Induction of Necrosis in Human Macrophage Cell Lines by Corynebacterium diphtheriae and Corynebacterium ulcerans Strains Isolated from Fatal Cases of Systemic Infections


Abstract
:1. Introduction
2. Results
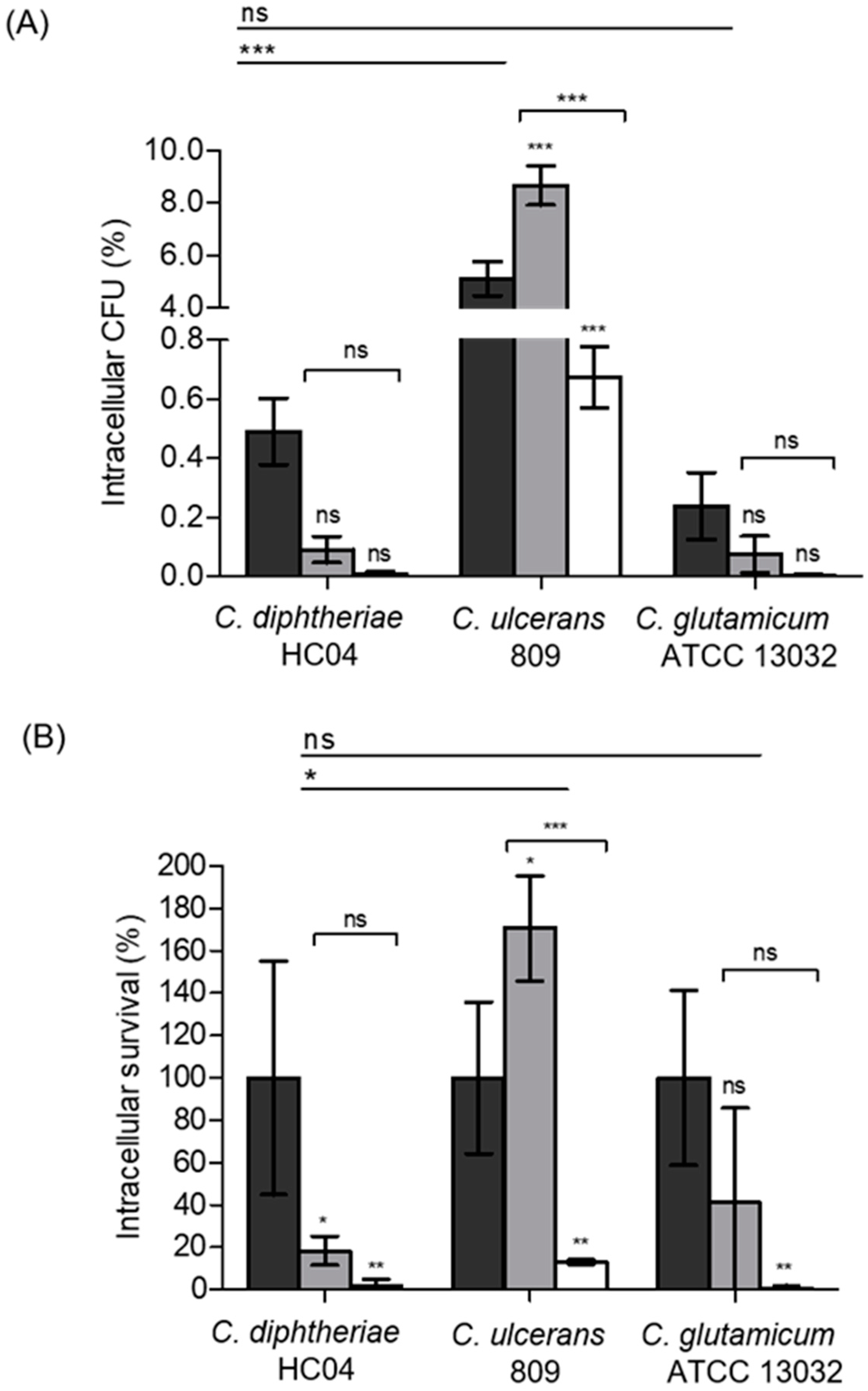
2.1. Species-Specific Proliferation and Elimination of Corynebacteria Inside Human Macrophages
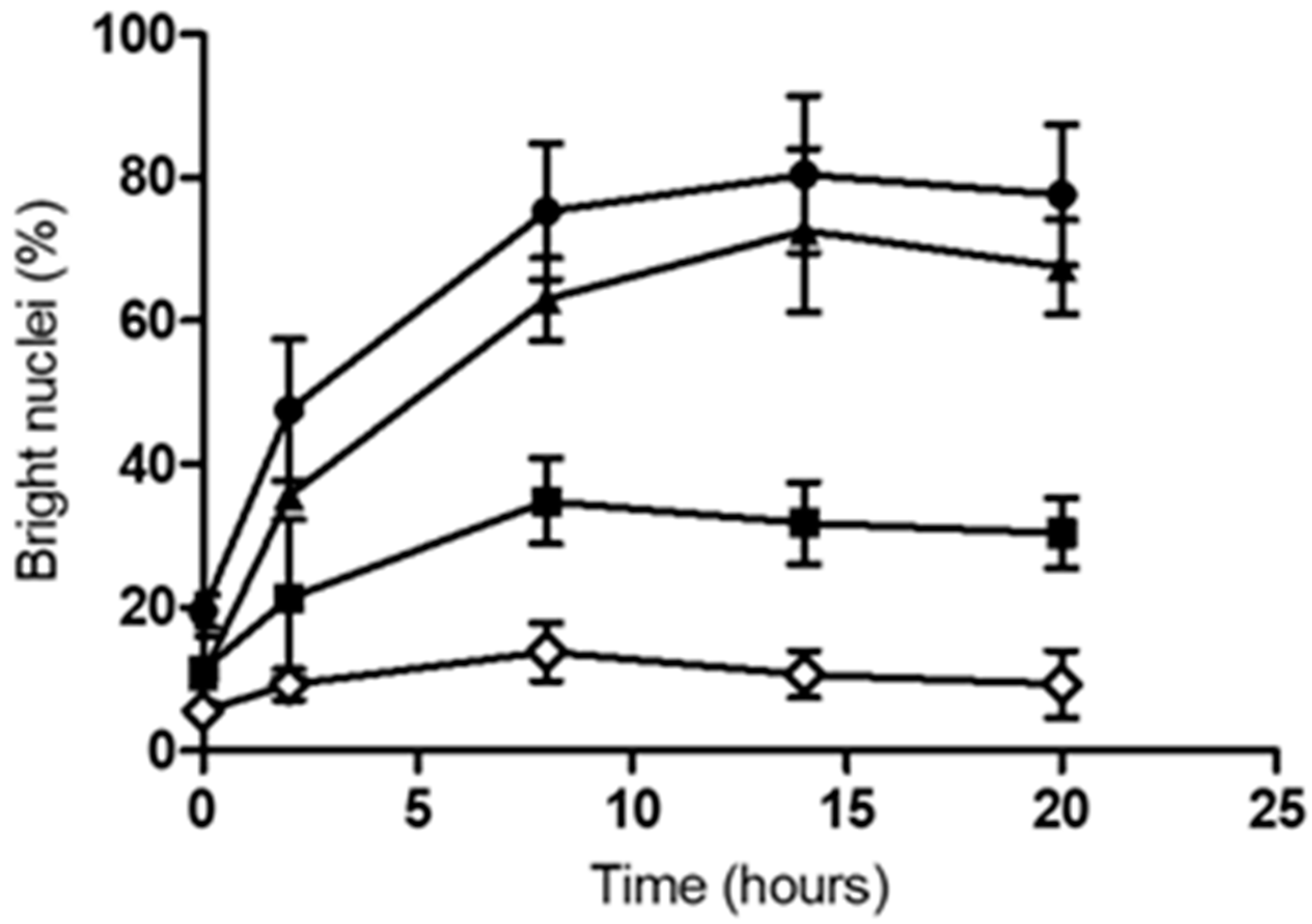
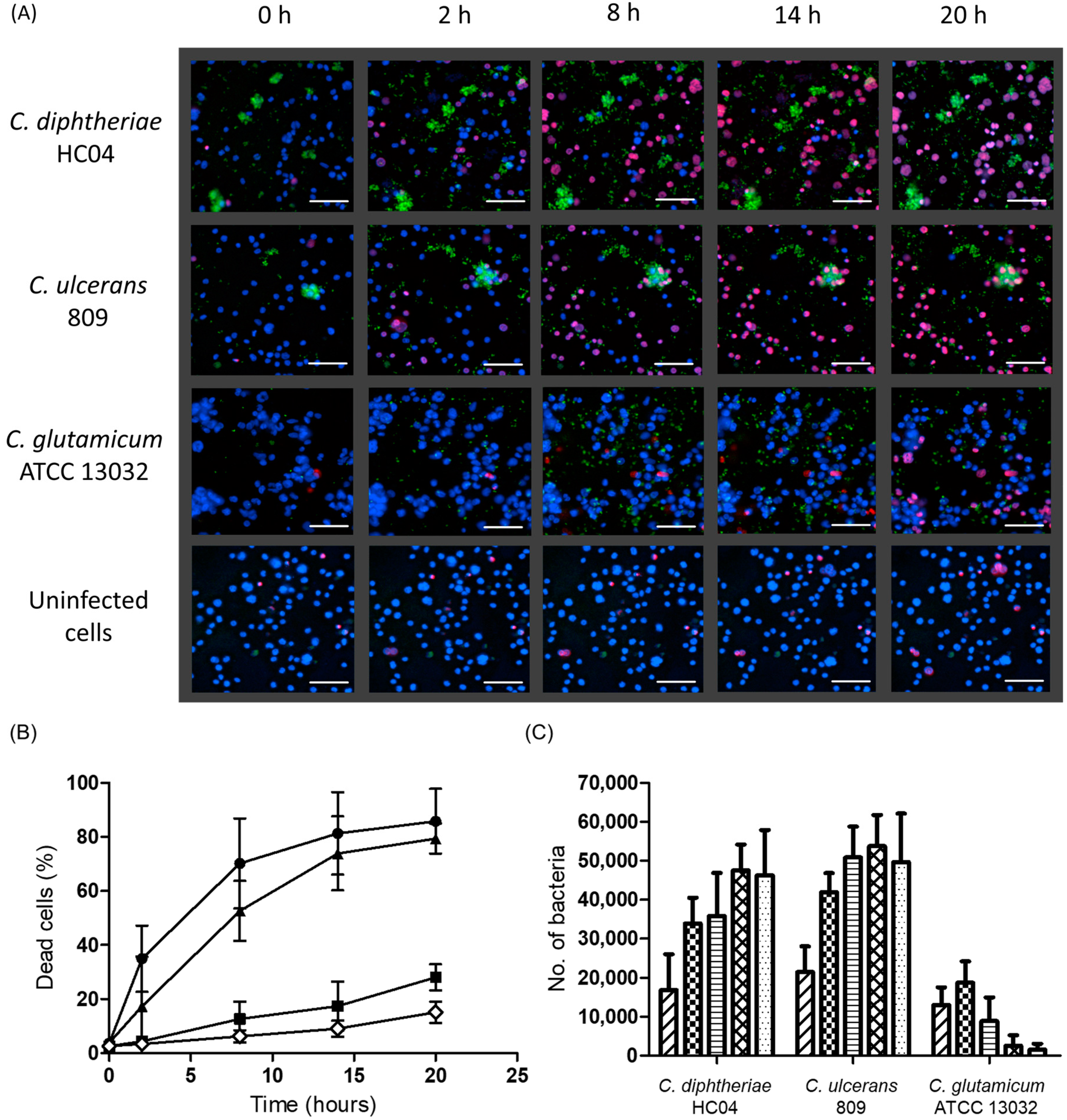
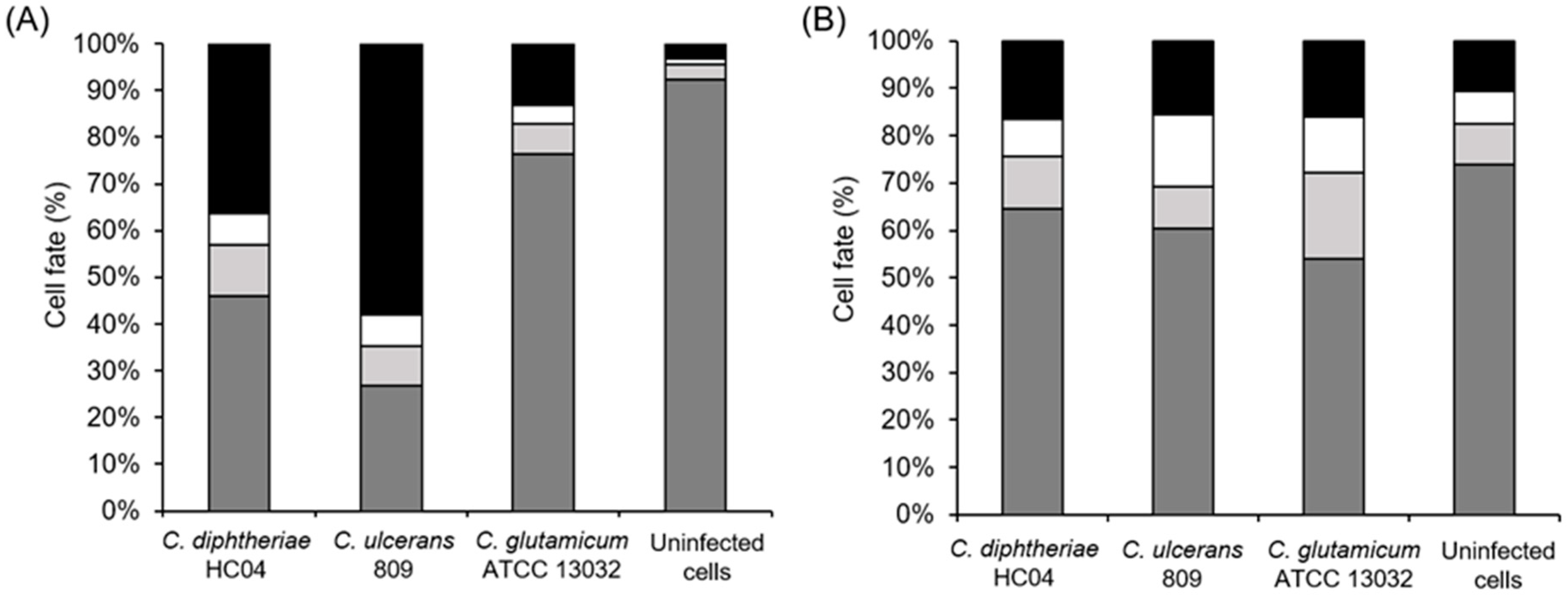
2.2. Infection with C. diphtheriae and C. ulcerans Results in Condensation of DNA in Macrophage Nuclei and Induces Cell Lysis
