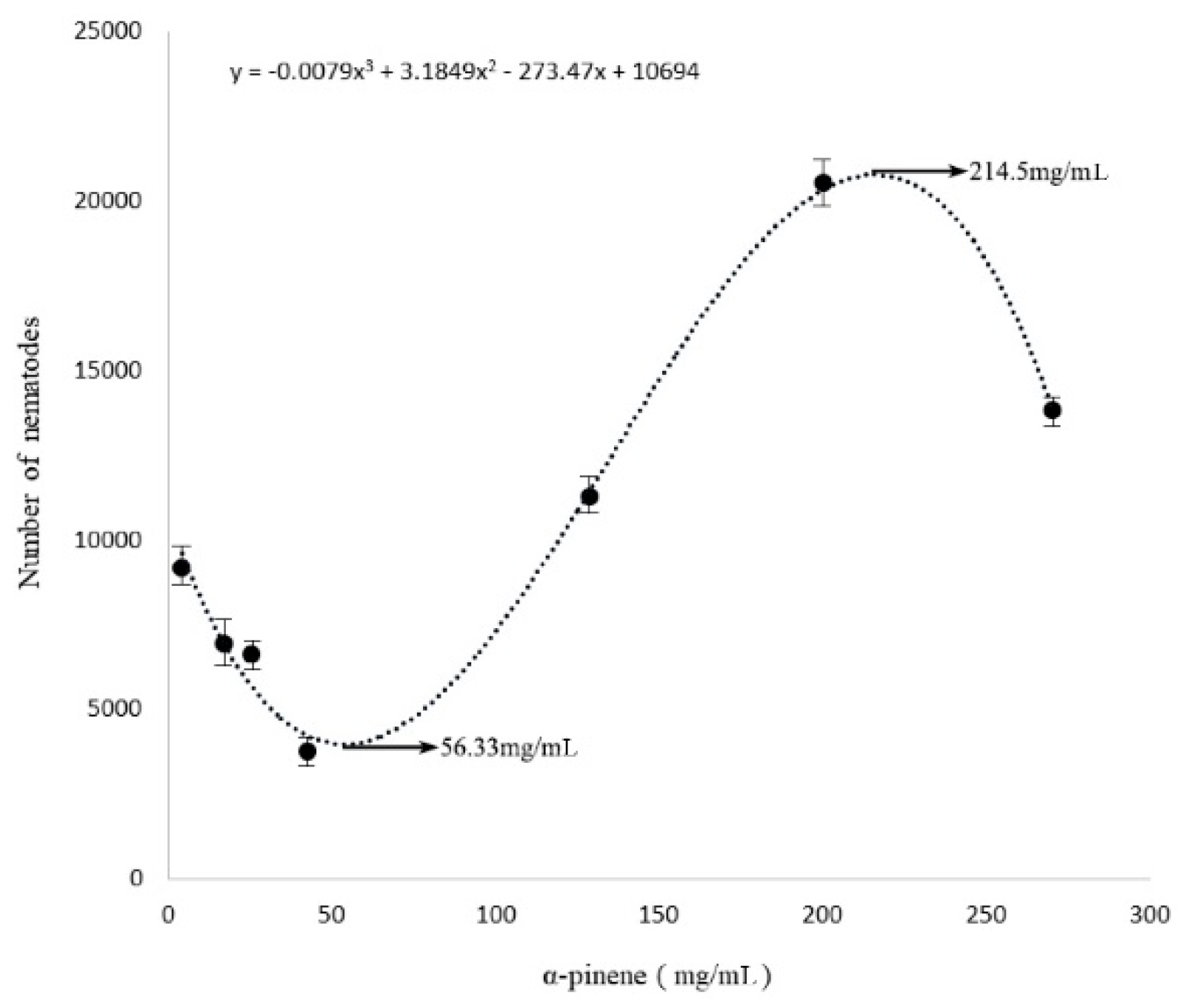
2.1. Toxic Effects by α-Pinene on B. xylophilus Viability and Reproduction
2.2. Transcriptome Sequencing and Data Assembly
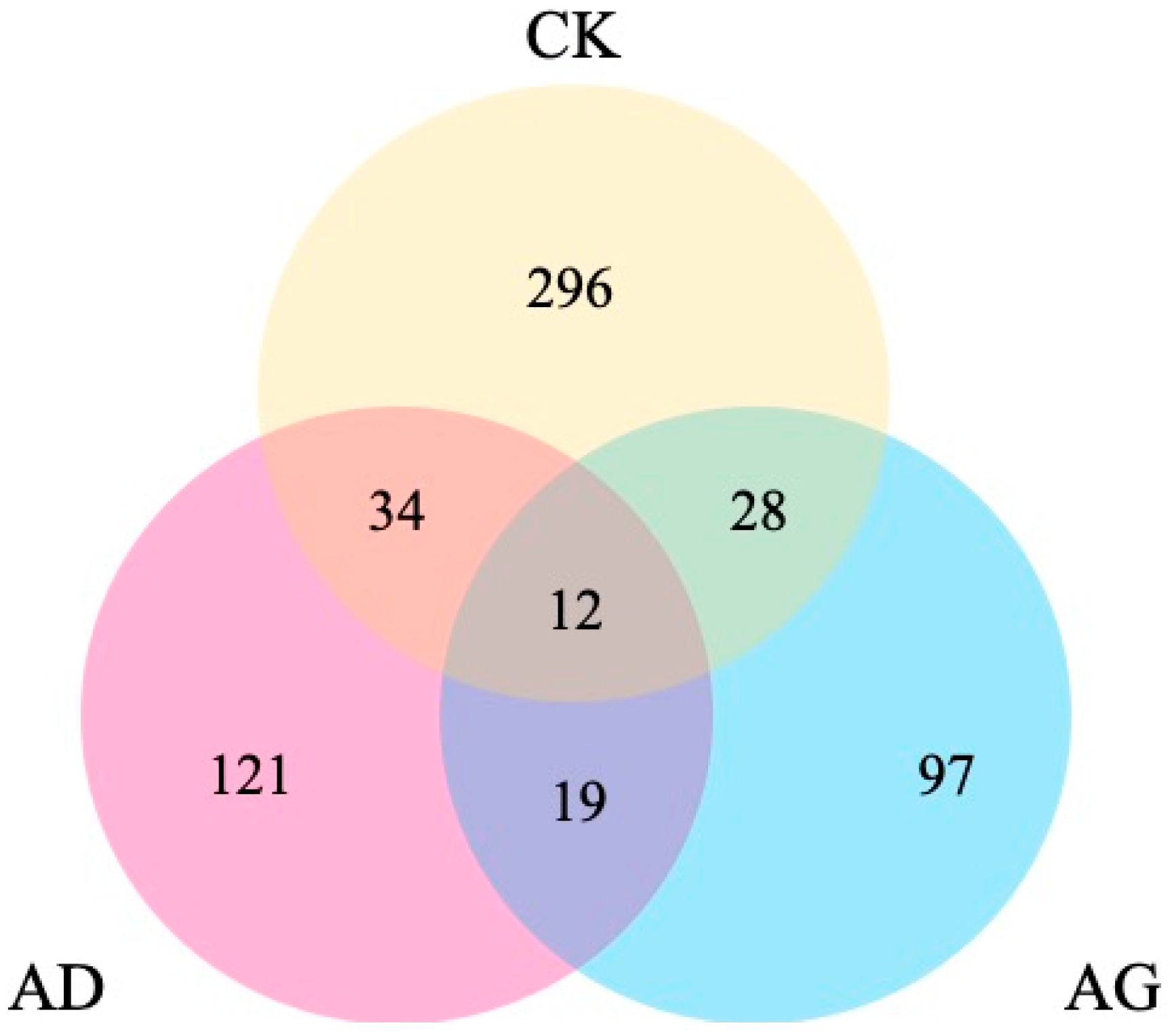
2.3. Differentially Expressed Genes (DEGs) Found Among the Transcriptomes
2.4. Functional Annotation of the DEGs
2.4.1. DEGs Involved in Detoxification
2.4.2. DEGs Involved in Transport
2.4.3. DEGs Encoding Receptors
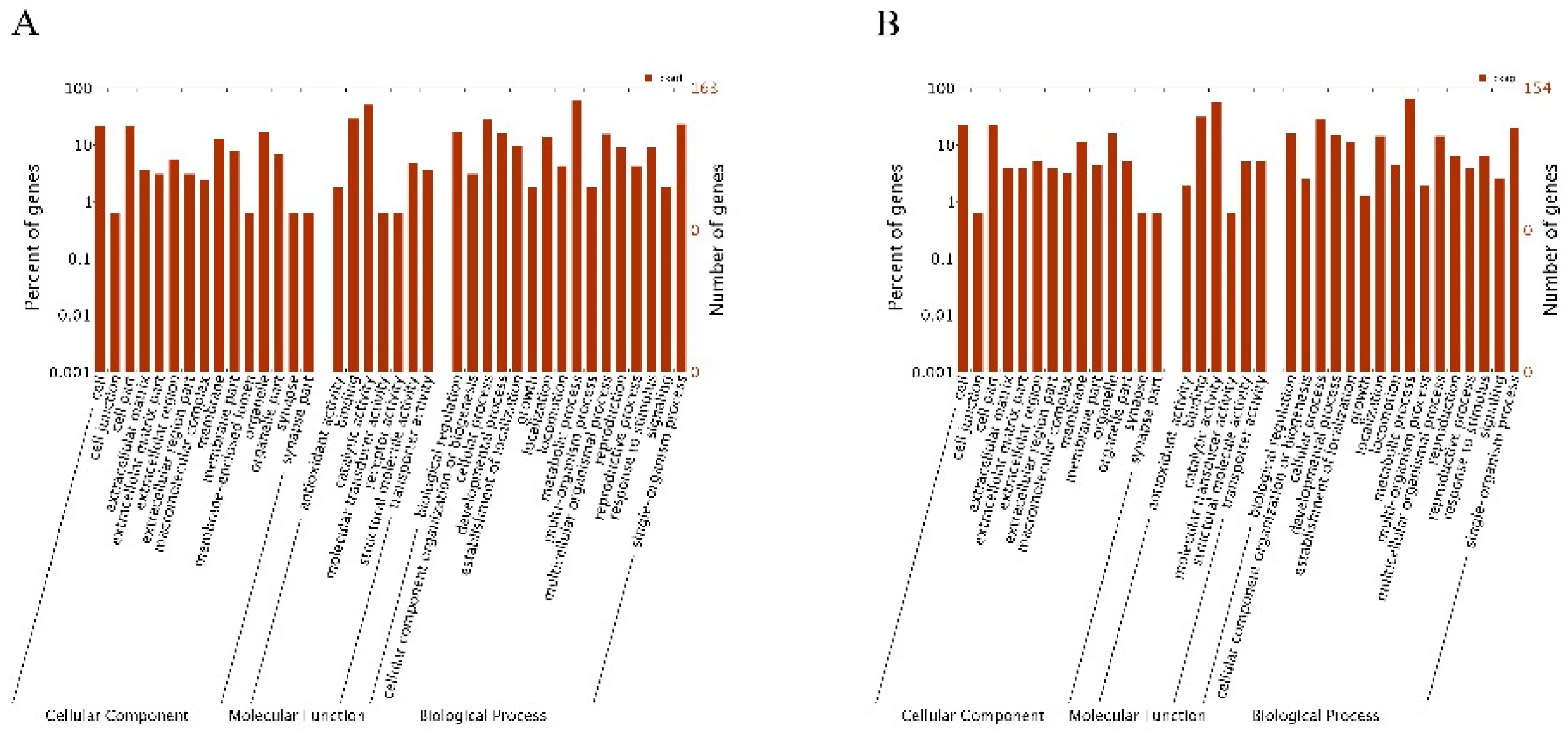
2.5. GO Enrichment Analysis of the DEGs
2.6. KEGG Functional Annotation of the DEGs
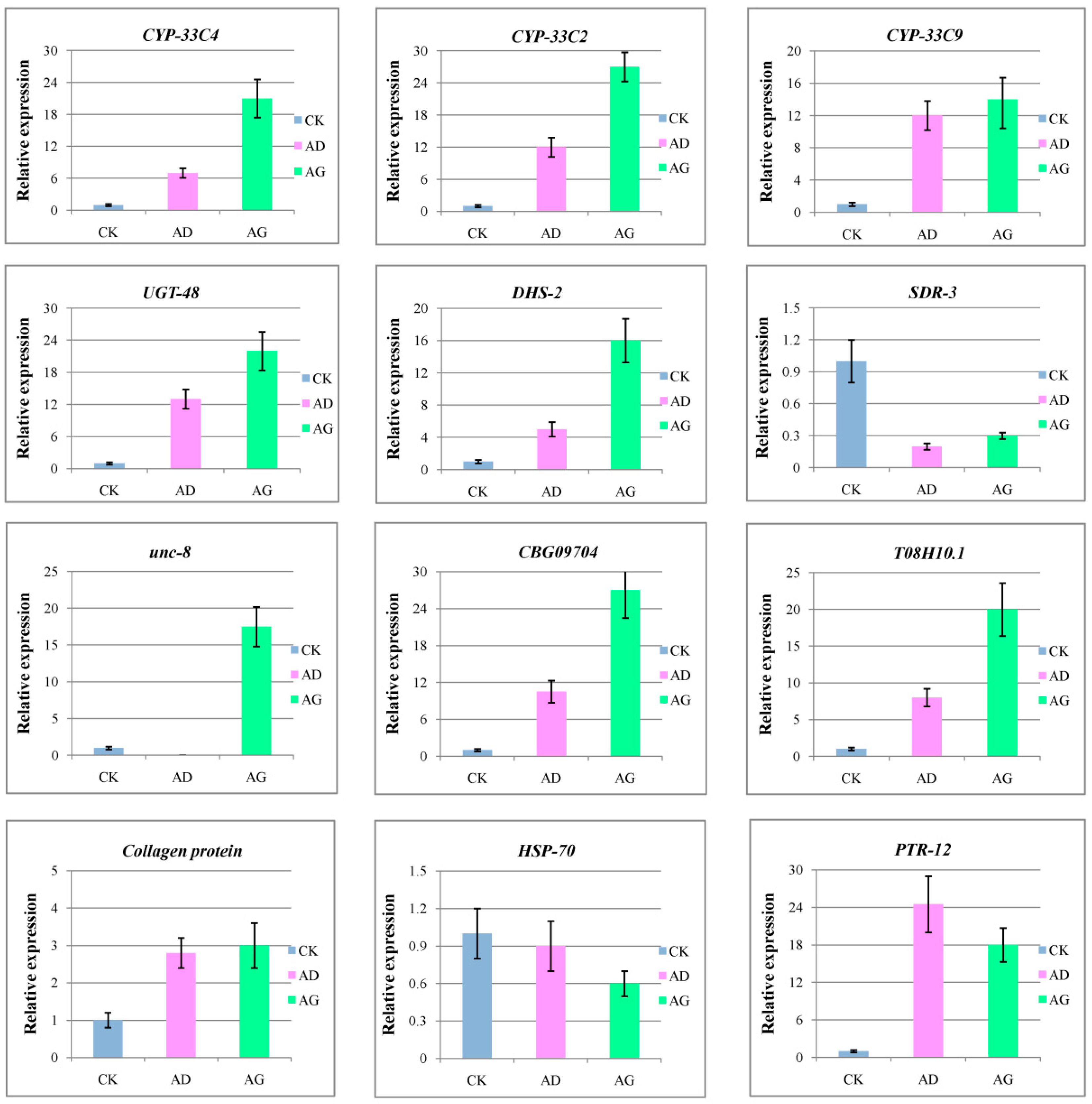
2.7. Verification of Gene Expression by qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Toxic Effects of Different α-Pinene Concentrations on the Mortality and Reproduction Rates of B. xylophilus
4.3. Preparation of PWNs Using Different Concentrations of α-Pinene for Transcriptome Analyses
4.4. Total RNA Isolation and cDNA Synthesis
4.5. Sequencing and Assemblage of B. xylophilus Transcriptome DEGs in Response to α-Pinene
4.6. Identification of Statistically Enriched GO Terms and KEGG Pathways
4.7. Quantitative RT-PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PWD | Pine wilt disease |
| PWN | Pine wood nematode |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
References
- Nickle, W.R.; Golden, A.M.; Mamiya, Y.; Wergin, W.P. On the taxonomy and morphology of the pine woodnematode, Bursaphelenchus xylophilus (Steiner &Buhrer 1934) Nickle 1970. J. Nematol. 1981, 13, 385–393. [Google Scholar] [PubMed]
- Branco, M.; Bragança, H.; Sousa, E.; Phillips, A.J. Pests and Diseases in Portuguese Forestry: Current and New Threats; Springer: New York, NY, USA, 2014. [Google Scholar]
- Mota, M.; Oliveira, S.; Vicente, C.; Vieira, P. Pine wilt disease in portugal: Recent progress and new approaches to the understanding and control of the pine wood nematode and its insect vector. J. Nematol. 2009, 41, 361. [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus xylophilus, causal agent of pine wilt disease on Pinus pinaster in northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.; Espada, M.; Vieira, P.; Mota, M. Pine Wilt Disease: A threat to European forestry. Eur. J. Plant Pathol. 2012, 133, 89–99. [Google Scholar] [CrossRef]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar] [PubMed]
- Togashi, K.; Shigesada, N. Spread of the pinewood nematode vectored by the Japanese pine sawyer: Modeling and analytical approaches. Popul. Ecol. 2006, 48, 271–283. [Google Scholar] [CrossRef]
