Roles of the INO80 and SWR1 Chromatin Remodeling Complexes in Plants


1
Key Laboratory of South China Agricultural Plant Molecular Analysis and Genetic Improvement, Guangdong Provincial Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
2
University of Chinese Academy of Sciences, Chinese Academy of Sciences, Beijing 100049, China
3
College of Light Industry and Food Science, Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China
4
Institute of Plant Biology, National Taiwan University, Taipei 106, Taiwan
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(18), 4591; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184591
Submission received: 21 August 2019
/
Revised: 12 September 2019
/
Accepted: 13 September 2019
/
Published: 17 September 2019
(This article belongs to the Special Issue Chromatin, Epigenetics and Plant Physiology)
Abstract
:Eukaryotic genes are packed into a dynamic but stable nucleoprotein structure called chromatin. Chromatin-remodeling and modifying complexes generate a dynamic chromatin environment that ensures appropriate DNA processing and metabolism in various processes such as gene expression, as well as DNA replication, repair, and recombination. The INO80 and SWR1 chromatin remodeling complexes (INO80-c and SWR1-c) are ATP-dependent complexes that modulate the incorporation of the histone variant H2A.Z into nucleosomes, which is a critical step in eukaryotic gene regulation. Although SWR1-c has been identified in plants, plant INO80-c has not been successfully isolated and characterized. In this review, we will focus on the functions of the SWR1-c and putative INO80-c (SWR1/INO80-c) multi-subunits and multifunctional complexes in Arabidopsis thaliana. We will describe the subunit compositions of the SWR1/INO80-c and the recent findings from the standpoint of each subunit and discuss their involvement in regulating development and environmental responses in Arabidopsis.
1. Introduction
Eukaryotic DNA is packaged with histones to form an inherently stable complex structure known as chromatin. The basic repeating unit of chromatin is the nucleosome, which consists of approximately 146 base pairs of DNA wrapped on a histone octamer containing two molecules each of histone H2A, H2B, H3, and H4 [1]. Therefore, to allow DNA processing, such as DNA replication, repair, and recombination, a dynamic chromatin environment is needed. Two main players, histone post-translational modifications by histone-modifying enzymes and nucleosome positioning by ATP-dependent chromatin-remodeling complexes (CRCs) are involved in the regulation of chromatin dynamics [2,3]. In general, CRCs can destabilize the interactions between histone octamers and DNA by using the energy derived from ATP hydrolysis [4], thus increasing the binding efficiency of transcription factors.
The catalytic subunit (ATPase) of CRCs belongs to the SWI/SNF family [5], which is part of a large superfamily of helicases and translocases called superfamily 2 (SF2). The SWI/SNF family is named after the first identified CRC from S. cerevisiae by the examination of mating type switching (SWI) and sucrose nonfermenting (SNF) mutants [6]. Further studies conducted with Drosophila, mammals, and plants revealed that CRCs widely exist in eukaryotes. Based on the evolutionarily conserved central ATPase subunit, CRCs are currently classified into four subfamilies: SWI/SNF, ISWI (imitation SWI), INO80 (Inositol Requiring 80) and CHD (chromo and DNA-binding domain) [7]. CRCs of the INO80 family include the yeast INO80 complex (INO80-c) and its orthologues in humans and plants, the yeast SWR1 (SWi2/snf2-Related 1) complex (SWR1-c) and its orthologues SRCAP (SNF2-RELATED CBP ACTIVATOR PROTEIN) (human) and SWR1 (Arabidopsis) (Table 1). Among those families, SWR1/INO80-c plays essential roles in DNA repair, checkpoint regulation, DNA replication, telomere maintenance and chromosome segregation [8,9].
Although the molecular mechanisms of SWR1/INO80-c are largely unknown, a large body of genetic and biochemical evidence strongly suggests that SWR1/INO80-c are also present in plants. In this review, we compare the functions and subunit compositions of the SWR1/INO80-c of plants and other organisms including yeasts and mammals. We also discuss the recent investigations underpinning the involvement of SWR1/INO80-c subunits in regulating key plant growth and development processes, as well as environmental responses.
2. Chromatin Remodeling Functions of INO80 and SWR1 CRCs
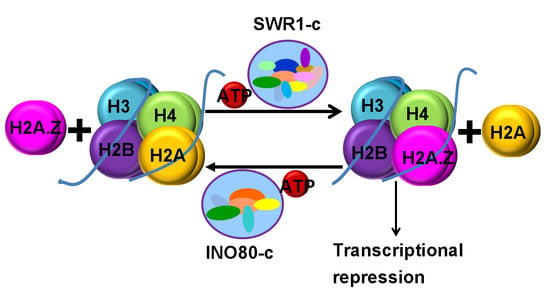
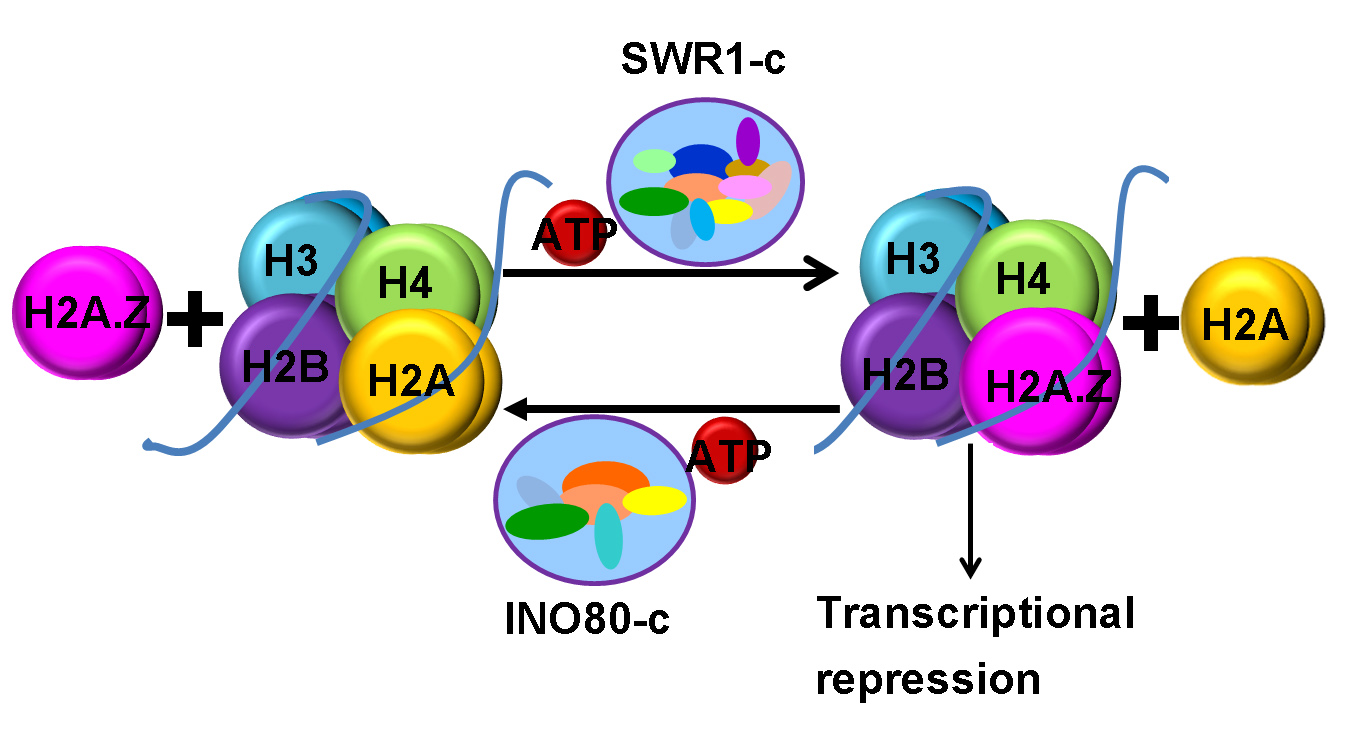
Biochemical analyses indicated that SWR1-c promotes the replacement of nucleosomal H2A/H2B dimers with H2A.Z/H2B [10], while INO80-c catalyzes the reverse dimer exchange reaction [11,12,13]. Further studies demonstrated that H3-K56Ac (H3K56 acetylation) or H3-K56Q (substitution H3-K56 for glutamine) switches the substrate specificity of SWR1-c, inhibiting H2A.Z deposition [13]. Consistently, it was observed that H2A.Z deposition is inhibited by H3-K56Q but not by SWR1-c or INO80-c [14]. However, recent direct EMSA assays indicated that INO80-c catalyzes the replacement of H2A.Z/H2B with H2A/H2B on an H2A.Z/H3-K56Q substrate [15]. The structural analysis of the core INO80-c from the fungus Chaetomium thermophilum showed that the two AAA+ ATPases, Rvb1 (RuvB-like protein 1) and Rvb2, form heterohexamers, acting as a ‘stator’ for the motor (ATPase) and nucleosome-gripping subunits [16]. The ATPase of INO80-c binds to nucleosomal DNA at superhelical location 6 (SHL6), unwraps approximately 15 base pairs, disrupts the H2A–DNA contacts and is poised to pump entry DNA into the nucleosome [16]. Arp5 (Actin-Related Protein 5)–Ies6 (Ino Eighty Subunit 6) binds at SHL2 and SHL 3 acting as a counter grip for the motor on the other side of the H2A–H2B dimer. On the other hand, yeast SWR1-c is also assembled around a heterohexameric core of the Rvb1 and Rvb2 subunits, and the motor subunit binds at SHL2, a position it shares in common with other remodelers [17].
