Proteomic Changes during MCMV Infection Revealed by iTRAQ Quantitative Proteomic Analysis in Maize


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
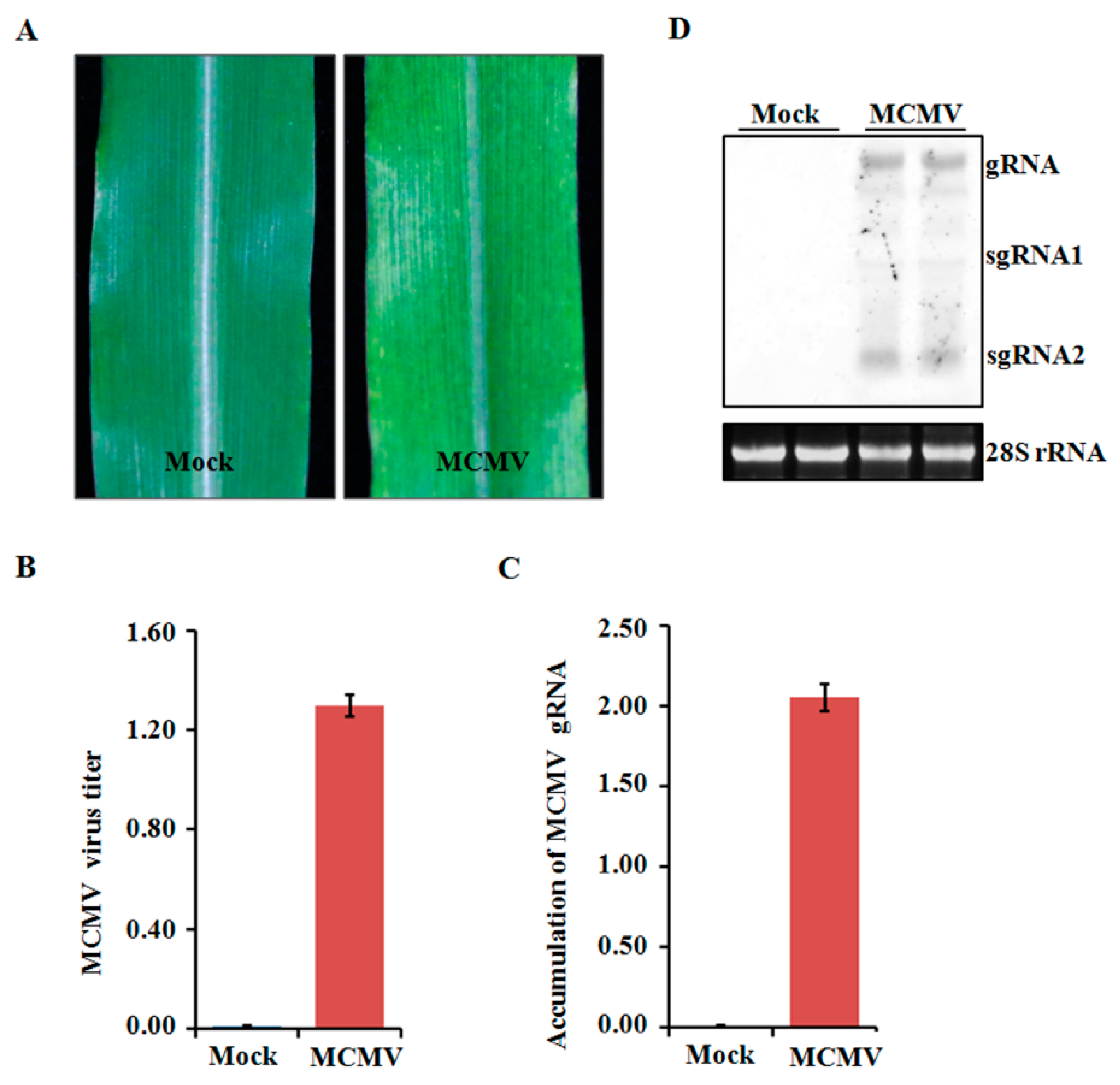
2.1. Phenotype Shown on MCMV-Infected Maize Plants
2.2. iTRAQ-Based Proteomic Analysis
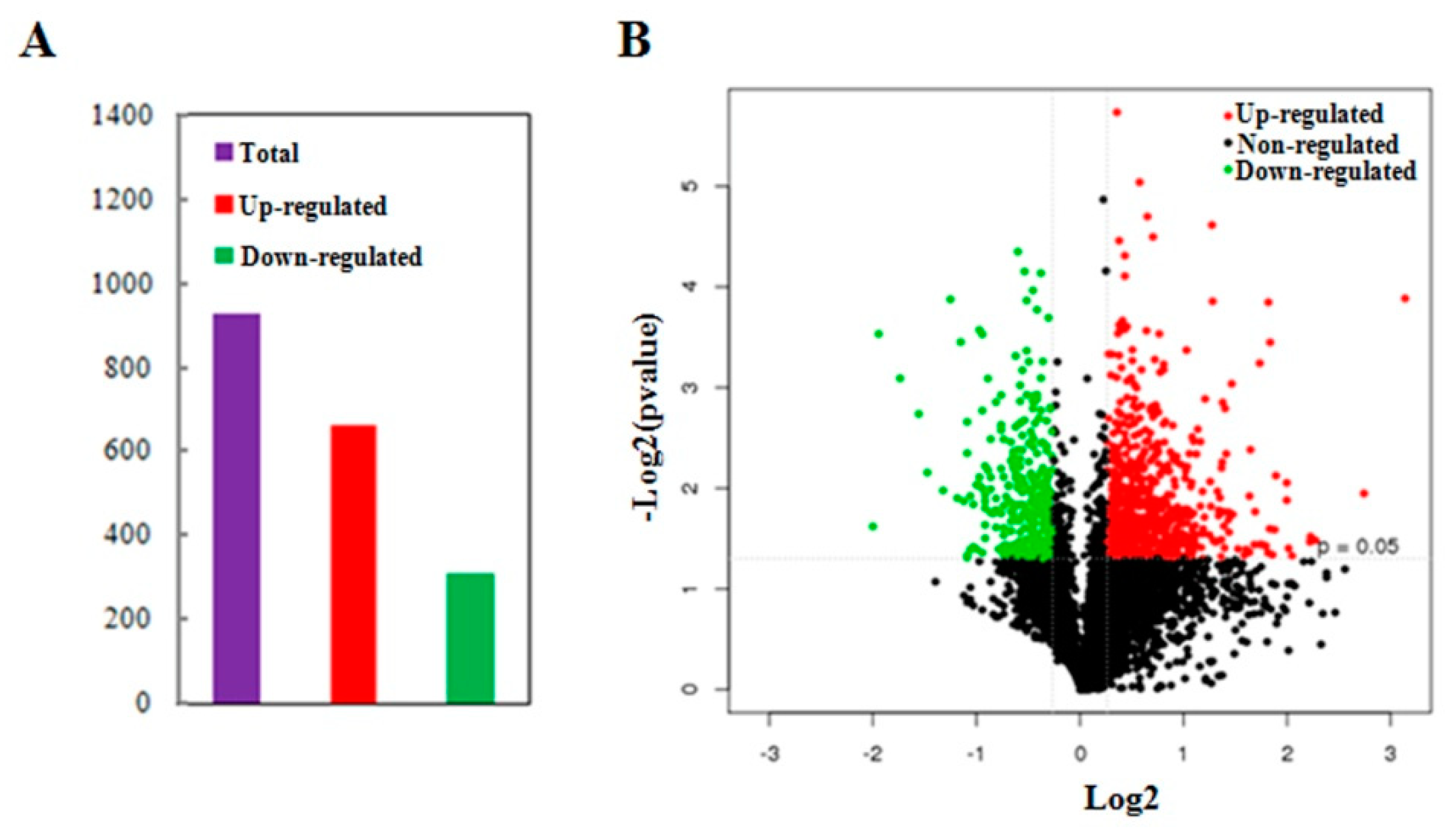
2.3. Impacts of MCMV Infection on the Maize Global Proteome
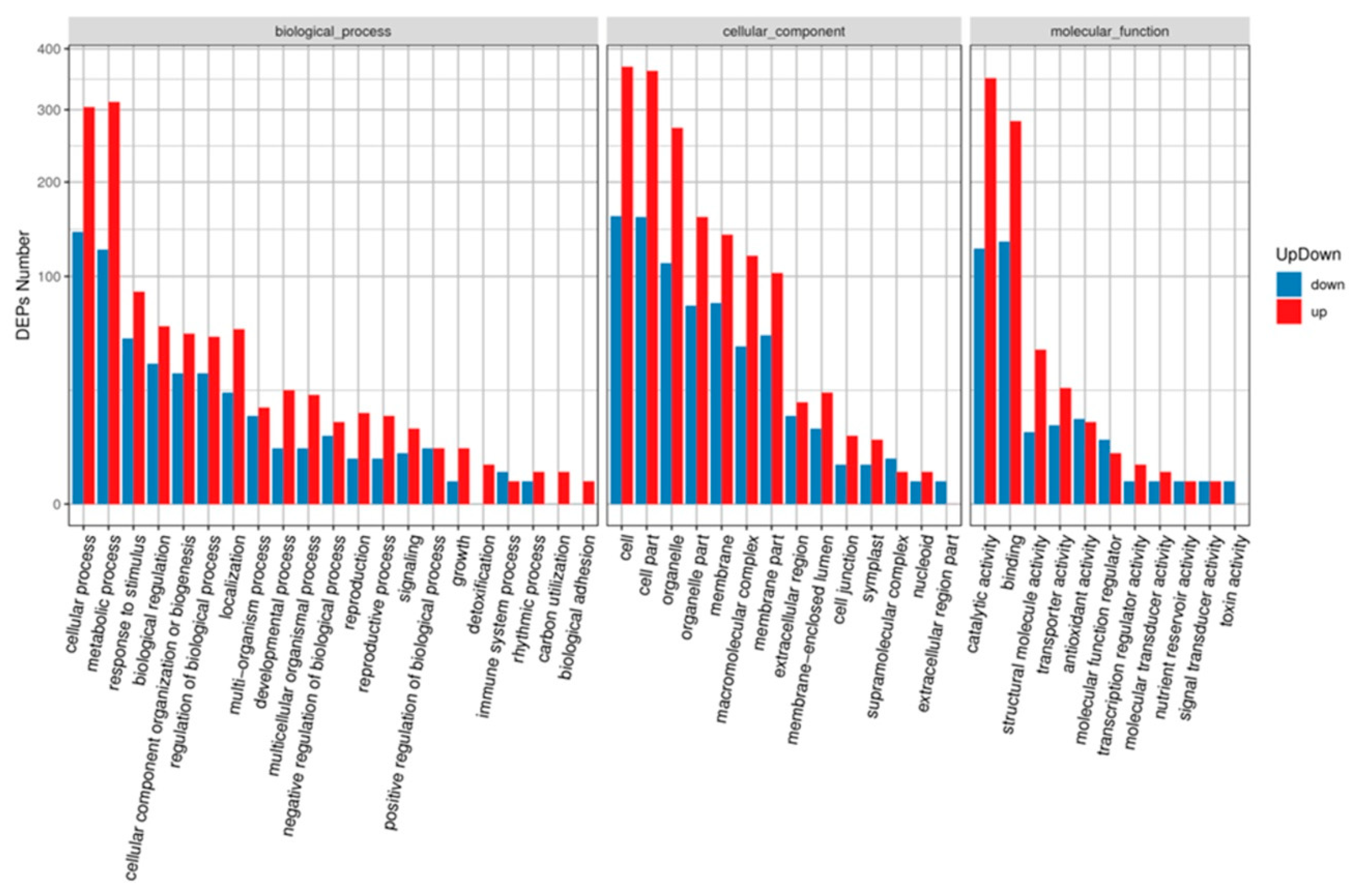
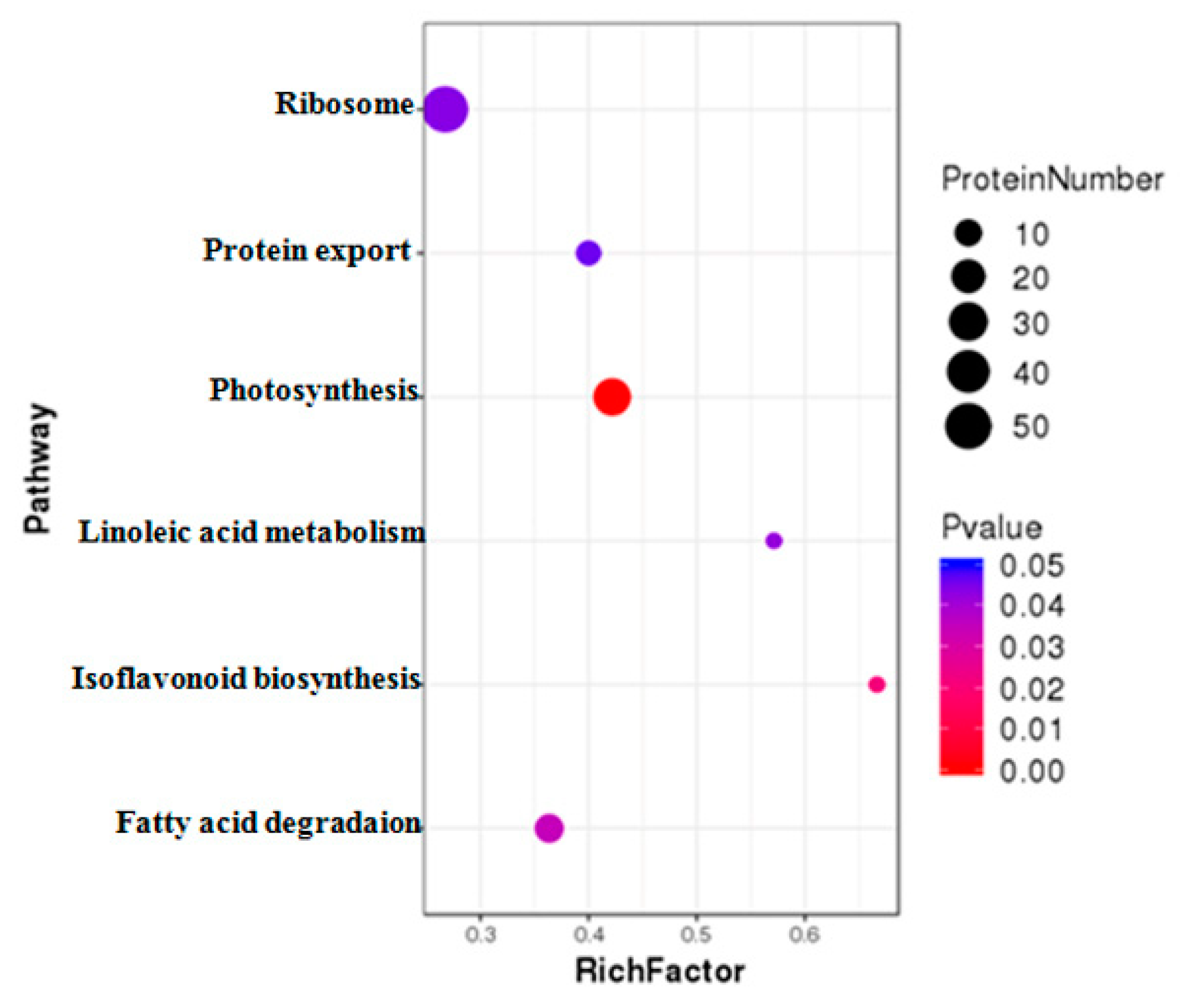
2.4. GO and KEGG Analysis of DAPs under MCMV Infection
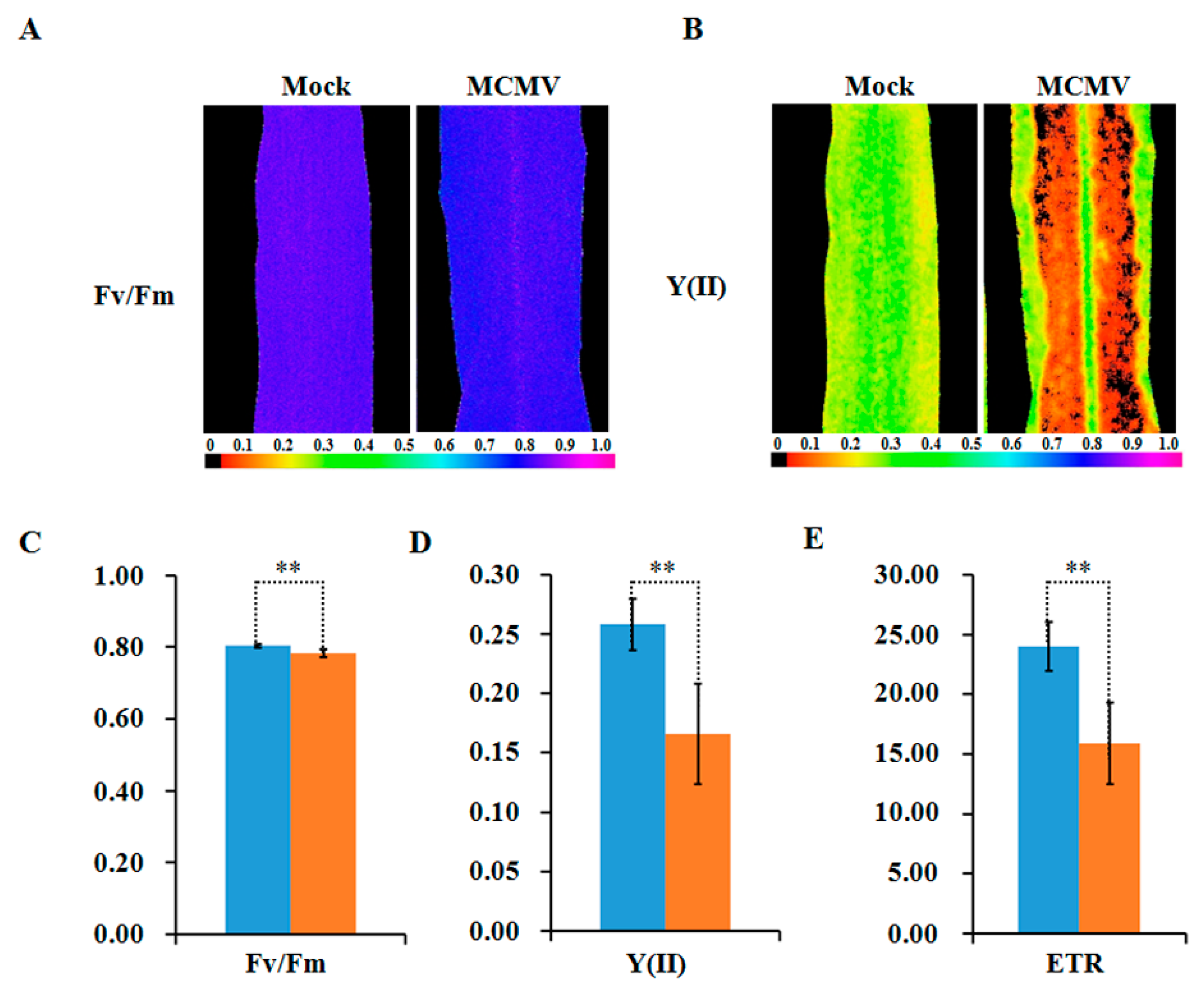
2.5. Photosynthetic Activity Decreased during MCMV Infection
2.6. Alteration of Ribosomal Protein Abundance during MCMV Infection
2.7. Enrichment of Stress Response-Related Proteins during MCMV Infection
2.8. Differential Regulation of Redox Homeostasis Related Proteins
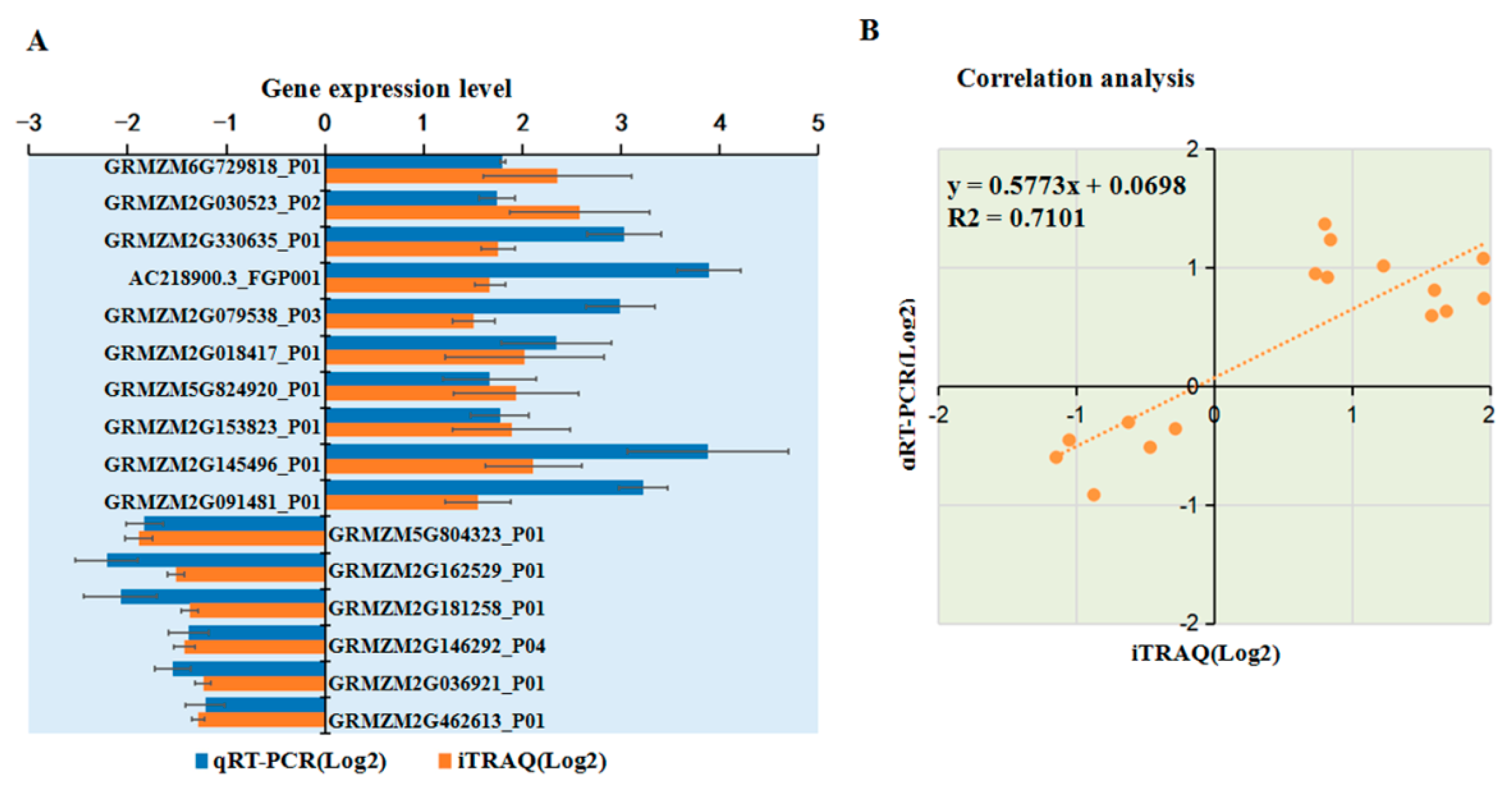
2.9. Transcriptional Level Analysis for Selected DAPs
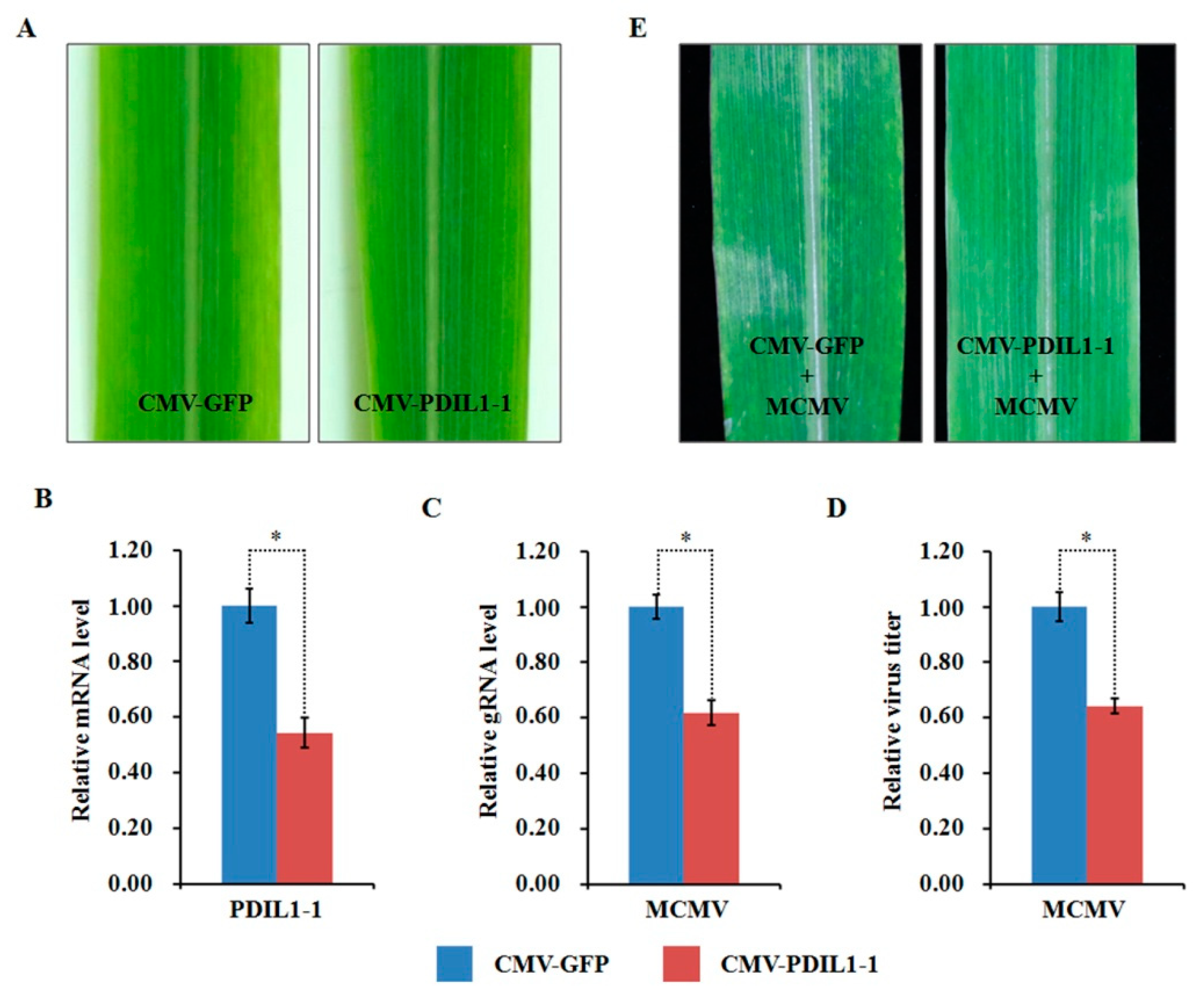
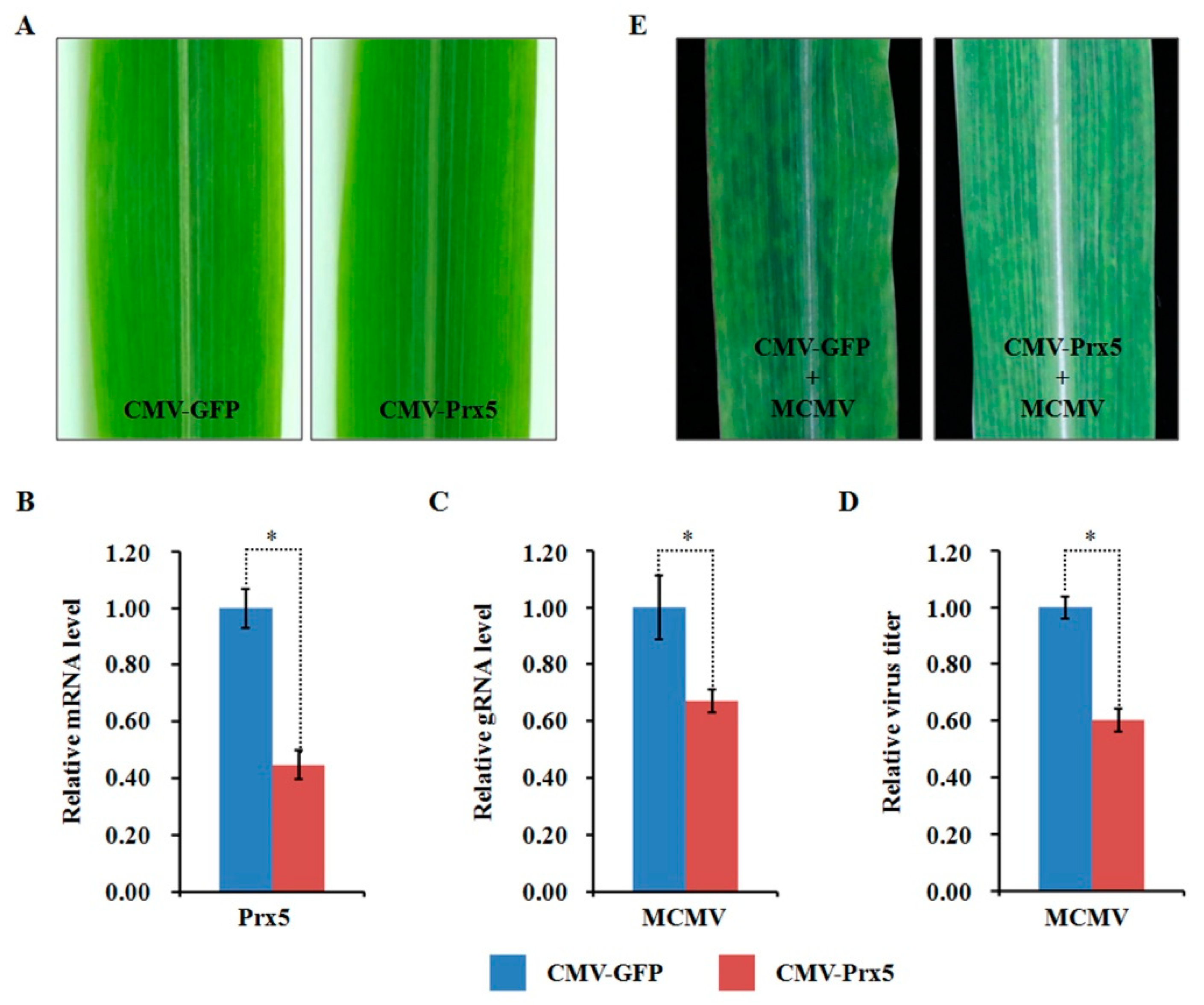
2.10. Functional Analyses of Two Selected DAPs through Virus-Induced Gene Silencing
3. Discussion
3.1. Photosynthesis Decreased during MCMV Infection
3.2. Ribosomal Proteins Commonly Affected by MCMV Infection
3.3. Stress Response-Related Proteins Regulated by MCMV Infection
3.4. Redox Regulation in Maize after MCMV Infection
3.5. ZmPDIL-1 Facilitates MCMV Infection
3.6. ZmPrx5 Is a Positive Regulator of MCMV Infection
4. Materials and Methods
4.1. Plant Growth and Virus Inoculation
4.2. Sample Preparation
4.3. iTRAQ Labeling and Strong-Cation-Exchange (SCX) Fractionation
4.4. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
4.5. Proteomics Data Analysis
4.6. Annotation of the Proteins
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Total RNA Extraction and qRT-PCR Assay
4.9. Measurements of Photosynthetic Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MCMV | chlorotic mottle virus |
| CMV | cucumber mosaic virus |
| SCMV | sugarcane mosaic virus |
| MDMV | maize dwarf mosaic virus |
| Mev | measles Virus |
| sgRNAs | subgenomic RNAs |
| LOP | lipoxygenase |
| ROS | reactive oxygen species |
| HSP | heat shock protein |
| CAT | catalases |
| POD | peroxidases |
| SOD | superoxide dismutases |
| PDI | protein disulfide isomerases |
| Prx | peroxiredoxin protein |
References
- Allie, F.; Rey, M.E.C. Transcriptional alterations in model host, Nicotiana benthamiana, in response to infection by South African cassava mosaic virus. Eur. J. Plant Pathol. 2013, 137, 765–785. [Google Scholar] [CrossRef]