- Yoshimura, A.; Kawasaki, K.; Takasu, F.; Togashi, K.; Futai, K.; Shigesada, N. Modeling the spread of wilt disease caused by nematodes with pine sawyers as vector. Ecology 1999, 80, 1691–1702. [Google Scholar] [CrossRef]
- Sun, Y.C. Pine wood nematode found at Dr. Sun Yat’s Mausoleum in Nanjing. Jiangsu For. Sci. Technol. 1982, 4, 47. [Google Scholar]
- Kiyohara, T.; Tokushige, Y. Inoculation experiments of a nematode, Bursaphelenchus sp., onto pine trees. J. Jpn. For. Soc. 2008, 53, 210–218. [Google Scholar]
- Mamiya, Y.; Enda, N. Transmission of Bursaphelenchus Lignicolus (Nematoda: Aphelenchoididae) by Monochamus Alternatus (Coleoptera: Cerambycidae). Nematologica 1972, 18, 159–162. [Google Scholar] [CrossRef]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Kai, M.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar]
- Morimoto, K.; Iwasaki, A. Role of Monochamus alternatus (Coleoptera: Cerambycidae) as a vector of Bursaphelenchus lignicolus (Nematoda : Aphelenchoididae). J. Jpn. For. Soc. 1972, 54, 177–183. [Google Scholar]
- Ryss, A.; Vieira, P.; Mota, M.; Kulinich, O. A synopsis of the genus Bursaphelenchus Fuchs, 1937 (Aphelenchida: Parasitaphelenchidae) with keys to species. Nematology 2005, 7, 393–458. [Google Scholar]
- Kuroda, K.; Yamada, T.; Mineo, K.; Tamura, H. Effects of cavitation on the development of pine wilt disease caused by Bursaphelenchus xylophilus. Jpn. J. Phytopathol. 1988, 54, 606–615. [Google Scholar] [CrossRef]
- Kuroda, K. Mechanism of cavitation development in the pine wilt disease. For. Pathol. 2010, 21, 82–89. [Google Scholar] [CrossRef]
- Ikeda, T.; Ohtsu, M. Detection of xylem cavitation in field-grown pine trees using the acoustic emission technique. Ecol. Res. 1992, 7, 391–395. [Google Scholar] [CrossRef]
- Kuroda, K.; Kanbara, Y.; Inoue, T.; Ogawa, A. Analysis of NMR-CT images to detect the xylem dysfunction and lesions in tree trunks. In Abstracts Pacific Regional Wood Anatomy Conference; FAO the United Nations Regional Office for Asia Pacific (FAO RAP): Bangkok, Thailand, 2002. [Google Scholar]
- Utsuzawa, S.; Fukuda, K.; Sakaue, D. Use of magnetic resonance microscopy for the nondestructive observation of xylem cavitation caused by pine wilt disease. Phytopathology 2005, 95, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Zulak, K.G.; Bohlmann, J. Terpenoid biosynthesis and specialized vascular cells of conifer defense. J. Intergr. Plant. Biol. 2010, 52, 86–97. [Google Scholar] [CrossRef]
