phyB and HY5 are Involved in the Blue Light-Mediated Alleviation of Dormancy of Arabidopsis Seeds Possibly via the Modulation of Expression of Genes Related to Light, GA, and ABA


Abstract
:1. Introduction
2. Results
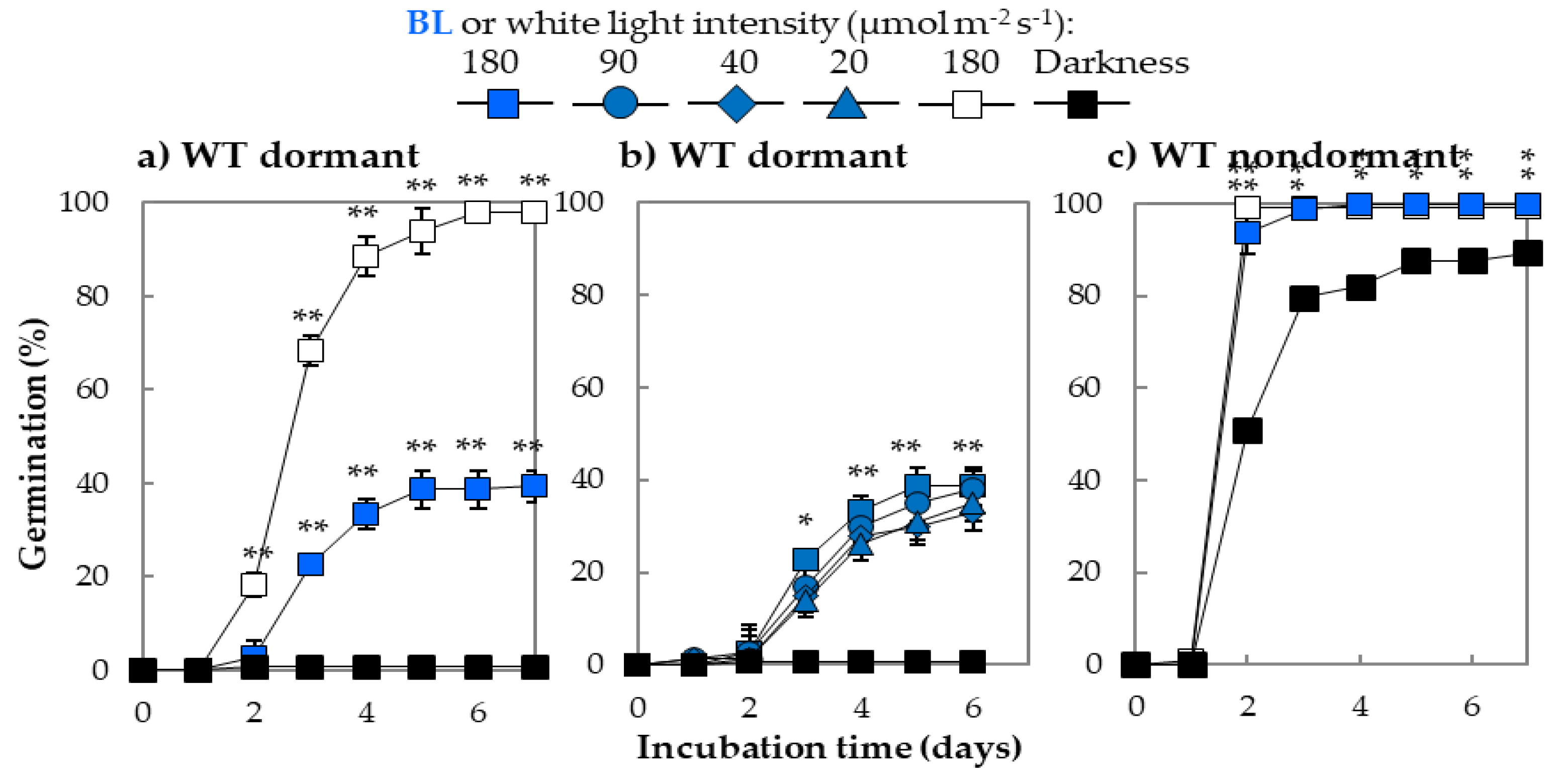
2.1. The Effect of Blue Light on Germination of Arabidopsis Seeds Characterized by Different Depths of Dormancy
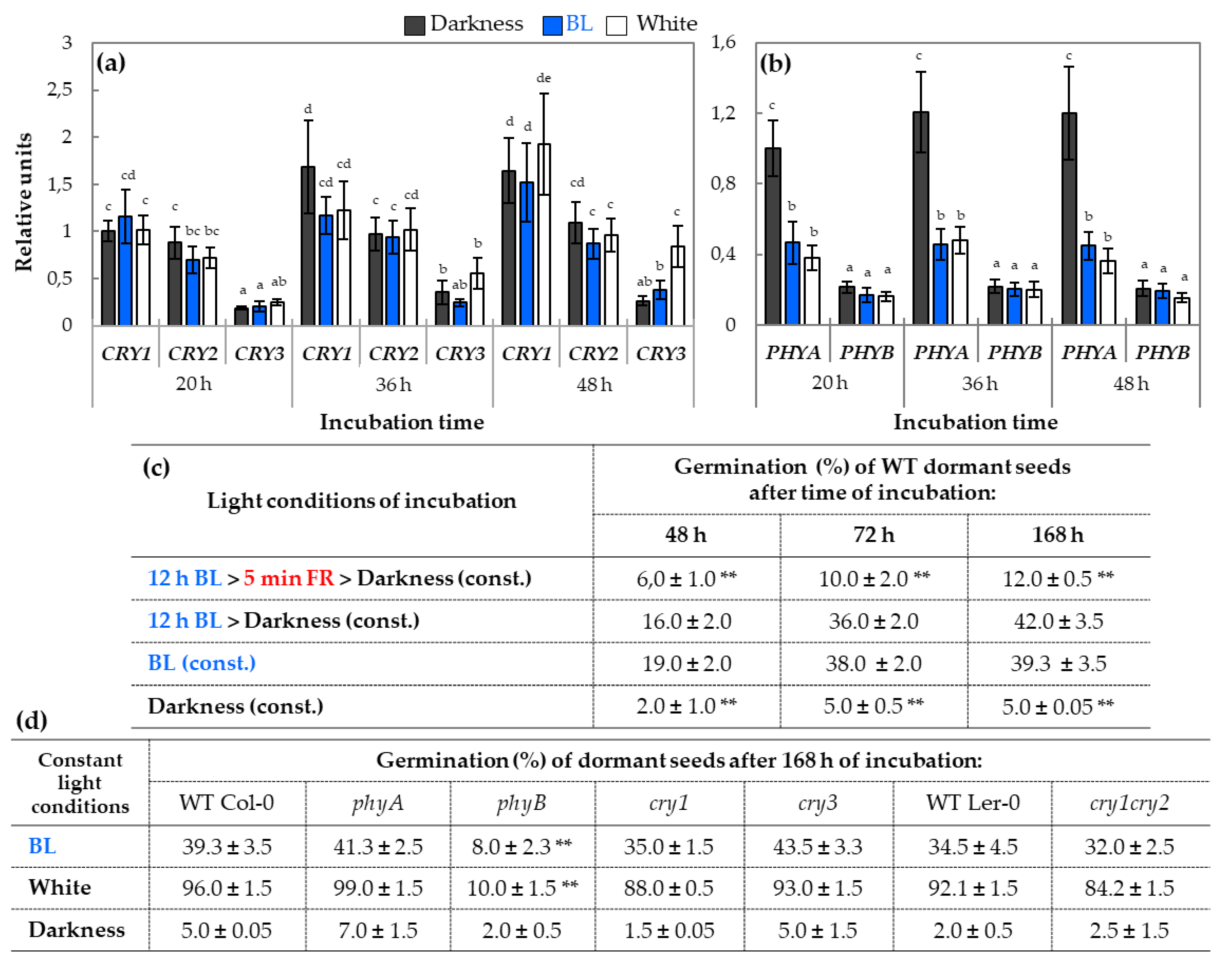
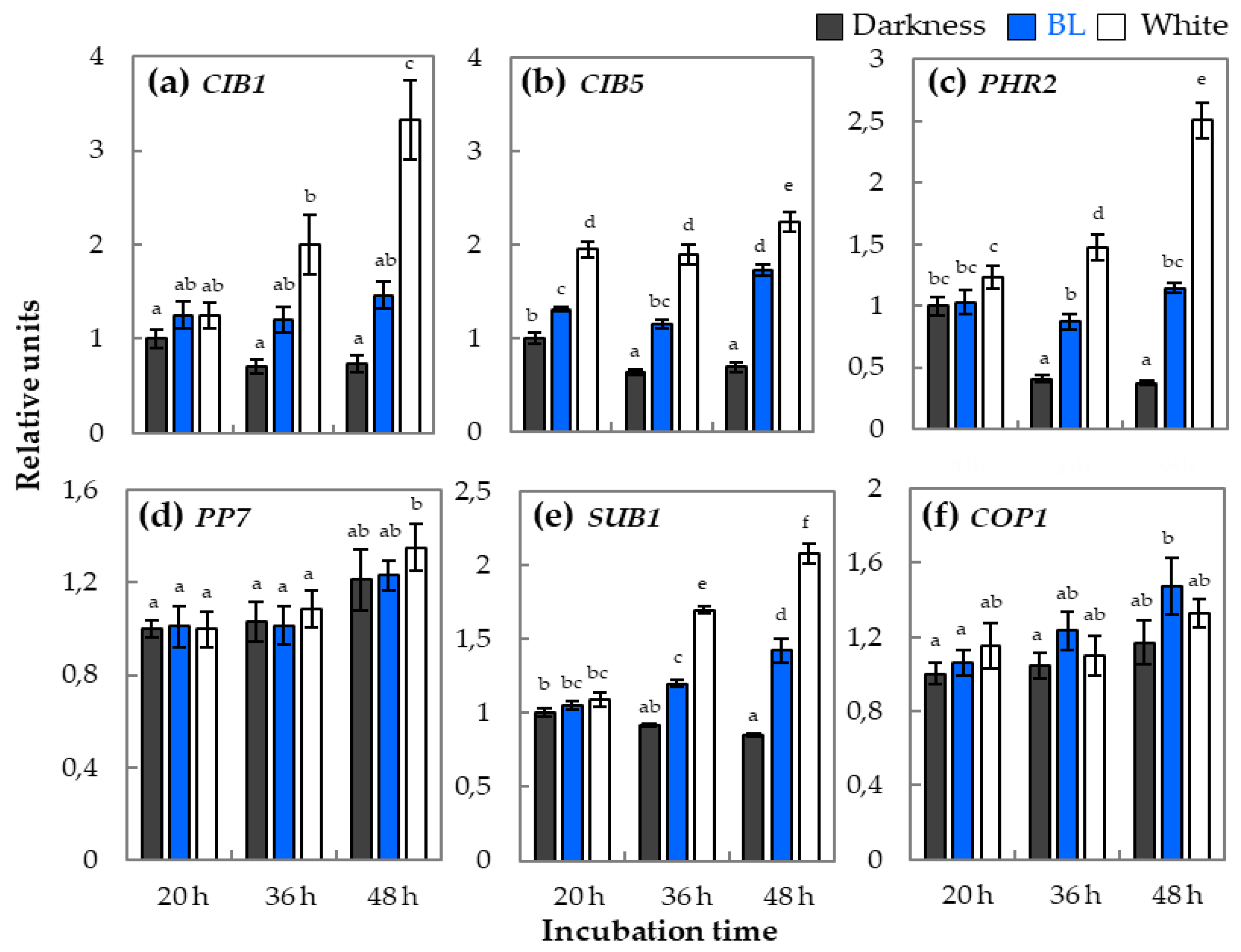
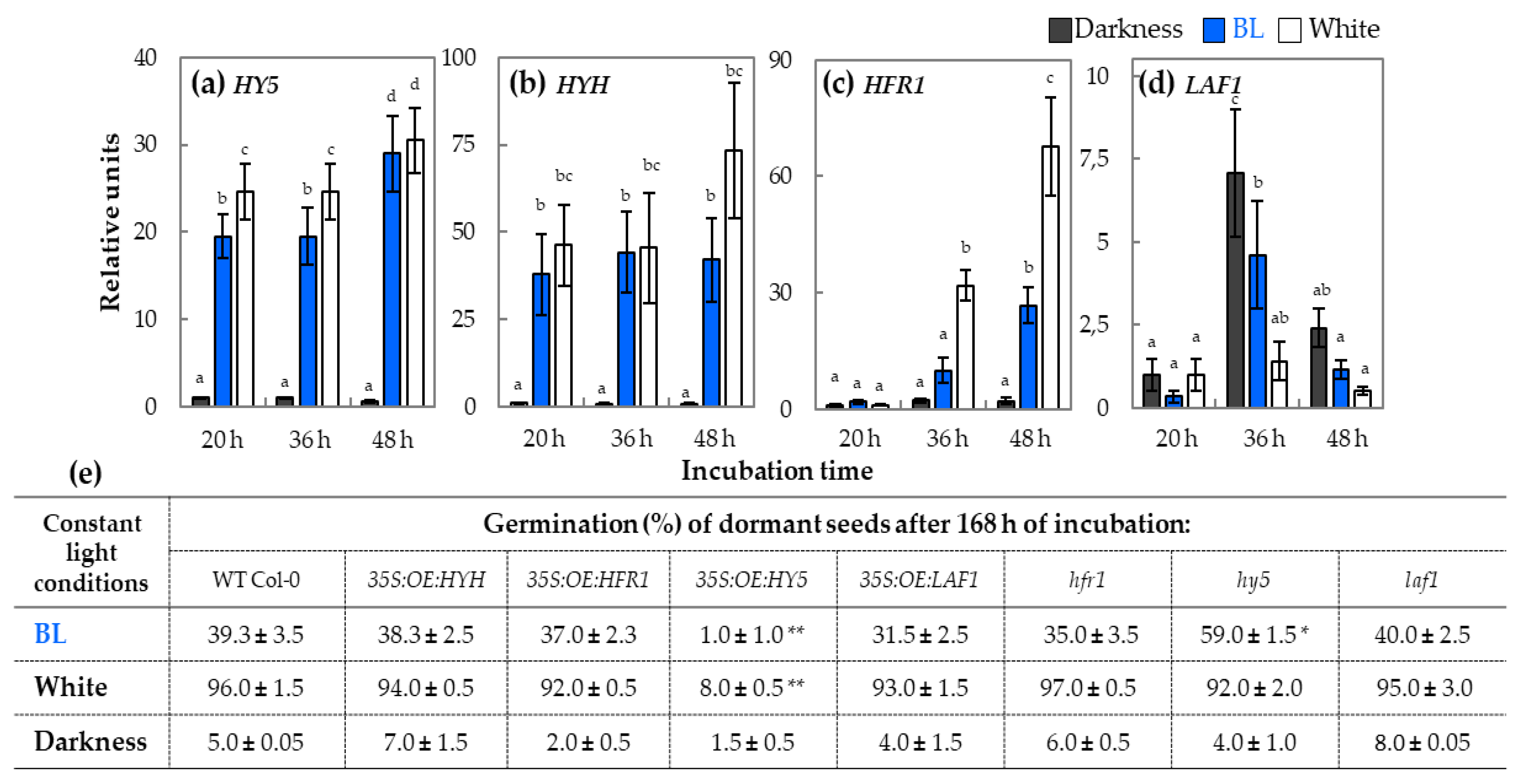
2.2. qRT-PCR Analysis of Relative Expression of Genes Involved in Light Perception and Signaling During Germination
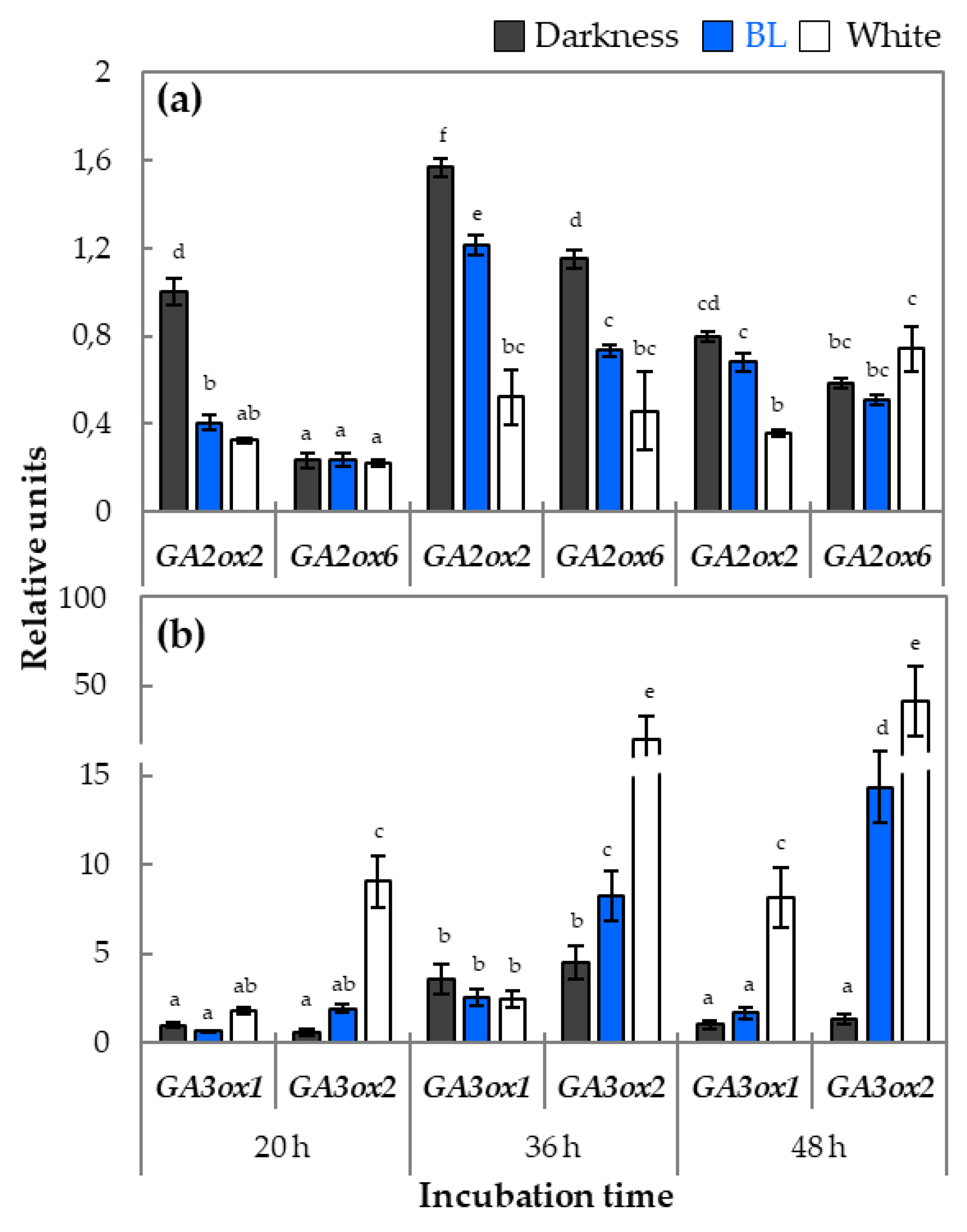
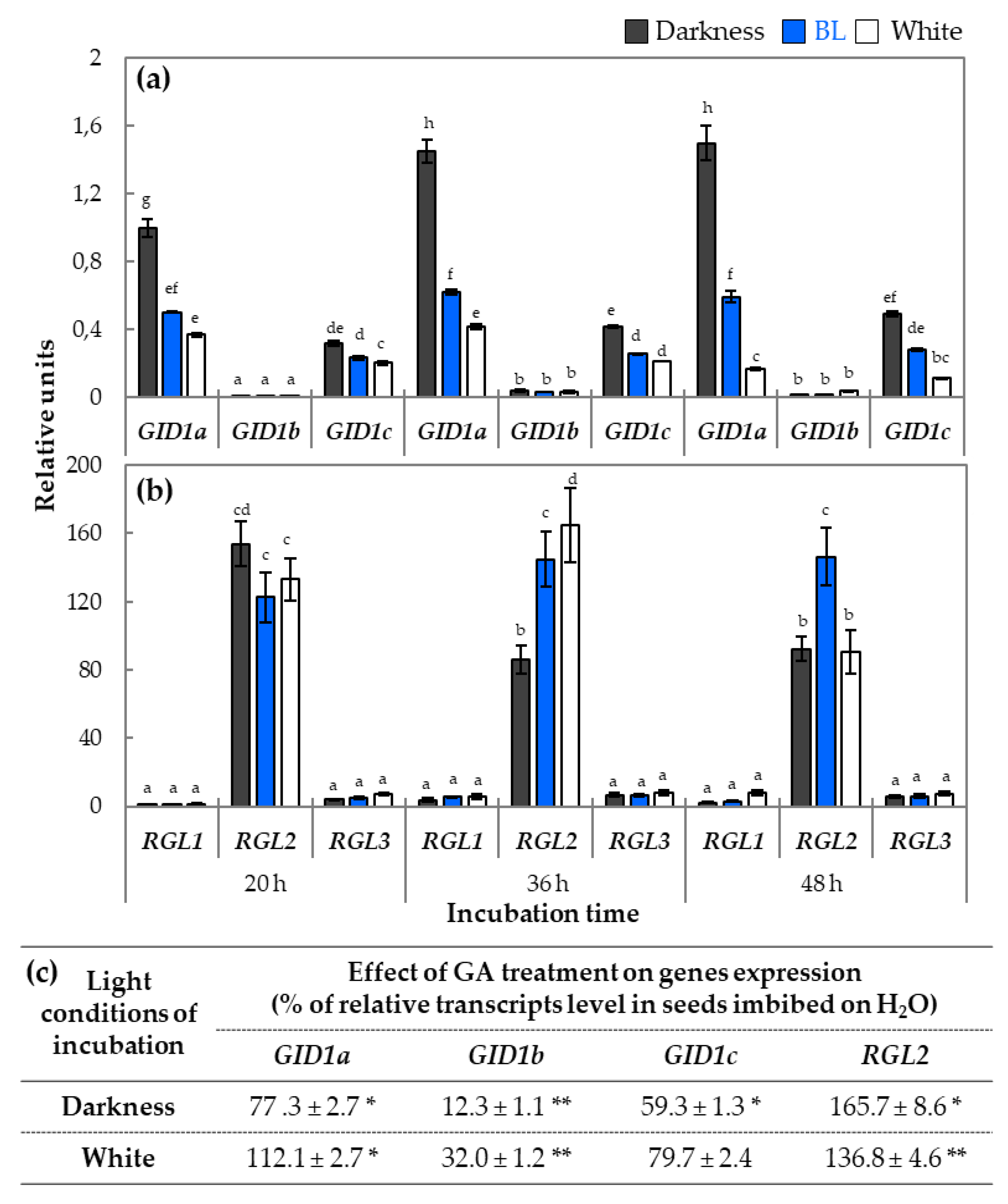
2.3. The Analysis of Expression Profiles of Genes Involved in GA Metabolism and Signaling in Light-Dependent Germination of Dormant Seeds
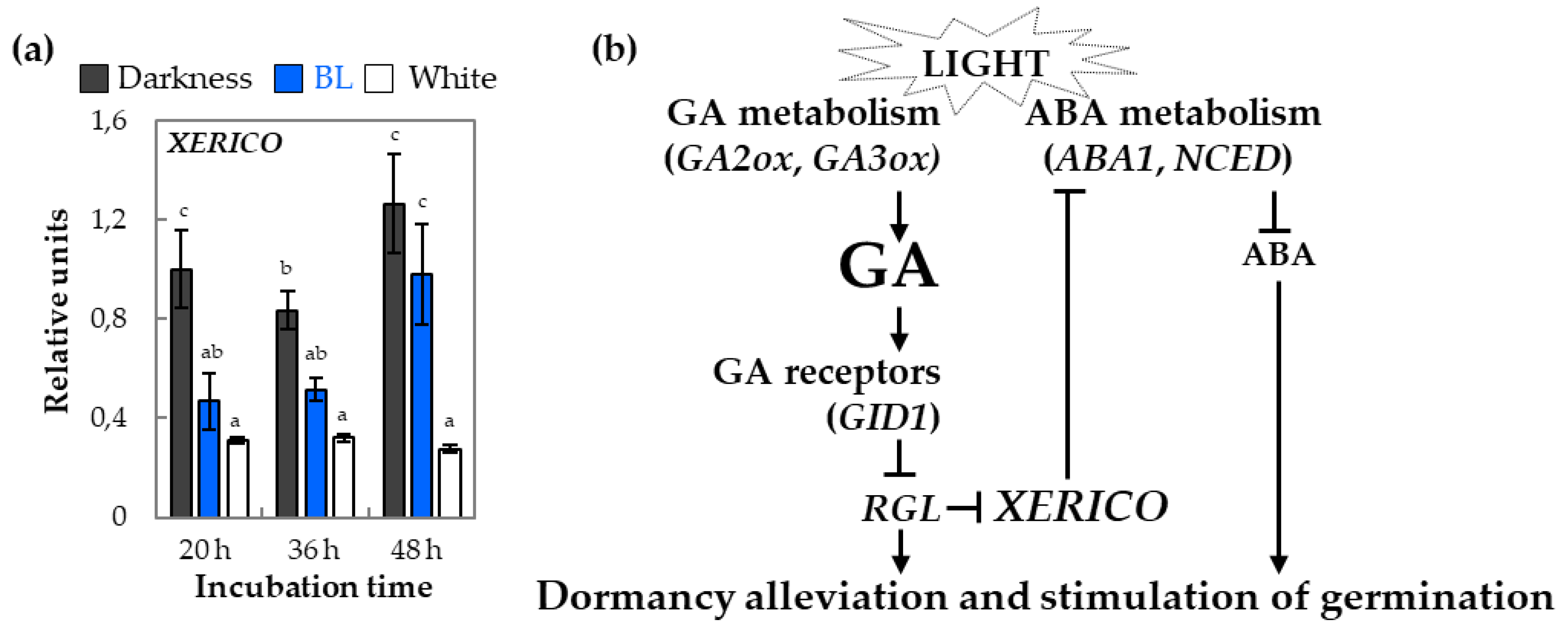
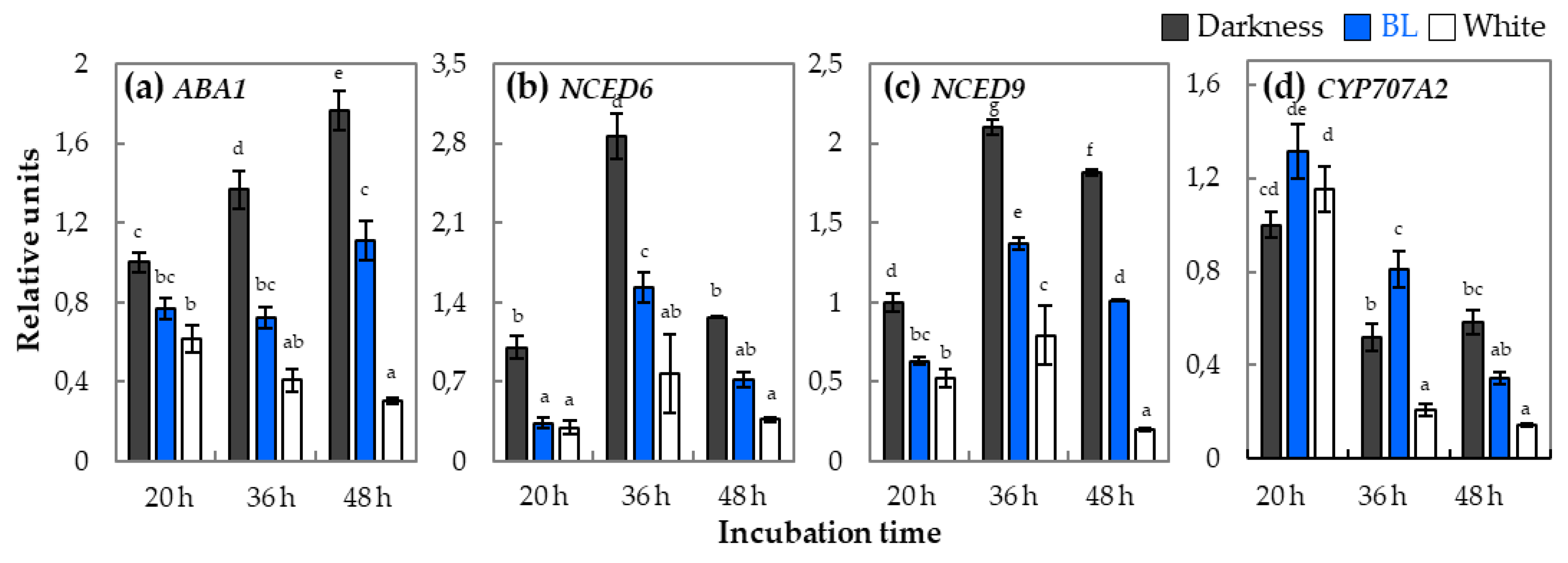
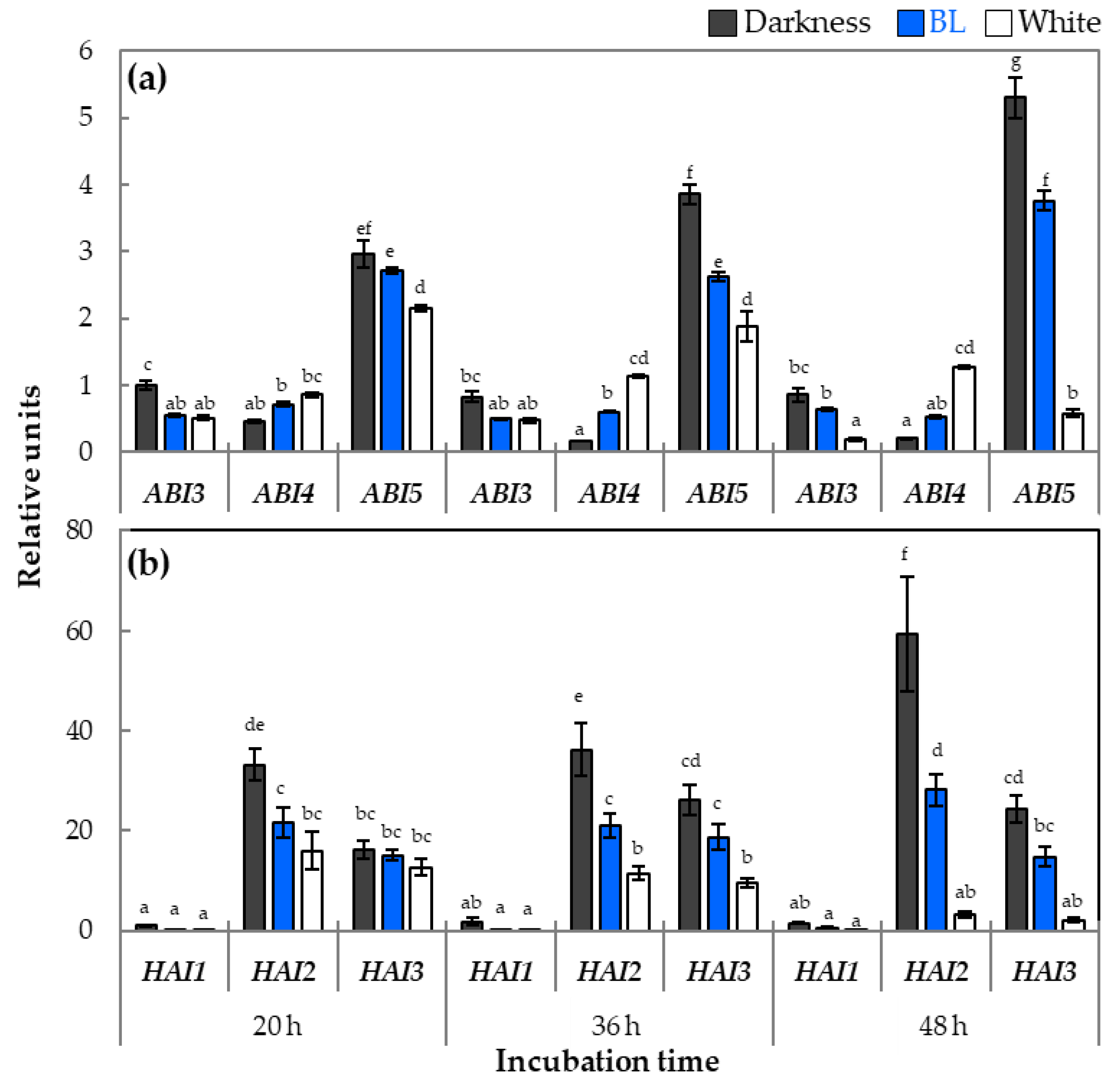
2.4. The Expression Patterns of Genes Involved in ABA Metabolism and Signaling in Light-Dependent Germination of Dormant Seeds
3. Discussion
3.1. Blue Light Alleviates Dormancy and Stimulates Germination of Arabidopsis Seeds
3.2. The phy Perceive Blue Light in Germinating Dormant Arabidopsis Seeds
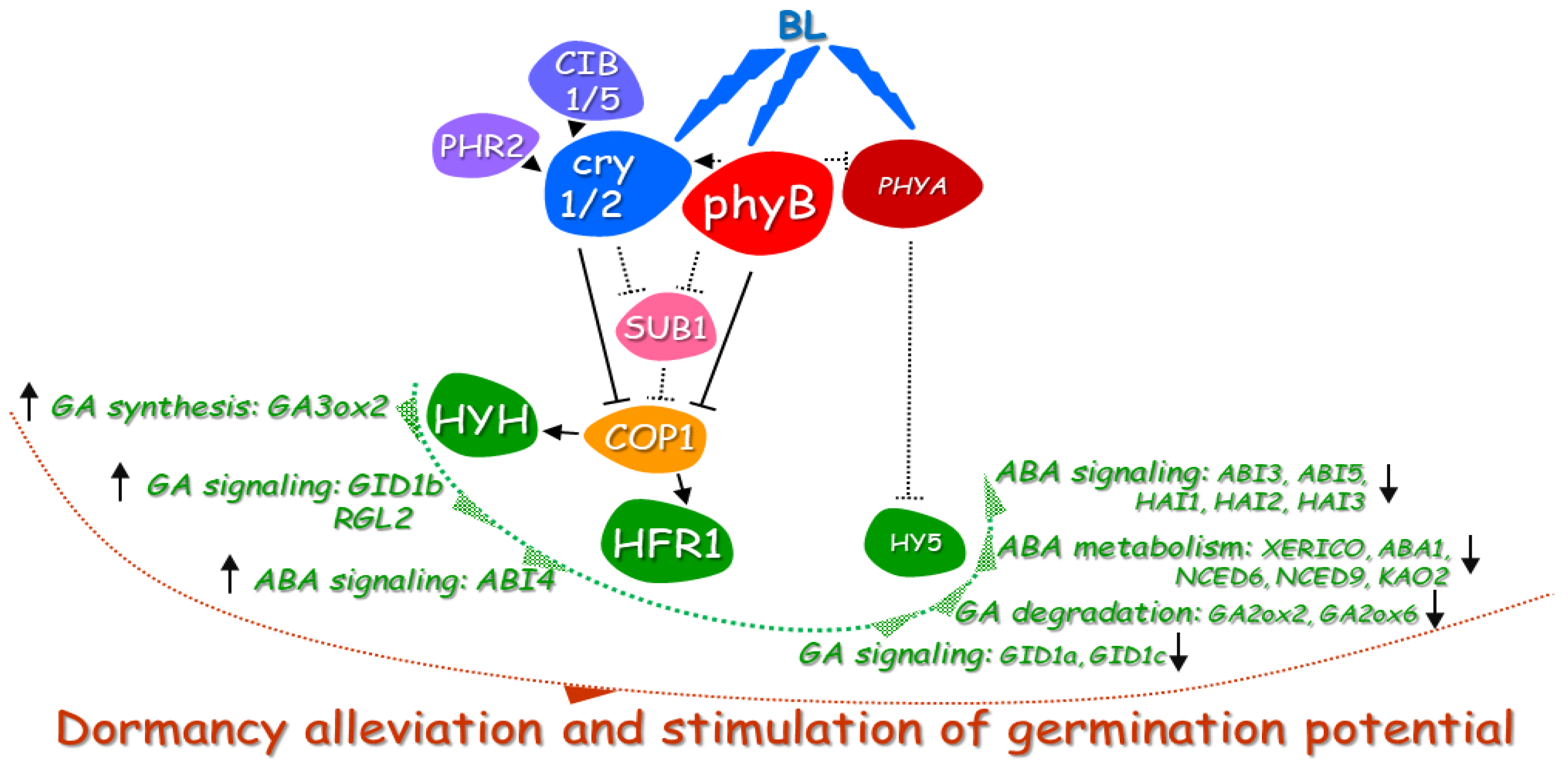
3.3. The phy–cry-Related Mechanism of Blue Light Action in Dormancy Alleviation and Germination of Arabidopsis Seeds Requires Interaction with Various Regulatory Proteins
3.4. HY5 Transcription Factor is the Most Probably Involved in Modulation of Blue Light-Mediated Dormancy Alleviation of Arabidopsis Seeds
3.5. Effect of Blue Light on GA and ABA Metabolism in Germinating Dormant Seeds of Arabidopsis
3.6. Effect of Blue Light on GA and ABA Signaling in Germinating Dormant Seeds of Arabidopsis
4. Materials and Methods
4.1. Plant Material and Germination Assays
4.2. Construct Preparation, Plant Transformation, and Phenotypic Analysis of Transgenic Plants
4.3. RNA Extraction and cDNA Synthesis
4.4. Quantitative Real Time–PCR Analysis
4.5. Statystical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABA | Abscisic Acid |
| ABA1 | ARABIDOPSIS THALIANA ABA DEFICIENT 1 |
| ABI3, 4 and 5 | ABA INSENSITIVE3, 4 and 5 |
| ACT7 | ACTIN 7 |
| APC2 | ANAPHASE PROMOTING COMPLEX 2 |
| BL | Blue Light |
| CIB1 and 5 | CRYPTOCHROME INTERACTING BASIC HELIX-LOOP-HELIX 1 and 5 |
| CRY1, 2 and 3 | CRYPTOCHROME 1, 2, and 3 |
| COP1 | CONSTITUTIVE PHOTOMORPHOGENIC 1 |