- Hou, H.W.; Hu, Y.; Wang, Q.; Xu, X.B.; Qian, Y.J.; Zhou, X.P. Gene Expression Profiling Shows That NbFDN1 Is Involved in Modulating the Hypersensitive Response-Like Cell Death Induced by the Oat dwarf virus RepA Protein. Mol. Plant Microbe Interact. 2018, 31, 1006–1020. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Luqman, S.; Masood, N.; Singh, D.K.; Saeed, S.T.; Samad, A. Eclipta yellow vein virus enhances chlorophyll destruction, singlet oxygen production and alters endogenous redox status in Andrographis paniculata. Plant Physiol. Biochem. 2016, 104, 165–173. [Google Scholar] [CrossRef]
- Li, J.J.; Henty-Ridilla, J.L.; Staiger, B.H.; Day, B.; Staiger, C.J. Capping protein integrates multiple MAMP signalling pathways to modulate actin dynamics during plant innate immunity. Nat. Commun. 2015, 6, 7206. [Google Scholar] [CrossRef] [Green Version]
- Pusztahelyi, T.; Holb, I.J.; Pocsi, I. Secondary metabolites in fungus-plant interactions. Front. Plant Sci. 2015, 6, 573. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Langlois-Meurinne, M.; Bellvert, F.; Garmier, M.; Didierlaurent, L.; Massoud, K.; Chaouch, S.; Marie, A.; Bodo, B.; Kauffmann, S.; et al. The differential spatial distribution of secondary metabolites in Arabidopsis leaves reacting hypersensitively to Pseudomonas syringae pv. tomato is dependent on the oxidative burst. J. Exp. Bot. 2010, 61, 3355–3370. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Qin, L.; Liu, G.S.; Peremyslov, V.V.; Dolja, V.V.; Wei, Y.D. Myosins XI modulate host cellular responses and penetration resistance to fungal pathogens. Proc. Natl. Acad. Sci. USA 2014, 111, 13996–14001. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.J.; He, B.; Chen, L.; Xiao, S.Q.; Fu, J.; Chen, Y.; Yu, T.Q.; Cheng, Z.Q.; Feng, H. Transcriptome analysis confers a complex disease resistance network in wild rice Oryza meyeriana against Xanthomonas oryzae pv. oryzae. Sci. Rep. 2016, 6, 38215. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.C.; Xu, L.P.; Wang, Z.Q.; Peng, Q.; Yang, Y.T.; Chen, Y.; Que, Y.X. Comparative proteomics reveals that central metabolism changes are associated with resistance against Sporisorium scitamineum in sugarcane. BMC Genom. 2016, 17, 800. [Google Scholar] [CrossRef] [Green Version]
- Das, P.P.; Chua, G.M.; Lin, Q.S.; Wong, S.M. iTRAQ-based analysis of leaf proteome identifies important proteins in secondary metabolite biosynthesis and defence pathways crucial to cross-protection against TMV. J. Proteom. 2019, 196, 42–56. [Google Scholar] [CrossRef]
- Yue, R.Q.; Lu, C.X.; Han, X.H.; Guo, S.L.; Yan, S.F.; Liu, L.; Fu, X.L.; Chen, N.N.; Guo, X.H.; Chi, H.F.; et al. Comparative proteomic analysis of maize (Zea mays L.) seedlings under rice black-streaked dwarf virus infection. BMC Plant Biol. 2018, 18, 191. [Google Scholar] [CrossRef] [Green Version]