In yeast, the histone variant H2A.Z (Htz1) is enriched in intergenic regions compared with coding regions, and it is involved in the activation of a subset of genes [18]. Meanwhile, incorporation of Htz1 into nucleosomes influences histone modifications and chromatin remodeling, supporting a role for Htz1 at inactive promoters [18]. In Arabidopsis, the high-resolution ChIP-exo assay showed that SWR1 and INO80-c are targeted selectively to the +1 nucleosome of essentially all genes via their subunits [19]. For instance, Swc4 (SWR complex subunit 4) and Swc5 occupy nucleosome positions in the body of genes in addition to +1, while Rvb1 and Swc2 have correlated co-occupancy levels that span a wide range over different promoters [19]. Interestingly, a previous study showed that SWR1/INO80-c may be involved in DNA methylation, since a strong anti-correlation between DNA methylation and H2A.Z deposition was observed [20]. However, the H2A.Z deposition at gene bodies is correlated with a lower transcription level, presumably by destabilizing constitutive gene expression rather than promoting gene body DNA methylation [21]. The genomic binding maps of the subunits of SWR1/INO80-c suggested that many subunits have a rather plastic organization that allows their subunits to exchange between the two complexes [19].
3. The Subunits of INO80 and SWR1 CRCs
yIno80, the ATPase of yeast INO80 complex, was identified in a screen for regulators of phospholipid biosynthesis [22]. Further studies found that yIno80 associates with 14 other proteins including Rvb1, Rvb2, Arp4, Arp5, Arp8, actin1, Taf14 (TATA binding protein-Associated Factor 14), Ies1-6, and Nhp10 (Non-Histone Protein 10) to form a complex (INO80 complex) of 1.2 Mda [22,23] (Table 1). ySwr1, the homolog of yIno80, was subsequently identified and found to catalyze the exchange of H2A for Htz1 [10,24]. Proteomic data indicated that SWR1-c consists of 14 subunits sharing 4 components (Rvb1, Rvb2, Arp4, Act1) with the INO80 complex (Table 1). Interestingly, 4 subunits (Arp4, Yaf9-YEAST ALL1-FUSED GENE FROM CHROMOSOME 9, Swc4, and Act1) of SWR1-c are also part of the NuA4 (Nucleosome Acetyltransferase of H4) complex that is a key regulator of transcription, cellular response to DNA damage and cell cycle control [25,26]. In addition to acetylation of histone H4, histone H2A and histone H2A variant Htz1 [27,28,29], the yeast NuA4 complex also targets non-histone proteins, which encompass metabolism, RNA processing and stress response [30]. In human, Tip60 (Tat-interactive protein 60), the homolog of acetyltransferase subunit of yNuA4 complex (Esa1, Essential for SAS Family Acetyltransferase), was originally identified as an interacting partner of the HIV1-tat protein [31]. Subsequently, the purified Tip60 complex contains at least 16 subunits harboring HAT activity towards histone H4, H2A and H2A.Z in chromatin [32,33,34]. The subunit composition analysis indicates that the Tip60 (NuA4) complex is a fusion form of yeast NuA4 and SWR1 complexes since it also has the enzymatic activity of ATP-dependent H2A.Z-H2B histone dimer exchange [35] (Table 1). A recent report showed that a dynamic merge and separation of NuA4 and SWR1 complexes control cell fate plasticity in C. albicans [36]. Indeed, the YEATS domain of Yaf9 can recognize the acetylation of Eaf1 K173 and mediate the Yaf9-Eaf1 interaction. Furthermore, the reversible acetylation and deacetylation of Eaf1 by Esa1 and Hda1 control the merge and separation of NuA4 and SWR1 complexes, and this regulation is triggered by Brg1 recruitment of Hda1 to chromatin in response to nutritional signals that sustain hyphal elongation [36].
To date, the structural insights into the plant SWR1/INO80-c are largely unknown. Based on sequence homology and protein-protein interacting data, the possible subunits orthologues of yeast and human SWR1/INO80-c were identified in Arabidopsis (Table 1). The plant ATPase of SWR1-c, PHOTOPERIOD-INDEPENDENT EARLY FLOWERING 1 (PIE1), was first characterized as an activator of FLOWERING LOCUS C (FLC) in flowering [37]. Further affinity purification followed by tandem mass spectrometry (TAP-MS) assays showed that SWR1-c contains at least 12 subunits, including RVB2A/B, YAF9A/B, and SWC6 in Arabidopsis [38]. Furthermore, a more recent study identified 11 conserved SWR1-c subunits in Arabidopsis using the conserved SWR1 subunit ARP6 as a bait in tandem affinity purification experiments [39]. Unlike SWR1-c, the compositions of Arabidopsis INO80-c are based on the sequence homology and protein-protein interacting data. The homologue of yINO80 was found in Arabidopsis as a factor in homologous recombination [40]. These data suggested that plants may also have the INO80 complex. Interestingly, recent studies indicated that the NuA4 complex may exist in Arabidopsis, since YAF9A, the yYAF9 homologue (a subunit of yeast NuA4 complex), targets to FLC chromatin and regulates the acetylation of H2A.Z and H4 [41]. The multiplication of the subunits in yeast, human and plant suggest that INO80/SWR1 CRCs are conserved in eukaryotes.
4. Involvement of INO80/SWR1-c in DNA Repair
In yeast, the ino80 and swr1 mutants are hypersensitive to DNA damage-inducing agents [42,43], suggesting a direct role for INO80 and SWR1 CRCs in DNA repair. Indeed, ino80 mutants have defects in both the homologous recombination (HR) and nonhomologous end-joining (NHEJ), two pathways for the repair of DNA double-strand breaks (DSBs), while swr1 mutants have defects in NHEJ alone [44]. Similarly, Arabidopsis INO80 also plays a role in DSBs (Table 2, Figure 1). Under standard growth conditions, the ino80 mutants show a reduction of the HR frequency compared with wild-type plants [40,45]; however, after genotoxic treatment, HR in the mutant increased, accompanied by more DNA double-strand breaks and stronger cellular responses [45]. Further analyses show that INO80 promotes HR downstream of the chaperone NRP1 (NAP1-RELATED PROTEIN 1) after the formation of γ-H2A.X foci during DNA damage repair [46]. Nevertheless, the functional coordination of INO80 and NRP1/2 was also observed in apical meristems during plant growth and development [47]. In addition to INO80, ARP5, the conserved subunit of the INO80-c, also plays key roles in the DNA repair. Indeed, ARPs are highly similar to actin, but they cannot polymerize and do not have ATPase activity. Currently, ARPs are classified into 11 subfamilies, and ARP4-ARP9 are predominantly localized in the nucleus [48]. As dedicated conserved subunits of SWI/SNF and INO80 CRCs, ARPs associate directly with ATPases via the conserved N-terminal HSA (helicase-SANT-associated) domain [49]. The arp5 plants were hypersensitive to DNA-damaging reagents [50], and the transcription levels of DNA repair genes RAD51/RAD54 were up-regulation in mutant plants [51].
Similar to INO80 and ARP5, SWR1-c is also important for DNA repair in Arabidopsis. Since mutations in genes for SWR1-c subunits PIE1, ARP6 and SWC6 cause hypersensitivity to various DNA damaging agents [68]. The reduced DNA repair capacity of those mutants is connected with impaired HR, in contrast with the hyper-recombinogenic phenotype of yeast SWR1 mutants [44,68]. This suggests functional diversification of SWR1-c between lower and higher eukaryotes. The arp5 arp6 double mutants displayed a higher percentage of DNA in tails than that of individual single mutants, suggesting that function of ARP5 in the INO80-c acts independently and/or synergistically with the ARP6-containing SWR1-c in DNA repair [51].
5. Functions of INO80/SWR1-c in Flowering
Successful reproduction in plants requires the coordinated transition from vegetative growth to reproductive development. Complex and intricate gene-regulatory networks of transcription factors and chromatin remodelers guide flowering time and flower development, while integrating both internal and external signals [75]. In Arabidopsis, two MADS box transcription factors MADS AFFECTING FLOWERING 4/5 (MAF4/5), FLC and SHORT VEGETATIVE PHASE (SVP) act as negative regulators of flowering time via directly repressing the expression of the floral pathway integrators FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1) [76,77,78].
The core subunit components of the Arabidopsis SWR1-c, including PIE1, ARP6, ARP4, SWC6/ SERRATED LEAVES AND EARLY FLOWERING (SEF), SWC4 and YAF9A/B have been shown to play important roles in regulating the proper growth and flowering. The Arabidopsis ATPase of SWR1-c, PIE1, was first identified as an activator of FLC [37]. Mutations in PIE1 repress the FLC-mediated delay of flowering, as a result of the presence of FRIGIDA [37]. Further research demonstrated that PIE1 associates with the SWR1-c subunits ARP6 (also known as EARLY IN SHORT DAYS 1 [ESD1] and SUPPRESSOR OF FRI 3 [SUF3]) and SWC6 in repression of flowering [53,60,61,66,67]. Indeed, loss of H2A.Z from chromatin in arp6 and pie1 mutants results in reduced FLC expression and premature flowering, indicating that H2A.Z is required for the expression of FLC [62], which suggested a link between PIE1 and H2A.Z incorporation.
Next to ARP6, ARP4 (present in both INO80-c and SWR1-c, Table 1) has also been shown to act in plant reproductive development. Knockdown of ARP4 results in strong pleiotropic phenotypes such as altered organization of plant organs, early flowering, delayed flower senescence and high levels of sterility, indicating its important roles in plant development [59]. Besides, TAP-MS assays showed that the multiple subunits of SWR1, NuA4, INO80 and SWI/SNF complexes copurified with ARP4, suggesting that ARP4 may be a core subunit of those complexes [56].
