Transcriptomic Analyses of Camellia oleifera ‘Huaxin’ Leaf Reveal Candidate Genes Related to Long-Term Cold Stress


Abstract
:1. Introduction
2. Results
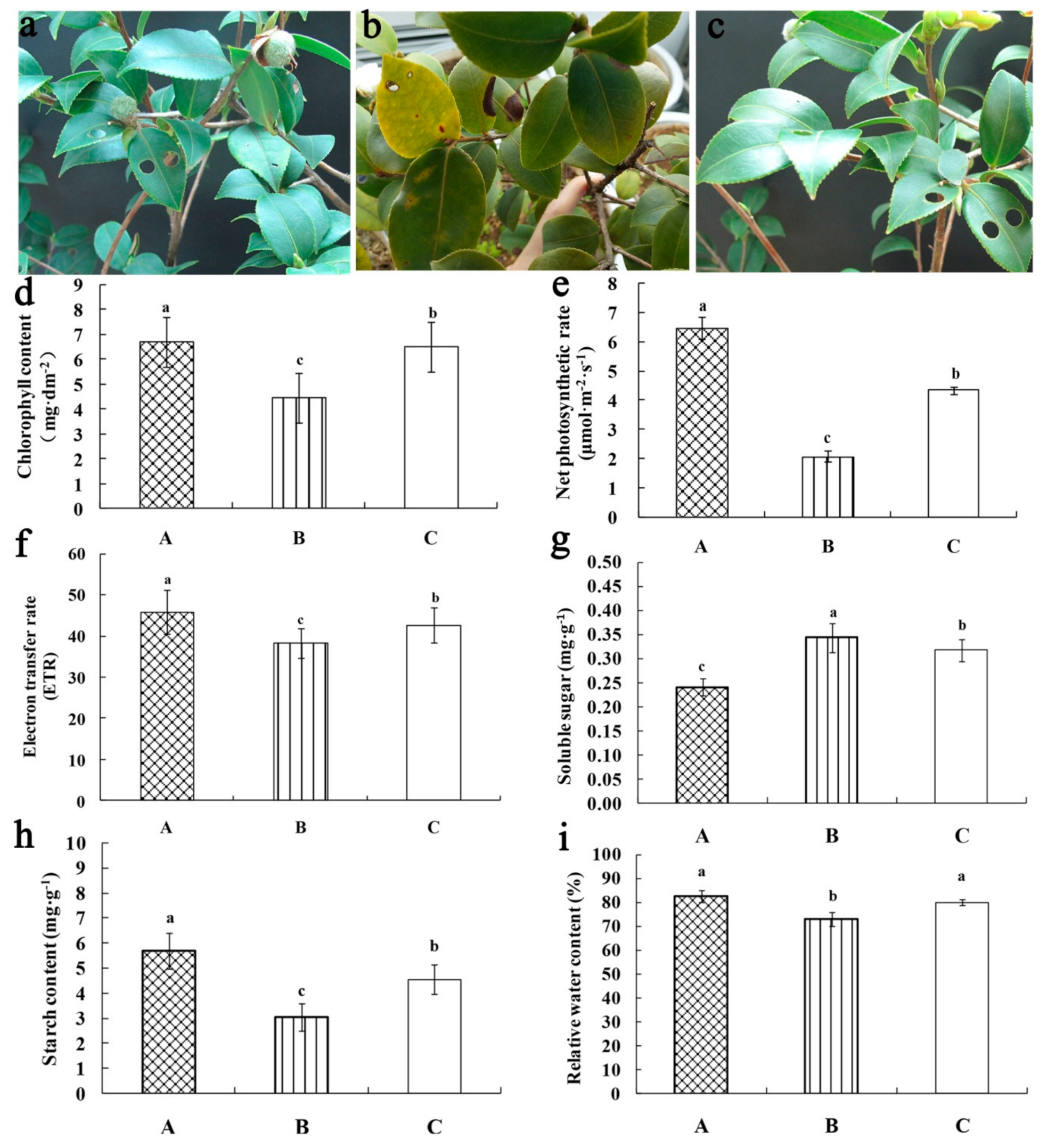
2.1. Morphological Observation and Physiological Indexes of C. oleifera ‘Huaxin’ Under Three Temperatures
2.2. Transcriptome Sequencing and Assembly
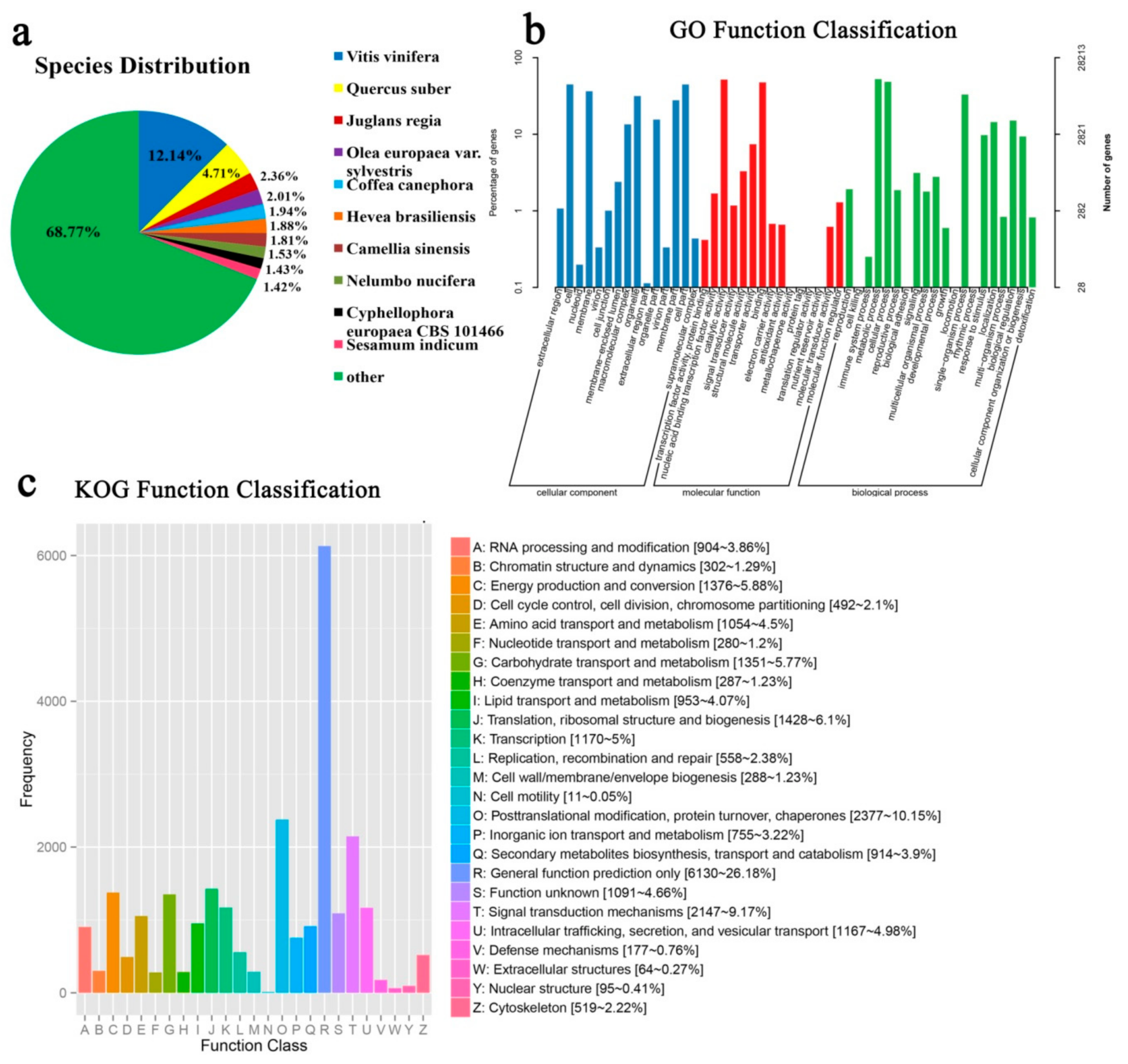
2.3. Gene Annotation and Functional Classification
2.4. Comparative Analyses of Differentially Expressed Genes (DEGs)
2.5. Enriched KEGG Pathway Analyses of DEGs in Response to Low Temperature
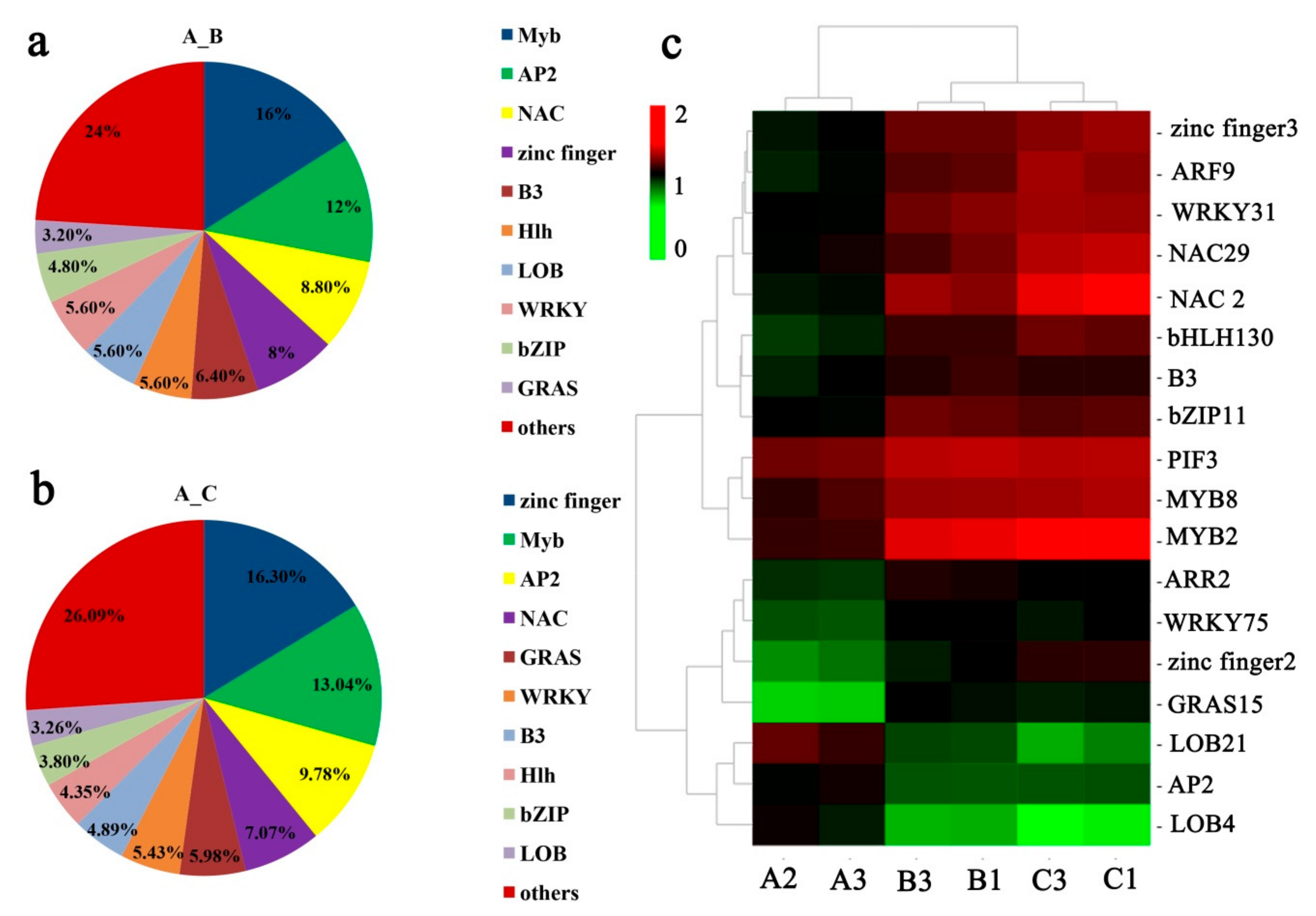
2.6. TFs and PK Responding to Low Temperature
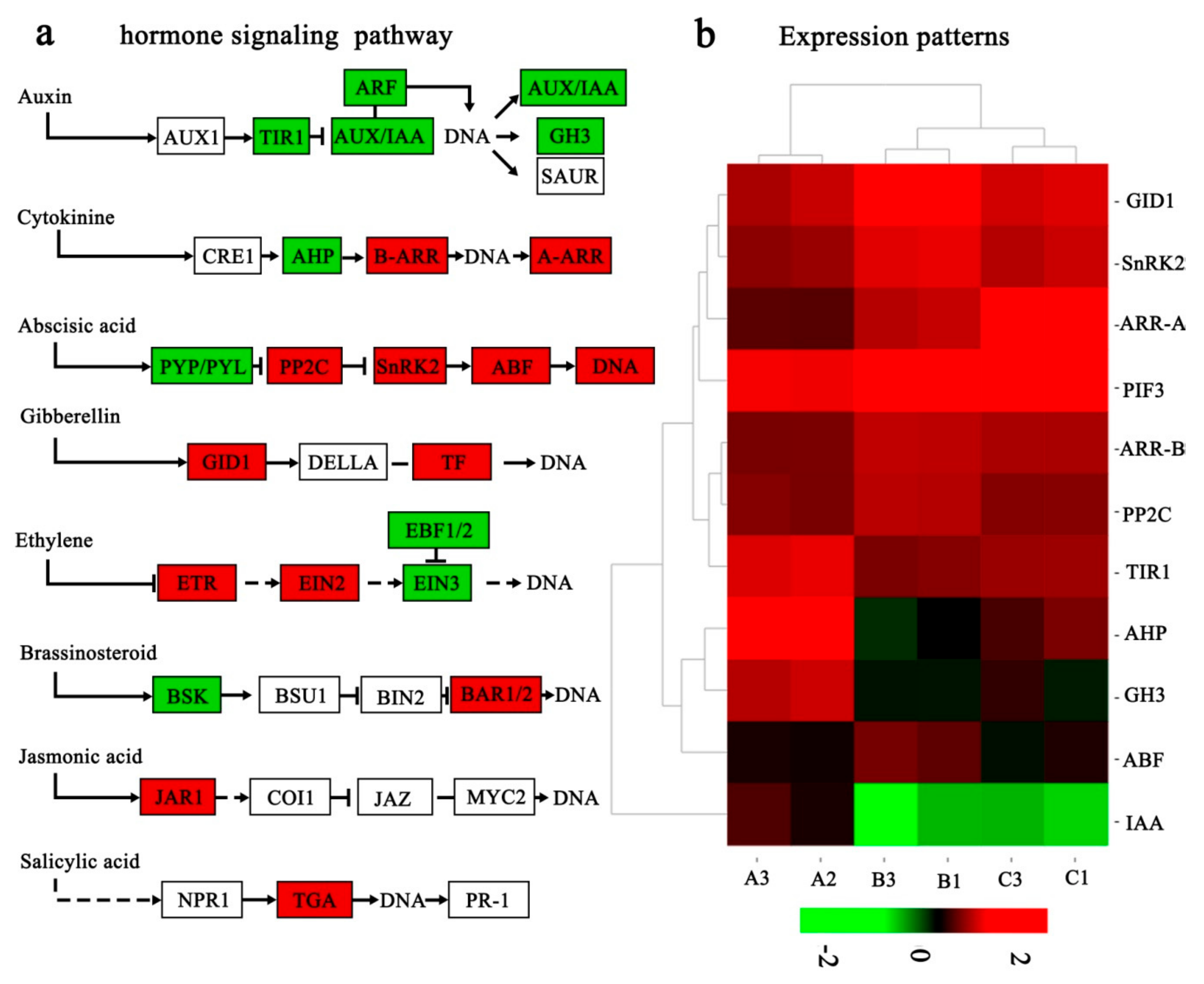
2.7. Hormone-Related DEGs Responding to Low Temperature
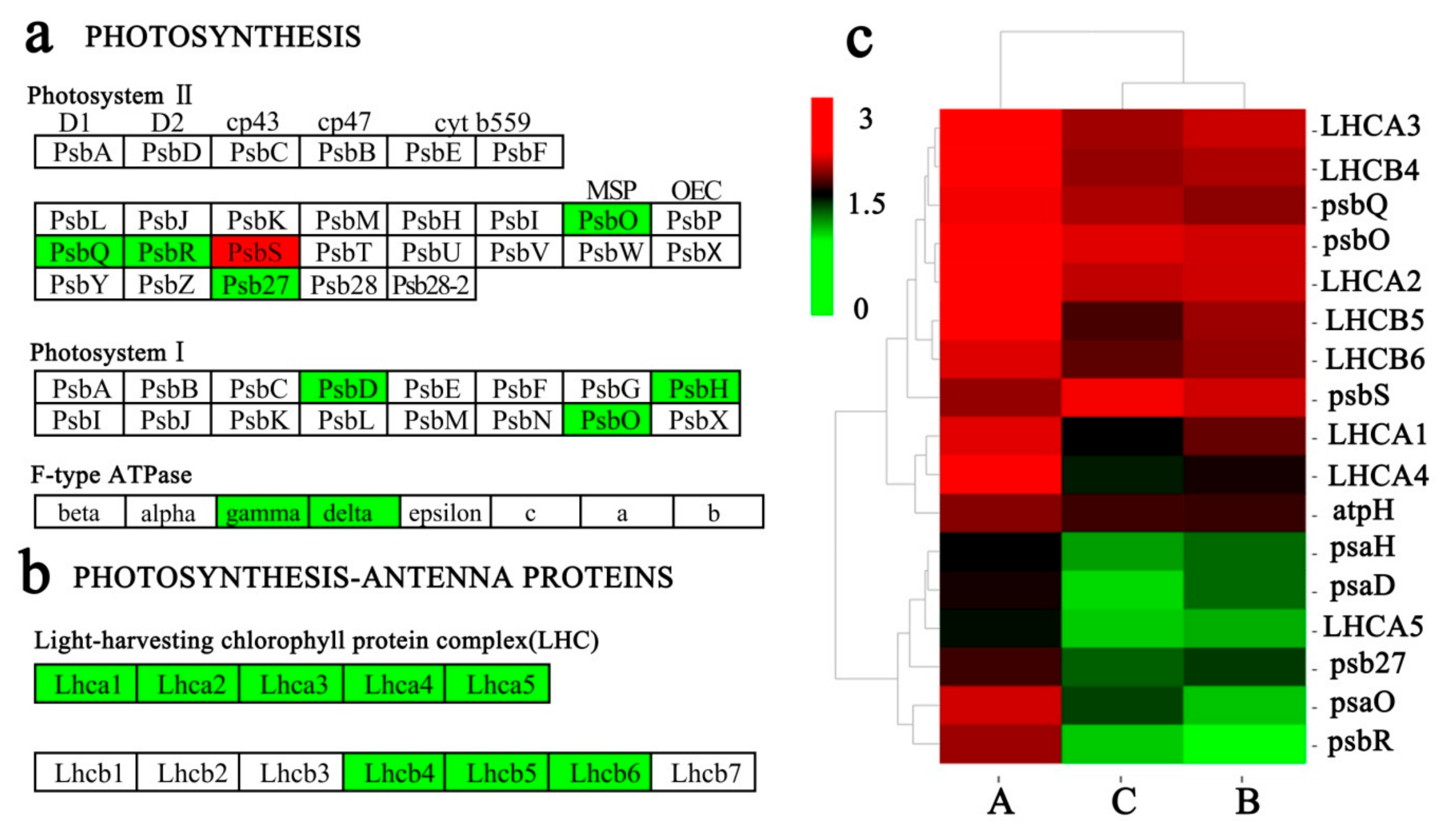
2.8. Key DEGs Related to Photosynthesis
2.9. Key DEGs Related to the Membrane System and Osmoregulation
2.10. Other Important DEGs Responding to Low Temperature
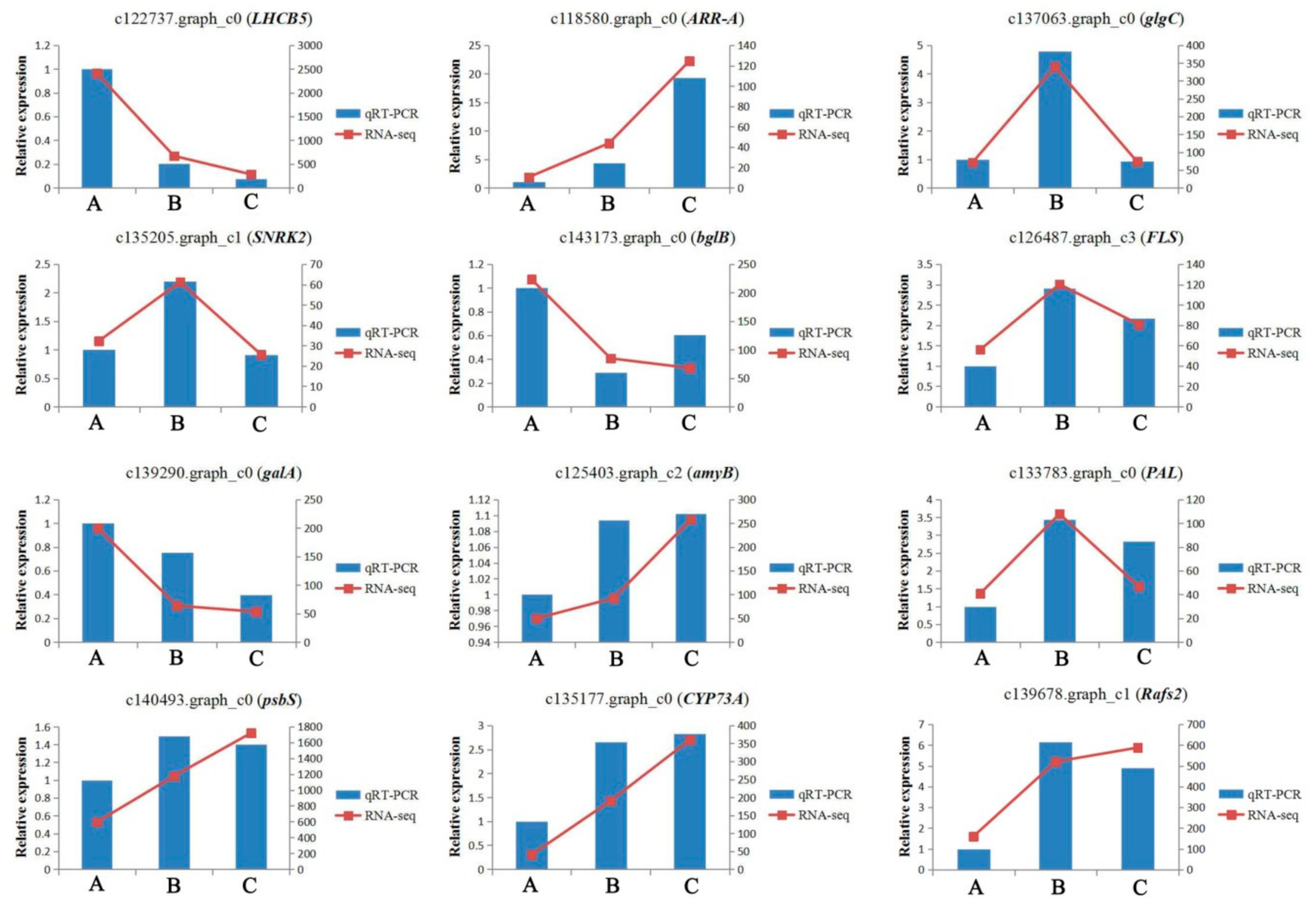
2.11. Quantitative RT-PCR Analysis
3. Discussion
3.1. Transcription Factors and Protein Kinase Responding to Low Temperature