- Achon, M.A.; Subira, J.; Sin, E. Seasonal occurrence of Laodelphax striatellus in Spain: Effect on the incidence of Maize rough dwarf virus. Crop Prot. 2013, 47, 1–5. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, C.; Wang, C.Y.; Qian, Y.J.; Li, Z.H.; Hong, J.; Zhou, X.P. Further characterization of Maize chlorotic mottle virus and its synergistic interaction with Sugarcane mosaic virus in maize. Sci. Rep. 2017, 7, 39960. [Google Scholar] [CrossRef]
- Scheets, K. Maize chlorotic mottle machlomovirus and wheat streak mosaic rymovirus concentrations increase in the synergistic disease corn lethal necrosis. Virology 1998, 242, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Nutter, R.C.; Scheets, K.; Panganiban, L.C.; Lommel, S.A. The Complete Nucleotide-Sequence of the Maize Chlorotic Mottle Virus Genome. Nucleic Acids Res. 1989, 17, 3163–3177. [Google Scholar] [CrossRef] [Green Version]
- Scheets, K. Analysis of gene functions in Maize chlorotic mottle virus. Virus Res. 2016, 222, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Scheets, K. Maize chlorotic mottle machlomovirus expresses its coat protein from a 1.47-kb subgenomic RNA and makes a 0.34-kb subgenomic RNA. Virology 2000, 267, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Boulila, M. Positive selection, molecular recombination structure and phylogenetic reconstruction of members of the family Tombusviridae: Implication in virus taxonomy. Genet. Mol. Biol. 2011, 34, 647–660. [Google Scholar] [CrossRef] [Green Version]
- Scheets, K.; Jordan, R.; White, K.A.; Hernandez, C. Pelarspovirus, a proposed new genus in the family Tombusviridae. Arch. Virol. 2015, 160, 2385–2393. [Google Scholar] [CrossRef] [Green Version]
- Shi, F.M.; Zhang, Y.H.; Wang, K.Q.; Meng, Q.L.; Liu, X.L.; Ma, L.G.; Li, Y.C.; Liu, J.; Ma, L. Expression profile analysis of maize in response to Setosphaeria turcica. Gene 2018, 659, 100–108. [Google Scholar] [CrossRef]
- Wang, Y.P.; Zhou, Z.J.; Gao, J.Y.; Wu, Y.B.; Xia, Z.L.; Zhang, H.Y.; Wu, J.Y. The Mechanisms of Maize Resistance to Fusarium verticillioides by Comprehensive Analysis of RNA-seq Data. Front. Plant Sci. 2016, 7, 1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, N.; Zhan, B.H.; Zhou, X.P. Nitric Oxide as a Downstream Signaling Molecule in Brassinosteroid-Mediated Virus Susceptibility to Maize Chlorotic Mottle Virus in Maize. Viruses 2019, 11, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kebede, A.Z.; Johnston, A.; Schneiderman, D.; Bosnich, W.; Harris, L.J. Transcriptome profiling of two maize inbreds with distinct responses to Gibberella ear rot disease to identify candidate resistance genes. BMC Genom. 2018, 19, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, J.; Berger, D.K.; Christensen, S.A.; Murray, S.L. RNA-Seq analysis of resistant and susceptible sub-tropical maize lines reveals a role for kauralexins in resistance to grey leaf spot disease, caused by Cercospora zeina. BMC Plant Biol. 2017, 17, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.J.; Han, Z.P.; Wang, S.X.; Wang, X.T.; Sun, A.G.; Zu, X.F.; Chen, Y.H. Comparative proteomic analysis of the plant-virus interaction in resistant and susceptible ecotypes of maize infected with sugarcane mosaic virus. J. Proteom. 2013, 89, 124–140. [Google Scholar] [CrossRef]