In yeast, Swc2 together with Swc6 and Arp6 forms SWR1 complex 2, which handles binding to the histone variant H2A.Z associated with the nucleosome and facilitating histone exchange [79]. To date, three SWCs including SWC2, SWC4 and SWC6 were found in Arabidopsis. SWC4-deficient mutants are embryo-lethal, while SWC4 knockdown lines display acceleration of flowering time, indicating that SWC4 controls post-embryonic processes [38]. However, unlike PIE1, SWC4 represses flowering via repressing FT [38]. Indeed, SWC4 mediates the recruitment of SWR1-c to the target chromatin regions through recognizing AT-rich DNA elements that are over-represented in the promoters of a subset of genes (FT, FUL, IAA19 and ERF9) where H2A.Z incorporation impairs transcription [38]. Intriguingly, like yeast SWC4, recent data demonstrated that SWC4 also associates with YAF9s in Arabidopsis by TAP-MS assays [41]. SWC6 was originally identified from sef mutants. The sef plants present a pleiotropic phenotype including serrated leaves, frequent absence of inflorescence internodes, bushy aspect, and early flowering [53,60]. Like PIE1, SWC6 also activates the transcription of FLC via interacting with SUF4 and FLX, two subunits of the FRIGIDA transcription activator complex [52]. Meanwhile, defects in the splicing of SWC6 pre-mRNA mediated by SKIP (SNW/SKI-INTERACTING PROTEIN) reduce H2A.Z enrichment at FLC, MAF4, and MAF5, resulting in reduced expression of these genes [80]. Interestingly, SWC6 interacts with HAM1 [38], a Myst family histone acetyltransferase of the putative Arabidopsis NuA4 complex (NuA4-c) [81]. Although yeast two-hybrid assays showed that SWC2 interacts with H2A.Z and SWC6, the function of SWC2 in Arabidopsis has not been characterized so far [61].
Yeast Yaf9 (YEAST ALL1-FUSED GENE FROM CHROMOSOME 9) contains an N-terminal YEATS domain and a C-terminal predicted coiled-coil sequence [82]. The YEATS domain proteins are conserved from yeast to human, and function as transcriptional regulators as a part of chromatin-modifying complexes [83]. Indeed, as a selective reader of H3K27ac, the YEATS domain of Yaf9 is essential for deposition of H2A.Z, gene transcription and the DNA-damage response [84]. Deletion of Yaf9 shows reduced telomere-proximal gene expression and sensitivity to DNA-damaging agents [85]. Additionally, Yaf9 has involved chromosome segregation, telomere silencing and response to spindle stress [82]. Moreover, SWR1-c and NuA4-c share 4 subunits (Act1, Arp4, Swc4, Yaf9), indicating a possible functional link between these two complexes in yeast [86]. The Arabidopsis Yaf9 homolog YAF9A acts as a negative regulator of flowering by controlling the H4 acetylation levels in FLC and FT chromatin [41,73]. Furthermore, CCA1 (CIRCADIAN CLOCK ASSOCIATED1) recruits YAF9A to the promoter region of GI (GIGANTEA), resulting in accumulation of H2A.Z and the acetylation of H4, thus increasing GI expression [74]. Indeed, recent data demonstrated that YAF9A and YAF9B have unequally redundant functions, and they regulate flowering time by both FLC-dependent and independent mechanisms [41]. Moreover, YAF9A interacts with HAM1, a putative catalytic subunit of NuA4-c, causing high level acetylation of H2A.Z and H4 of FLC chromatin without affecting the deposition of H2A.Z at the FLC locus [41]. YAF9 proteins also associate with other speculative SWR1-c subunits in Arabidopsis [38,56]. Together, these data suggest functional conservation of the SWR1-c-NuA4-c module in plants and yeast. In addition to flowering, SWR1-c subunit loss-of-function mutants also exhibit pleiotropic flower phenotypes including frequent absence of inflorescence internodes, bushy growth and flowers with altered organ number and size [37,60,62,66,69].
Unlike the SWR1-c, Arabidopsis INO80-c plays a positive role in flowering. INO80, the ATPase of INO80-c is involved in flowering time control via interacting with H2A.Z. Indeed, INO80 binds within the gene body, enhances H2A.Z enrichment and maintains low expression levels of the key flowering repressor genes FLC and MAF4/5, thus ino80 mutants displaying late flowering phenotype [45]. Recent analysis demonstrated that INO80 interacts with ARP5 and acts in concert with ARP5 during plant cellular proliferation and replication stress response [51]. Nevertheless, ARP5 is not required for INO80-mediated control of flowering time and related transcriptional regulation of FLC and MAF4/MAF5 [51]. Taken together, these data indicated that SWR1-c and INO80-c may play an opposite role in flowering, which is consistent with the biochemical activity of SWR1-c and INO80-c.
6. Functions of INO80/SWR1-c in Immunity Response
Plants need to precisely coordinate defense with growth and development to optimize environmental conditions. A previous study showed that 65% of the differentially expressed genes in hta9 hta11 mutants (loss of two major H2A.Z encoding genes out of the possible three in Arabidopsis) were also misregulated in pie1 plants, and the majority of misregulated genes were related to systemic acquired resistance (SAR) [63]. These data indicated that SWR1-c plays a role in maintaining a repressive state of the SAR response. However, further phenotypic analysis showed that pie1, swc6, and hta9 hta11 mutants displayed more macroscopic disease symptoms and increased susceptibility toward Pst DC3000 compared with the wild-type, whereas the arp6 mutant showed increased resistance [57]. These observations suggest that H2A.Z, PIE1, and SWC6 are essential for basal resistance in Arabidopsis, whereas ARP6 has an opposite function. Moreover, although the arp6 and swc6 mutants have similar morphological and developmental phenotypes (early flowering and serrated leaves), and ARP6 and SWC6 have been shown to physically interact in Arabidopsis [53,60], the RNA-seq data did not show a strong correlation in these mutants [57]. This observation is consistent with the defense phenotypes, suggesting that ARP6 and SWC6 could have distinct functions in Arabidopsis.
To data, little is known about INO80-c in plant immunity response. However, Arabidopsis RVB1/RIN1 (RESISTANCE TO PSEUDOMONAS SYRINGAE PV MACULICOLA INTERACTOR 1), the common subunit of INO80/SWR1-c, interacts with both RPM1 (CC-NB-LRR) and RPP5 (TIR-NB-LRR) proteins to negatively regulate the expression of disease resistance (R) genes [58]. Indeed, RVB1 belongs to the members of a larger family of proteins known as the AAA+ class chaperone-like ATPases [87]. In bacteria, Rvb1 and Rvb2 form a double hexamer around Holliday junctions to promote their migration during homologous recombination in DSB repair [88]. In yeast, Rvb1 and Rvb2 are present in both SWR1/INO80-c and are homologous to Arabidopsis RVB1and RVB2A/B. RVB1 is essential for Arabidopsis development during both female gametophyte and sporophyte development [58]. Meanwhile, ectopic expression of RVB1 in yeast yRvb1 deletion alleles can rescue yeast viability [58]. Intriguingly, although TAP-MS experiments showed that RVB1 may not be the subunit of Arabidopsis SWR1-c [39], the interaction between RVB1 and YAF9A still suggested that RVB1 associates with SWR1-c [41]. Compared with yeast, little is known about RVB2A/B functions in plants. Recent data showed that RVB2A/B physically interact with the EAF1 subunit of the NuA4 complex [56], which is a homolog of yeast Eaf1. Further research is required to investigate the molecular mechanism of how INO80-c is involved in plant defense.
7. Involvement of INO80/SWR1-c in the Regulation of MicroRNA Expression
MicroRNAs (miRNAs) repress protein production post-transcriptionally. In flowering plants, modules of miRNAs and their target transcription factors, such as the miR156-SPLs/miR172-AP2 LIKEs and miR319-TCPs/miR164-CUCs modules, control diverse developmental processes including phase transitions, leaf shape, and floral organ identity [89,90]. Previous studies suggest that SWR1-c contributes to the fine control of plant development by generating a balance between miRNAs and target mRNAs at the transcriptional level. In the mutants of arp6, sef, and pie1, miR156 and miR164 levels are reduced at the transcriptional level, which results in the accumulation of target mRNAs and associated morphological changes [54]. Further investigation showed that arp6, sef, and hta9 hta11 mutant plants reduce the expression of MIR156A and MIR156C via facilitating the deposition of H3K4me3, rather than decreasing nucleosome occupancy [55].
The functions of INO80-c in miRNA regulation are largely unknown. However, in mammals, INO80 is a target gene of miR148a in the cancer stem cells (CSCs) of anaplastic thyroid carcinoma (ATC) [91]. The expression of INO80 was upregulated in ATC-CSCs and downregulated upon miRNA148 overexpression. Moreover, overexpression of miRNA148a and knockdown of INO80 acted synergistically to decrease the expression of stem cell marker genes as well as to attenuate stem cell-specific properties including the ability to form tumors [91]. These results suggested that in addition to miRNA regulation, miRNA may also affect the functions of INO80-c or SWR1-c.