- Lewinsohn, E.; Savage, T.J.; Croteau, M.G.R. Simultaneous analysis of monoterpenes and diterpenoids of conifer oleoresin. Phytochem. Anal. 1993, 4, 220–225. [Google Scholar] [CrossRef]
- Ning, T.; Fan, J.; Fang, Y.; Sun, J. Changes in contents of host volatile terpenes under different damaged states and electroantennogram response of Monochamus alternatus Hope to these volatiles. Acta Entomol. Sin. 2006, 49, 179–188. [Google Scholar]
- Zhen, Z.; Xi, H.U.; Dong, L.I.; Fu, X.U.; Gui, H.U.; Zhen, S.; Xian, L. Study on contents variation of neutral terpenoids of resistant provenance of P. massoniana after inoculating Bursaphelenchus xylophilus. Chem. Ind. For. Prod. 2001, 21, 52–58. [Google Scholar]
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.L.; Wei, W.; Kang, L.; Sun, J.H. Chemotaxis of the pinewood nematode, Bursaphelenchus xylophilus, to volatiles associated with host pine, Pinus massoniana, and its vector Monochamus alternatus. J. Chem. Ecol. 2007, 33, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Zhao, L.; Lu, M.; Zhang, S.; Sun, J. The ratio and concentration of two monoterpenes mediate fecundity of the pinewood nematode and growth of Its associated fungi. PLoS ONE 2013, 7, e31716. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [PubMed]
- Ryoji, S.; Hironobu, M.; Taisei, K.; Yuko, T.; Mitsuyoshi, U.; Kazuyoshi, F. Secretome analysis of the pine wood nematode Bursaphelenchus xylophilus reveals the tangled roots of parasitism and Its potential for molecular mimicry. PLoS ONE 2013, 8, e67377. [Google Scholar]
- Matsye, P.D.; Kumar, R.; Hosseini, P.; Jones, C.M.; Tremblay, A.; Alkharouf, N.W.; Matthews, B.F.; Klink, V.P. Mapping cell fate decisions that occur during soybean defense responses. Plant Mol. Biol. 2011, 77, 513–528. [Google Scholar] [CrossRef] [PubMed]
- Matsye, P.D.; Lawrence, G.W.; Youssef, R.M.; Kim, K.H.; Lawrence, K.S.; Matthews, B.F.; Klink, V.P. The expression of a naturally occurring, truncated allele of an α-SNAP gene suppresses plant parasitic nematode infection. Plant Mol. Biol. 2012, 80, 131–155. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, Z.Y.; Zhang, K.; Lu, Q.; Liang, J.; Zhang, X.Y. Characterization of the Pinus massoniana transcriptional response to Bursaphelenchus xylophilus infection using Suppression Subtractive Hybridization. Int J. Mol. Sci. 2013, 14, 11356–11375. [Google Scholar] [CrossRef]