| CUL4 | CULLIN4 |
| CYP707A2 | CYTOCHROME P450, FAMILY 707, SUBFAMILY A, POLYPEPTIDE 2 |
| DELLA | DELLA factors encoded by RGL genes |
| FKF1 | FLAVIN-BINDING, KELCH REPEAT, F-BOX 1 |
| FR | Far Red Light |
| FT | FLOWERING LOCUS |
| GA | Gibberellins |
| GA3ox1 and 2 | GA 3–OXIDASE 1 and 2 |
| GA2ox2 and 6 | GA 2–OXIDASE 2 and 6 |
| GID1a, 1b and 1c | GA INSENSITIVE DWARF 1a, 1b, and 1c |
| HAI1, 2 and 3 | HIGHLY ABA–INDUCED PP2C 1, 2, and 3 |
| HBT | HOBBIT |
| HFR1 | LONG HYPOCOTYL IN FAR-RED 1 |
| HY5 | ELONGATED HYPOCOTYL 5 |
| HYH | HY5–HOMOLOG |
| KAO2 | ENT–KAURENOIC ACID OXYDASE 2 |
| LAF1 | LONG AFTER FAR-RED LIGHT 1 |
| LFR | Low Fluence Response |
| LKP2 | LOV KELCH PROTEIN 2 |
| LLP BL | LOV/LOV domain-containing protein |
| LOV | Light, Oxygen, or Voltage photosensory domain |
| NCED6 and 9 | NINE–CIS EPOXYCAROTENOID DIOXYGENASE 6 and 9 |
| ND | Nondormant |
| PAC | Paclobutrazol |
| phot1 and 2 | phototropin 1 and 2 |
| PHR2 | PHOTOLYASE/BLUE LIGHT RECEPTOR 2 |
| PHYA and B | PHYTOCHROMEA and B |
| PIF | PHYTOCHROME INTERACTING FACTOR |
| PIL5 | PIF3-LIKE 5 |
| PP7 | Serine/Threonine Phosphatase 7 |
| PP2C | PROTEIN PHOSPHATASE TYPE 2C |
| PYR/PYL/RCAR | PYRABACTIN RESISTANCE/PYR1-LIKE/REGULATORY COMPONENT OF ABA RECEPTOR |
| R | Red Light |
| RGL1, 2 and 3 | REPRESSOR OF GA-LIKE 1, 2, and 3 |
| SnRK2 | SNF1-related protein kinases 2 |
| SPA | SUPRESS PHYTOCHROME A-105 |
| SUB1 | CALCIUM ION-BINDING PROTEIN SUB1 |
| VLFR | Very Low Fluence Response |
| WT | Wild Type |
| XERICO | gene encoding RING/U-box superfamily protein XERICO |
| ZTL | ZEITLUPE |
References
- Song, J.; Liu, Q.; Hu, B.; Wu, W. Photoreceptor PhyB Involved in Arabidopsis Temperature Perception and Heat–Tolerance Formation. Int. J. Mol. Sci. 2017, 18, 1194. [Google Scholar] [CrossRef] [PubMed]
- Mérai, Z.; Graeber, K.; Wilhelmsson, P.; Ullrich, K.K.; Arshad, W.; Grosche, C.; Tarkowská, D.; Turečková, V.; Strnad, M.; Rensing, S.A.; et al. Aethionema arabicum: A novel model plant to study the light control of seed germination. J. Exp. Bot. 2019, 70, 3313–3328. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Nambara, E.; Choi, G.; Yamaguchi, S. Interaction of light and hormone signals in germinating seeds. Plant Mol. Biol. 2009, 69, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Footitt, S.; Tang, A.; Finch-Savage, W.E. Predicted global warming scenarios impact on the mother plant to alter seed dormancy and germination behaviour in Arabidopsis. Plant Cell Environ. 