8. Other Functions of Core Subunits of INO80/SWR1-c
In addition to the functions of INO80/SWR1-c described above, the core subunits of INO80/SWR1-c have other roles in plants development. For instance, the accumulation of anthocyanin in H2A.Z deposition-deficient mutants such as pie1 is associated with increased H3K4me3 of anthocyanin biosynthetic genes, which is negatively associated with the presence of H2A.Z [64]. These results reveal an antagonistic relationship between H2A.Z and H3K4me3 in the regulation of anthocyanin biosynthesis genes. However, genome-wide occupancy assays of H2A.Z, H3K4me3 and H3K27me3 showed that H2A.Z preferentially associates with H3K4me3 at promoters and H3K27me3 at enhancers in Arabidopsis inflorescence. Moreover, H2A.Z represses enhancer activity by promoting H3K27me3 and preventing H3K4me3 histone modifications [92]. A recent study demonstrated that H3K27me3-enriched chromatin is dependent on the prior action of PIE1 and H2A.Z. Indeed, the H2A.Z- and H3K27me3-enriched chromatin is subsequently stabilized during transcription by the CHD remodeler PICKLE and the histone methyltransferase CURLY LEAF [65]. Collectively, these data suggested that there is a cross-talk between H2A.Z deposition and histone modifications.
Moreover, ARP6 also regulates the female meiosis of prophase I by activating the expression of meiotic recombination related genes such as DMC1 (DISRUPTED MEIOTIC cDNA1) [69]. Subsequent research revealed that ARP6 associates with SWR1-c and promotes the expression of WRKY28 through H2A.Z deposition in megaspore mother cells (MMC) [70]. Intriguingly, arp6 null alleles exhibit a delayed germination rate in osmotic stress conditions, suggesting a positive role of ARP6 in osmotic stress responses [71]. Meanwhile, arp6 plants also display an apparent phosphate starvation response (PSR), when grown in a phosphate replete medium, since ARP6 is required for proper H2A.Z deposition at a number of PSR genes [72]. Moreover, overexpressing ARP6s from other plant species such as Physcomitrella patens and rice (Oryza sativa) could rescue the early flowering phenotype of Arabidopsis arp6 mutants, suggesting that the function of ARP6 is conserved in plants [93].
9. Concluding Remarks and Future Perspectives
The roles of SWR1/INO80-c in nuclear activities are quite diverse ranging from DSB repair to the regulation of gene expression. The mutants of INO80-c and SWR1-c subunits display a variety of organism-specific phenotypes. Moreover, phenotypic differences among the single and double mutants of INO80-c and SWR1-c subunit genes are also observed. For instance, the phenotype of pie1 plants is more severe than that of other SWR1 component mutants, suggesting that PIE1 may be a scaffolding component of different complexes. Indeed, like human p400, PIE1 contains HSA, ATPase, and SANT domains, which are found separately in the Swr1 and Eaf1 proteins, implying that PIE1 may also be an ortholog of p400. TAP-MS data showed that Arabidopsis TRA1 (also a subunit of Spt-Ada-Gcn5-acetyltransferase [SAGA] complex), the homolog of the yeast NuA4 subunit Tra1 and the mammalian Tip 60 subunit TRRAP, also presents in SWR1-c. Collectively, these data suggest that the same subunit (like PIE1) may exist in different distinct complexes. The SWR1-c components also play different roles in resistance to different pathogens. For instance, pie1 and swc6 plants display reduced basal resistance, while loss of ARP6 fucntion results in enhanced resistance [57]. Further in-depth analyses are needed to uncover the functional specificity of SWR1/INO80-c and NuA4-c subunits and their interaction in plants.
Although SWR1/INO80-c are associated with histone variants in vivo [45,63] (Table 2; Figure 1), the precise chromatin-remodeling mechanisms of these complexes in diverse cellular processes are largely unknown. For instances, the mechanism of how SWR1/INO80-c are recruited to their target loci such as FLC is still unclear [37,45,60,61]. It is possible that there is a cross-talk between chromatin modifications and histone variant exchange. In yeast, the NuA4-c mediated acetylation of specific histones in the nucleosome is important for SWR1-c targeting to chromatin and H2A.Z incorporation [28,94]. Moreover, it was shown that Bdf1, a bromodomain-containing subunit of yeast SWR1-c, recruits the complex to chromatin by recognizing acetylated H4 tails [28,95]. Consistently, loss-of-function of Bdf1 results in global reduction of H2A.Z in chromatin [96]. Indeed, the Yaf9 subunit of SWR1-c (also a subunit of NuA4-c) targets histone H3K27ac through its YEATS domain in yeast [84]. The Arabidopsis Yaf9 homologs YAF9A and YAF9B are also novel histone readers that bind to unmodified and acetylated histone H3 [41]. Meanwhile, MBD9, a bromodomain and homeodomains (PHD)-containing unique subunit of Arabidopsis SWR1-c, also recognizes the acetylated histones [97]. Further studies demonstrated that in mbd9 plants, the level of H2A.Z incorporation is significantly reduced, and H2A.Z sites depending on MBD9 has higher levels of H3K9Ac and lower levels of H3K4me3 and H3K36me3 [39]. These data suggest that MBD9, like Bdf1 in yeast, could target SWR1-c via its bromodomain by recognizing acetylated histone marks, such asH3K9Ac. Further research is required to investigate whether acetylated residues on histones can directly recruit SWR1-c to facilitate distinct nuclear processes. In addition, genome-wide binding analyses of INO80/ SWR1-c combined with transcriptome analysis will be useful for the identification of the direct target genes regulated by SWR1/INO80-c, which may provide a comprehensive understanding of how SWR1/INO80-c control plant development and stress responses.
Author Contributions
J.W. and S.G. conceived the idea and wrote the manuscript. X.P. and S.Y. designed and organized the tables and figure. S.Y. and K.W. conceived the idea and revised and organized the manuscript.
Funding
This work was supported by the Guangdong Natural Science Funds for Distinguished Young Scholars (2016A030306047); Pearl River S&T Nova Program of Guangzhou (201610010138); the Youth Innovation Promotion Association, CAS (2017398), and the National Natural Science Foundation of China (No. 31771423). This work was also supported by the Ministry of Science and Technology of Taiwan (105-2311-B-002-012-MY3 and 106-2313-B-002-003) and National Taiwan University (NTU-AS- 108L104310).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Act1 | Actin1 |
| ARP4 | Actin-Related Protein 4 |
| ARP5 | Actin-Related Protein 5 |
| ARP6 | Actin-Related Protein 6 |
| ARP8 | Actin-Related Protein 8 |
| ARP9 | Actin-Related Protein 9 |
| ATC | anaplastic thyroid carcinoma |
| ATP | adenosine triphosphate |
| ATPases | adenosine triphosphatase |
| Bdf1 | Bromodomain factor 1 |
| Brg1 | brahma-related gene 1 |
| CCA1 | CIRCADIAN CLOCK ASSOCIATED1 |
| CHD | chromo and DNA-binding domain |
| CRCs | chromatin-remodeling complexes |
| CSCsCUCs | cancer stem cellsCUP SHAPED COTYLEDONs |
| DMC1 | DISRUPTED MEIOTIC cDNA1 |
| DNA | DeoxyriboNucleic Acid |
| DSBs | DNA double-strand breaks |
| EAF1 | Esa1-associated factor 1 |
| ERF9 | ethylene response factor 9 |
| Esa1 | Essential for SAS Family Acetyltransferase 1 |
| ESD1 | EARLY IN SHORT DAYS 1 |
| FLC | FLOWERING LOCUS C |
| FLX | FLC EXPRESSOR |
| FT | FLOWERING LOCUS T |
| FUL | FRUITFULL |
| GI | GIGANTEA |
| HAM1 | HAIRY MERISTEM 1 |
| Hda1 | Histone deacetylase 1 |
| HR | homologous recombination |
| HSA | helicase/SANT-associated |
| hta11 | HISTONE H2A 11 |
| hta9 | HISTONE H2A 9 |
| IAA19 | AUXIN-RESPONSIVE PROTEIN 19 |
| Ies1-6 | Ino Eighty Subunit 1-6 |
| INO80 | Inositol Requiring 80 |
| INO80-c | INO80 complex |
