Comprehensive Influences of Overexpression of a MYB Transcriptor Regulating Anthocyanin Biosynthesis on Transcriptome and Metabolome of Tobacco Leaves

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Phenotype and Anthocyanin Content of Transgenic Line Overexpressing LrAN2
2.2. Chemical Compounds Difference in Transgenic Lines and Wild-Type (WT)
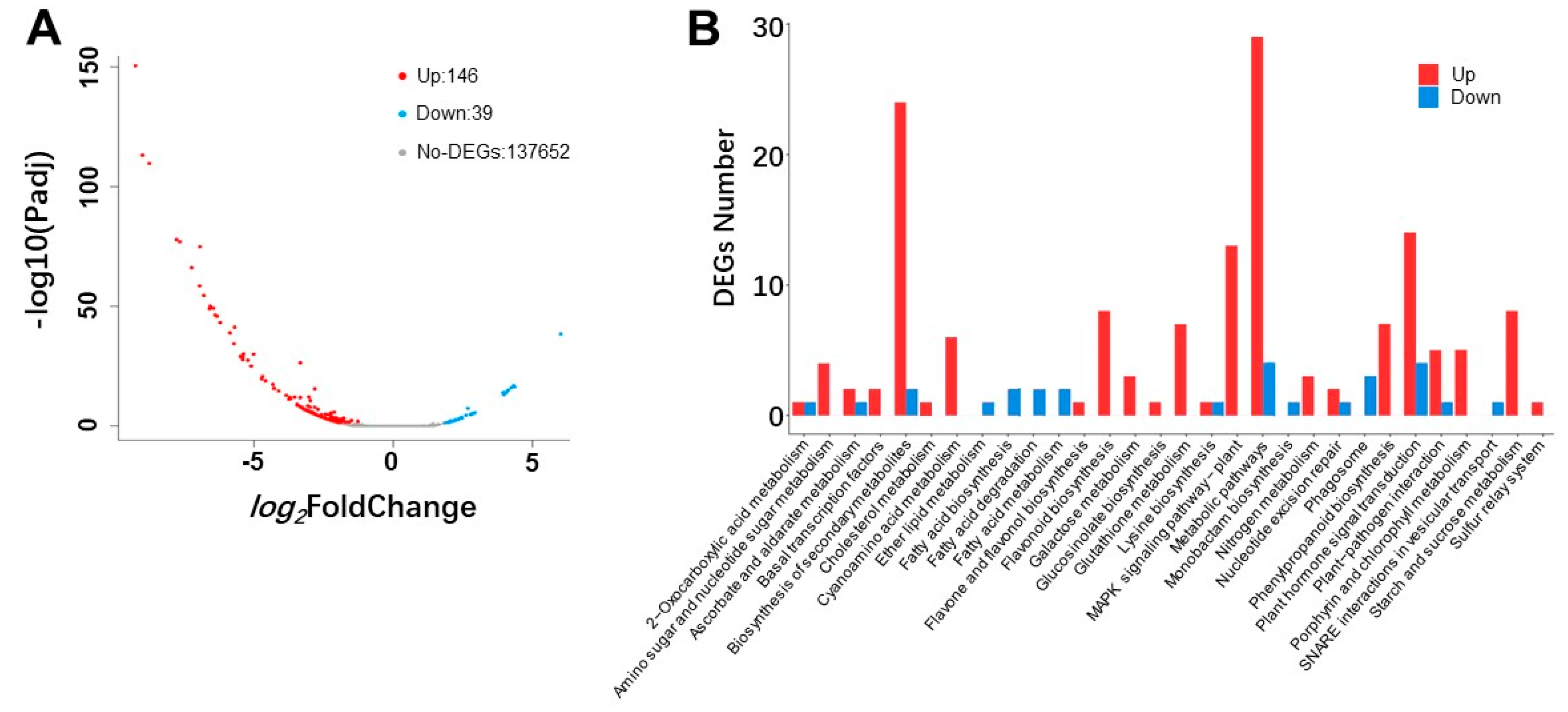
2.3. Differently-Expressed Genes in Transgenic Lines and WT
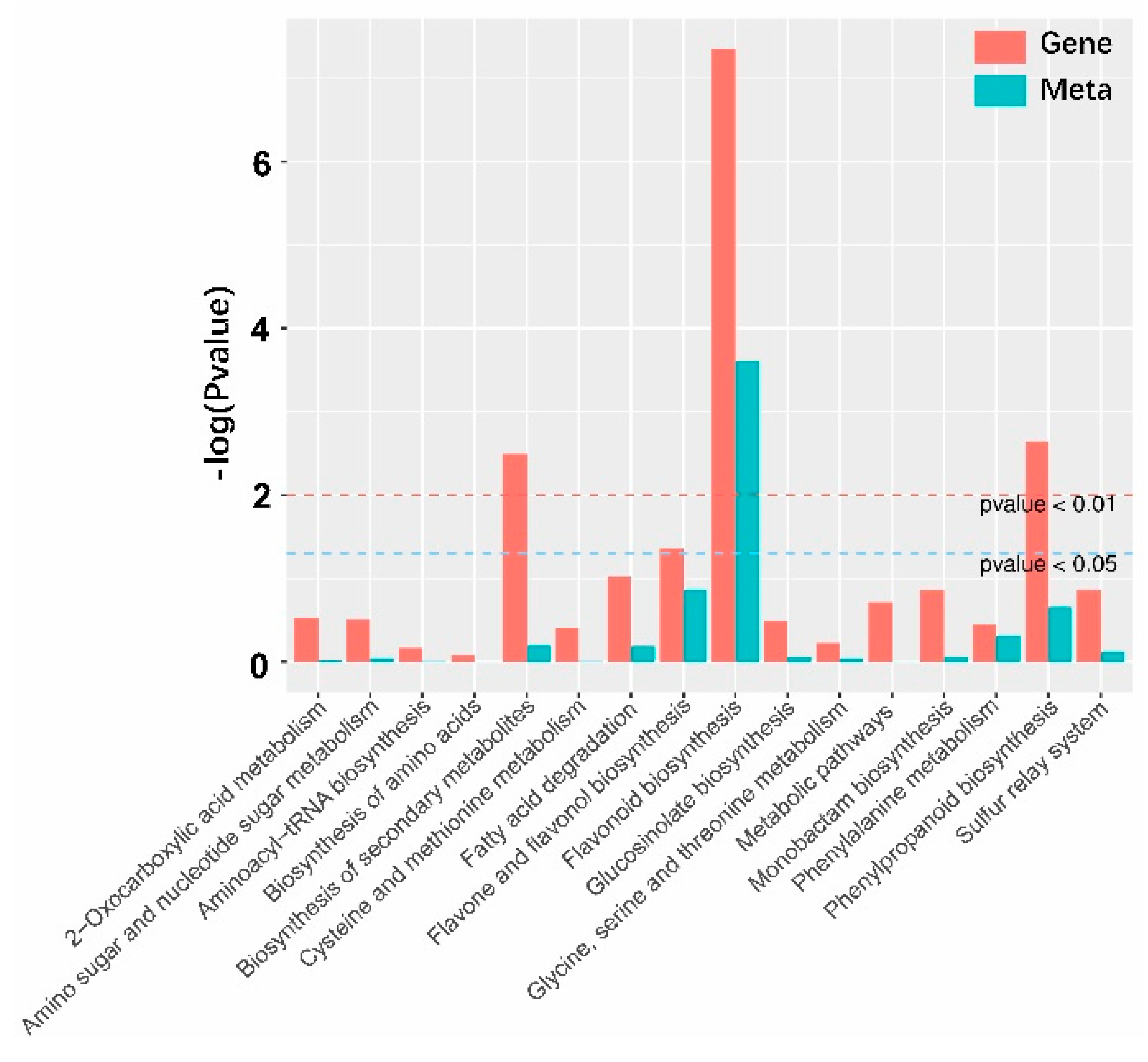
2.4. Conjoint Analysis of Transcriptome and Metabolome
3. Discussion
3.1. Transcriptome Turbulence Was Relatively Simple and Metabolome Influence Was Extensive
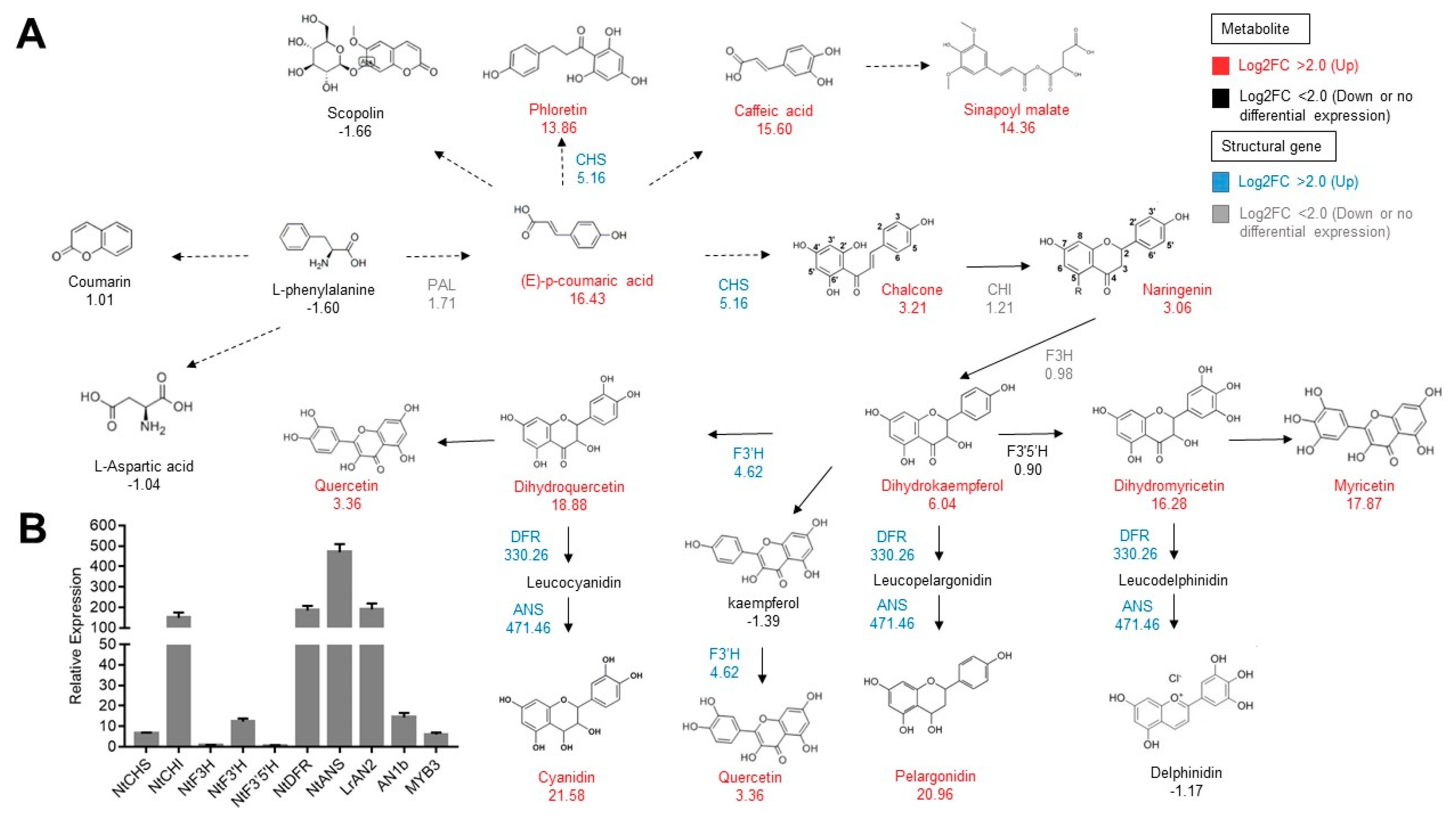
3.2. Overexpression of MYB Transcription Factors Influences Phenylpropanoid Metabolism
3.3. Amino Acid Biosynthesis is Influenced by the Overexpression of MYB Transcription Factor
4. Materials and Methods
4.1. Materials
4.2. Obtaining Transgenic Lines with Overexpression of MYB Transcription Factor
4.3. Anthocyanin Content Measurement
4.4. Analysis of Chemical Contents
4.5. Illumina Sequencing and Data Analysis
4.6. Quantitative Reverse-Transcription-PCR
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 4CL | 4-coumarate-CoA ligase |
| CHS | chalcone synthase |
| CHI | chalcone isomeras |
| F3H | flavanone 3-hydroxylase |
| F3′H | flavonoid 3-hydroxylase |
| F3′5′H | flavonoid 3′,5′-hydroxylase |
| DFR | dihydroflavonol 4-reductase |
| ANS | anthocyanidin synthase |
| OMT | O-methyltransferase |
| MBW | MYB-bHLH-WD40 |
| KEGG | kyoto encyclopedia of genes and genomes |
| MAPK | mitogen-activated protein kinase |
| SNARE | soluble N -ethylmaleimide-sensitive fusion protein attachment protein receptors |
| AOAC | association of official analytical chemists |
| COG | clusters of orthologous groups of proteins |
| GO | gene ontology |
References