2.3. Detrimental Effects Depend on Endocytosis of C. diphtheriae HC04 and C. ulcerans 809 and Bacterial Replication within Macrophages
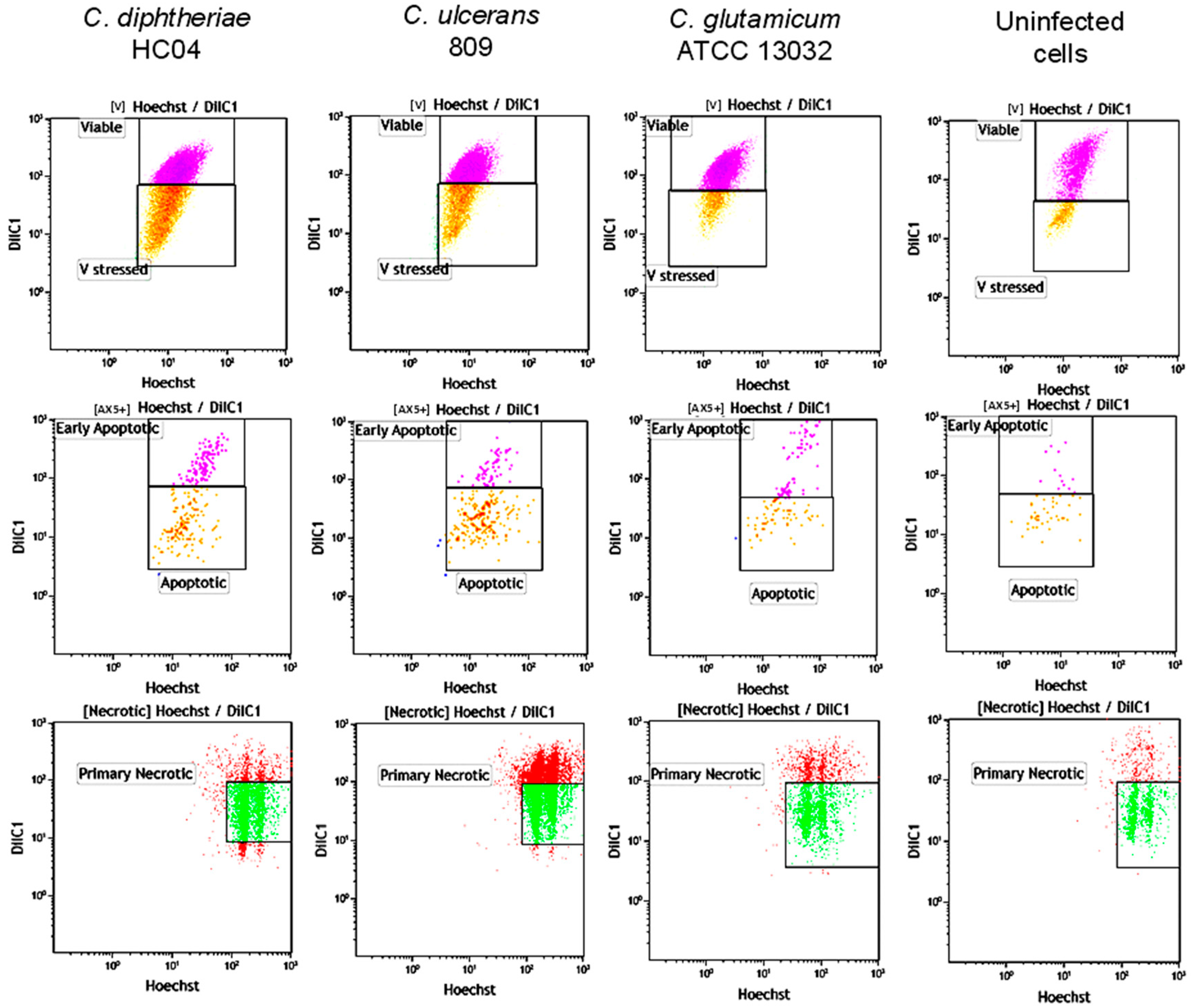
2.4. Induction of Necrosis in Human Macrophages by C. diphtheriae and C. ulcerans
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Phagocytosis and Survival Assay
4.3. Live Cell Imaging
4.4. Flow Cytometry
4.5. Cytotoxicity Assay
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CFU | Colony-forming unit(s) |
