Dietary Supplementation with Sea Bass (Lateolabrax maculatus) Ameliorates Ulcerative Colitis and Inflammation in Macrophages through Inhibiting Toll-Like Receptor 4-Linked Pathways

 ,
, 

Abstract
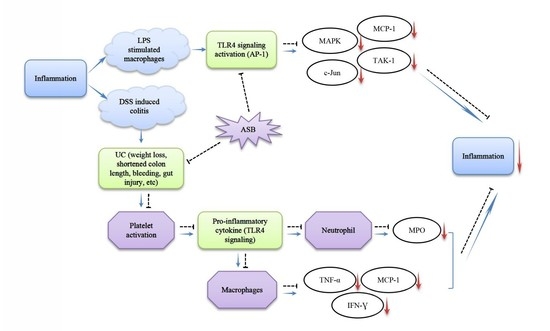
:
1. Introduction
2. Results
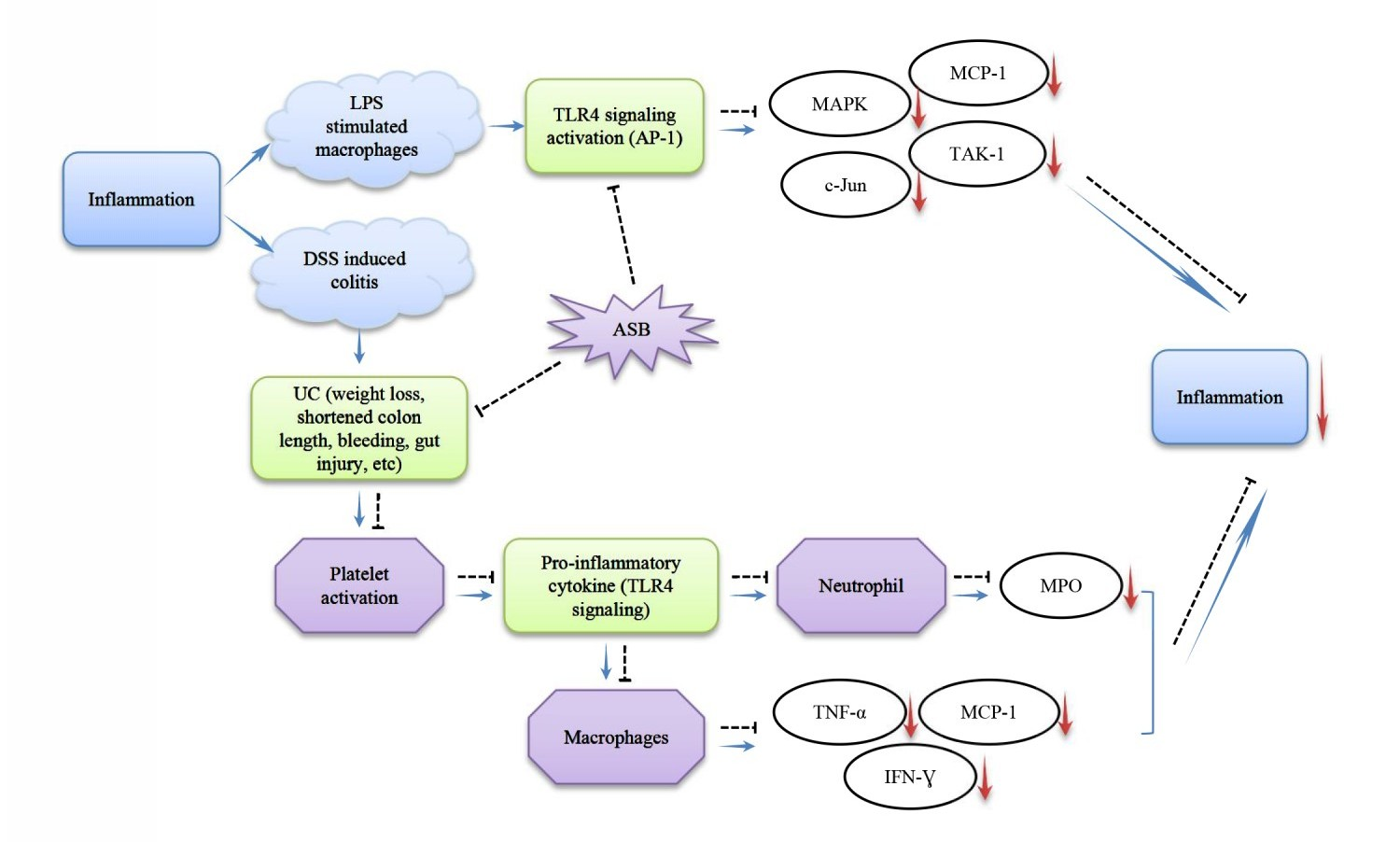
2.1. Characterization of the Aqueous Extract of Sea Bass (ASB)
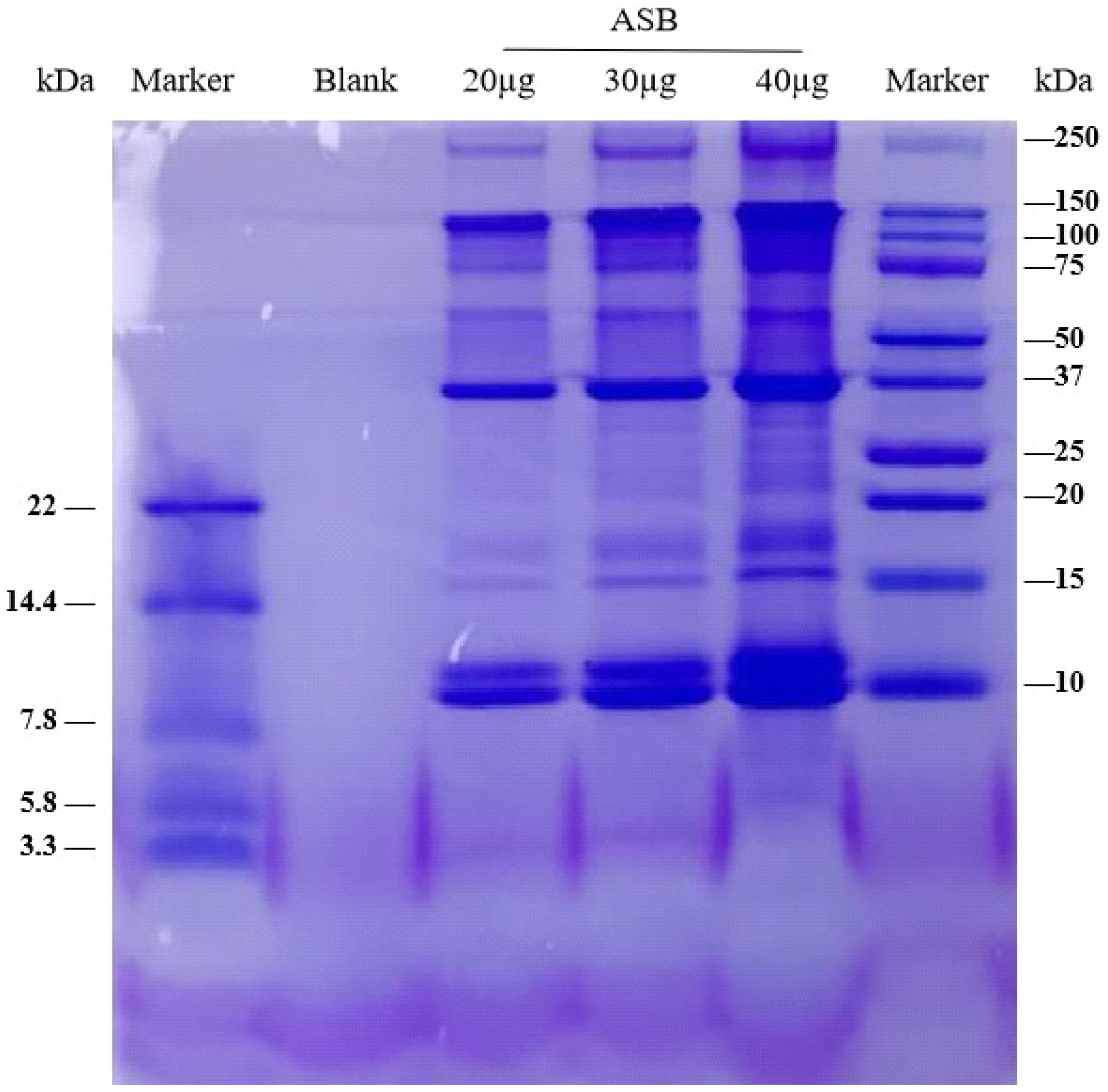
2.2. ASB Ameliorated DSS-Induced Colitis
2.3. Hematological Parameters
2.4. ASB Reduced Intestinal Permeability
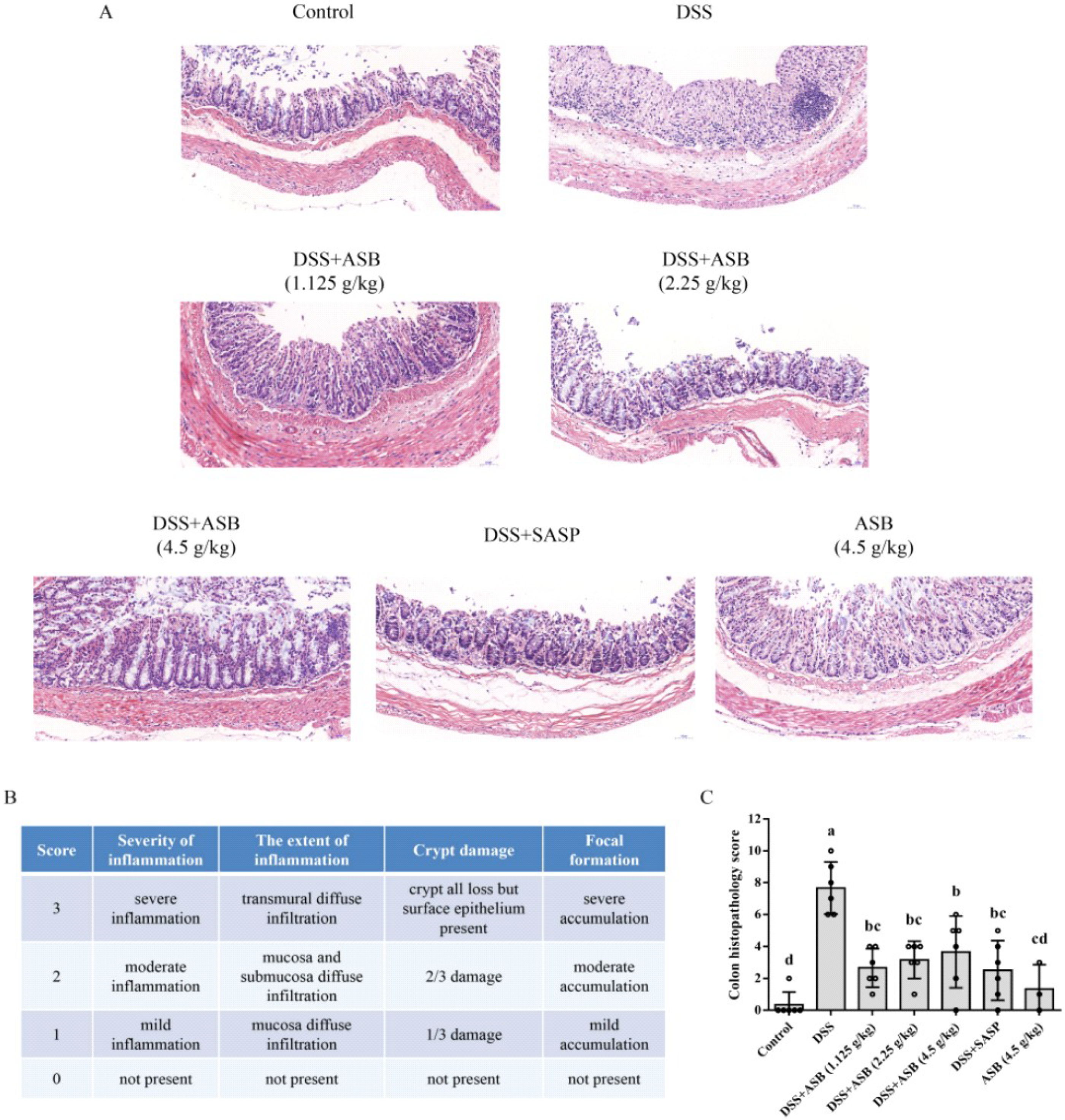
2.5. ASB Reduced Colonic Tissue Damage
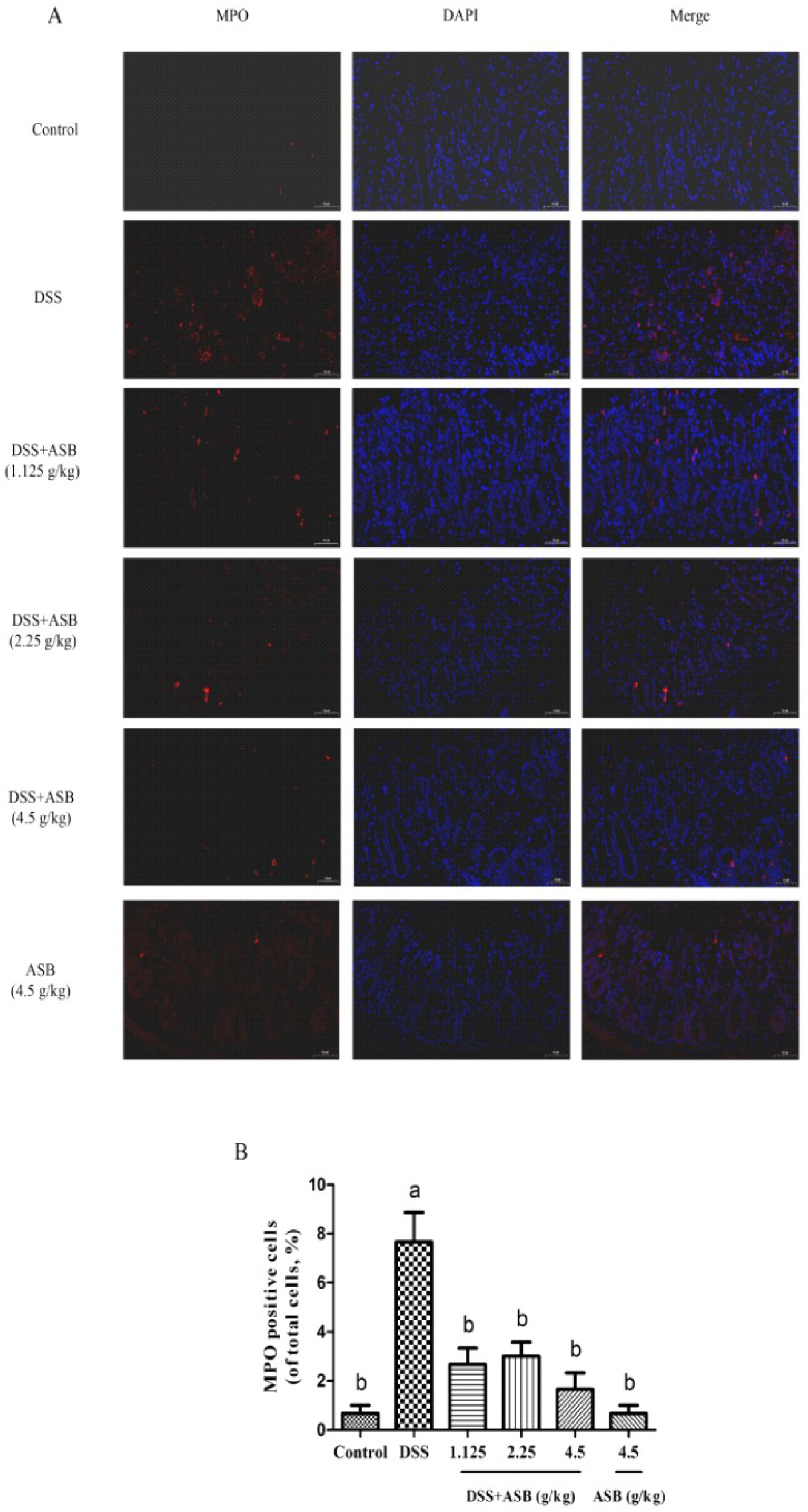
2.6. ASB Inhibited the Neutrophil Infiltration in Impaired Colon
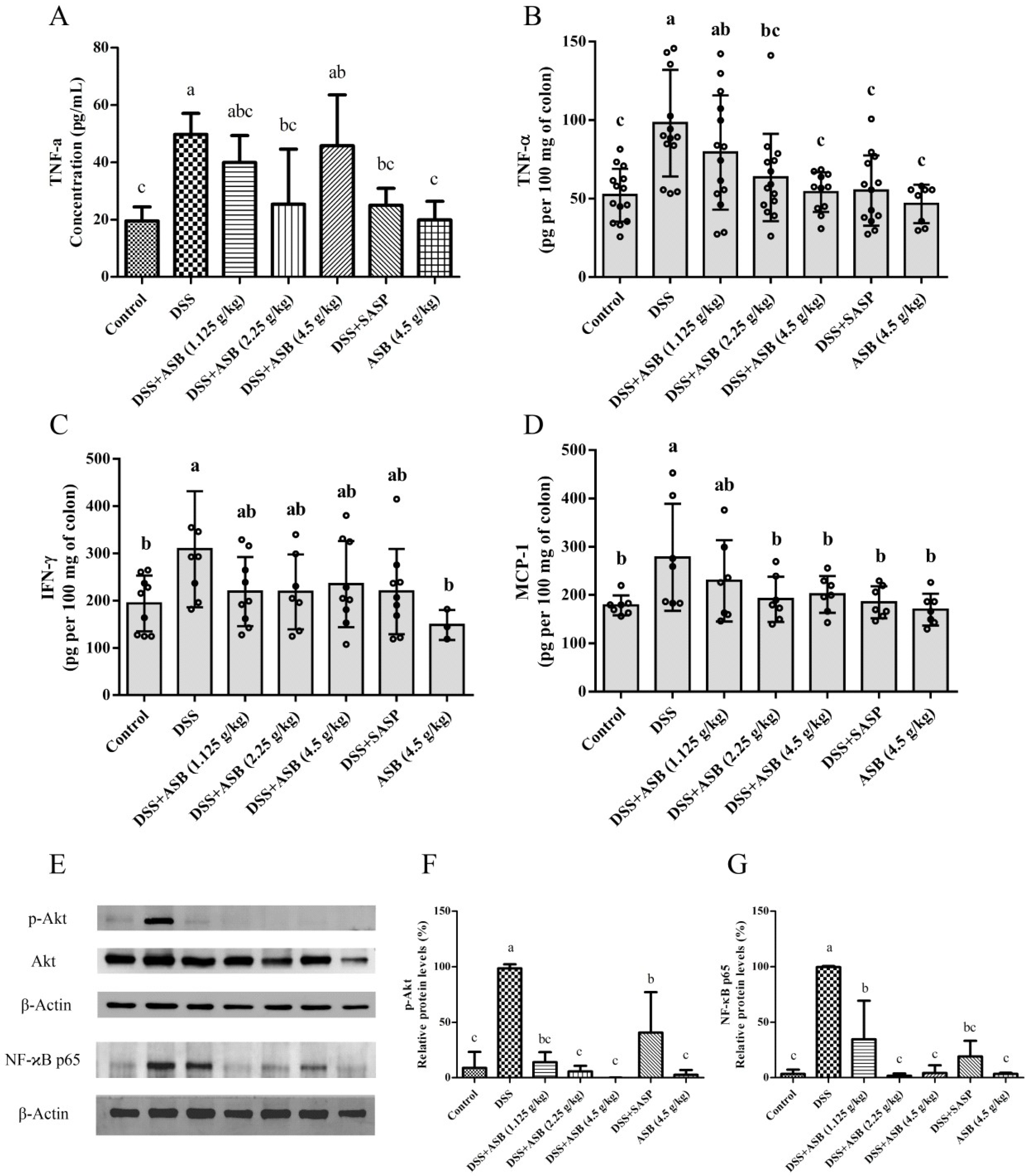
2.7. ASB Suppressed the Production of Pro-Inflammatory Mediators in the Impaired Colon
2.8. ASB Improved UC through TLR4 Signaling Inhibition
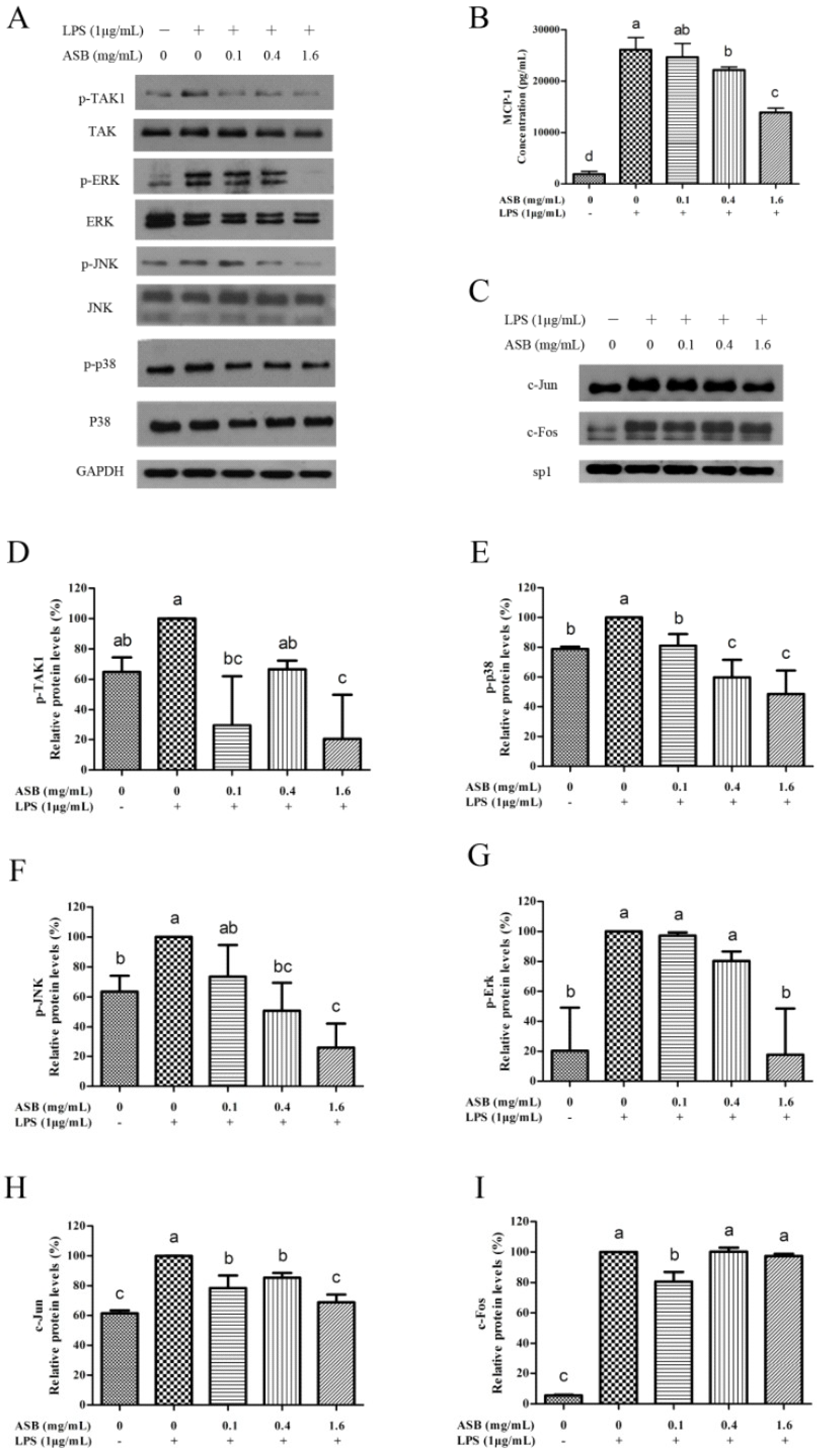
2.9. ASB Down-Regulated the Expression Levels of Inflammatory Mediators in LPS-Activated Macrophages through TLR4 Signaling Inhibition
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Sea Bass Materials and Preparation of Aqueous Extract of Sea Bass
4.3. Characterization of the Aqueous Extract of Sea Bass (ASB)
4.4. Animals
4.5. Establishment of Ulcerative Colitis (UC) Model
4.6. Intestinal Permeability In Vivo
4.7. Disease Activity Index (DAI)
4.8. Histology and Immunofluorescence
4.9. Cell Culture
4.10. Western Blotting
4.11. Enzyme-Linked Immunosorbent Assay (ELISA) Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASB | Aqueous extract of sea bass |
| DSS | Dextran sulphate sodium |
| ELISA | Enzyme-linked immunosorbent assay |
| IBD | Inflammatory bowel disease |
| IFN-γ | Interferon gamma |
| IF | Immunofluorescence |
| LPS | Lipopolysaccharide |
| MPO | Myeloperoxidase |
| MCP-1 | Monocyte chemoattractant protein-1 |
| SASP | Sulfasalazine |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor alpha |
| UC | Ulcerative colitis |
References