- Chen, H.; Cao, Y.Y.; Li, Y.Q.; Xia, Z.H.; Xie, J.P.; Carr, J.P.; Wu, B.M.; Fan, Z.F.; Zhou, T. Identification of differentially regulated maize proteins conditioning Sugarcane mosaic virus systemic infection. New Phytol. 2017, 215, 1156–1172. [Google Scholar] [CrossRef] [Green Version]
- Mostafavi, F.S.; Sabbagh, S.K.; Yamchi, A.; Nasrollanejad, S.; Panjehkeh, N. Differential molecular response of maize and Johnson grass against maize dwarf mosaic virus and bermuda grass southern mosaic virus. Acta Virol. 2019, 63, 70–79. [Google Scholar] [CrossRef]
- Zhao, J.P.; Zhang, X.; Hong, Y.G.; Liu, Y.L. Chloroplast in Plant-Virus Interaction. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.F.; Wu, J.X.; Lu, L.N.; Xu, Y.; Zhou, X.P. Interaction between Rice stripe virus Disease-Specific Protein and Host PsbP Enhances Virus Symptoms. Mol. Plant 2014, 7, 691–708. [Google Scholar] [CrossRef] [Green Version]
- Graf, M.; Arenz, S.; Huter, P.; Donhofer, A.; Novacek, J.; Wilson, D.N. Cryo-EM structure of the spinach chloroplast ribosome reveals the location of plastid-specific ribosomal proteins and extensions. Nucleic Acids Res. 2017, 45, 2887–2896. [Google Scholar] [CrossRef]
- Mower, J.P.; Bonen, L. Ribosomal protein L10 is encoded in the mitochondrial genome of many land plants and green algae. BMC Evol. Biol. 2009, 9, 265. [Google Scholar] [CrossRef] [PubMed]
- Gleason, C.; Huang, S.B.; Thatcher, L.F.; Foley, R.C.; Anderson, C.R.; Carroll, A.J.; Millar, A.H.; Singh, K.B. Mitochondrial complex II has a key role in mitochondrial-derived reactive oxygen species influence on plant stress gene regulation and defense. Proc. Natl. Acad. Sci. USA 2011, 108, 10768–10773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braidwood, L.; Quito-Avila, D.F.; Cabanas, D.; Bressan, A.; Wangai, A.; Baulcombe, D.C. Maize chlorotic mottle virus exhibits low divergence between differentiated regional sub-populations. Sci. Rep. 2018, 8, 1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.W.; Penning, B.W.; Jamann, T.M.; Glaubitz, J.C.; Romay, C.; Buckler, E.S.; Redinbaugh, M.G. Diverse Chromosomal Locations of Quantitative Trait Loci for Tolerance to Maize chlorotic mottle virus in Five Maize Populations. Phytopathology 2018, 108, 748–758. [Google Scholar] [CrossRef] [Green Version]
- Das, P.P.; Lin, Q.S.; Wong, S.M. Comparative proteomics of Tobacco mosaic virus-infected Nicotiana tabacum plants identified major host proteins involved in photosystems and plant defence. J. Proteom. 2019, 194, 191–199. [Google Scholar] [CrossRef]