3.2. Hormone-Related DEGs Responding to Low Temperature
3.3. Key DEGs Related to Photosynthesis
3.4. Key DEGs Related to the Membrane System and Osmoregulation
3.5. Other Important DEGs Responding to Low Temperature
4. Materials and Methods
4.1. Plant Materials and Low-Temperature Treatments
4.2. Determination of Physiological Indexes of C. oleifera ‘Huaxin’ under Low Temperatures
4.3. Sampling for RNA-Seq and RNA Preparation
4.4. Library Preparation and RNA Sequencing
4.5. Transcriptome Assembly and Gene Functional Annotation
4.6. Differential Expression Analyses
4.7. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tan, X.F.; Jiang, G.X.; Tan, F.Y. Research report on industrialization development strategy of Vernicia fordii in China. Non Wood Forest Res. 2011, 29, 1–5. [Google Scholar]
- Li, Z.; Tan, X.F.; Liu, Z.M.; Lin, Q.; Zhang, L.; Yuan, J.; Zeng, Y.L.; Wu, L.L. In vitro propagation of Camellia oleifera abel. using hypocotyl, cotyledonary node, and radicle explants. Hort Sci. 2016, 51, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.J.; Wang, H.; Chen, M.; Liao, J.; Yuan, J.; Niu, G.H. Drought