| ISWI | imitation SWI |
| MAF4/5 | MADS AFFECTING FLOWERING 4/5 |
| MBD9 | Methyl-CpG-binding domain 9 |
| miRNAs | MicroRNAs |
| MMC | megaspore mother cells |
| NHEJ | nonhomologous end-joining |
| Nhp10 | Non-Histone Protein 10 |
| NRP1 | NAP1-RELATED PROTEIN 1 |
| NuA4 | nucleosome acetyltransferase of H4 |
| NuA4-c | NuA4 complex |
| PHD | plant homeodomain |
| PIE1 | PHOTOPERIOD-INDEPENDENT EARLY FLOWERING 1 |
| PSR | phosphate starvation responses |
| RAD51/RAD54 | RADiation sensitive 51/54 |
| RPM1 | RESISTANCE TO P. SYRINGAE PV MACULICOLA 1 |
| RPP5 | RECOGNITION OF PERONOSPORA PARASITICA 5 |
| Rvb1 | RuvB-like protein 1 |
| RVB1/RIN1 | RESISTANCE TO PSEUDOMONAS SYRINGAE PV MACULICOLA INTERACTOR 1 |
| Rvb2 | RuvB-like protein 2 |
| RVB2A/B | RuvB-like protein 2A/B |
| SAGA | Spt-Ada-Gcn5-acetyltransferase |
| SANT | Swi3-Ada2-NCoR-TFIIIB |
| SAR | systemic acquired resistance |
| SEF | SERRATED LEAVES AND EARLY FLOWERING |
| SF2 | superfamily 2 |
| SHL 3 | superhelical location 3 |
| SHL2 | superhelical location 2 |
| SHL6 | superhelical location 6 |
| SKIP | SNW/SKI-INTERACTING PROTEIN |
| SOC1 | SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 |
| SPLs | SQUAMOSA PROMOTER BINDING PROTEIN (SBP)-DOMAIN TRANSCRIPTION FACTORs |
| SRCAP | SNF2-RELATED CBP ACTIVATOR PROTEIN (human) |
| SUF3 | SUPPRESSOR OF FRI 3 |
| SUF4 | SUPPRESSOR OF FRI 4 |
| SVP | SHORT VEGETATIVE PHASE |
| Swc2-6 | SWR complex subunit 2-6 |
| SWI | mating type switching |
| SWR1 | SWi2/snf2-Related 1 |
| SWR1-c | SWR1 complex |
| Taf14 | TATA binding protein-Associated Factor 14 |
| TAP-MS | tandem mass spectrometry |
| TCPs | TEOSINTE BRANCHED 1, CYCLOIDEA AND PCF TRANSCRIPTION FACTORs |
| Tip60 | Tat-interactive protein 60 |
| Tra1 | transfer region 1 |
| TRRAP | transformation/transcription domain-associated protein |
| Yaf9 | YEAST ALL1-FUSED GENE FROM CHROMOSOME 9 |
| YAF9A | YEAST ALL1-FUSED GENE FROM CHROMOSOME 9A |
| YAF9A/B | YEAST ALL1-FUSED GENE FROM CHROMOSOME 9A/B |
| YAF9B | YEAST ALL1-FUSED GENE FROM CHROMOSOME 9B |
References
- Luger, K.; Mader, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 a resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Shilatifard, A. Histone modification: Cause or cog? Trends Genet. 2011, 27, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Jerzmanowski, A. Swi/snf chromatin remodeling and linker histones in plants. Biochim. Biophys. Acta 2007, 1769, 330–345. [Google Scholar] [CrossRef] [PubMed]
- Imbalzano, A.N. Energy-dependent chromatin remodelers: Complex complexes and their components. Crit. Rev. Eukar. Gene 1998, 8, 225–255. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Koonin, E.V. Helicases-amino-acid-sequence comparisons and structure-function-relationships. Curr. Opin. Struc. Biol. 1993, 3, 419–429. [Google Scholar] [CrossRef]
- Abrams, E.; Neigeborn, L.; Carlson, M. Molecular analysis of snf2 and snf5, genes required for expression of glucose-repressible genes in saccharomyces-cerevisiae. Mol. Cell Biol. 1986, 6, 3643–3651. [Google Scholar] [CrossRef] [PubMed]
- Clapier, C.R.; Cairns, B.R. The biology of chromatin remodeling complexes. Annu Rev. Biochem. 2009, 78, 273–304. [Google Scholar] [CrossRef]
- Morrison, A.J.; Shen, X.T. Chromatin remodelling beyond transcription: The ino80 and swr1 complexes. Nat. Rev. Mol. Cell Biol. 2009, 10, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Gerhold, C.B.; Gasser, S.M. Ino80 and swr complexes: Relating structure to function in chromatin remodeling. Trends Cell Biol. 2014, 24, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, G.; Shen, X.; Landry, J.; Wu, W.H.; Sen, S.; Wu, C. Atp-driven exchange of histone h2az variant catalyzed by swr1 chromatin remodeling complex. Science 2004, 303, 343–348. [Google Scholar] [CrossRef]
- Papamichos-Chronakis, M.; Watanabe, S.; Rando, O.J.; Peterson, C.L. Global regulation of h2a.Z localization by the ino80 chromatin-remodeling enzyme is essential for genome integrity. Cell 2011, 144, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Tan, D.; Lakshminarasimhan, M.; Washburn, M.P.; Hong, E.J.; Walz, T.; Peterson, C.L. Structural analyses of the chromatin remodelling enzymes ino80-c and swr-c. Nat. Commun. 2015, 6, 7108. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Radman-Livaja, M.; Rando, O.J.; Peterson, C.L. A histone acetylation switch regulates h2a.Z deposition by the swr-c remodeling enzyme. Science 2013, 340, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ranjan, A.; Wei, D.B.; Wu, C. Comment on “a histone acetylation switch regulates h2a.Z deposition by the swr-c remodeling enzyme”. Science 2016, 353, 358. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Peterson, C.L. Response to comment on “a histone acetylation switch regulates h2a.Z deposition by the swr-c remodeling enzyme”. Science 2016, 353, 358. [Google Scholar] [CrossRef] [PubMed]
- Eustermann, S.; Schall, K.; Kostrewa, D.; Lakomek, K.; Strauss, M.; Moldt, M.; Hopfner, K.P. Structural basis for atp-dependent chromatin remodelling by the ino80 complex. Nature 2018, 556, 386–390. [Google Scholar] [CrossRef]
- Willhoft, O.; Ghoneim, M.; Lin, C.L.; Chua, E.Y.D.; Wilkinson, M.; Chaban, Y.; Ayala, R.; McCormack, E.A.; Ocloo, L.; Rueda, D.S.; et al. Structure and dynamics of the yeast swr1-nucleosome complex. Science 2018, 362, eaat7716. [Google Scholar] [CrossRef]
- Li, B.; Pattenden, S.G.; Lee, D.; Gutierrez, J.; Chen, J.; Seidel, C.; Gerton, J.; Workman, J.L. Preferential occupancy of histone variant h2az at inactive promoters influences local histone modifications and chromatin remodeling. Proc. Natl. Acad. Sci. USA 2005, 102, 18385–18390. [Google Scholar] [CrossRef]
- Yen, K.; Vinayachandran, V.; Pugh, B.F. Swr-c and ino80 chromatin remodelers recognize nucleosome-free regions near +1 nucleosomes. Cell 2013, 154, 1246–1256. [Google Scholar] [CrossRef]
- Zilberman, D.; Coleman-Derr, D.; Ballinger, T.; Henikoff, S. Histone h2a.Z and DNA methylation are mutually antagonistic chromatin marks. Nature 2008, 456, 125–129. [Google Scholar] [CrossRef]
- Coleman-Derr, D.; Zilberman, D. Deposition of histone variant h2a.Z within gene bodies regulates responsive genes. PLoS Genet. 2012, 8, e1002988. [Google Scholar] [CrossRef] [PubMed]
- Ebbert, R.; Birkmann, A.; Schuller, H.J. The product of the snf2/swi2 paralogue ino80 of saccharomyces cerevisiae required for efficient expression of various yeast structural genes is part of a high-molecular-weight protein complex. Mol. Microbiol. 1999, 32, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Mizuguchi, G.; Hamiche, A.; Wu, C. A chromatin remodelling complex involved in transcription and DNA processing. Nature 2000, 406, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.H.; Wu, C.H.; Ladurner, A.; Mizuguchi, G.; Wei, D.; Xiao, H.; Luk, E.; Ranjan, A.; Wu, C. N terminus of swr1 binds to histone h2az and provides a platform for subunit assembly in the chromatin remodeling complex. J. Biol. Chem. 2009, 284, 6200–6207. [Google Scholar] [CrossRef] [PubMed]
- Allard, S.; Utley, R.T.; Savard, J.; Clarke, A.; Grant, P.; Brandl, C.J.; Pillus, L.; Workman, J.L.; Cote, J. Nua4, an essential transcription adaptor/histone h4 acetyltransferase complex containing esa1p and the atm-related cofactor tra1p. Embo J. 1999, 18, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Doyon, Y.; Cote, J. The highly conserved and multifunctional nua4 hat complex. Curr. Opin. Genet. Dev. 2004, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, L.; Lambert, J.P.; Gerdes, M.; Al-Madhoun, A.S.; Skerjanc, I.S.; Figeys, D.; Baetz, K. Functional dissection of the nua4 histone acetyltransferase reveals its role as a genetic hub and that eaf1 is essential for complex integrity. Mol. Cell Biol. 2008, 28, 2244–2256. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.; Auger, A.; Monnet-Saksouk, J.; Brodeur, J.; Piquet, S.; Cramet, M.; Bouchard, N.; Lacoste, N.; Utley, R.T.; Gaudreau, L.; et al. Nua4-dependent acetylation of nucleosomal histones h4 and h2a directly stimulates incorporation of h2a.Z by the swr1 complex. J. Biol. Chem. 2010, 285, 15966–15977. [Google Scholar] [CrossRef] [PubMed]