- Novakova, S.; Danchenko, M.; Skultety, L.; Fialova, I.; Leskova, A.; Beke, G.; Flores-Ramirez, G.; Glasa, M. Photosynthetic and Stress Responsive Proteins Are Altered More Effectively in Nicotiana benthamiana Infected with Plum pox virus Aggressive PPV-CR versus Mild PPV-C Cherry-Adapted Isolates. J. Proteome Res. 2018, 17, 3114–3127. [Google Scholar] [CrossRef]
- Wrzesinska, B.; Vu, L.D.; Gevaert, K.; De Smet, I.; Obrepalska-Steplowska, A. Peanut Stunt Virus and Its Satellite RNA Trigger Changes in Phosphorylation in N-benthamiana Infected Plants at the Early Stage of the Infection. Int. J. Mol. Sci. 2018, 19, 3223. [Google Scholar] [CrossRef] [Green Version]
- Di Carli, M.; Villani, M.E.; Bianco, L.; Lombardi, R.; Perrotta, G.; Benvenuto, E.; Donini, M. Proteomic Analysis of the Plant-Virus Interaction in Cucumber Mosaic Virus (CMV) Resistant Transgenic Tomato. J. Proteome Res. 2010, 9, 5684–5697. [Google Scholar] [CrossRef]
- Gnanasekaran, P.; Ponnusamy, K.; Chakraborty, S. A geminivirus betasatellite encoded beta C1 protein interacts with PsbP and subverts PsbP-mediated antiviral defence in plants. Mol. Plant Pathol. 2019, 20, 943–960. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Herva, J.J.; Gonzalez-Melendi, P.; Cuartas-Lanza, R.; Antunez-Lamas, M.; Rio-Alvarez, I.; Li, Z.; Lopez-Torrejon, G.; Diaz, I.; del Pozo, J.C.; Chakravarthy, S.; et al. A bacterial cysteine protease effector protein interferes with photosynthesis to suppress plant innate immune responses. Cell. Microbiol. 2012, 14, 669–681. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Li, X.; Zhou, Y.J. Ribosomal protein L18 is an essential factor that promote rice stripe virus accumulation in small brown planthopper. Virus Res. 2018, 247, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.; Daros, J.A. Tobacco Etch Virus Protein P1 Traffics to the Nucleolus and Associates with the Host 60S Ribosomal Subunits during Infection. J. Virol. 2014, 88, 10725–10737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, C.M.; Santos, A.A.; Pires, S.R.; Rocha, C.S.; Saraiva, D.I.; Machado, J.P.B.; Mattos, E.C.; Fietto, L.G.; Fontes, E.P.B. Regulated Nuclear Trafficking of rpL10A Mediated by NIK1 Represents a Defense Strategy of Plant Cells against Virus. PLoS Pathog. 2008, 4, e1000247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraj, S.; Senthil-Kumar, M.; Ramu, V.S.; Wang, K.R.; Mysore, K.S. Plant Ribosomal Proteins, RPL12 and RPL19, Play a Role in Nonhost Disease Resistance against Bacterial Pathogens. Front. Plant Sci. 2016, 6, 1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.C.; Schwab, W. Cloning and characterization of a 9-lipoxygenase gene induced by pathogen attack from Nicotiana benthamiana for biotechnological application. BMC Biotechnol. 