- Cui, J.; Li, Y.; Zhang, W.; Wang, X.; Pan, L.; Feng, Y.; Zhang, X. The population strucuture and sex ratios of Bursaphelenchus xylophilus under-pinene stress. J. For. Res. 2018. [Google Scholar] [CrossRef]
- Kong, J.O.; Park, I.K.; Choi, K.S.; Shin, S.C.; Ahn, Y.J. Nematicidal and propagation activities of thyme red and white oil compounds toward Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae). J. Nematol. 2007, 39, 237. [Google Scholar] [PubMed]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural products (secondary metabolites). Biochem. Mol. Biol. Plants 2000, 24, 1250–1319. [Google Scholar]
- Langenheim, J.H. Higher plant terpenoids: A phytocentric overview of their ecological roles. J. Chem. Ecol. 1994, 20, 1223–1280. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Kanzaki, N.; Futai, K. Volatile compounds in pine stands suffering from pine wilt disease: Qualitative and quantitative evaluation. Nematology 2006, 8, 869–879. [Google Scholar]
- Yan, X.; Cheng, X.Y.; Wang, Y.S.; Luo, J.; Mao, Z.C.; Ferris, V.R.; Xie, B.Y. Comparative transcriptomics of two pathogenic pinewood nematodes yields insights into parasitic adaptation to life on pine hosts. Gene 2012, 505, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Thiel, L.; Hendricks, F. Study into the Establishment of an Aroma and Fragrance Fine Chemicals Value Chain in South Africa, Part III: Aroma Chemicals Derived from Petrochemical Feedstocks, Tender Number T79/07/03; Government Tender Bulletins; National Economic Development and Labor Council: Pretoria, South Africa, 2004. [Google Scholar]
- Espada, M.; Silva, A.C.; van den Akker, S.E.; Cock, P.J.; Mota, M.; Jones, J.T. Identification and characterization of parasitism genes from the pinewood nematode Bursaphelenchus xylophilus reveals a multilayered detoxification strategy. Mol. Plant Pathol. 2016, 17, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.Y.; Tian, X.L.; Wang, Y.S.; Lin, R.M.; Mao, Z.C.; Chen, N.; Xie, B.Y. Metagenomic analysis of the pinewood nematode microbiome reveals a symbiotic relationship critical for xenobiotics degradation. Sci. Rep. 2013, 3, 1869. [Google Scholar] [CrossRef] [PubMed]
- Shreffler, W.; Magardino, T.; Shekdar, K.; Wolinsky, E. The unc-8 and sup-40 genes regulate ion channel function in Caenorhabditis elegans motorneurons. Genetics 1995, 139, 1261–1272. [Google Scholar]
- Stein, L.D.; Bao, Z.; Blasiar, D.; Blumenthal, T.; Brent, M.R.; Chen, N.; Chinwalla, A.; Clarke, L.; Clee, C.; Coghlan, A. The genome sequence of Caenorhabditis briggsae: A platform for comparative genomics. PLoS Biol. 2003, 1, E45. [Google Scholar] [CrossRef]