- Asen, S.; Stewart, R.N.; Norris, K.H. Copigmentation of Anthocyanins in Plant-Tissues and Its Effect on Color. Phytochemistry 1972, 11, 1139–1144. [Google Scholar] [CrossRef]
- Florenciano, E.G.; Calderon, A.A.; Munoz, R.; Barcelo, A.R. Patterns of Anthocyanin Deposition in Vacuoles of Suspension Cultured Grapevine Cells. Phyton-Int. J. Exp. Bot 1992, 53, 47–50. [Google Scholar]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The Case for Anthocyanin Consumption to Promote Human Health: A Review. Compr. Rev. Food Sci. F. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Li, S.S.; Strid, A. Anthocyanin accumulation and changes in CHS and PR-5 gene expression in Arabidopsis thaliana after removal of the inflorescence stem (decapitation). Plant Physiol. Bioch. 2005, 43, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Honda, C.; Kotoda, N.; Wada, M.; Kondo, S.; Kobayashi, S.; Soejima, J.; Zhang, Z.L.; Tsuda, T.; Moriguchi, T. Anthocyanin biosynthetic genes are coordinately expressed during red coloration in apple skin. Plant Physiol. Bioch. 2002, 40, 955–962. [Google Scholar] [CrossRef]
- Sun, W.; Meng, X.Y.; Liang, L.J.; Li, Y.Q.; Zhou, T.T.; Cai, X.Q.; Wang, L.; Gao, X. Overexpression of a Freesia hybrida flavonoid 3-O-glycosyltransferase gene, Fh3GT1, enhances transcription of key anthocyanin genes and accumulation of anthocyanin and flavonol in transgenic petunia (Petunia hybrida). In Vitro Cell Dev.-Pl. 2017, 53, 478–488. [Google Scholar] [CrossRef]
- Zhu, Z.G.; Li, G.R.; Liu, L.; Zhang, Q.T.; Han, Z.; Chen, X.S.; Li, B. A R2R3-MYB Transcription Factor, VvMYBC2L2, Functions as a Transcriptional Repressor of Anthocyanin Biosynthesis in Grapevine (Vitis vinifera L.). Molecules 2019, 24, 92. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.D.; Meng, F.L.; Zheng, Y.N. Study of Anthocyanins in fruit of different Vaccinium genotypes. Acta Hortic. 2006, 715, 589–594. [Google Scholar] [CrossRef]