- Babiarz, J.E.; Halley, J.E.; Rine, J. Telomeric heterochromatin boundaries require nua4-dependent acetylation of histone variant h2a.Z in saccharomyces cerevisiae. Gene Dev. 2006, 20, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Lu, J.Y.; Zhang, J.M.; Walter, W.; Dang, W.W.; Wan, J.; Tao, S.C.; Qian, J.; Zhao, Y.M.; Boeke, J.D.; et al. Protein acetylation microarray reveals that nua4 controls key metabolic target regulating gluconeogenesis. Cell 2009, 136, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Kamine, J.; Elangovan, B.; Subramanian, T.; Coleman, D.; Chinnadurai, G. Identification of a cellular protein that specifically interacts with the essential cysteine region of the hiv-1 tat transactivator. Virology 1996, 216, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Ikura, T.; Ogryzko, V.V.; Grigoriev, M.; Groisman, R.; Wang, J.; Horikoshi, M.; Scully, R.; Qin, J.; Nakatani, Y. Involvement of the tip60 histone acetylase complex in DNA repair and apoptosis. Cell 2000, 102, 463–473. [Google Scholar] [CrossRef]
- Legube, G.; Linares, L.K.; Lemercier, C.; Scheffner, M.; Khochbin, S.; Trouche, D. Tip60 is targeted to proteasome-mediated degradation by mdm2 and accumulates after uv irradiation. Embo J. 2002, 21, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Giaimo, B.D.; Ferrante, F.; Vallejo, D.M.; Hein, K.; Gutierrez-Perez, I.; Nist, A.; Stiewe, T.; Mittler, G.; Herold, S.; Zimmermann, T.; et al. Histone variant h2a.Z deposition and acetylation directs the canonical notch signaling response. Nucleic Acids Res. 2018, 46, 8197–8215. [Google Scholar] [CrossRef] [PubMed]
- Auger, A.; Galarneau, L.; Altaf, M.; Nourani, A.; Doyon, Y.; Utley, R.T.; Cronier, D.; Allard, S.; Cote, J. Eaf1 is the platform for nua4 molecular assembly that evolutionarily links chromatin acetylation to atp-dependent exchange of histone h2a variants. Mol. Cell Biol. 2008, 28, 2257–2270. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, W.; Chang, P.; Wu, H.; Liu, H.; Chen, J. Merge and separation of nua4 and swr1 complexes control cell fate plasticity in candida albicans. Cell Discov. 2018, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Noh, Y.S.; Amasino, R.M. Pie1, an iswi family gene, is required for flc activation and floral repression in arabidopsis. Plant Cell 2003, 15, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Zambrano, Á.; Crevillén, P.; Franco-Zorrilla, J.M.; López, J.A.; Santos-González, J.; Roszak, P.; Santos-Gonzalez, J.; Jurado, S.; Vázquez, J.; Köhler, C.; et al. Arabidopsis swc4 binds DNA and recruits the swr1 complex to modulate histone h2a.Z deposition at key regulatory genes. Mol. Plant 2018, 11, 815–832. [Google Scholar] [CrossRef] [PubMed]
- Sijacic, P.; Holder, D.H.; Bajic, M.; Deal, R.B. Methyl-cpg-binding domain 9 (mbd9) is required for h2a.Z incorporation into chromatin at a subset of h2a.Z-enriched regions in the arabidopsis genome. PLoS Genet. 2019, 15, e1008326. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, O.; Benvenuto, G.; Bowler, C.; Molinier, J.; Hohn, B. The ino80 protein controls homologous recombination in arabidopsis thaliana. Mol. Cell 2004, 16, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Crevillén, P.; Gómez-Zambrano, Á.; López, J.A.; Vázquez, J.; Piñeiro, M.; Jarillo, J.A. Arabidopsis yaf9 histone readers modulate flowering time through nua4-complex-dependent h4 and h2a.Z histone acetylation at flc chromatin. NEW Phytol. 2019, 222, 1893–1908. [Google Scholar] [CrossRef] [PubMed]
- Van Attikum, H.; Fritsch, O.; Hohn, B.; Gasser, S.M. Recruitment of the ino80 complex by h2a phosphorylation links atp-dependent chromatin remodeling with DNA double-strand break repair. Cell 2004, 119, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Morrison, A.J.; Highland, J.; Krogan, N.J.; Arbel-Eden, A.; Greenblatt, J.F.; Haber, J.E.; Shen, X. Ino80 and gamma-h2ax interaction links atp-dependent chromatin remodeling to DNA damage repair. Cell 2004, 119, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Van Attikum, H.; Fritsch, O.; Gasser, S.M. Distinct roles for swr1 and ino80 chromatin remodeling complexes at chromosomal double-strand breaks. Embo J. 2007, 26, 4113–4125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cao, L.; Rong, L.; An, Z.; Zhou, W.; Ma, J.; Shen, W.H.; Zhu, Y.; Dong, A. The chromatin-remodeling factor atino80 plays crucial roles in genome stability maintenance and in plant development. Plant J. 2015, 82, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Gao, J.; Ma, J.; Cao, L.; Zhang, C.; Zhu, Y.; Dong, A.; Shen, W.H. Distinct roles of the histone chaperones nap1 and nrp and the chromatin-remodeling factor ino80 in somatic homologous recombination in arabidopsis thaliana. Plant J. 2016, 88, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Ma, J.; Wu, D.; Shen, W.H.; Zhu, Y. Functional coordination of the chromatin-remodeling factor atino80 and the histone chaperones nrp1/2 in inflorescence meristem and root apical meristem. Front. Plant Sci. 2019, 10, 115. [Google Scholar] [CrossRef]
- Muller, J.; Oma, Y.; Vallar, L.; Friederich, E.; Poch, O.; Winsor, B. Sequence and comparative genomic analysis of actin-related proteins. Mol. Biol. Cell 2005, 16, 5736–5748. [Google Scholar] [CrossRef] [PubMed]
- Szerlong, H.; Hinata, K.; Viswanathan, R.; Erdjument-Bromage, H.; Tempst, P.; Cairns, B.R. The hsa domain binds nuclear actin-related proteins to regulate chromatin-remodeling atpases. Nat. Struct. Mol. Biol. 2008, 15, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, M.K.; McKinney, E.C.; Deal, R.B.; Smith, A.P.; Meagher, R.B. Arabidopsis actin-related protein arp5 in multicellular development and DNA repair. Dev. Biol. 2009, 335, 22–32. [Google Scholar] [CrossRef]
- Kang, H.; Zhang, C.; An, Z.; Shen, W.H.; Zhu, Y. Atino80 and atarp5 physically interact and play common as well as distinct roles in regulating plant growth and development. NEW Phytol. 2019, 223, 336–353. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Kim, J.; Hwang, H.J.; Kim, S.; Park, C.; Kim, S.Y.; Lee, I. The frigida complex activates transcription of flc, a strong flowering repressor in arabidopsis, by recruiting chromatin modification factors. Plant Cell 2011, 23, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, A.; Gómez-Zambrano, A.; López-González, L.; Piñeiro, M.; Jarillo, J.A. Mutations in the arabidopsis swc6 gene, encoding a component of the swr1 chromatin remodelling complex, accelerate flowering time and alter leaf and flower development. J. Exp. Bot. 2008, 59, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Kim, J.; Muller, S.Y.; Oh, M.; Underwood, C.; Henderson, I.; Lee, I. Regulation of microrna-mediated developmental changes by the swr1 chromatin remodeling complex. Plant Physiol. 2016, 171, 1128–1143. [Google Scholar] [PubMed]
- Xu, M.L.; Leichty, A.R.; Hu, T.Q.; Poethig, R.S. H2a.Z promotes the transcription of mir156a and mir156c in arabidopsis by facilitating the deposition of h3k4me3. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Bieluszewski, T.; Galganski, L.; Sura, W.; Bieluszewska, A.; Abram, M.; Ludwikow, A.; Ziolkowski, P.A.; Sadowski, J. Ateaf1 is a potential platform protein for arabidopsis nua4 acetyltransferase complex. BMC Plant Biol. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Berriri, S.; Gangappa, S.N.; Kumar, S.V. Swr1 chromatin-remodeling complex subunits and h2a.Z have non-overlapping functions in immunity and gene regulation in arabidopsis. Mol. Plant 2016, 9, 1051–1065. [Google Scholar] [CrossRef] [PubMed]