- Consortium, C.E.S. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018. [Google Scholar]
- Ingham, P.W.; Nakano, Y.; Seger, C. Mechanisms and functions of Hedgehog signalling across the metazoa. Nat. Rev. Genet. 2011, 12, 393. [Google Scholar] [CrossRef] [PubMed]
- Nüssleinvolhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef]
- Mohler, J. Requirements for hedgehog, a segmental polarity gene, in patterning larval and adult cuticle of Drosophila. Genetics 1988, 120, 1061–1072. [Google Scholar] [PubMed]
- Kang, J.S.; Lee, H.; Moon, I.S.; Lee, Y.; Koh, Y.H.; Je, Y.H.; Lee, S.H. Construction and characterization of subtractive stage-specific expressed sequence tag (EST) libraries of the pinewood nematode Bursaphelenchus xylophilus. Genomics 2009, 94, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Kim, J.; Shin, S.; Park, I. Nematicidal activity of monoterpenoids against the pine wood nematode (Bursaphelenchus xylophilus). Russ. J. Nematol. 2007, 15, 35. [Google Scholar]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293. [Google Scholar] [CrossRef]
- Su, Y.L.; Li, J.M.; Li, M.; Luan, J.B.; Ye, X.D.; Wang, X.W.; Liu, S.S. Transcriptomic analysis of the salivary glands of an invasive whitefly. PLoS ONE 2012, 7, e39303. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Martin, J.; Abubucker, S.; Scott, A.L.; Mccarter, J.P.; Wilson, R.K.; Jasmer, D.P.; Mitreva, M. Intestinal transcriptomes of nematodes: Comparison of the parasites Ascaris suum and Haemonchus contortus with the free-living Caenorhabditis elegans. PLoS Neglect. Trop. Dis. 2008, 2, e269. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, H.; Pei, J.; Dong, Y.; Wang, F.; Chen, H.; Sun, Y.; Wang, N.; Li, H.; Li, Y. De novo sequencing and comparative analysis of the blueberry transcriptome to discover putative genes related to antioxidants. Gene 2012, 511, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Livak, J.K.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Data (bp) | Reads | Q20 (%) | Q30 (%) | Mapped (%) | ExpGene |
|---|---|---|---|---|---|---|
| CK | 2,121,994,600 | 21,219,946 | 96.495 | 89.795 | 96.77 | 14,796 |
| AD | 2,798,113,800 | 27,981,138 | 96.705 | 90.255 | 97.06 | 15,159 |
| AG | 2,267,290,600 | 22,672,906 | 96.650 | 90.095 | 96.94 | 14,998 |
| Condition | ID | Fold Change | p-Value | Annotation |
|---|---|---|---|---|
| CK–AD-up | BUX.s01144.121 | 29.0980 | 2.4758 × 10−13 | CYP-33C4,Oxidoreductase |