- Fumery, M.; Singh, S.; Dulai, P.S.; Gower-Rousseau, C.; Peyrin-Biroulet, L.; Sandborn, W.J. Natural history of adult ulcerative colitis in population-based cohorts: A systematic review. Clin. Gastroenterol. Hepatol. 2018, 16, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Qiu, Y.; Chen, B.; Li, Y.; Xiong, S.; Zhang, S.; He, Y.; Zeng, Z.R.; Ben-Horin, S.; Chen, M.H.; Mao, R. Risk factors and long-term outcome of disease extent progression in Asian patients with ulcerative colitis: A retrospective cohort study. BMC Gastroenterol. 2019, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Loddo, I.; Romano, C. Inflammatory bowel disease: Genetics, epigenetics, and pathogenesis. Front. Immunol. 2015, 6, 551. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N.; Xavier, R.J.; Podolsky, D.K. Inflammatory Bowel Diseases: Pathogenesis. In Yamada’s Textbook of Gastroenterol; Wiley-Blackwell: New York, NY, USA, 2015; pp. 1364–1377. [Google Scholar]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008, 453, 620–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, L.; Ley, R.E.; Volchkov, P.Y.; Stranges, P.B.; Avanesyan, L.; Stonebraker, A.C.; Hu, C.Y.; Wong, S.S.; Szot, G.L.; Bluestone, J.A.; et al. Innate immunity and intestinal microbiota in the development of type 1 diabetes. Nature 2008, 455, 1109–1113. [Google Scholar] [CrossRef]
- Hakansson, A.; Badia Tormo, N.; Baridi, A.; Xu, J.; Molin, G.; Hagslatt, M.L.; Karlsson, C.; Jeppsson, B.; Cilio, C.M.; Ahrne, S. Immunological alteration and changes of gut microbiota after dextran sulfate sodium (DSS) administration in mice. Clin. Exp. Med. 2015, 15, 107–120. [Google Scholar] [CrossRef]
- Heinsbroek, S.E.M.; Williams, D.L.; Welting, O.; Meijer, S.L.; Gordon, S.; Jonge, W.J. Orally delivered β-glucans aggravate dextran sulfate sodium (DSS)—Induced intestinal inflammation. Nutr. Res. 2015, 35, 1106–1112. [Google Scholar] [CrossRef]
- Chen, L.; Wilson, J.E.; Koenigsknecht, M.J.; Chou, W.C.; Montgomery, S.A.; Truax, A.D.; Brickey, W.J.; Packey, C.D.; Maharshak, N.; Matsushima, G.K.; et al. NLRP12 attenuates colon inflammation by maintaining colonic mircobial diversity and promoting protective commensal bacterial growth. Nat. Immunol. 2017, 18, 541–551. [Google Scholar] [CrossRef]
- Zhu, W.H.; Winter, M.G.; Byndloss, M.X.; Spiga, L.; Duerkop, B.A.; Hughes, E.R.; Buttner, L.; Romao, E.D.L.; Behrendt, C.L.; Lopez, C.A.; et al. Precision editing of the gut microbiota ameliorates colitis. Nature 2018, 553, 208–211. [Google Scholar] [CrossRef]