- Holt, B.F.; Boyes, D.C.; Ellerstrom, M.; Siefers, N.; Wiig, A.; Kauffman, S.; Grant, M.R.; Dangl, J.L. An evolutionarily conserved mediator of plant disease resistance gene function is required for normal arabidopsis development. Dev. Cell 2002, 2, 807–817. [Google Scholar] [CrossRef]
- Kandasamy, M.K.; Deal, R.B.; McKinney, E.C.; Meagher, R.B. Silencing the nuclear actin-related protein atarp4 in arabidopsis has multiple effects on plant development, including early flowering and delayed floral senescence. Plant J. 2005, 41, 845–858. [Google Scholar] [CrossRef]
- March-Díaz, R.; García-Domínguez, M.; Florencio, F.J.; Reyes, J.C. Sef, a new protein required for flowering repression in arabidopsis, interacts with pie1 and arp6. Plant Physiol. 2007, 143, 893–901. [Google Scholar] [CrossRef]
- Choi, K.; Park, C.; Lee, J.; Oh, M.; Noh, B.; Lee, I. Arabidopsis homologs of components of the swr1 complex regulate flowering and plant development. Development 2007, 134, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
- Deal, R.B.; Topp, C.N.; McKinney, E.C.; Meagher, R.B. Repression of flowering in arabidopsis requires activation of flowering locus c expression by the histone variant h2a.Z. Plant Cell 2007, 19, 74–83. [Google Scholar] [CrossRef] [PubMed]
- March-Díaz, R.; García-Domínguez, M.; Lozano-Juste, J.; León, J.; Florencio, F.J.; Reyes, J.C. Histone h2a.Z and homologues of components of the swr1 complex are required to control immunity in arabidopsis. Plant J. 2008, 53, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Zhang, M.; Chai, M.; He, Q.; Huang, X.; Zhao, L.; Qin, Y. Epigenetic regulation of anthocyanin biosynthesis by an antagonistic interaction between h2a.Z and h3k4me3. New Phytol. 2019, 221, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.; Bishop, B.; Ho, K.K.; Huang, R.; Jia, W.; Zhang, H.; Pascuzzi, P.E.; Deal, R.B.; Ogas, J. The chromatin remodelers pkl and pie1 act in an epigenetic pathway that determines h3k27me3 homeostasis in arabidopsis. Plant Cell 2018, 30, 1337–1352. [Google Scholar] [CrossRef]
- Choi, K.; Kim, S.; Kim, S.Y.; Kim, M.; Hyun, Y.; Lee, H.; Choe, S.; Kim, S.G.; Michaels, S.; Lee, I. Suppressor of frigida3 encodes a nuclear actin-related protein6 required for floral repression in arabidopsis. Plant Cell 2005, 17, 2647–2660. [Google Scholar] [CrossRef]
- Martin-Trillo, M.; Lazaro, A.; Poethig, R.S.; Gomez-Mena, C.; Pineiro, M.A.; Martinez-Zapater, J.M.; Jarillo, J.A. Early in short days 1 (esd1) encodes actin-related protein 6 (atarp6), a putative component of chromatin remodelling complexes that positively regulates flc accumulation in arabidopsis. Development 2006, 133, 1241–1252. [Google Scholar] [CrossRef]
- Rosa, M.; Von Harder, M.; Cigliano, R.A.; Schlogelhofer, P.; Mittelsten Scheid, O. The arabidopsis swr1 chromatin-remodeling complex is important for DNA repair, somatic recombination, and meiosis. Plant Cell 2013, 25, 1990–2001. [Google Scholar] [CrossRef]
- Qin, Y.; Zhao, L.H.; Skaggs, M.I.; Andreuzza, S.; Tsukamoto, T.; Panoli, A.; Wallace, K.N.; Smith, S.; Siddiqi, I.; Yang, Z.B.; et al. Actin-related protein6 regulates female meiosis by modulating meiotic gene expression in arabidopsis. Plant Cell 2014, 26, 1612–1628. [Google Scholar] [CrossRef]
- Zhao, L.; Cai, H.; Su, Z.; Wang, L.; Huang, X.; Zhang, M.; Chen, P.; Dai, X.; Zhao, H.; Palanivelu, R.; et al. Klu suppresses megasporocyte cell fate through swr1-mediated activation of wrky28 expression in arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E526–E535. [Google Scholar] [CrossRef]
- Sura, W.; Kabza, M.; Karlowski, W.M.; Bieluszewski, T.; Kus-Slowinska, M.; Paweloszek, L.; Sadowski, J.; Ziolkowski, P.A. Dual role of the histone variant h2a.Z in transcriptional regulation of stress-response genes. Plant Cell 2017, 29, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.P.; Jain, A.; Deal, R.B.; Nagarajan, V.K.; Poling, M.D.; Raghothama, K.G.; Meagher, R.B. Histone h2a.Z regulates the expression of several classes of phosphate starvation response genes but not as a transcriptional activator. Plant Physiol. 2010, 152, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Zacharaki, V.; Benhamed, M.; Poulios, S.; Latrasse, D.; Papoutsoglou, P.; Delarue, M.; Vlachonasios, K.E. The arabidopsis ortholog of the yeats domain containing protein yaf9a regulates flowering by controlling h4 acetylation levels at the flc locus. Plant Sci. 2012, 196, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.H.; Wang, S.L.; Zhang, F.; Zheng, H.; Liu, Y.N.; Huang, T.T.; Ding, Y. Phosphorylation of histone h2a at serine 95: A plant-specific mark involved in flowering time regulation and h2a.Z deposition. Plant Cell 2017, 29, 2197–2213. [Google Scholar] [CrossRef] [PubMed]
- Wils, C.R.; Kaufmann, K. Gene-regulatory networks controlling inflorescence and flower development in arabidopsis thaliana. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yoo, S.J.; Park, S.H.; Hwang, I.; Lee, J.S.; Ahn, J.H. Role of svp in the control of flowering time by ambient temperature in arabidopsis. Genes Dev. 2007, 21, 397–402. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.; Shen, L.; Wu, Y.; Chen, H.; Robertson, M.; Helliwell, C.A.; Ito, T.; Meyerowitz, E.; Yu, H. A repressor complex governs the integration of flowering signals in arabidopsis. Dev. Cell 2008, 15, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Putterill, J.; Laurie, R.; Macknight, R. It’s time to flower: The genetic control of flowering time. Bioessays 2004, 26, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Kobor, M.S.; Venkatasubrahmanyam, S.; Meneghini, M.D.; Gin, J.W.; Jennings, J.L.; Link, A.J.; Madhani, H.D.; Rine, J. A protein complex containing the conserved swi2/snf2-related atpase swr1p deposits histone variant h2a.Z into euchromatin. Plos Biol. 2004, 2, 587–599. [Google Scholar] [CrossRef]
- Cui, Z.; Tong, A.; Huo, Y.; Yan, Z.; Yang, W.; Yang, X.; Wang, X.X. Skip controls flowering time via the alternative splicing of sef pre-mrna in arabidopsis. BMC Biol. 2017, 15, 80. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, H.; Xing, L.; Xu, S.; Liu, H.; Chong, K.; Xu, Y. Requirement of histone acetyltransferases ham1 and ham2 for epigenetic modification of flc in regulating flowering in arabidopsis. J. Plant Physiol. 2013, 170, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Le Masson, I.; Yu, D.Y.; Jensen, K.; Chevalier, A.; Courbeyrette, R.; Boulard, Y.; Smith, M.M.; Mann, C. Yaf9, a novel nua4 histone acetyltransferase subunit, is required for the cellular response to spindle stress in yeast. Mol. Cell Biol. 2003, 23, 6086–6102. [Google Scholar] [CrossRef] [PubMed]
- Schulze, J.M.; Wang, A.Y.; Kobor, M.S. Yeats domain proteins: A diverse family with many links to chromatin modification and transcription. Biochem. Cell Biol. 2009, 87, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.J.; Ahmad, S.; Vann, K.R.; Andrews, F.H.; Mayo, Z.A.; Bourriquen, G.; Bridgers, J.B.; Zhang, J.Y.; Strahl, B.D.; Cote, J.; et al. Yaf9 subunit of the nua4 and swr1 complexes targets histone h3k27ac through its yeats domain. Nucleic Acids Res. 2018, 46, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Richardson, D.O.; Roberts, D.N.; Utley, R.; Erdjument-Bromage, H.; Tempst, P.; Cote, J.; Cairns, B.R. The yaf9 component of the swr1 and nua4 complexes is required for proper gene expression, histone h4 acetylation, and htz1 replacement near telomeres. Mol. Cell Biol. 2004, 24, 9424–9436. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.Y.T.; Levesque, N.; Kobor, M.S. Nua4 and swr1-c: Two chromatin-modifying complexes with overlapping functions and components. Biochem Cell Biol. 2009, 87, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Neuwald, A.F.; Aravind, L.; Spouge, J.L.; Koonin, E.V. Aaa+: A class of chaperone-like atpases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999, 9, 27–43. [Google Scholar] [PubMed]