2018, 41, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Leymarie, J.; Vitkauskaité, G.; Hoang, H.H.; Gendreau, E.; Chazoule, V.; Meimoun, P.; Corbineau, F.; El-Maarouf-Bouteau, H.; Bailly, C. Role of Reactive Oxygen Species in the Regulation of Arabidopsis Seed Dormancy. Plant Cell Physiol. 2012, 53, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; Karpiński, S. Phytohormones Signaling Pathways and ROS Involvement in Seed Germination. Front. Plant Sci. 2016, 7, 864. [Google Scholar] [CrossRef]
- Shinomura, T.; Nagatani, A.; Chory, J.; Furuya, M. The Induction of Seed Germination in Arabidopsis thaliana Is Regulated Principally by Phytochrome B and Secondarily by Phytochrome A. Plant Physiol. 1994, 104, 363–371. [Google Scholar] [CrossRef]
- Oracz, K.; Stawska, M. Cellular Recycling of Proteins in Seed Dormancy Alleviation and Germination. Plant Sci. 2016, 1128. [Google Scholar] [CrossRef]
- Oracz, K.; Voegele, A.; Tarkowská, D.; Jacquemoud, D.; Turecková, V.; Urbanová, T.; Strnad, M.; Sliwinska, E.; Leubner–Metzger, G. Myrigalone A inhibits Lepidium sativum seed germination by interference with gibberellin metabolism and apoplastic superoxide production required for embryo extension growth and endosperm rupture. Plant Cell Physiol. 2012, 53, 81–95. [Google Scholar] [CrossRef]
- Chen, M.; Chory, J.; Fankhauser, C. Light Signal Transduction in Higher Plants. Annu. Rev. Genet. 2004, 38, 87–117. [Google Scholar] [CrossRef]
- Ito, S.; Song, Y.H.; Imaizumi, T. LOV Domain-Containing F-Box Proteins: Light—Dependent Protein Degradation Modules in Arabidopsis. Mol. Plant 2012, 5, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Zoltowski, B.D.; Imaizumi, T. Structure and function of the ZTL/FKF1/LKP2 group proteins in Arabidopsis. Enzymes 2014, 35, 213–239. [Google Scholar] [PubMed]
- Klar, T.; Pokorny, R.; Moldt, J.; Batschauer, A.; Essen, L.-O. Cryptochrome 3 from Arabidopsis thaliana: Structural and Functional Analysis of its Complex with a Folate Light Antenna. J. Mol. Biol. 2007, 366, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The Cryptochrome Blue Light Receptors. Arab. Book Am. Soc. Plant Biol. 2010, 8. [Google Scholar] [CrossRef]
- Stawska, M.; Oracz, K. Sieć powiązań szlaków fitochromowych, kryptochromowych oraz indukowanych przez regulatory wzrostu i rozwoju w biologii nasion. Post. Biol. Kom. 2015, 42, 687–706. [Google Scholar]