| EDTA | Ethylenediaminetetraacetic acid |
| FACS | Fluorescence-activated cell sorting |
| FCS | Fetal calf serum |
| FITC | Fluorescein isothiocyanate |
| GFP | Green-fluorescent protein |
| HI | Heart infusion |
| LDH | Lactate dehydrogenase |
| MOI | Multiplicity of infection |
| NEC-1 | Necrostatin-1 |
| OD600 | Optical density at 600 nm wavelength |
| PBS | Phosphate-buffered saline |
| PI | Propidium iodide |
| PLD | Phospholipase D |
| PMA | phorbol 12-myristate 13-acetate |
| RPMI | Roswell Park Memorial Institute |
References
- Tauch, A.; Sandbote, J. The family Corynebacteriaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 239–277. [Google Scholar]
- Burkovski, A. Diphtheria. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 237–245. [Google Scholar]
- Burkovski, A. Diphtheria and its etiological agents. In Corynebacterium diphtheriae and Related Toxigenic Species; Burkovski, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–14. [Google Scholar]
- Rappuoli, R.; Malito, E. History of diphtheria vaccine development. In Corynebacterium diphtheriae and Related Toxigenic Species; Burkovski, A., Ed.; Springer: Dordrecht, Germany, 2014; pp. 225–238. [Google Scholar]
- Burkovski, A. Pathogenesis of Corynebacterium diphtheriae and Corynebacterium ulcerans. In Human Emerging and Re-emerging Infections: Viral and Parasitic Infections; Singh, S.K., Ed.; Wiley Blackwell Press: Hoboken, NJ, USA, 2015; pp. 699–709. [Google Scholar]
- World Health Organization: Immunization, Vaccines and Biologicals. Available online: https://apps.who.int/immunization_monitoring/globalsummary/timeseries/tsincidencediphtheria.html (accessed on 16 July 2019).