stress–induced physiological and metabolic changes in leaves of two oil tea cultivars. J. Amer. Soc. Hort. 2019, 144, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Yuan, D.Y.; Yang, Y.; Wang, B.F.; Liu, D.M.; Zou, F. Pollen tube growth and double fertilization in Camellia oleifera. J. Amer. Soc. Hort. 2015, 140, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Yang, X.Q.; Huang, X.M.; Duan, S.H.; Long, C.; Chen, J.K.; Rong, J. Leaf transcriptome analysis of a subtropical evergreen broadleaf plant, wild oil-tea camellia (Camellia oleifera), revealing candidate genes for cold acclimation. BMC Genom. 2017, 18, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.Y.; Rong, J.; Zhang, W.J.; Chen, J.K. Cultivation history of Camellia oleifera and genetic resources in the Yangtze River Basin. Biodivers. Sci. 2018, 26, 384–395. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Liao, Y.F. Research summary on meteorological influence indicators of oil tea Camellia. Chin. Agric. Sci. Bull. 2015, 31, 179–183. [Google Scholar]
- Andreas, T.; Christophe, C.; Essaïd, A.B. Physiological and molecular changes in plants grown at low temperatures. Planta 2012, 235, 1091–1105. [Google Scholar]
- Wu, Z.J.; Li, X.H.; Liu, Z.W.; Li, H.; Wang, Y.X.; Zhuang, J. Transcriptome-based discovery of AP2/ERF transcription factors related to temperature stress in tea plant (Camellia sinensis). Funct. Integr. Genomic. 2015, 15, 741–752. [Google Scholar] [CrossRef]
- Zeng, Y.R.; Ni, Z.J. Effects of the climate during flowering period on post-flowering fruit setting in Camellia oleifera. J. Zhejiang For. Coll. 2010, 27, 323–328. [Google Scholar]
- Gu, Y.Y.; Zhang, F.H.; Zeng, Y.L.; Zhang, L.; Tan, X.F.; Cao, H.P.; Li, Z. Physiological responses of tung tree (Vernicia fordii) saplings to different red, white and blue light-emitting diodes. Intl. J. Agric. Biol. 2019, 22, 569–577. [Google Scholar]
- Knight, M.R.; Knight, H. Low temperature perception leading to gene expression and cold tolerance in higher plants. New Phytol. 2012, 195, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Furumoto, T. Cold signaling and cold response in plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, K.; Uemura, M.; Tsurumi, S.; Rahman, A. Auxin response in Arabidopsis under cold stress: underlying molecular mechanisms. Plant Cell 2009, 21, 3823–3838. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 0–158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.X.; Liu, L.F.; Wang, P.; Zhu, H.J. The features of the climate adaptability about Camellia oleifera planting in hilly red soil region of southern China: a case study of Changning City Hunan Province. Ecol. Sci. 2015, 34, 61–67. [Google Scholar]
- Tan, X.F.; Yuan, D.Y.; Zou, F.; Yuan, J.; Xie, P.; Su, Y.; Wang, Y.; Yang, D.T.; Peng, J.T. An Elite Variety: Camellia oleifera ‘Huaxin’. Sci. Silvae Sin. 2012, 48, 170–174. [Google Scholar]
- Peng, S.F.; Chen, Y.Z. Freezing injury to oil-tea Camellia and measures to its prevention and remedy. Sci. Silv. Sin. 2008, 24, 184–188. [Google Scholar]
- Wu, L.L.; Li, J.A.; Gu, Y.Y.; Zhang, F.H.; Gu, L.; Tan, X.F.; Shi, M.W. Effect of chilling temperature on florescence, leaf anatomical structure, and physiological and biochemical characteristics of two Camellia oleifera cultivars. Intl. J. Agric. Biol. accepted.