- Podolec, R.; Ulm, R. Photoreceptor–mediated regulation of the COP1/SPA E3 ubiquitin ligase. Curr. Opin. Plant Biol. 2018, 45, 18–25. [Google Scholar] [CrossRef]
- Shinomura, T.; Nagatani, A.; Hanzawa, H.; Kubota, M.; Watanabe, M.; Furuya, M. Action spectra for phytochrome A– and B–specific photoinduction of seed germination in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1996, 93, 8129–8133. [Google Scholar] [CrossRef]
- Saijo, Y.; Zhu, D.; Li, J.; Rubio, V.; Zhou, Z.; Shen, Y.; Hoecker, U.; Wang, H.; Deng, X.W. Arabidopsis COP1/SPA1 complex and phytochrome A signaling intermediates associate with distinct phosphorylated forms of phytochrome A in balancing signal propagation and attenuation. Mol. Cell 2008, 31, 607–613. [Google Scholar] [CrossRef]
- Seo, M.; Hanada, A.; Kuwahara, A.; Endo, A.; Okamoto, M.; Yamauchi, Y.; North, H.; Marion-Poll, A.; Sun, T.; Koshiba, T.; et al. Regulation of hormone metabolism in Arabidopsis seeds: Phytochrome regulation of abscisic acid metabolism and abscisic acid regulation of gibberellin metabolism. Plant J. 2006, 48, 354–366. [Google Scholar] [CrossRef]
- Ibarra, S.E.; Auge, G.; Sánchez, R.A.; Botto, J.F. Transcriptional programs related to phytochrome A function in Arabidopsis seed germination. Mol. Plant 2013, 6, 1261–1273. [Google Scholar] [CrossRef]
- Griffiths, J.; Murase, K.; Rieu, I.; Zentella, R.; Zhang, Z.-L.; Powers, S.J.; Gong, F.; Phillips, A.L.; Hedden, P.; Sun, T.; et al. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell 2006, 18, 3399–3414. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. GA Signaling: Direct Targets of DELLA Proteins. Plant Cell 2007, 19, 2970. [Google Scholar] [CrossRef]
- Tischer, S.V.; Wunschel, C.; Papacek, M.; Kleigrewe, K.; Hofmann, T.; Christmann, A.; Grill, E. Combinatorial interaction network of abscisic acid receptors and coreceptors from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, 10280–10285. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.-L.; He, S.-B.; Zhang, Y.-C.; Zhu, D.-M.; Zhang, J.-Y.; Jia, K.-P.; Sun, S.-X.; Li, L.; Yang, H.-Q. Blue-light—Dependent interaction of cryptochrome 1 with SPA1 defines a dynamic signaling mechanism. Genes Dev. 2011, 25, 1023–1028. [Google Scholar] [CrossRef]
- Liu, B.; Zuo, Z.; Liu, H.; Liu, X.; Lin, C. Arabidopsis cryptochrome 1 interacts with SPA1 to suppress COP1 activity in response to blue light. Genes Dev. 2011, 25, 1029–1034. [Google Scholar] [CrossRef]