- Lloyd, A.; Brockman, A.; Aguirre, L.; Campbell, A.; Bean, A.; Cantero, A.; Gonzalez, A. Advances in the MYB-bHLH-WD Repeat (MBW) Pigment Regulatory Model: Addition of a WRKY Factor and Co-option of an Anthocyanin MYB for Betalain Regulation. Plant Cell Physiol. 2017, 58, 1431–1441. [Google Scholar] [CrossRef]
- Chiu, L.W.; Zhou, X.; Burke, S.; Wu, X.; Prior, R.L.; Li, L. The purple cauliflower arises from activation of a MYB transcription factor. Plant Physiol. 2010, 154, 1470–1480. [Google Scholar] [CrossRef]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.Y.; Ahn, Y.O.; Kim, S.H.; Kim, Y.H.; Lee, H.S.; Catanach, A.S.; Jacobs, J.M.; Conner, A.J.; Kwak, S.S. The sweet potato IbMYB1 gene as a potential visible marker for sweet potato intragenic vector system. Physiol. Plant 2010, 139, 229–240. [Google Scholar] [PubMed]
- Yamagishi, M.; Shimoyamada, Y.; Nakatsuka, T.; Masuda, K. Two R2R3-MYB genes, homologs of Petunia AN2, regulate anthocyanin biosyntheses in flower Tepals, tepal spots and leaves of asiatic hybrid lily. Plant Cell Physiol. 2010, 51, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Pattanaik, S.; Kong, Q.; Zaitlin, D.; Werkman, J.R.; Xie, C.H.; Patra, B.; Yuan, L. Isolation and functional characterization of a floral tissue-specific R2R3 MYB regulator from tobacco. Planta 2010, 231, 1061–1076. [Google Scholar] [CrossRef]
- Cho, J.S.; Nguyen, V.P.; Jeon, H.W.; Kim, M.H.; Eom, S.H.; Lim, Y.J.; Kim, W.C.; Park, E.J.; Choi, Y.I.; Ko, J.H. Overexpression of PtrMYB119, a R2R3-MYB transcription factor from Populus trichocarpa, promotes anthocyanin production in hybrid poplar. Tree Physiol. 2016, 36, 1162. [Google Scholar] [CrossRef]
- Li, Y.; Tu, K.; Zheng, S.; Wang, J.; Li, Y.; Hao, P.; Li, X. Association of feature gene expression with structural fingerprints of chemical compounds. J. Bioinform. Comput. Biol. 2011, 9, 503–519. [Google Scholar] [CrossRef]