- Wang, X.C.; Zhao, Q.Y.; Ma, C.L.; Zhang, Z.H.; Yang, Y.J. Global transcriptome profiles of Camellia sinensis during cold acclimation. BMC Genom. 2013, 14, 415–429. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.J.; Li, X.H.; Liu, Z.W.; Xu, Z.S.; Zhuang, J. De novo assembly and transcriptome characterization: novel insights into catechins biosynthesis in Camellia sinensis. Bmc Plant Biol. 2014, 14, 227–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Wang, X.W.; Ban, Q.Y.; Zhu, X.X.; Jiang, C.J.; Wei, C.L.; Jeffrey, L.B. Comparative transcriptomic analysis reveals gene expression associated with cold adaptation in the tea plant Camellia sinensis. BMC Genom. 2019, 20, 624–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, E.H.; Jiang, J.J.; Huang, H.; Zhang, L.P.; Zhang, H.B.; Gu, L.Z. Transcriptome Analysis of the Oil-Rich Tea Plant, Camellia oleifera, Reveals Candidate Genes Related to Lipid Metabolism. PloS ONE 2014, 9, e104150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Wang, M.J.; Zhao, S.T.; Hu, J.J.; Lu, M.Z. Changes in freezing tolerance in hybrid poplar caused by up- and down-regulation of PtFAD2 gene expression. Transgenic Res. 2010, 19, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Takahashi, D.; Kawamura, Y.; Uemura, M. Comparison of plasma membrane proteomic changes of Arabidopsis suspension-cultured cells (T87 line) after cold and ABA treatment in association with freezing tolerance development. Plant Cell Physiol. 2012, 53, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Aiello, A.S. Seeking Cold-Hardy Camellias. Arnoldia 2009, 67, 20–30. [Google Scholar]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [CrossRef]
- Buskirk, H.A.; Thomashow, M.F. Arabidopsis transcription factors regulating cold acclimation. Physiol. Plant. 2006, 126, 72–80. [Google Scholar] [CrossRef]
- Cominelli, E.; Gusmaroli, G.; Conti, L.; Allegra, D.; Petroni, K.; Tonelli, C. Role of Arabidopsis MYB transcription factors in osmotic stress. Plant Toler. Abiotic Stresses Agric. Role Genet. Eng. 2000. [Google Scholar] [CrossRef]
- Ma, J.; Wang, F.; Li, M.Y.; Jiang, Q.; Tan, G.F.; Xiong, A.S. Genome wide analysis of the NAC transcription factor family in Chinese cabbage to elucidate responses to temperature stress. Sci. Hortic. 2014, 165, 82–90. [Google Scholar] [CrossRef]
- Cheong, Y.H.; Kim, K.N.; Pandey, G.K.; Gupta, R.; Grant, J.J.; Luan, S. CBL1, a calcium sensor that differentially regulates salt, drought and cold responses in Arabidopsis. Plant Cell 2003, 15, 1833–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denu, J.M.; Stuckey, J.A.; Saper, M.A.; Dixon, J.E. Form and function in protein dephosphorylation. Cell 1996, 87, 361–364. [Google Scholar] [CrossRef] [Green Version]
- Lehti-Shiu, M.D.; Zou, C.; Hanada, K.; Shiu, S.H. Evolutionary history and stress regulation of plant receptor-like kinase/pelle genes. Plant Physiol. 2009, 150, 12–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Los, D.A.; Zorina, A.A.; Sinetova, M.; Kryazhov, S.; Mironov, K.; Zinchenko, V.V. Stress sensors and signal transducers in Cyanobacteria. Sensors 2010, 10, 2386–2415. [Google Scholar] [CrossRef] [Green Version]
- Zorina, A.A. Eukaryotic protein kinases in Cyanobacteria. Russ. J. Plant Physiol. 2013, 60, 589–596. [Google Scholar] [CrossRef]
- Zorina, A.A.; Bedbenov, V.S.; Novikova, G.V.; Panichkin, V.B.; Los, D.A. Involvement of serine/threonine protein kinases in the cold stress response in the Cyanobacteriumsynechocystissp. pcc 6803: functional characterization of SpkE protein kinase. Mol. Biol. 2014, 48, 390–398. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Yadav, S.K. Cold stress tolerance mechanisms in plants. In Sustainable Agriculture Volume 2; Springer: Dordrecht, The Netherlands, 2011. [Google Scholar]
- Muller, B.; Sheen, J. Advances in cytokinin signaling. Science 2007, 318, 68–69. [Google Scholar] [CrossRef]
- Ha, S.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Cytokinins: metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci. 2012, 17, 172–179. [Google Scholar] [CrossRef]
- Danquah, A.; De Zelicourt, J.; Colcombet,, A.; Hirt, H. The role of ABA and MAPK signaling pathways in plant abiotic stress responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef]
- Sijmons, P.C.; Grundler, F.M.W.; Mende, N.; Burrows, P.R.; Wyss, U. Arabidopsis thaliana as a new model for plant-parasitic nematodes. Plant J. 1991, 1, 245–254. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caffarri, S.; Frigerio, S.; Olivieri, E.; Righetti, P.G.; Bassi, R. Differential accumulation of lhcb gene products in thylakoid membranes of Zea mays plants grown under contrasting light and temperature conditions. Proteomics 2005, 5. [Google Scholar]
- Barth, C.; Krause, G.H. Inhibition of photosystem I and II in chilling-sensitive and chilling-tolerant plants under light and low temperature stress. Naturforsch 1999, 54, 645–657. [Google Scholar] [CrossRef]
- Gabriella, S.; Majláth, I.; Pál, M.; Gondor, O.K.; Rudnóy, S.; Oláh, C. Janus-faced nature of light in the cold acclimation processes of Maize. Front. Plant Sci. 2018, 9, 850. [Google Scholar]