- Wang, Q.; Zuo, Z.; Wang, X.; Liu, Q.; Gu, L.; Oka, Y.; Lin, C. Beyond the photocycle—How cryptochromes regulate photoresponses in plants? Curr. Opin. Plant Biol. 2018, 45, 120–126. [Google Scholar] [CrossRef]
- Liu, H.; Yu, X.; Li, K.; Klejnot, J.; Yang, H.; Lisiero, D.; Lin, C. Photoexcited CRY2 Interacts with CIB1 to Regulate Transcription and Floral Initiation in Arabidopsis. Science 2008, 322, 1535–1539. [Google Scholar] [CrossRef]
- Barrero, J.M.; Downie, A.B.; Xu, Q.; Gubler, F. A Role for Barley CRYPTOCHROME1 in Light Regulation of Grain Dormancy and Germination. Plant Cell 2014, 26, 1094–1104. [Google Scholar] [CrossRef]
- Gubler, F.; Hughes, T.; Waterhouse, P.; Jacobsen, J. Regulation of Dormancy in Barley by Blue Light and After-Ripening: Effects on Abscisic Acid and Gibberellin Metabolism. Plant Physiol. 2008, 147, 886–896. [Google Scholar] [CrossRef]
- Strasser, B.; Sánchez-Lamas, M.; Yanovsky, M.J.; Casal, J.J.; Cerdán, P.D. Arabidopsis thaliana life without phytochromes. Proc. Natl. Acad. Sci. USA 2010, 107, 4776–4781. [Google Scholar] [CrossRef]
- Hofmann, N. Cryptochromes and Seed Dormancy: The Molecular Mechanism of Blue Light Inhibition of Grain Germination. Plant Cell 2014, 26, 846. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.V.; Barrero, J.M.; Hughes, T.; Julkowska, M.; Taylor, J.M.; Xu, Q.; Gubler, F. Roles for blue light, jasmonate and nitric oxide in the regulation of dormancy and germination in wheat grain Triticum aestivum L. Planta 2013, 238, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Casal, J.J.; Candia, A.N.; Sellaro, R. Light perception and signalling by phytochrome A. J. Exp. Bot. 2014, 65, 2835–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neff, M.M.; Fankhauser, C.; Chory, J. Light: An indicator of time and place. Genes Dev. 2000, 14, 257–271. [Google Scholar] [PubMed]
- Pokorny, R.; Klar, T.; Hennecke, U.; Carell, T.; Batschauer, A.; Essen, L.-O. Recognition and repair of UV lesions in loop structures of duplex DNA by DASH–type cryptochrome. Proc. Natl. Acad. Sci. USA 2008, 105, 21023–21027. [Google Scholar] [CrossRef] [Green Version]
- Kleine, T.; Lockhart, P.; Batschauer, A. An Arabidopsis protein closely related to Synechocystis cryptochrome is targeted to organelles. Plant J. Cell Mol. Biol. 2003, 35, 93–103. [Google Scholar] [CrossRef]
- Clack, T.; Mathews, S.; Sharrock, R.A. The phytochrome apoprotein family in Arabidopsis is encoded by five genes: The sequences and expression of PHYD and PHYE. Plant Mol. Biol. 1994, 25, 413–427. [Google Scholar] [CrossRef]