- Fujii, K.; Murase, T.; Beppu, S.; Saida, K.; Takino, H.; Masaki, A.; Ijichi, K.; Kusafuka, K.; Iida, Y.; Onitsuka, T.; et al. MYB, MYBL1, MYBL2 and NFIB gene alterations and MYC overexpression in salivary gland adenoid cystic carcinoma. Histopathology 2017, 71, 823–834. [Google Scholar] [CrossRef]
- Janssen, B.J.; Lund, L.; Sinha, N. Overexpression of a homeobox gene, LeT6, reveals indeterminate features in the tomato compound leaf. Plant Physiol. 1998, 117, 771–786. [Google Scholar] [CrossRef]
- Wei, C.; Liang, G.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Jie, L. A Novel Integrated Method for Large-Scale Detection, Identification, and Quantification of Widely Targeted Metabolites: Application in the Study of Rice Metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [Green Version]
- Zhang, N.; Liu, B.; Ma, C.; Zhang, G.; Chang, J.; Si, H.; Wang, D. Transcriptome characterization and sequencing-based identification of drought-responsive genes in potato. Mol. Biol. Rep. 2014, 41, 505–517. [Google Scholar] [CrossRef]
- Dorn, K.M.; Fankhauser, J.D.; Wyse, D.L.; Marks, M.D. De novo assembly of the pennycress (Thlaspi arvense) transcriptome provides tools for the development of a winter cover crop and biodiesel feedstock. Plant J. 2013, 75, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; LaBonte, D.; Villordon, A.; Kfir, Y.; Solis, J.; Lapis, E.; Perlman, T.S.; Doron-Faigenboim, A.; Hetzroni, A.; Althan, L. Transcriptional profiling of sweetpotato (Ipomoea batatas) roots indicates down-regulation of lignin biosynthesis and up-regulation of starch biosynthesis at an early stage of storage root formation. BMC Genomics 2013, 14, 460. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Zhu, X.; Liu, Z.; Xi, X.; Li, G.; Cao, D.; Wei, L.; Li, J.; Liu, B. Functional MYB transcription factor encoding gene AN2 is associated with anthocyanin biosynthesis in Lycium ruthenicum Murray. BMC Plant Biol. 2019, 19, 169. [Google Scholar] [CrossRef] [PubMed]