2011, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- Christensen, S.A.; Nemchenko, A.; Park, Y.S.; Borrego, E.; Huang, P.C.; Schmelz, E.A.; Kunze, S.; Feussner, I.; Yalpani, N.; Meeley, R.; et al. The Novel Monocot-Specific 9-Lipoxygenase ZmLOX12 Is Required to Mount an Effective Jasmonate-Mediated Defense against Fusarium verticillioides in Maize. Mol. Plant Microbe Interact. 2014, 27, 1263–1276. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, R.; Citores, L.; Ragucci, S.; Russo, R.; Di Maro, A.; Ferreras, J.M. Biological and antipathogenic activities of ribosome-inactivating proteins from Phytolacca dioica L. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 1256–1264. [Google Scholar] [CrossRef]
- Park, C.J.; Seo, Y.S. Heat Shock Proteins: A Review of the Molecular Chaperones for Plant Immunity. Plant Pathol. J. 2015, 31, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.L.; Liu, R.X.; Li, Y.H.; Wang, W.; Tai, F.J.; Xue, R.L.; Li, C.H. Heat shock protein 70 regulates the abscisic acid-induced antioxidant response of maize to combined drought and heat stress. Plant Growth Regul. 2010, 60, 225–235. [Google Scholar] [CrossRef]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant. 2010, 138, 414–429. [Google Scholar] [CrossRef]
- Xu, E.J.; Brosche, M. Salicylic acid signaling inhibits apoplastic reactive oxygen species signaling. BMC Plant Biol. 2014, 14, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindermayr, C.; Durner, J. Interplay of Reactive Oxygen Species and Nitric Oxide: Nitric Oxide Coordinates Reactive Oxygen Species Homeostasis. Plant Physiol. 2015, 167, 1209–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, F.; Mizoguchi, T.; Yoshida, R.; Ichimura, K.; Shinozaki, K. Calmodulin-Dependent Activation of MAP Kinase for ROS Homeostasis in Arabidopsis. Mol. Cell 2011, 41, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Magbanua, Z.V.; De Moraes, C.M.; Brooks, T.D.; Williams, W.P.; Luthe, D.S. Is catalase activity one of the factors associated with maize resistance to Aspergillus flavus? Mol. Plant Microbe Interact. 2007, 20, 697–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, P.; Ibanez, S.G.; Agostini, E.; Etcheverry, M. Effects of maize inoculation with Fusarium verticillioides and with two bacterial biocontrol agents on seedlings growth and antioxidative enzymatic activities. Appl. Soil Ecol. 2011, 51, 52–59. [Google Scholar] [CrossRef]
- Huang, X.L.; Liu, L.X.; Chen, J.; Zhai, Y.H. Comparative proteomic analysis of the response in resistant and susceptible maize inbred lines to infection by Curvularia lunata. Prog. Nat. Sci. Mater. Int. 2009, 19, 845–850. [Google Scholar] [CrossRef]