- Sharrock, R.A.; Quail, P.H. Novel phytochrome sequences in Arabidopsis thaliana: Structure, evolution, and differential expression of a plant regulatory photoreceptor family. Genes Dev. 1989, 3, 1745–1757. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Yamaguchi, S.; Kamiya, Y.; Bae, G.; Chung, W.I.; Choi, G. Light activates the degradation of PIL5 protein to promote seed germination through gibberellin in Arabidopsis. Plant J. 2006, 47, 124–139. [Google Scholar] [CrossRef]
- Guo, H.; Mockler, T.; Duong, H.; Lin, C. SUB1, an Arabidopsis Ca2+—Binding Protein Involved in Cryptochrome and Phytochrome Coaction. Science 2001, 291, 487–490. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Jarillo, J.A.; Cashmore, A.R. PHR2: A novel Arabidopsis gene related to the blue-light. Plant Physiol. 1998, 117, 718–719. [Google Scholar]
- Andreeva, A.V.; Kutuzov, M.A. Nuclear localization of the plant protein Ser/Thr phosphatase PP7. Mol. Cell Biol. Res. Commun. MCBRC 2001, 4, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Higa, L.A.; Wu, M.; Ye, T.; Kobayashi, R.; Sun, H.; Zhang, H. CUL4–DDB1 ubiquitin ligase interacts with multiple WD40–repeat proteins and regulates histone methylation. Nat. Cell Biol. 2006, 8, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Arias, E.E.; Chen, J.; Harper, J.W.; Walter, J.C. A Family of Diverse Cul4–Ddb1–Interacting Proteins Includes Cdt2, which Is Required for S Phase Destruction of the Replication Factor Cdt1. Mol. Cell 2006, 23, 709–721. [Google Scholar] [CrossRef]
- Deng, X.-W.; Matsui, M.; Wei, N.; Wagner, D.; Chu, A.M.; Feldmann, K.A.; Quail, P.H. COP1, an Arabidopsis regulatory gene, encodes a protein with both a zinc-binding motif and a Gβ homologous domain. Cell 1992, 71, 791–801. [Google Scholar] [CrossRef]
- Wang, H.; Ma, L.-G.; Li, J.-M.; Zhao, H.-Y.; Deng, X.W. Direct Interaction of Arabidopsis Cryptochromes with COP1 in Light Control Development. Science 2001, 294, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, B.; Zhao, C.; Pepper, M.; Lin, C. The action mechanisms of plant cryptochromes. Trends Plant Sci. 2011, 16, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Yi, C.; Deng, X.W. COP1—From plant photomorphogenesis to mammalian tumorigenesis. Trends Cell Biol. 2005, 15, 618–625. [Google Scholar] [CrossRef]