| BUX.s00351.319 | 23.1279 | 0.0364 | unc-8,Membrane Ion channel and transport | |
| BUX.s00713.666 | 17.9516 | 5.798 × 10−12 | F21A3.11,Acid phosphatase activity | |
| BUX.s00460.317 | 17.2096 | 3.57851 × 10−6 | ugt-48,Glycosyltransferase | |
| BUX.s01063.115 | 15.4635 | 8.46364 × 10−13 | CYP-33C2,Oxidoreductase | |
| BUX.s00116.700 | 14.3489 | 1.16933 × 10−11 | CYP-33C9,Oxidoreductase | |
| BUX.s01337.92 | 12.1480 | 0.0176 | PTR-12,hedgehog receptor activity | |
| BUX.s00110.26 | 11.6515 | 8.98925 × 10−6 | CBG09704,oxidoreductase activity | |
| BUX.s00460.348 | 11.4609 | 2.44898 × 10−8 | CBG01395,steroid hormone receptor activity | |
| BUX.s00460.315 | 10.2344 | 6.54149 × 10−10 | ugt-48,Glycosyltransferase | |
| CK–AD-down | BUX.s01337.103 | 34.2444 | 0.016867135 | Dimethylaniline monooxygenase |
| BUX.s01147.198 | 33.0521 | 3.56339 × 10−16 | Alcohol dehydrogenase 1 | |
| BUX.s00055.304 | 32.8176 | 0.020311806 | CBR-CUTI-1,Unknown | |
| BUX.s00116.698 | 29.7579 | 7.6292 × 10−16 | CYP-33E2,Oxidoreductase | |
| BUX.s00116.725 | 21.7595 | 1.37374 × 10−8 | Oxidoreductase dhs-27 | |
| BUX.s00508.7 | 21.4028 | 0.011669587 | LMOD1,Cytoskeleton | |
| BUX.c09083.1 | 20.3836 | 1.98965 × 10−5 | GST-1,Peroxidase and Transferase activity | |
| BUX.s01147.18 | 18.8661 | 8.05309 × 10−6 | SCAF14537,small GTPase | |
| BUX.s00609.89 | 17.8356 | 0.026861087 | Chia-prov protein (Fragment) | |
| BUX.s01254.326 | 17.5406 | 2.43072 × 10−12 | Elo-6,transferase activity | |
| CK–AG-up | BUX.s00116.700 | 13.2087 | 3.07567 × 10−20 | CYP-33C9,Oxidoreductase |
| BUX.s01063.115 | 10.4479 | 3.01485 × 10−19 | CYP-33C2,Oxidoreductase | |
| BUX.s01092.190 | 8.7500 | 0.0431 | CBN-SRV-15,Unknown | |
| BUX.s01337.92 | 8.4583 | 0.0493 | PTR-12,hedgehog receptor activity | |
| BUX.s01144.121 | 8.2639 | 3.21387 × 10−7 | CYP-33C4,Oxidoreductase | |
| BUX.s00460.348 | 7.5662 | 7.77552 × 10−9 | CBG01395,steroid hormone receptor activity | |
| BUX.s00460.317 | 7.3889 | 0.0021 | ugt-48,Glycosyltransferase | |
| BUX.s01144.118 | 7.2917 | 1.18745 × 10−6 | CYP-33C1,Oxidoreductase | |
| BUX.s01092.48 | 6.8873 | 6.21794 × 10−11 | CYP-32A1,Oxidoreductase | |
| BUX.s01259.89 | 6.8056 | 0.0040 | GANA-1,Glycosidase and Hydrolase | |
| CK–AG-down | BUX.s01337.103 | 9.1429 | 0.0475 | Dimethylaniline monooxygenase |
| BUX.s01268.31 | 7.1579 | 8.58 × 10−5 | Alcohol dehydrogenase 1 | |
| BUX.s00116.725 | 6.0621 | 2.60 × 10−6 | Oxidoreductase dhs-27 | |
| BUX.s01149.11 | 6.0280 | 1.65 × 10−11 | RHY-1,Transferase activity | |