- Turano, C.; Coppari, S.; Altieri, F.; Ferraro, A. Proteins of the PDI family: Unpredicted non-ER locations and functions. J. Cell. Physiol. 2002, 193, 154–163. [Google Scholar] [CrossRef]
- Kamauchi, S.; Wadahama, H.; Iwasaki, K.; Nakamoto, Y.; Nishizawa, K.; Ishimoto, M.; Kawada, T.; Urade, R. Molecular cloning and characterization of two soybean protein disulfide isomerases as molecular chaperones for seed storage proteins. FEBS J. 2008, 275, 2644–2658. [Google Scholar] [CrossRef]
- Rawarak, N.; Suttitheptumrong, A.; Reamtong, O.; Boonnak, K.; Pattanakitsakul, S.N. Protein Disulfide Isomerase Inhibitor Suppresses Viral Replication and Production during Antibody-Dependent Enhancement of Dengue Virus Infection in Human Monocytic Cells. Viruses 2019, 11, 155. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Chang, K.O. Protein disulfide isomerases as potential therapeutic targets for influenza A and B viruses. Virus Res. 2018, 247, 26–33. [Google Scholar] [CrossRef]
- Oono, Y.; Wakasa, Y.; Hirose, S.; Yang, L.J.; Sakuta, C.; Takaiwa, F. Analysis of ER stress in developing rice endosperm accumulating beta-amyloid peptide. Plant Biotechnol. J. 2010, 8, 691–718. [Google Scholar] [CrossRef] [PubMed]
- Caplan, J.L.; Zhu, X.H.; Mamillapalli, P.; Marathe, R.; Anandalakshmi, R.; Dinesh-Kumar, S.P. Induced ER Chaperones Regulate a Receptor-like Kinase to Mediate Antiviral Innate Immune Response, in Plants. Cell Host Microbe 2009, 6, 457–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Lupken, T.; Habekuss, A.; Hensel, G.; Steuernagel, B.; Kilian, B.; Ariyadasa, R.; Himmelbach, A.; Kumlehn, J.; Scholz, U.; et al. PROTEIN DISULFIDE ISOMERASE LIKE 5-1 is a susceptibility factor to plant viruses. Proc. Natl. Acad. Sci. USA 2014, 111, 2104–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pena-Ahumada, A.; Kahmann, U.; Dietz, K.J.; Baier, M. Regulation of peroxiredoxin expression versus expression of Halliwell-Asada-Cycle enzymes during early seedling development of Arabidopsis thaliana. Photosynth. Res. 2006, 89, 99–112. [Google Scholar] [CrossRef]
- Watanabe, A.; Yoneda, M.; Ikeda, F.; Sugai, A.; Sato, H.; Kai, C. Peroxiredoxin 1 Is Required for Efficient Transcription and Replication of Measles Virus. J. Virol. 2011, 85, 2247–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Gan, X.; Ito, M.; Chen, M.; Aly, H.H.; Matsui, C.; Abe, T.; Watashi, K.; Wakita, T.; Suzuki, T.; et al. Peroxiredoxin 1, a Novel HBx-Interacting Protein, Interacts with Exosome Component 5 and Negatively Regulates Hepatitis B Virus (HBV) Propagation through Degradation of HBV RNA. J. Virol. 2019, 93, e02203-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.H.; Xu, N.; Li, X.; Jin, W.W.; Tian, Q.; Gu, S.Y.; Sun, G.Y. Overexpression of 2-Cys Prx Increased Salt Tolerance of Photosystem. in Tobacco. Int. J. Agric. Biol. 2017, 19, 735–745. [Google Scholar] [CrossRef]