- Li, X.G.; Meng, Q.W.; Jiang, G.Q.; Zou, Q. The susceptibility of cucumber and sweet pepper to chilling under low irradiance is related to energy dissipation and water-water cycle. Photosynthetica 2003, 41, 259–265. [Google Scholar] [CrossRef]
- Nield, J.; Barber, J. Refinement of the structural model for the Photosystem II super complex of higher plants. Biochim. Biophys. Acta (BBA)-Bioenerg. 2006, 1757, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.Z.; Zeng, X.C.; Sun, W.C.; Wu, J.Y.; Fang, Y.; Liu, Z.G.; Ma, L. Cloning of PsbR gene in winter turnip rape (Brassica campestris) and its expression analysis under low temperature stress. J. Agricultural Biotechnol. 2017, 25, 1423–1433. [Google Scholar]
- Cui, C.; Wang, L.J.; Zhou, Q.Y.; Tan, Z.F.; Qu, C.M.; Zhang, Z.S. Expression profiling of genes related to photosynthesis and antioxidant capacity in flue-cured tobacco seedlings subjected to chilling stress. Acta Ecol. Sin. 2014, 34, 6076–6089. [Google Scholar]
- Rorat, T.; Havaux, M.; Irzykowski, W.; Stéphan, C.; Noëlle, B.; Rey, P. PSII-S gene expression, photosynthetic activity and abundance of plastid thioredoxin-related and lipid-associated proteins during chilling stress in solanum species differing in freezing resistance. Physiol. Plant. 2001, 113, 72–78. [Google Scholar] [CrossRef]
- Wang, K.; Bai, Z.Y.; Liang, Q.Y.; Liu, Q.L.; Zhang, L.; Pan, Y.Z.; Liu, G.L.; Jiang, B.B.; Zhang, F.; Jia, Y. Transcriptome analysis of Chrysanthemum (Dendranthemagrandiflorum) in response to low temperature stress. BMC Genom. 2018, 19, 319–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hincha, D.K. Cryoprotectin: a plant lipid-transfer protein homologue that stabilizes membranes during freezing. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 909–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, F.; Guy, C.L. β-amylase induction and the protective role of maltose during temperature shock. Plant Physiol. 2004, 135, 1674–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tordachescu, M.; Imai, R. Trehalose Biosynthesis in Response to Abiotic Stresses. J. Integr. Plant Biol. 2010, 50, 1223–1229. [Google Scholar] [CrossRef]
- Konrádová, H.; Gricová, M.; Lipavská, H. Cold-induced accumulation of raffinose family oligosaccharides in somatic embryos of norway spruce (Piceaabies). In Vitro Cell. Dev. Biol.-Plant. 2003, 39, 425–427. [Google Scholar]
- Sprenger, N.; Keller, F. Allocation of raffinose family oligosaccharides to transport and storage pools in Ajugareptans: the roles of two distinct galactinol synthases. Plant J. 2010, 21, 249–258. [Google Scholar] [CrossRef]
- Arti, R.; Ruchi, S.; Arvind, S.P.; Deo, T.R.; Kumar, T.P.; Debasis, C. Expression of rice cyp450-like gene (os08g01480) in Arabidopsis modulates regulatory network leading to heavy metal and other abiotic stress tolerance. PloS ONE 2015, 10, e0138574. [Google Scholar]
- Tamiru, M.; Undan, J.R.; Takagi, H.; Abe, A.; Yoshida, K.; Undan, J.Q.; Natsume, S.; Uemura, A.; Saitoh, H.; Matsumura, H.; et al. A cytochrome P450 (OsDSS1) is involved in growth and drought stress responses in rice (Oryzasatival.). Plant Mol. Biol. 2015, 88, 85–99. [Google Scholar] [CrossRef]
- Wang, C.; Yang, Y.; Wang, H.; Ran, X.; Li, B.; Zhang, J.; Zhang, H. Ectopic expression of a cytochrome P450 monooxygenase gene PtCYP714A3 from Populustrichocarpa reduces shoot growth and improves tolerance to salt stress in transgenic rice. Plant Biotechnol. J. 2016, 14, 1838–1851. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Kumar, A.; Malik, A.K. Flavonoids biosynthesis in plants and its further analysis by capillary electrophoresis. Electrophoresis 2017, 38, 820–832. [Google Scholar] [CrossRef]
- Taulavuori, E.; Tahkokorpi, M.; Taulavuori, K.; Laine, K. Anthocyanins and glutathione S-transferase activities in response to low temperature and frost hardening in Vacciniummyrtillus (L.). J. Plant Physiol. 2004, 161, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Gondor, O.K.; Szalai, G.; Kovács, V.; Janda, T.; Pál, M. Relationship between polyamines and other cold-induced response mechanisms in different cereal species. J. Agro. Crop. Sci. 2016, 202, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Leyva, A. Low temperature induces the accumulation of phenylalanine ammonia-lyase and chalcone synthase mRNAs of Arabidopsis thaliana in a light-dependent manner. Plant Physiol. 1995, 108. [Google Scholar] [CrossRef]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.R.; Hincha, D.K. Natural variation in flavonol and anthocyanin metabolism during cold acclimation in Arabidopsis thaliana accessions. Plant Cell Environ. 2015, 38, 1658–1672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Li, C.; Jiang, Q.T.; Wei, Y.; Yao, H.; Chen, H.; Wu, Q. Cloning and characterization of a cold inducible Pal promoter from Fagopyrumtataricum. Cent. Eur. J. Biol. 2014, 9, 290–297. [Google Scholar]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by over accumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef]
- Watanabe, M.; Ayugase, J. Effect of low temperature on flavonoids, oxygen radical absorbance capacity values and major components of winter sweet spinach (Spinaciaoleracea, L.). J. Sci. Food. Agric. 2015, 95, 2095–2104. [Google Scholar] [CrossRef]