| BUX.s01281.195 | 5.6961 | 1.53 × 10−9 | Alcohol dehydrogenase 1 | |
| BUX.s01281.74 | 5.4611 | 8.24 × 10−12 | F44E5.4,ATP binding | |
| BUX.s00560.3 | 5.4286 | 0.0034 | TTR-28,extracellular space | |
| BUX.s00364.138 | 5.3184 | 1.22 × 10−10 | F27D9.2,transmembrane transport | |
| BUX.s00961.158 | 4.9979 | 7.15 × 10−11 | HSP-70,ATP binding | |
| BUX.s00364.193 | 4.7950 | 2.06 × 10−10 | Acyl-coenzyme A oxidase; |
| Classification | Genes | Function | Fold Change | |
|---|---|---|---|---|
| CK–AD | CK–AG | |||
| Detoxification | cyp-32a1 | Monooxygenase activity | 9.4349 | 6.8873 |
| cyp-13a8 | Monooxygenase activity | 0.3504 | 0.4425 | |
| cyp-33c1 | Monooxygenase activity | 6.2375 | 7.2917 | |
| cyp-33c2 | Monooxygenase activity | 15.4635 | 10.4479 | |
| cyp-33c4 | Monooxygenase activity | 29.098 | 8.2639 | |
| cyp-33c9 | Monooxygenase activity | 14.3489 | 13.2087 | |
| cyp-33e2 | Monooxygenase activity | 0.0336 | 0.2401 | |
| ugt-47 | UDP-glucuronosyl Transferase | 4.4247 | 2.6057 | |
| ugt-48 | UDP-glucuronosyl Transferase | 17.2096 | 7.3889 | |
| ugt-49 | UDP-glucuronosyl Transferase | 3.2175 | - | |
| sdr-1 | Dehydrogenase/reductase | 5.011 | 3.325 | |
| sdr-4 | Dehydrogenase/reductase | 0.1948 | 0.3949 | |
| dhs-2 | Dehydrogenase/reductase | 7.528 | 3.8671 | |
| dhs-27 | Dehydrogenase/reductase | 4.4334 | 3.1952 | |
| gst-33 | Glutathione S-Transferase | 0.1312 | 0.4045 | |
| gst-39 | Glutathione S-Transferase | 0.1734 | 0.4042 | |
| Transport | unc-8 | Ion channel and transport | 23.1279 | - |
| CBG06849 | Integral to membrane | 4.9059 | 5.0114 | |
| CBG15937 | Integral to membrane | 3.399 | 3.5515 | |
| T09B9.2 | Transmembrane transport | 2.6854 | 3.0404 | |
| TSP-11 | Protein; Transmembrane | 3.6887 | - | |
| TWK-8 | Potassium channel activity | 3.9715 | 2.1194 | |
| ATPase | Sodium/potassium transport | 2.3976 | - | |
| ABC | ABC transporter; | 4.5513 | 2.3998 | |
| F44E7.7 | Transmembrane transport | - | - | |
| K09E9.1 | Transmembrane transport | - | 2.254 | |
| F27D9.2 | Transmembrane transport | 0.1588 | - | |
| unc-49 | Channel activity | 2.2265 | - | |
| T19D12.9 | Transmembrane transport | 0.3451 | - | |
| Y19D10A.11 | Transmembrane transport | 0.2002 | - | |
| Receptor | CBG01395 | Steroid hormone receptor activity | 11.4609 | 7.5662 |
| IFTA-2 | Cell growth and regulatory | 2.2312 | - | |
| Exostosin-2 | Multicellular organismal development | 2.9168 | - | |
| PTR-12 | Hedgehog receptor activity | 12.148 | 8.4583 | |