- Hessling, M.; Feiertag, J.; Hoenes, K. Pathogens provoking most deaths worldwide: A review. Biosci. Biotech. Res. Comm. 2017, 10, 1–7. [Google Scholar]
- Gilbert, R.; Stewart, F.C. Corynebacterium ulcerans: A pathogenic microorganism resembling Corynebacterium diphtheriae. J. Lab. Clin. Med. 1927, 12, 756–761. [Google Scholar]
- Zakikhany, K.; Efstratiou, A. Diphtheria in Europe: Current problems and new challenges. Future Microbiol. 2012, 5, 595–607. [Google Scholar] [CrossRef]
- Meinel, D.M.; Margos, G.; Konrad, R.; Krebs, S.; Blum, H.; Sing, A. Next generation sequencing analysis of nine Corynebacterium ulcerans isolates reveals zoonotic transmission and a novel putative diphtheria toxin-encoding pathogenicity island. Genome Med. 2014, 6, 113. [Google Scholar] [CrossRef] [PubMed]
- Meinel, D.M.; Konrad, R.; Berger, A.; König, C.; Schmidt-Wieland, T.; Hogardt, M.; Bischoff, H.; Ackermann, N.; Hörmansdorfer, S.; Krebs, S.; et al. Zoonotic transmission of toxigenic Corynebacterium ulcerans strain, Germany, 2012. Emerg. Infect Dis. 2015, 21, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Hacker, E.; Ott, L.; Hasselt, K.; Mattos-Guaraldi, A.L.; Tauch, A.; Burkovski, A. Colonization of human epithelial cell lines by Corynebacterium ulcerans from human and animal sources. Microbiology 2015, 161, 1582–1591. [Google Scholar] [CrossRef]
- Hirata, R., Jr.; Pereira, G.A.; Filardy, A.A.; Gomes, D.L.R.; Damasco, P.V.; Rosa, A.C.; Nagao, P.E.; Pimenta, F.P.; Mattos-Guaraldi, A.L. Potential pathogenic role of aggregative-adhering Corynebacterium diphtheriae of different clonal groups in endocarditis. Braz. J. Med. Biol. Res. 2008, 41, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, R.S.; Hacker, E.; Antunes, C.A.; Weerasekera, D.; Dias, A.A.S.O.; Martins, C.A.; Hirata, R., Jr.; dos Santos, K.R.N.; Burkovski, A.; Mattos-Guaraldi, A.L. Pathogenic properties of a Corynebacterium diphtheriae strain isolated from a case of osteomyelitis. J. Med. Microbiol. 2016, 65, 1311–1321. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Antunes, C.A.; Lourêdo, L.S.; Viana, V.G.; dos Santos, C.S.; da Silva, J.F.R.; Hirata, R., Jr.; Hacker, E.; Mattos-Guaraldi, A.L.; Burkovski, A. Functional characterization of the collagen-binding protein DIP2093 and its influence on host-pathogen interaction and arthritogenic potential of Corynebacterium diphtheriae. Microbiology 2017, 163, 692–701. [Google Scholar] [CrossRef]
- Mattos-Guaraldi, A.L.; Sampaio, J.L.; Santos, C.S.; Pimenta, F.P.; Pereira, G.A.; Pacheco, L.G.; Miyoshi, A.; Azevedo, V.; Moreira, L.O.; Gutierrez, F.L.; et al. First detection of Corynebacterium ulcerans producing a diphtheria-like toxin in a case of human with pulmonary infection in the Rio de Janeiro metropolitan area, Brazil. Mem. Inst. Oswaldo. Cruz. 2008, 103, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Trost, E.; Al-Dilaimi, A.; Papavasiliou, P.; Schneider, J.; Viehoever, P.; Burkovski, A.; Soares, S.C.; Almeida, C.S.; Dorella, F.A.; Miyoshi, A.; et al. Comparative analysis of two complete Corynebacterium ulcerans genomes and detection of candidate virulence factors. BMC Genomics 2011, 12, 383. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, C.S.; dos Santos, L.S.; de Souza, M.C.; dos Santos, D.F.; de Souza, O.D.A.A.; Sabbadini, P.S.; Pereira, G.A.; Cabral, M.C.; Hirata, R., Jr.; Mattos-Guaraldi, A.L. Non-opsonic phagocytosis of homologous non-toxigenic and toxigenic Corynebacterium diphtheriae strains by human U-937 macrophages. Microbiol. Immunol. 2010, 54, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hacker, E.; Ott, L.; Schulze-Luehrmann, J.; Lührmann, A.; Wiesmann, V.; Wittenberg, T.; Burkovski, A. The killing of macrophages by Corynebacterium ulcerans. Virulence 2016, 7, 45–55. [Google Scholar] [CrossRef]