- Cui, L.; Feng, L.; Zhang, Z.H.; Jia, X.B. The anti-inflammation effect of baicalin on experimental colitis through inhibiting TLR4/NF-κB pathway activation. Int. Immunopharmacol. 2014, 23, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Lee, I.A.; Gu, W.; Hyam, S.R.; Kim, D.H. β-Sitosterol attenuates high fat diet induced intestinal inflammation in mice by inhibiting the binding of lipopolysaccharide to toll-like receptor 4 in the NF-κB pathway. Mol. Nutr. Food Res. 2014, 58, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.X.; Shao, M.J.; Qi, Q.; Xu, Y.S.; Yang, X.Q.; Zhu, F.H.; He, S.J.; He, P.L.; Feng, C.L.; Wu, Y.W.; et al. Artemisinin analogue SM934 ameliorates DSS-induced mouse ulcerative colitis via suppressing neutrophils and macrophages. Acta Pharmacol. Sin. 2018, 39, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, M.; Minami, M.; Seno, H.; Yasui, M.; Komekado, H.; Higuchi, S.; Fujikawa, R.; Nakanishi, Y.; Fukuda, A.; Kawada, K.; et al. EP4 receptor—Associated protein in macrophages ameliorates colitis and colitis-associated tumorigenesis. PLoS Genet. 2015, 11, e1005542. [Google Scholar] [CrossRef] [PubMed]
- Haribhai, D.; Ziegelbauer, J.; Jia, S.; Upchurch, K.; Yan, K.; Schmitt, E.G.; Salzman, N.H.; Simpson, P.; Hessner, M.J.; Chatila, T.A.; et al. Alternatively activated macrophages boost induced regulatory T and Th17 cell responses during immunotherapy for colitis. J. Immunol. 2016, 196, 3305–3317. [Google Scholar] [CrossRef]
- Liu, Y.T.; Fang, S.L.; Li, X.Y.; Feng, J.; Du, J.; Guo, L.J.; Su, Y.Y.; Zhou, J.; Ding, G.; Bai, Y.X.; et al. Aspirin inhibits LPS-induced macrophage activation via the NF-κB pathway. Sci. Rep. 2017, 7, 11549. [Google Scholar] [CrossRef]
- Pham, T.H.; Kim, M.S.; Le, M.Q.; Song, Y.S.; Bak, Y.; Ryu, H.W.; Oh, S.R.; Yoon, D.Y. Fargesin exerts anti-inflammatory effects in THP-1 monocytes by suppressing PKC-dependent AP-1 and NF-ĸB signaling. Phytomedicine 2017, 24, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Lin, Y.J.; Ho, C.T.; Ye, G.C. The inhibitory effect of pterostilbene on inflammatory responses during the interaction of 3T3-L1 adipocytes and RAW264.7 macrophages. J. Agric. Food Chem. 2013, 61, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.J.; Choi, S.H.; Yu, G.J.; Hong, S.H.; Chung, Y.H.; Kim, C.H.; Yoon, H.M.; Kim, G.Y.; Kim, B.W.; Choi, Y.H. Anti-inflammatory potential of total saponins derived from the roots of Panax ginseng in lipopolysaccharide—Activated RAW 264.7 macrophages. Exp. Ther. Med. 2016, 11, 1109–1115. [Google Scholar] [CrossRef]