- Bhatt, I.; Tripathi, B.N. Plant peroxiredoxins: Catalytic mechanisms, functional significance and future perspectives. Biotechnol. Adv. 2011, 29, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Alam, I.; Lee, K.W.; Sharmin, S.A.; Kwak, S.S.; Lee, S.Y.; Lee, B.H. Enhanced tolerance of transgenic tall fescue plants overexpressing 2-Cys peroxiredoxin against methyl viologen and heat stresses. Biotechnol. Lett. 2010, 32, 571–576. [Google Scholar] [CrossRef]
- Kiba, A.; Nishihara, M.; Tsukatani, N.; Nakatsuka, T.; Kato, Y.; Yamamura, S. A peroxiredoxin Q homolog from gentians is involved in both resistance against fungal disease and oxidative stress. Plant Cell Physiol. 2005, 46, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Sheoran, I.S.; Ross, A.R.S.; Olson, D.J.H.; Sawhney, V.K. Compatibility of plant protein extraction methods with mass spectrometry for proteome analysis. Plant Sci. 2009, 176, 99–104. [Google Scholar] [CrossRef]
- Lau, E.; Lam, M.P.Y.; Siu, S.O.; Kong, R.P.W.; Chan, W.L.; Zhou, Z.J.; Huang, J.R.; Lo, C.; Chu, I.K. Combinatorial use of offline SCX and online RP-RP liquid chromatography for iTRAQ-based quantitative proteomics applications. Mol. Biosyst. 2011, 7, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.X.; Wang, Q.; Liu, H.; Qian, Y.J.; Xie, Y.; Zhou, X.P. Monoclonal antibody-based serological methods for maize chlorotic mottle virus detection in China. J. Zhejiang Univ. Sci. B 2013, 14, 555–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.X.; Chen, Z.X.; Zhou, X.P. Production of a monoclonal antibody to Sugarcane mosaic virus and its application for virus detection in China. J. Phytopathol. 2003, 151, 361–364. [Google Scholar] [CrossRef]
- Wang, R.; Yang, X.X.; Wang, N.; Liu, X.D.; Nelson, R.S.; Li, W.M.; Fan, Z.F.; Zhou, T. An efficient virus-induced gene silencing vector for maize functional genomics research. Plant J. 2016, 86, 102–115. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, M.; Cheng, Q.; Hu, Y.; Wu, J.; Zhou, X.; Qian, Y. Proteomic Changes during MCMV Infection Revealed by iTRAQ Quantitative Proteomic Analysis in Maize. Int. J. Mol. Sci. 2020, 21, 35. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010035
Dang M, Cheng Q, Hu Y, Wu J, Zhou X, Qian Y. Proteomic Changes during MCMV Infection Revealed by iTRAQ Quantitative Proteomic Analysis in Maize. International Journal of Molecular Sciences. 2020; 21(1):35. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010035
Chicago/Turabian StyleDang, Mingqing, Qi Cheng, Ya Hu, Jianxiang Wu, Xueping Zhou, and Yajuan Qian. 2020. "Proteomic Changes during MCMV Infection Revealed by iTRAQ Quantitative Proteomic Analysis in Maize" International Journal of Molecular Sciences 21, no. 1: 35. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010035