- Christopher Dean, G.; Paula, C.; Virginia, W. A multidrug resistance-associated protein involved in anthocyanin transport in Zea mays. Plant Cell 2004, 16, 1812–1826. [Google Scholar]
- De Vetten, N.; Ter Horst, J.; Van Schaik, H.P.; De Boer, A.; Mol, J.; Koes, R. A cytochrome b5 is required for full activity of flavonoid 3′, 5′-hydroxylase, a cytochrome P450 involved in the formation of blue flower colors. Proc. Natl. Acad. Sci. USA 1999, 96, 778–783. [Google Scholar] [CrossRef]
- Vincenzo, D.A.; Riccardo, A.; Giorgia, B.; Immacolata, C.; Mar, C.M.; Ana Beatriz, C.S.; Pasquale, C.; Carlo, F.; Francesca, P.; Domenico, C. High AN1 variability and interaction with basic helix-loop-helix co-factors related to anthocyanin biosynthesis in potato leaves. Plant J. 2015, 80, 527–540. [Google Scholar]
- Blank, M.L.; Lee, T.; Fitzgerald, V.; Snyder, F. A specific acetylhydrolase for 1-alkyl-2-acetyl-sn-glycero-3-phosphocholine (a hypotensive and platelet-activating lipid). J. Biol. Chem. 1981, 256, 175–178. [Google Scholar]
- Li, X.; Hao, F.; Hu, X.; Wang, H.; Dai, B.; Wang, X.; Liang, H.; Cang, M.; Liu, D. Generation of Tbeta4 knock-in Cashmere goat using CRISPR/Cas9. Int. J. Biol. Sci. 2019, 15, 1743–1754. [Google Scholar] [CrossRef]
- Zhu, Q.; Sui, S.; Lei, X.; Yang, Z.; Lu, K.; Liu, G.; Liu, Y.G.; Li, M. Ectopic Expression of the Coleus R2R3 MYB-Type Proanthocyanidin Regulator Gene SsMYB3 Alters the Flower Color in Transgenic Tobacco. PLoS ONE 2015, 10, e0139392. [Google Scholar] [CrossRef]