- Chen, H.; Sohn, J.; Zhang, L.; Tian, J.; Chen, S.; Bjeldanes, L.F. Anti-inflammatory effects of chicanine on murine macrophage by down-regulating LPS-induced inflammatory cytokines in IκBα/MAPK/ERK signaling pathways. Eur. J. Pharmacol. 2014, 724, 168–174. [Google Scholar] [CrossRef]
- Cheng, B.C.Y.; Ma, X.Q.; Kwan, H.Y.; Tse, K.W.; Cao, H.H.; Su, T.; Shu, X.; Wu, Z.Z.; Yu, Z.L. A herbal formula consisting of Rosae Multiflorae Fructus and Lonicerae Japonicae FLoS inhibits inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. J. Ethnopharmacol. 2014, 153, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Yang, Y.D.; Bian, Z.X.; Xu, B.J. Characterization and anti-inflammatory potential of and exopolysaccharide from submerged mycelial culture of Schizophyllum commune. Front. Pharmacol. 2017, 8, 252. [Google Scholar] [CrossRef] [PubMed]
- Delhase, M. Kappa B kinase and NF-kappa B signaling in response to pro-inflammatory cytokines. Methods Mol. Biol. 2003, 225, 7–17. [Google Scholar] [PubMed]
- Ahn, C.B.; Jung, W.K.; Park, S.J.; Kim, Y.T.; Kim, W.S.; Je, J.Y. Gallic acid-g-chitosan modulates inflammatory responses in LPS stimulated RAW264.7 cells via NF-ĸB, AP-1 and MAPK pathways. Inflammation 2015, 39, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Joos, G.; Boon, L.; Tournoy, K.; Provoost, S.; Maes, T. Role of tumor necrosis factor—α and its receptors in diesel exhaust particle-induced pulmonary inflammation. Sci. Rep. 2017, 7, 711508. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, Z.; Su, J.; Pan, N.; Song, Q. Anti-inflammatory activity of 3β-hydroxycholest-5-en-7-one isolated from Hippocampus trimaculatus leach via inhibiting iNOS, TNF-α, and 1L-1β of LPS induced RAW 264.7 macrophage cells. Food Funct. 2017, 8, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.C.Y.; Yu, H.; Su, T.; Fu, X.Q.; Guo, H.; Li, T.; Cao, H.H.; Tse, A.K.W.; Kwan, H.Y.; Yu, Z.L. A herbal formula comprising Rosae Multiflorae Fructus and Lonicerae Japonicae FLoS inhibits the production of inflammatory mediators and the IRAK-1/TAK1 and TBK1/IRF3 pathways in RAW 264.7 and THP-1 cells. J. Ethnopharmacol. 2015, 174, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.X.; Yang, L.; Tian, L.; Li, W.Y.; Yang, L.; Li, L.Y. Macrophage migration inhibitor factor up-regulates MCP-1 expression in an autocrine manner in hepatocytes during acute mouse liver injury. Sci. Rep. 2016, 6, 27665. [Google Scholar] [CrossRef]