| PTR-13 | Hedgehog receptor activity | - | 2.1063 | |
| SREBP-1c | Sterol regulatory element-binding | 4.4802 | - | |
| START | START domain protein; cell division | - | - | |
| shr-86 | Steroid hormone receptor activity | 6.7994 | 3.1469 | |
| nhr-19 | Nuclear receptor family member | 2.1711 | - | |
| nhr-3 | Nuclear receptor family member | 2.1206 | - | |
| nhr-40 | Nuclear receptor family member | 2.706 | - | |
| nhr-62 | Nuclear receptor family member | 2.4684 | 2.0637 | |
| nhr-70 | Nuclear receptor family member | 3.1917 | 2.5895 | |
| nhr-10 | Nuclear receptor family member | 2.649 | 2.9513 | |
| nhr-31 | Nuclear receptor family member | - | - | |
| KEGG (KO) Term | NO. Genes | |
|---|---|---|
| CK–AD | CK–AG | |
| Metabolism | ||
| Carbohydrate metabolism | 67 | 90 |
| Energy metabolism | 14 | 18 |
| Lipid metabolism | 77 | 66 |
| Amino acid metabolism | 38 | 68 |
| Glycan biosynthesis and metabolism | 16 | 17 |
| Metabolism of cofactors and vitamins | 42 | 53 |
| Metabolism of terpenoids and polyketides | 3 | 4 |
| Biosynthesis of other secondary metabolites | 11 | 13 |
| Xenobiotics biodegradation and metabolism | 114 | 112 |
| Cellular processes | ||
| Transport and catabolism | 57 | 26 |
| Organismal systems | ||
| Immune system | 8 | |
| Digestive system | 12 | |
| Endocrine system | 14 | 9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Meng, F.; Deng, X.; Wang, X.; Feng, Y.; Zhang, W.; Pan, L.; Zhang, X. Comparative Transcriptome Analysis of the Pinewood Nematode Bursaphelenchus xylophilus Reveals the Molecular Mechanism Underlying Its Defense Response to Host-Derived α-pinene. Int. J. Mol. Sci. 2019, 20, 911. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040911
Li Y, Meng F, Deng X, Wang X, Feng Y, Zhang W, Pan L, Zhang X. Comparative Transcriptome Analysis of the Pinewood Nematode Bursaphelenchus xylophilus Reveals the Molecular Mechanism Underlying Its Defense Response to Host-Derived α-pinene. International Journal of Molecular Sciences. 2019; 20(4):911. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040911
Chicago/Turabian StyleLi, Yongxia, Fanli Meng, Xun Deng, Xuan Wang, Yuqian Feng, Wei Zhang, Long Pan, and Xingyao Zhang. 2019. "Comparative Transcriptome Analysis of the Pinewood Nematode Bursaphelenchus xylophilus Reveals the Molecular Mechanism Underlying Its Defense Response to Host-Derived α-pinene" International Journal of Molecular Sciences 20, no. 4: 911. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040911