- Weerasekera, D.; Fastner, T.; Lang, R.; Burkovski, A.; Ott, L. Of mice and men: Interaction of Corynebacterium diphtheriae strains with murine and human phagocytes. Virulence 2019, 10, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Weerasekera, D.; Möller, J.; Kraner, M.E.; Antunes, C.A.; Mattos-Guaraldi, A.L.; Burkovski, A. Beyond diphtheria toxin: Cytotoxic proteins of Corynebacterium ulcerans and Corynebacterium diphtheriae. Microbiology 2019. [Google Scholar] [CrossRef]
- Wallez, Y.; Bouillot, S.; Soleilhac, E.; Huber, P.; Attrée, I.; Faudry, E. CLIQ-BID: A method to quantify bacteria-induced damage to eukaryotic cells by automated live-imaging of bright nuclei. Sci Rep 2018, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Walker, N.I.; Harmon, B.V.; Gobe, G.C.; Kerr, J.F. Patterns of cell death. Methods Achiev. Exp. Pathol. 1988, 13, 18–54. [Google Scholar]
- Munoz, L.E.; Maueroder, C.; Chaurio, R.; Berens, C.; Herrmann, M.; Janko, C. Colourful death: Six-parameter classification of cell death by flow cytometry-dead cells tell tales. Autoimmunity 2013, 46, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Ott, L.; Hacker, E.; Kunert, T.; Karrington, I.; Etschel, P.; Lang, R.; Wiesmann, V.; Wittenberg, T.; Singh, A.; Varela, C.; et al. Analysis of Corynebacterium diphtheriae macrophage interaction: Dispensability of corynomycolic acids for inhibition of phagolysosome maturation and identification of a new gene involved in synthesis of the corynomycolic acid layer. PLoS ONE 2017, 12, e0180105. [Google Scholar] [CrossRef] [PubMed]
- Trost, E.; Blom, J.; Soares, S.C.; Huang, I.; Al-Dilaimi, A.; Schröder, J.; Jaenicke, S.; Dorella, F.A.; Rocha, F.S.; Miyoshi, A.; et al. Pangenomic study of Corynebacterium diphtheriae that provides insights into the genomic diversity of pathogenic isolates from cases of classical diphtheria, endocarditis, and pneumonia. J. Bacteriol. 2012, 194, 3199–3215. [Google Scholar] [CrossRef]
- Bittel, M.; Gastiger, S.; Amin, B.; Hofmann, J.; Burkovski, A. Surface and extracellular proteome of the emerging pathogen Corynebacterium ulcerans. Proteomes 2018, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, A.L.; Krywult, J.; Corner, L.A.; Rothel, J.S.; Radford, A.J. Rational attenuation of Corynebacterium pseudotuberculosis: Potential cheesy gland vaccine and live delivery vehicle. Infect Immun. 1992, 60, 2900–2905. [Google Scholar]
- McNamara, P.J.; Bradley, G.A.; Songer, J.G. Targeted mutagenesis of the phospholipase D gene results in decreased virulence of Corynebacterium pseudotuberculosis. Mol. Microbiol. 1994, 12, 921–930. [Google Scholar] [CrossRef] [PubMed]
- McKean, S.C.; Davies, J.K.; Moore, R.J. Expression of phospholipase D, the major virulence factor of Corynebacterium pseudotuberculosis, is regulated by multiple environmental factors and plays a role in macrophage death. Microbiology 2007, 153, 2203–2211. [Google Scholar] [CrossRef] [PubMed]
- Exton, J.H. Phospholipase D: Enzymology, mechanisms of regulation, and function. Physiol. Rev. 1997, 77, 303–320. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.L.; Olsen, M.R.; Jagannath, C.; Actor, J.K. Multiple roles of cord factor in the pathogenesis of primary, secondary, and cavitary tuberculosis, including a revised description of the pathology of secondary disease. Ann. Clin. Lab. Sci. 2006, 36, 371–385. [Google Scholar] [PubMed]
- Schick, J.; Etschel, P.; Bailo, R.; Ott, L.; Bhatt, A.; Lepenies, B.; Kirschning, C.; Burkovski, A.; Lang, R. Toll-like receptor 2 and mincle cooperatively sense corynebacterial cell wall glycolipids. Infect Immun. 2017, 85, e00075-17. [Google Scholar] [CrossRef] [PubMed]
- Tauch, A.; Burkovski, A. Molecular armory or niche factors: Virulence determinants of Corynebacterium species. FEMS Microbiol. Lett. 2015, 362, fnv185. [Google Scholar] [CrossRef] [PubMed]