- Wang, B.M.; Chen, J.; Chen, L.; Wang, X.; Wang, R.; Ma, L.; Peng, S.; Luo, J.; Chen, Y. Combined drought and heat stress in Camellia oleifera cultivars: leaf characteristics, soluble sugar and protein contents, and Rubisco gene expression. Trees 2015, 29, 1483–1492. [Google Scholar] [CrossRef]
- Li, Z.; Tan, X.F.; Lu, K.; Liu, Z.M.; Wu, L.L. The effect of CaCl2on calcium content, photosynthesis, and chlorophyll fluorescence of tung tree seedlings under drought conditions. Photosynthetica 2017, 55, 553–560. [Google Scholar] [CrossRef]
- Li, W.; Xiang, F.; Zhong, M.; Zhou, L.; Liu, H.; Li, S.; Wang, X. Transcriptome andmetabolite analysis identifies nitrogen utilization genes in tea plant (Camellia sinensis). Sci. Rep. 2017, 7, 1693. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Dewey, C.N.; Bo, L. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Length Range | Transcript | Unigene |
|---|---|---|
| 200–300 | 42,879 (22.43%) | 35,944 (35.69%) |
| 300–500 | 35,226 (18.43%) | 21,988 (21.83%) |
| 500–1000 | 43,723 (22.87%) | 18,930 (18.80%) |
| 1000–2000 | 42,472 (22.22%) | 14,590 (14.49%) |
| 2000+ | 26,850 (14.05%) | 9251 (9.19%) |
| Total number | 191,150 | 100,703 |
| Total length | 197,720,185 | 79,598,970 |
| N50 length | 1690 | 1413 |
| Mean length | 1034.37 | 790.43 |
| Annotated Databases | Annotated Number | Length <= 300 bp | Length >= 1000 bp |
|---|---|---|---|
| COG | 13,161 | 3273 | 7010 |
| GO | 28,213 | 9019 | 12,485 |
| KEGG | 14,824 | 4606 | 7355 |
| KOG | 23,416 | 7249 | 11,409 |
| Pfam | 26,264 | 7364 | 14,330 |
| Swiss-Prot | 24,203 | 7277 | 13,150 |
| eggNOG | 40,098 | 13,082 | 17,864 |
| NR | 43,518 | 14,758 | 18,596 |
| All | 44,610 | 14,946 | 18,668 |
| DEF-Sets | Pathway ID | Pathway | DEGs in Pathway | All Genesin Pathway | p-Value |
|---|---|---|---|---|---|
| A_B | ko00196 | Photosynthesis—antenna proteins | 10 | 30 | 9.95 × 10−7 |
| ko00941 | Flavonoid biosynthesis | 11 | 48 | 1.68 × 10−5 | |
| ko00130 | Ubiquinone and other terpenoid–quinone biosynthesis | 11 | 59 | 0.000127 | |
| ko00195 | Photosynthesis | 13 | 86 | 0.00029 | |
| ko00360 | Phenylalanine metabolism | 12 | 76 | 0.000327 | |
| ko00940 | Phenylpropanoid biosynthesis | 20 | 184 | 0.000769 | |
| ko04075 | Plant hormone signal transduction | 26 | 271 | 0.000916 | |
| ko00052 | Galactose metabolism | 13 | 107 | 0.002352 | |
| ko00945 | Stilbenoid, diarylheptanoid, and gingerol biosynthesis | 5 | 20 | 0.002448 | |
| ko00350 | Tyrosine metabolism | 10 | 72 | 0.002749 | |
| A_C | ko00196 | Photosynthesis—antenna proteins | 15 | 30 | 6.93 × 10−10 |
| ko00500 | Starch and sucrose metabolism | 44 | 281 | 2.45 × 10−6 | |
| ko04075 | Plant hormone signal transduction | 40 | 271 | 2.95 × 10−5 | |
| ko04070 | Phosphatidylinositol signaling system | 16 | 103 | 0.004376 | |
| ko00940 | Phenylpropanoid biosynthesis | 24 | 184 | 0.005951 | |
| ko00350 | Tyrosine metabolism | 12 | 72 | 0.007305 | |
| ko00460 | Cyanoamino acid metabolism | 12 | 72 | 0.007305 | |
| ko04712 | Circadian rhythm—plant | 12 | 74 | 0.00909 | |
| ko00360 | Phenylalanine metabolism | 12 | 76 | 0.011198 | |
| ko00960 | Tropane, piperidine, and pyridine alkaloid biosynthesis | 8 | 42 | 0.012192 | |
| B_C | ko03010 | Ribosome | 35 | 635 | 5.29 × 10−5 |
| ko00500 | Starch and sucrose metabolism | 19 | 281 | 0.000262 | |
| ko00941 | Flavonoid biosynthesis | 7 | 48 | 0.000307 | |
| ko00052 | Galactose metabolism | 9 | 107 | 0.002673 | |
| ko00196 | Photosynthesis—antenna proteins | 4 | 30 | 0.008731 | |
| ko00940 | Phenylpropanoid biosynthesis | 11 | 184 | 0.012611 | |
| ko04626 | Plant–pathogen interaction | 14 | 262 | 0.01331 | |
| ko00945 | Stilbenoid, diarylheptanoid, and gingerol biosynthesis | 3 | 20 | 0.016597 | |
| ko00520 | Amino sugar and nucleotide sugar metabolism | 11 | 213 | 0.033104 | |
| ko00905 | Brassinosteroid biosynthesis | 2 | 14 | 0.055219 |
| Pathway | Pathway ID | A_B | A_C | B_C |
|---|---|---|---|---|
| Photosynthesis—antenna proteins | ko 00196 | 10 | 15 | 4 |
| Photosynthesis | ko 00195 | 13 | 11 | 2 |
| Phenylalanine metabolism | ko 00360 | 12 | 12 | 5 |
| Plant hormone signal transduction | ko 04075 | 26 | 40 | 11 |
| Galactose metabolism | ko 00052 | 13 | 13 | 9 |
| Phenylpropanoid biosynthesis | ko00940 | 20 | 24 | 11 |
| Starch and sucrose metabolism | ko 00500 | 23 | 44 | 19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.; Li, J.; Li, Z.; Zhang, F.; Tan, X. Transcriptomic Analyses of Camellia oleifera ‘Huaxin’ Leaf Reveal Candidate Genes Related to Long-Term Cold Stress. Int. J. Mol. Sci. 2020, 21, 846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030846
Wu L, Li J, Li Z, Zhang F, Tan X. Transcriptomic Analyses of Camellia oleifera ‘Huaxin’ Leaf Reveal Candidate Genes Related to Long-Term Cold Stress. International Journal of Molecular Sciences. 2020; 21(3):846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030846
Chicago/Turabian StyleWu, Lingli, Jian’an Li, Ze Li, Fanhang Zhang, and Xiaofeng Tan. 2020. "Transcriptomic Analyses of Camellia oleifera ‘Huaxin’ Leaf Reveal Candidate Genes Related to Long-Term Cold Stress" International Journal of Molecular Sciences 21, no. 3: 846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030846