- Zhu, H.-F.; Fitzsimmons, K.; Khandelwal, A.; Kranz, R.G. CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Mol. Plant 2009, 2, 790–802. [Google Scholar]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A Simple and General Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Cabau, C.; Escudie, F.; Djari, A.; Guiguen, Y.; Bobe, J.; Klopp, C. Compacting and correcting Trinity and Oases RNA—Seq de novo assemblies. PeerJ 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Iseli, C.; Jongeneel, C.V.; Bucher, P. ESTScan: A program for detecting, evaluating, and reconstructing potential coding regions in EST sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1999, 1999, 138–148. [Google Scholar]
- Romualdi, C.; Bortoluzzi, S.; D’Alessi, F.; Danieli, G.A. IDEG6: A web tool for detection of differentially expressed genes in multiple tag sampling experiments. Physiol. Genomics 2003, 12, 159–162. [Google Scholar] [CrossRef]
- Rivals, I.; Personnaz, L.; Taing, L.; Potier, M.-C. Enrichment or depletion of a GO category within a class of genes: Which test? Bioinformatics 2007, 23, 401–407. [Google Scholar] [CrossRef]
- Hurley, J.H.; Dasgupta, A.; Andrews, P.; Crowell, A.M.; Ringelberg, C.; Loros, J.J.; Dunlap, J.C. A Tool Set for the Genome-Wide Analysis of Neurospora crassa by RT-PCR. G3 (Bethesda) 2015, 5, 2043–2049. [Google Scholar] [CrossRef]
- An, S.L.; Mo, Y.X.; Ou, C.Q. [Probit analysis with SPSS 10.0 software]. Di Yi Jun Yi Da Xue Xue Bao 2002, 22, 1019–1021. [Google Scholar]
- An, S.; Zhang, Y.; Chen, Z. [Analysis of binary classification repeated measurement data with GEE and GLMMs using SPSS software]. Nan Fang Yi Ke Da Xue Xue Bao 2012, 32, 1777–1780. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Total Compounds | Differently Accumulated Compounds | Percent (%) | Up-Accumulation Compounds | Down-Accumulation Compounds |
|---|---|---|---|---|---|
| Anthocyanins | 9 | 7 | 77.78 | 6 | 1 |
| Polyphenol | 12 | 8 | 66.67 | 7 | 1 |
| Flavonol | 29 | 17 | 58.62 | 12 | 5 |
| Flavonoid | 18 | 10 | 55.56 | 8 | 2 |