- Lefebvre, V.; North, H.; Frey, A.; Sotta, B.; Seo, M.; Okamoto, M.; Nambara, E.; Marion-Poll, A. Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy. Plant J. 2006, 45, 309–319. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Ogawa, M.; Kuwahara, A.; Hanada, A.; Kamiya, Y.; Yamaguchi, S. Activation of Gibberellin Biosynthesis and Response Pathways by Low Temperature during Imbibition of Arabidopsis thaliana Seeds. Plant Cell 2004, 16, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Ariizumi, T.; Lawrence, P.K.; Steber, C.M. The Role of Two F-Box Proteins, SLEEPY1 and SNEEZY, in Arabidopsis Gibberellin Signaling. Plant Physiol. 2011, 155, 765–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murase, K.; Hirano, Y.; Sun, T.; Hakoshima, T. Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 2008, 456, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Voegele, A.; Graeber, K.; Oracz, K.; Tarkowská, D.; Jacquemoud, D.; Turečková, V.; Urbanová, T.; Strnad, M.; Leubner-Metzger, G. Embryo growth, testa permeability, and endosperm weakening are major targets for the environmentally regulated inhibition of Lepidium sativum seed germination by myrigalone A. J. Exp. Bot. 2012, 63, 5337–5350. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, J.; Neff, M.M.; Hong, S.-W.; Zhang, H.; Deng, X.-W.; Xiong, L. Integration of light and abscisic acid signaling during seed germination and early seedling development. Proc. Natl. Acad. Sci. USA 2008, 105, 4495–4500. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Park, J.; Lee, N.; Jeong, J.; Toh, S.; Watanabe, A.; Kim, J.; Kang, H.; Kim, D.H.; Kawakami, N.; et al. ABA–INSENSITIVE3, ABA–INSENSITIVE5, and DELLAs Interact to Activate the Expression of SOMNUS and Other High-Temperature-Inducible Genes in Imbibed Seeds in Arabidopsis. Plant Cell 2013, 25, 4863–4878. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Ji, Q.; Huang, Y.; Jiang, Z.; Bao, M.; Wang, H.; Lin, R. FAR–RED ELONGATED HYPOCOTYL3 and FAR–RED IMPAIRED RESPONSE1 Transcription Factors Integrate Light and Abscisic Acid Signaling in Arabidopsis. Plant Physiol. 2013, 163, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Zhang, H.; Wang, S.; Chen, M.; Wu, Y.; Tang, S.; Liu, C.; Feng, Y.; Cao, X.; Xie, Q. ABI4 Regulates Primary Seed Dormancy by Regulating the Biogenesis of Abscisic Acid and Gibberellins in Arabidopsis. PLoS Genet. 2013, 9, e1003577. [Google Scholar] [CrossRef] [Green Version]
- Kurup, S.; Jones, H.D.; Holdsworth, M.J. Interactions of the developmental regulator ABI3 with proteins identified from developing Arabidopsis seeds. Plant J. 2000, 21, 143–155. [Google Scholar] [CrossRef]