- Peskar, B.M.; Dreyling, K.W.; Peskar, B.A.; May, B.; Goebell, H. Enhanced formation of sulfidopeptide-leukotrienes in ulcerative colitis and Crohn’s disease: Inhibition by sulfasalazine and 5-aminosalicylic acid. Agents Actions Suppl. 1986, 18, 381–383. [Google Scholar] [CrossRef]
- Sutherland, L.R.; MacDonald, J.K. Oral 5-aminosalicylic acid for induction of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2006. [Google Scholar] [CrossRef]
- Du, B.; Yang, Y.D.; Bian, Z.X.; Xu, B.J. Molecular weight and helix conformation determine intestinal anti-inflammatory effects of exopolysaccharide from Schizophyllum commune. Carbohydr. Polym. 2017, 172, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Gascon, M.J.Y.; Selma, M.V.; Sarrias, A.G.; Toti, S.; Ceron, J.J.; Barberan, F.T.; Dolara, P.; Espin, J.C. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in DSS-induced colitis rat model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.T.; Lin, C.Y.; Ho, D.H.H.; Peng, J.; Chen, Y.; Tsang, S.W.; Wong, M.; Zhang, X.J.; Zhang, M.; Bian, Z.X. Inhibitory effect of the gallotannin corilagin on dextran sulfate sodium-induced murine ulcerative colitis. J. Nat. Prod. 2013, 76, 2120–2125. [Google Scholar] [CrossRef] [PubMed]
- Markovic, B.S.; Nikolic, A.; Gazdic, M.; Bojic, S.; Vucicevic, L.; Kosic, M.; Mitrovic, S.; Milosavljevic, M.; Besra, G.; Trajkovic, V.; et al. Gal-3 plays an important pro-inflammatory role in the induction phase of acute colitis by promoting activation of NLRP3 inflammasome and production of IL-1β in macrophages. J. Crohns Colitis 2016, 10, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Possemiers, S.; Wiele, T.V.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Zhang, H.; Wu, H.; Li, H.; Liu, L.; Guo, J.; Li, C.Y.; Shih, D.Q.S.; Zhang, X.L. Protective role of 1, 25 (OH)2 vitamin D3 in the mucosal injury and epithelial barrier disruption in DSS-induced acute colitis in mice. BMC Gastroenterol. 2012, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- Muthas, D.; Reznichenko, A.; Balendran, C.A.; Böttcher, G.; Clausen, I.G.; Kärrman Mårdh, C.; Ottosson, T.; Uddin, M.; MacDonald, T.T.; Danese, S.; et al. Neutrophils in ulcerative colitis: A review of selected biomarkers and their potential therapeutic implications. Scand. J. Gastroenterol. 2017, 52, 125–135. [Google Scholar] [CrossRef]