| Flavone | 49 | 27 | 55.10 | 11 | 16 |
| Flavanone | 19 | 9 | 47.37 | 7 | 2 |
| Phenylpropanoids | 53 | 25 | 47.17 | 18 | 7 |
| Lipids | 68 | 31 | 45.59 | 10 | 21 |
| Vitamins and derivatives | 15 | 6 | 40.00 | 3 | 3 |
| Terpene | 17 | 6 | 35.29 | 2 | 4 |
| Organic acids and derivatives | 97 | 34 | 35.05 | 17 | 17 |
| Phenolamides | 16 | 5 | 31.25 | 1 | 4 |
| Others | 23 | 5 | 21.74 | 1 | 4 |
| Amino acid and derivatives | 82 | 16 | 19.51 | 0 | 16 |
| Nucleotide and derivates | 47 | 8 | 17.02 | 4 | 4 |
| Isoflavone | 6 | 1 | 16.67 | 0 | 1 |
| Carbohydrates | 19 | 3 | 15.79 | 1 | 2 |
| Alkaloids | 33 | 5 | 15.15 | 1 | 4 |
| Indole derivatives | 7 | 1 | 14.29 | 0 | 1 |
| Alcohols | 13 | 0 | 0.00 | 0 | 0 |
| Quinones | 2 | 0 | 0.00 | 0 | 0 |
| Sterides | 5 | 0 | 0.00 | 0 | 0 |
| Total | 639 | 224 | 35.05 | 109 | 115 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zong, Y.; Li, S.; Xi, X.; Cao, D.; Wang, Z.; Wang, R.; Liu, B. Comprehensive Influences of Overexpression of a MYB Transcriptor Regulating Anthocyanin Biosynthesis on Transcriptome and Metabolome of Tobacco Leaves. Int. J. Mol. Sci. 2019, 20, 5123. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205123
Zong Y, Li S, Xi X, Cao D, Wang Z, Wang R, Liu B. Comprehensive Influences of Overexpression of a MYB Transcriptor Regulating Anthocyanin Biosynthesis on Transcriptome and Metabolome of Tobacco Leaves. International Journal of Molecular Sciences. 2019; 20(20):5123. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205123
Chicago/Turabian StyleZong, Yuan, Shiming Li, Xinyuan Xi, Dong Cao, Zhong Wang, Ran Wang, and Baolong Liu. 2019. "Comprehensive Influences of Overexpression of a MYB Transcriptor Regulating Anthocyanin Biosynthesis on Transcriptome and Metabolome of Tobacco Leaves" International Journal of Molecular Sciences 20, no. 20: 5123. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205123