- Ahmad, M.; Grancher, N.; Heil, M.; Black, R.C.; Giovani, B.; Galland, P.; Lardemer, D. Action spectrum for cryptochrome–dependent hypocotyl growth inhibition in Arabidopsis. Plant Physiol. 2002, 129, 774–785. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium—Mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Report. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Cadman, C.S.C.; Toorop, P.E.; Hilhorst, H.W.M.; Finch-Savage, W.E. Gene expression profiles of Arabidopsis Cvi seeds during dormancy cycling indicate a common underlying dormancy control mechanism. Plant J. 2006, 46, 805–822. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, B.J.W.; Willems, L.; Bassel, G.W.; van Bolderen-Veldkamp, R.P.; Ligterink, W.; Hilhorst, H.W.M.; Bentsink, L. Identification of Reference Genes for RT–qPCR Expression Analysis in Arabidopsis and Tomato Seeds. Plant Cell Physiol. 2012, 53, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constant Light Conditions | Treatment | Germination (%) of WT Dormant Seeds after Time of Incubation | |
|---|---|---|---|
| 48 h | 72 h | ||
| BL | H2O | 14.33 ± 0.5 | 44.4 ± 2.5 |
| PAC (1 µM) | 12.0 ± 0.5 | 15.0 ± 1.5 * | |
| ABA (1 µM) | 3.0 ± 0.33 * | 5.0 ± 0.33 ** | |
| Darkness | H2O | 0.7 ± 0.5 | 0.7 ± 0.5 |
| PAC (1 µM) | 2.0 ± 0.12 | 2.0 ± 0.5 | |
| ABA (1 µM) | 0.0 ± 0.0 | 1.0 ± 0.33 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stawska, M.; Oracz, K. phyB and HY5 are Involved in the Blue Light-Mediated Alleviation of Dormancy of Arabidopsis Seeds Possibly via the Modulation of Expression of Genes Related to Light, GA, and ABA. Int. J. Mol. Sci. 2019, 20, 5882. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235882
Stawska M, Oracz K. phyB and HY5 are Involved in the Blue Light-Mediated Alleviation of Dormancy of Arabidopsis Seeds Possibly via the Modulation of Expression of Genes Related to Light, GA, and ABA. International Journal of Molecular Sciences. 2019; 20(23):5882. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235882
Chicago/Turabian StyleStawska, Marlena, and Krystyna Oracz. 2019. "phyB and HY5 are Involved in the Blue Light-Mediated Alleviation of Dormancy of Arabidopsis Seeds Possibly via the Modulation of Expression of Genes Related to Light, GA, and ABA" International Journal of Molecular Sciences 20, no. 23: 5882. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235882