- Fukata, M.; Shang, L.; Santaolalla, R.; Sotolongo, J.; Pastorini, C.; España, C.; Ungaro, R.; Harpaz, N.; Cooper, H.S.; Elson, G.; et al. Constitutive activation of epithelial TLR4 augments inflammatory responses to mucosal injury and drives colitis-associated tumorigenesis. Inflamm. Bowel Dis. 2010, 17, 1464–1473. [Google Scholar] [CrossRef]
- Abreu, M.T.; Fukata, M.; Arditi, M. TLR signaling in the gut in health and disease. J. Immunol. 2005, 174, 4453–4460. [Google Scholar] [CrossRef]
- Bantel, H.; Berg, C.; Vieth, M.; Stolte, M.; Kruis, W.; Schulze-Osthoff, K. Mesalazine inhibits activation of transcription factor NF-κB in inflamed mucosa of patients with ulcerative colitis. Am. J. Gastroenterol. 2000, 95, 3452. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Endale, M.; Park, S.C.; Kim, S.; Kim, S.H.; Yang, Y.; Cho, J.Y.; Rhee, M.H. Quercetin disrupts tyrosine-phosphorylated phosphatidylinositol 3-kinase and myeloid differentiation factor-88 association, and inhibits MAPK/AP-1 and IKK/NF-κB-induced inflammatory mediators production in RAW 264.7 cells. Immunobiology 2013, 218, 1452–1467. [Google Scholar] [CrossRef] [PubMed]
- Demon, D.; Kuchmiy, A.; Fossoul, A.; Zhu, Q.; Kanneganti, T.D.; Lamkanfi, M. Caspase-11 is expressed in the colonic mucosa and protects against dextran sodium sulfate-induced colitis. Nature 2014, 7, 1480–1491. [Google Scholar] [Green Version]
- Chen, Q.Y.; Duan, X.Y.; Fan, H.; Xu, M.; Tang, Q.; Zhang, L.J.; Shou, Z.X.; Liu, X.X.; Zuo, D.M.; Yang, J.; et al. Oxymatrine protects against DSS-induced colitis via inhibiting the PI3K/AKT signaling pathway. Int. Immunopharmacol. 2017, 53, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Jayachandran, M.; Xu, B.J.; Yu, Z.L. Sea bass (Lateolabrax maculatus) accelerates wound healing: A transition from inflammation to proliferation. J. Ethnopharmacol. 2019, 236, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.L. Kjeldahl method for total nitrogen. Anal. Chem. 1950, 22, 354–358. [Google Scholar] [CrossRef]