- Antunes, C.A.; dos Santos, L.S.; Hacker, E.; Köhler, S.; Bösl, K.; Ott, L.; de Luna, M.D.; Hirata, R., Jr.; Azevedo, V.A.; Mattos-Guaraldi, A.L.; et al. Characterization of DIP0733, a multi-functional virulence factor of Corynebacterium diphtheriae. Microbiology 2015, 161, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Dangel, A.; Peters, M.; Mühldorfer, K.; Braune, S.; Eisenberg, T.; Szentiks, C.A.; Rau, J.; Konrad, R.; Hörmansdorfer, S.; et al. Tox-positive Corynebacterium ulcerans in hedgehogs, Germany. Emerg. Microbes Infect 2019, 8, 211–217. [Google Scholar] [CrossRef]
- Abe, S.; Takayama, K.; Kinoshita, S. Taxonomical studies on glutamic acid-producing bacteria. J. Gen. Appl. Microbiol. 1967, 13, 279–301. [Google Scholar] [CrossRef]
- Dias, A.A.; Silva, F.C., Jr.; Pereira, G.A.; Souza, M.C.; Camello, T.C.; Damasceno, J.A.; Pacheco, L.G.; Miyoshi, A.; Azevedo, V.A.; Hirata, R., Jr.; et al. Corynebacterium ulcerans isolated from an asymptomatic dog kept in an animal shelter in the metropolitan area of Rio de Janeiro, Brazil. Vector Borne Zoonotic Dis. 2010, 10, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer. 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Knoppová, M.; Phensaijai, M.; Veselý, M.; Zemanová, M.; Nesvera, J.; Pátek, M. Plasmid vectors for testing in vivo promoter activities in Corynebacterium glutamicum and Rhodococcus erythropolis. Curr. Microbiol. 2007, 55, 234–239. [Google Scholar] [CrossRef] [PubMed]




 ), 2 h (
), 2 h (  ), 8 h (
), 8 h (  ), 14 h (
), 14 h (  ), and 20 h (
), and 20 h (  ) are shown. Data shown are the mean values of at least three independent biological replicates each performed in triplicate ± standard deviations.
), 2 h ( ), 8 h ( ), 14 h ( ), and 20 h ( ) are shown. Data shown are the mean values of at least three independent biological replicates each performed in triplicate ± standard deviations.
) are shown. Data shown are the mean values of at least three independent biological replicates each performed in triplicate ± standard deviations.
), 2 h ( ), 8 h ( ), 14 h ( ), and 20 h ( ) are shown. Data shown are the mean values of at least three independent biological replicates each performed in triplicate ± standard deviations.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Description/Genotype | Reference/Source |
| C. glutamicum ATCC 13032 | type-strain, non-pathogenic | [37] |
| C. diphtheriae HC04 | Isolated from a 7 year old female with a fatal endocarditic infection, wild-type, non-toxigenic (tox−) | [13,26] |
| C. ulcerans 809 | Isolated from an 80 year old woman with a fatal pulmonary infection, non-toxigenic (tox−) | [38] |
| Cell lines | Description | Reference/Source |
| THP-1 | Human leukemic monocytic cells | [39] |
| THP1-Blue NF-κB | THP-1 cells with stable integrated NF-κB inducible SEAP reporter construct | Invivogen |
| Plasmids | Description | Reference/Source |
| pEPR1-p45gfp | P45, gfpuv, KmR, rep, per, T1, T2 | [40] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weerasekera, D.; Hahn, J.; Herrmann, M.; Burkovski, A. Induction of Necrosis in Human Macrophage Cell Lines by Corynebacterium diphtheriae and Corynebacterium ulcerans Strains Isolated from Fatal Cases of Systemic Infections. Int. J. Mol. Sci. 2019, 20, 4109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174109
Weerasekera D, Hahn J, Herrmann M, Burkovski A. Induction of Necrosis in Human Macrophage Cell Lines by Corynebacterium diphtheriae and Corynebacterium ulcerans Strains Isolated from Fatal Cases of Systemic Infections. International Journal of Molecular Sciences. 2019; 20(17):4109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174109
Chicago/Turabian StyleWeerasekera, Dulanthi, Jonas Hahn, Martin Herrmann, and Andreas Burkovski. 2019. "Induction of Necrosis in Human Macrophage Cell Lines by Corynebacterium diphtheriae and Corynebacterium ulcerans Strains Isolated from Fatal Cases of Systemic Infections" International Journal of Molecular Sciences 20, no. 17: 4109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174109