- Kanemaki, M.; Kurokawa, Y.; Matsu-ura, T.; Makino, Y.; Masani, A.; Okazaki, K.; Morishita, T.; Tamura, T. Tip49b, a new ruvb-like DNA helicase, is included in a complex together with another ruvb-like DNA helicase, tip49a. J. Biol. Chem. 1999, 274, 22437–22444. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Guo, Z.; Li, L. Evolutionary conservation of microrna regulatory programs in plant flower development. Dev. Biol. 2013, 380, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Samad, A.F.A.; Sajad, M.; Nazaruddin, N.; Fauzi, I.A.; Murad, A.M.A.; Zainal, Z.; Ismail, I. Microrna and transcription factor: Key players in plant regulatory network. Front. Plant Sci. 2017, 8, 565. [Google Scholar] [CrossRef]
- Sheng, W.; Chen, Y.; Gong, Y.; Dong, T.; Zhang, B.; Gao, W. Mir-148a inhibits self-renewal of thyroid cancer stem cells via repressing ino80 expression. Oncol. Rep. 2016, 36, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Bai, Y.; Zhao, L.; Dou, X.; Liu, Y.; Wang, L.; Li, Y.; Li, W.; Hui, Y.; Huang, X.; et al. H2a.Z represses gene expression by modulating promoter nucleosome structure and enhancer histone modifications in arabidopsis. Mol. Plant 2017, 10, 1274–1292. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Park, C.; Choi, K.; Lee, D.; Seok, C.; Lee, I. Molecular evolution of actin related protein 6, a component of swr1 complex in arabidopsis. J. Plant Biol. 2016, 59, 467–477. [Google Scholar] [CrossRef]
- Cheng, X.; Auger, A.; Altaf, M.; Drouin, S.; Paquet, E.; Utley, R.T.; Robert, F.; Cote, J. Eaf1 links the nua4 histone acetyltransferase complex to htz1 incorporation and regulation of purine biosynthesis. Eukaryot Cell 2015, 14, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Ladurner, A.G.; Inouye, C.; Jain, R.; Tjian, R. Bromodomains mediate an acetyl-histone encoded antisilencing function at heterochromatin boundaries. Mol. Cell 2003, 11, 365–376. [Google Scholar] [CrossRef]
- Durant, M.; Pugh, B.F. Nua4-directed chromatin transactions throughout the saccharomyces cerevisiae genome. Mol. Cell Biol. 2007, 27, 5327–5335. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J. Lysine acetylation and the bromodomain: A new partnership for signaling. Bioessays 2004, 26, 1076–1087. [Google Scholar] [CrossRef]
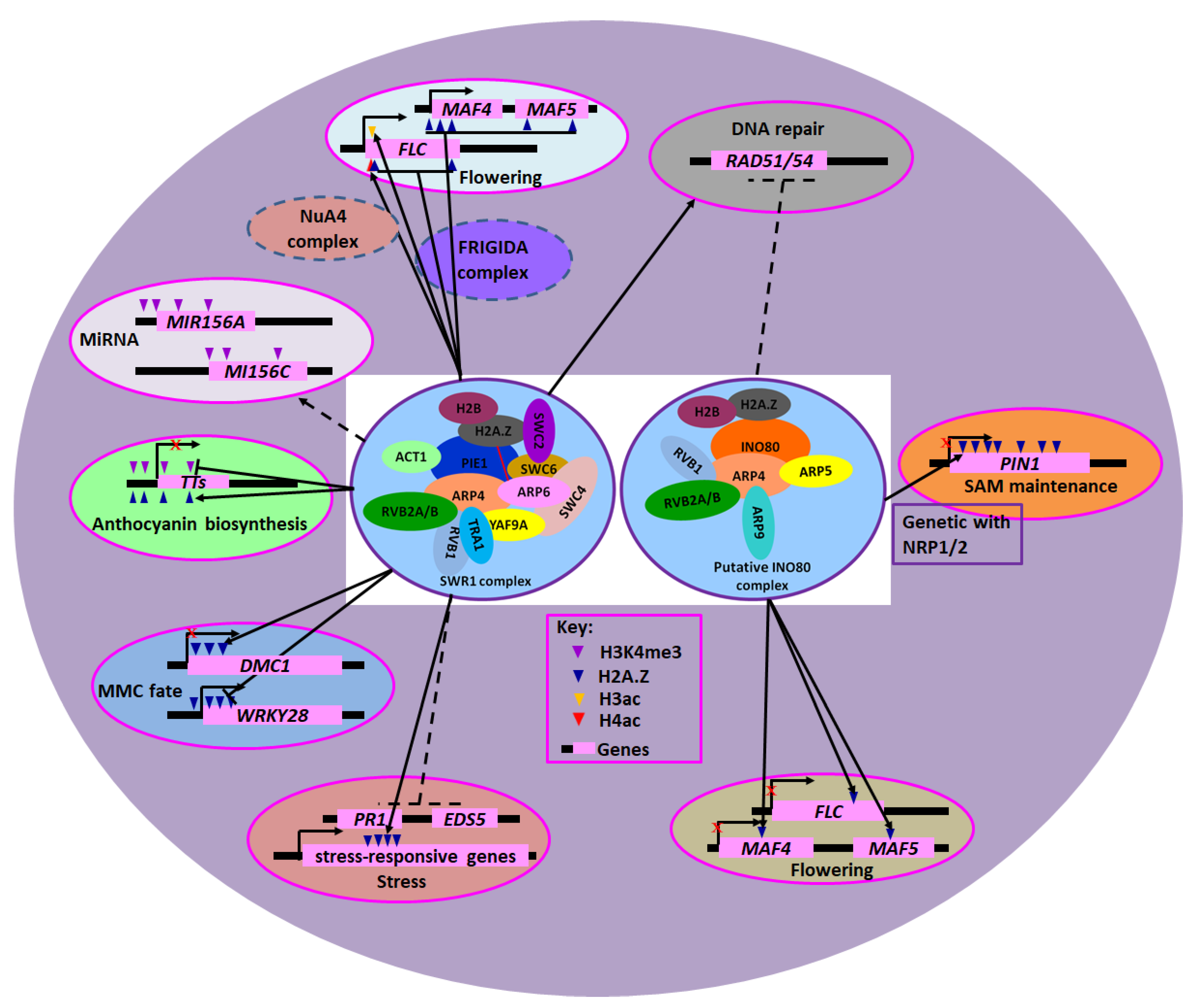
Figure 1.
INO80 and SWR1 Chromatin Remodeling Complexes in Cellular Processes and Plant Development. The compositions of putative INO80 and SWR1 complex are indicated. The purple oval represents the nucleus of plant cells. The overlapped ovals and red lines represent interacting subunits of the INO80 and SWR1 complex. Lines with perpendicular bars and red cross denote repression, while arrows indicate enhancement or activation. Broken lines indicate relationships not proven to be direct.
Figure 1.
INO80 and SWR1 Chromatin Remodeling Complexes in Cellular Processes and Plant Development. The compositions of putative INO80 and SWR1 complex are indicated. The purple oval represents the nucleus of plant cells. The overlapped ovals and red lines represent interacting subunits of the INO80 and SWR1 complex. Lines with perpendicular bars and red cross denote repression, while arrows indicate enhancement or activation. Broken lines indicate relationships not proven to be direct.


{kind=link}
{kind=link}
Table 1.
Compositions of INO80, SWR1 and NuA4 Complexes in Arabidopsis.
| Complex | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Organism | Yeast | Human | Arabidopsis | ||||||
| Family and Composition | INO80 | SWR1 | NuA4 | INO80 | SWR1/SRCAP | NuA4/Tip60 | INO80 | SWR1 | NuA4 |
| ATPase or Acetyltransferase | Ino80 | Swr1 | Eaf1 **, Esa1 * | hIno80 | SRCAP | p400 **, Tip60 * | INO80 | PIE1 | HAM1/2 *, EAF1 ** |
| Noncatalytic homologous subunits | Rvb1, Rvb2 | Tip49a, Tip49b | RVB1/RIN1 | ||||||
| RVB2A, RVB2B | |||||||||
| Arp4, Actin1 | BAF53a | ARP4, ACT1 | |||||||
| Arp5, Arp8 | Arp6 | Arp5, Arp8 | Arp6 | Actin | ARP5, ARP9 | ARP6 | |||
| Taf14 | Yaf9 | GAS41 | GAS41/YAF9A, TAF14/YAF9B | ||||||
| Ies2, Ies6 | hIes2, hIes6 | ||||||||
| Swc4/Eaf2 | DMAP1 | SWC4 | |||||||
| Swc2/Vps72 | YL-1 | SWC2 | |||||||
| Swc6/Vps71 | ZnF-HIT1 | SWC6 | |||||||
| Bdf1 | Brd8/TRCp120 | ||||||||
| H2A.Z, H2B | H2A.Z, H2B | H2A.Z, H2B | |||||||
| Tra1 | TRRAP | TRA1 | |||||||
| Epl1 | EPC1 | ||||||||
| Yng2 | ING3 | ING1, ING2 | |||||||
| Eaf3 | MRG15 | MRG1, MRG2 | |||||||
| Eaf5 | |||||||||
| Eaf7 | MRGBP | ||||||||
| Eaf6 | hEaf6 | ||||||||
| Unique | Ies1, Ies3, Ies 4, Ies5, Nhp10 | Swc3,5,7 | Amida, NFRKB, MCRS1, FLJ90652, FLJ20309 | MRGX, FLJ11730, MRGBP, EPC1, EPC-like | MBD9, AL5-7 | ||||
* and ** represent the acetyltransferase and ATPase subunits of NuA4-c, respectively. The subunits of INO80 and NuA4 complexes in Arabidopsis are based on the sequence homology and protein-protein interacting data.
Table 2.
The Functions of core SWR1/INO80-c Subunits in Arabidopsis.
| Gene | Interacting Proteins * | Functions | Reference | |
|---|---|---|---|---|
| The common subunits of INO80/SWR1-c | SWC2 | unknown | ||
| SWC4 | Flowering time control; Leaf cell proliferation and expansion | [38] | ||
| SWC6/SEF | SUF4, FLX, TAF14 | Flowering time control | [52,53] | |
| MicroRNA expression | [54,55] | |||
| HAM1, EAF1 | [38,56] | |||
| Immunity response | [57] | |||
| RVB1/RIN1 | FLX, SUF4, FES1, FRI | [52] | ||
| RPM1, RPP5 | Sporophyte and female gametophyte; Disease resistance | [58] | ||
| RVB2A, RVB2B | EAF1 | [56] | ||
| ARP4 | Multiple effects on plant development | [59] | ||
| Core subunit of SWR1, NuA4, INO80 and SWI/SNF complexes | [56] | |||
| Unique subunits of INO80-c | INO80 | Controls homologous recombination | [40,45,46,51] | |
| Flowering time control | [45,51] | |||
| Apical meristems maintenance | [47] | |||
| ARP5 | organ development; DNA repair | [50,51] | ||
| ARP9 | unknown | |||
| Unique subunits of SWR1-c | PIE1 | Flowering time control | [37,60,61,62] | |
| Immunity response | [57,63] | |||
| MicroRNA expression | [54] | |||
| Anthocyanin biosynthesis | [64] | |||
| Maintenance of H3K27me3 | [65] | |||
| ARP6 | Flowering time control | [61,62,66,67] | ||
| MicroRNA expression | [54,55] | |||
| Immunity response | [57] | |||
| DNA repair | [68] | |||
| Female meiosis regulation; Germ-line specification | [69,70] | |||
| Osmotic stress; Phosphate starvation response | [71,72] | |||
| YAF9A | CCA1, HAM1 | Flowering time control | [41,73,74] | |
| YAF9B |
* The interactions among the core subunits of SWR1/INO80-c are shown in Figure 1.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, J.; Gao, S.; Peng, X.; Wu, K.; Yang, S. Roles of the INO80 and SWR1 Chromatin Remodeling Complexes in Plants. Int. J. Mol. Sci. 2019, 20, 4591. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184591
AMA Style
Wang J, Gao S, Peng X, Wu K, Yang S. Roles of the INO80 and SWR1 Chromatin Remodeling Complexes in Plants. International Journal of Molecular Sciences. 2019; 20(18):4591. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184591
Chicago/Turabian StyleWang, Jianhao, Sujuan Gao, Xiuling Peng, Keqiang Wu, and Songguang Yang. 2019. "Roles of the INO80 and SWR1 Chromatin Remodeling Complexes in Plants" International Journal of Molecular Sciences 20, no. 18: 4591. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184591
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.