- Jambrak, A.R.; Mason, T.J.; Lelas, V.; Paniwnyk, L.; Herceg, Z. Effect of ultrasound treatment on particle size and molecular weight of whey proteins. J. Food Eng. 2014, 121, 15–23. [Google Scholar] [CrossRef]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host derived nitrate boosts growth of E. coli in the inflamed gut. Science 2013, 339, 708–771. [Google Scholar] [CrossRef]
- Xu, X.L.; Yin, P.; Wan, C.R.; Chong, X.L.; Liu, M.J.; Cheng, P.; Chen, J.J.; Liu, F.H.; Xu, J.Q. Punicalagin inhibits inflammation in LPS-induced RAW264.7 macrophages via the suppression of TLR4-mediated MAPKs and NF-κB activation. Inflammation 2014, 37, 956–965. [Google Scholar] [CrossRef]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z. Indirubin inhibits LPS-induced inflammation via TLR4 abrogation mediated by the NF-κB and MAPK signaling pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hematological Parameters | Control | DSS | DSS+ASB (1.125 g/kg) | DSS+ASB (2.25 g/kg) | DSS+ASB (4.5 g/kg) | DSS+SASP | NVR |
|---|---|---|---|---|---|---|---|
| RBC (1012/L) | 7.39 ± 0.55 a | 5.37 ± 1.25 c | 6.67 ± 1.15 ab | 6.23 ± 0.64 bc | 6.01 ± 0.90 bc | 6.07 ± 0.56 bc | 6.68~8.28 |
| HGB (g/L) | 115 ± 8 a | 83 ± 20 c | 101 ± 17 ab | 95 ± 10 bc | 93 ± 14 bc | 90 ± 8 bc | 106~129 |
| HCT (%) | 37.2 ± 2.6 a | 26.3 ± 6.1 c | 32.8 ± 5.7 ab | 30.5 ± 3.2 bc | 29.3 ± 4.5 bc | 29.3 ± 2.5 bc | 33.9~41.8 |
| MCV (fL) | 50.4 ± 0.8 a | 49.0 ± 1.0 b | 49.2 ± 0.7 b | 48.9 ± 0.5 b | 48.8 ± 0.5 b | 48.4 ± 1.7 b | 49.2~51.3 |
| MCH (pg) | 15.5 ± 0.2 a | 15.4 ± 0.2 ab | 15.2 ± 0.1 b | 15.3 ± 0.1 ab | 15.5 ± 0.2 a | 14.9 ± 0.6 c | 15.2~15.9 |
| MCHC (g/L) | 308 ± 4 bc | 314 ± 8 ab | 309 ± 4 bc | 314 ± 5 ab | 318 ± 5 a | 308 ± 4 bc | 299~313 |
| PDW | 14.8 ± 0.1 a | 14.3 ± 0.2 c | 14.6 ± 0.2 b | 14.5 ± 0.2 b | 14.5 ± 0.2 b | 14.4 ± 0.1 bc | 14.7~14.9 |
| WBC (109/L) | 3.76 ± 0.65 ab | 3.30 ± 0.87 b | 5.14 ± 1.10 a | 4.07 ± 2.34 ab | 2.59 ± 0.95 b | 4.02 ± 1.29 ab | 2.53~4.62 |
| Neu# (109/L) | 2.08 ± 0.81 abc | 1.52 ± 0.34 bc | 2.84 ± 0.61 a | 2.29 ± 1.91 ab | 1.15 ± 0.52 c | 1.86 ± 0.63 abc | 0.47~3.01 |
| Lymph# (109/L) | 1.68 ± 0.35 ab | 1.78 ± 0.61 ab | 2.29 ± 0.62 a | 1.77 ± 0.60 ab | 1.43 ± 0.64 b | 2.16 ± 0.72 a | 1.23~2.31 |
| Mon# (109/L) | 0 ± 0 b | 0 ± 0 b | 0.01 ± 0.01 a | 0 ± 0 b | 0 ± 0 b | 0 ± 0 b | 0 |
| Eos# (109/L) | 0 ± 0.01 a | 0 ± 0 a | 0 ± 0 a | 0.01 ± 0.01 a | 0 ± 0 a | 0 ± 0 a | 0~0.02 |
| Bas# (109/L) | 0 ± 0 b | 0 ± 0 b | 0 ± 0 b | 0 ± 0 b | 0.01 ± 0.01 a | 0 ± 0 b | 0 |
| Neu% (%) | 53.3 ± 16.5 a | 46.8 ± 6.3 a | 55.5 ± 6.7 a | 51.6 ± 12.7 a | 45.2 ± 13.2 a | 46.2 ± 4.9 a | 39.6~66.1 |
| Lymph% (%) | 46.5 ± 16.4 a | 53.1 ± 6.3 a | 44.2 ± 6.8 a | 48.0 ± 12.4 a | 54.1 ± 13.2 a | 53.7 ± 5.0 a | 33.7~81.3 |
| Mon% (%) | 0 ± 0 b | 0.04 ± 0.1 ab | 0.2 ± 0.2 a | 0.1 ± 0.1 ab | 0.1 ± 0.2 ab | 0.1 ± 0.1 ab | 0 |
| Eos% (%) | 0.2 ± 0.2 a | 0 ± 0 a | 0.1 ± 0.1 a | 0.2 ± 0.4 a | 0.2 ± 0.2 a | 0 ± 0 a | 0~0.7 |
| Bas% (%) | 0 ± 0.1 b | 0.1 ± 0.1 b | 0 ± 0.1 b | 0.2 ± 0.2 b | 0.4 ± 0.4 a | 0 ± 0 b | 0~0.2 |
| RDW-CV (%) | 13.3 ± 0.8 b | 13.6 ± 2.6 b | 13.6 ± 1.3 b | 13.7 ± 1.3 b | 13.1 ± 0.6 b | 17.2 ± 1.6 a | 12.6~15.1 |
| PLT (109/L) | 611 ± 53 a | 576 ± 146 a | 681 ± 122 a | 631 ± 94 a | 569 ± 128 a | 688 ± 91 a | 526~662 |
| MPV (fL) | 5.3 ± 0.2 a | 5.3 ± 0.2 a | 5.3 ± 0.2 a | 5.4 ± 0.2 a | 5.3 ± 0.1 a | 5.3 ± 0.1 a | 5.2~5.7 |
| PCT (%) | 0.325 ± 0.032 a | 0.304 ± 0.072 a | 0.359 ± 0.067 a | 0.339 ± 0.058 a | 0.299 ± 0.065 a | 0.363 ± 0.048 a | 0.274~0.362 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Jayachandran, M.; Zhang, W.; Chen, L.; Du, B.; Yu, Z.; Xu, B. Dietary Supplementation with Sea Bass (Lateolabrax maculatus) Ameliorates Ulcerative Colitis and Inflammation in Macrophages through Inhibiting Toll-Like Receptor 4-Linked Pathways. Int. J. Mol. Sci. 2019, 20, 2907. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122907
Chen J, Jayachandran M, Zhang W, Chen L, Du B, Yu Z, Xu B. Dietary Supplementation with Sea Bass (Lateolabrax maculatus) Ameliorates Ulcerative Colitis and Inflammation in Macrophages through Inhibiting Toll-Like Receptor 4-Linked Pathways. International Journal of Molecular Sciences. 2019; 20(12):2907. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122907
Chicago/Turabian StyleChen, Jiali, Muthukumaran Jayachandran, Wenxia Zhang, Lingyuqing Chen, Bin Du, Zhiling Yu, and Baojun Xu. 2019. "Dietary Supplementation with Sea Bass (Lateolabrax maculatus) Ameliorates Ulcerative Colitis and Inflammation in Macrophages through Inhibiting Toll-Like Receptor 4-Linked Pathways" International Journal of Molecular Sciences 20, no. 12: 2907. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122907