Highly Efficient Protoplast Isolation and Transient Expression System for Functional Characterization of Flowering Related Genes in Cymbidium Orchids

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
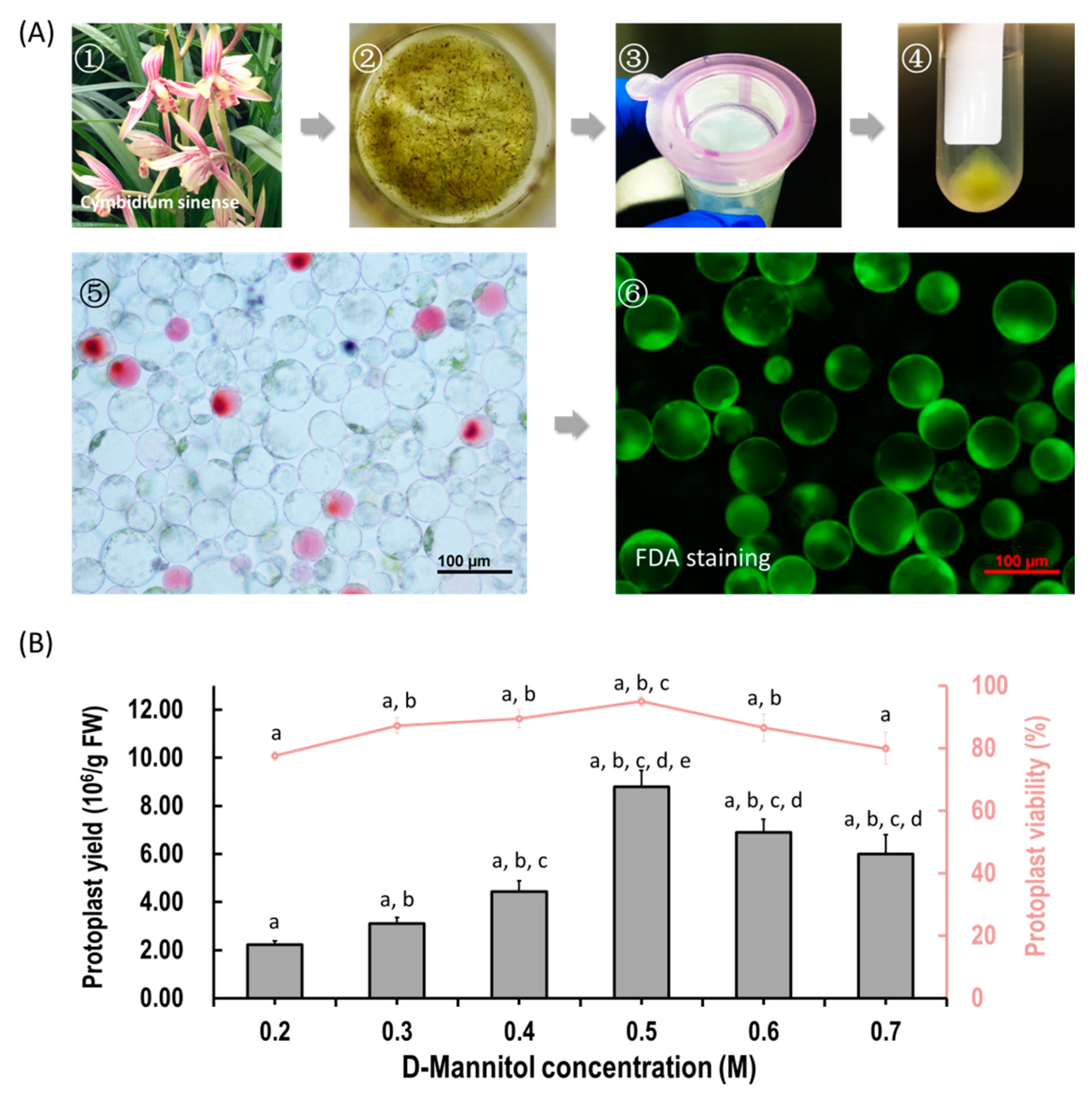
2.1. Protoplast Isolation from Cymbidium Flower Petals
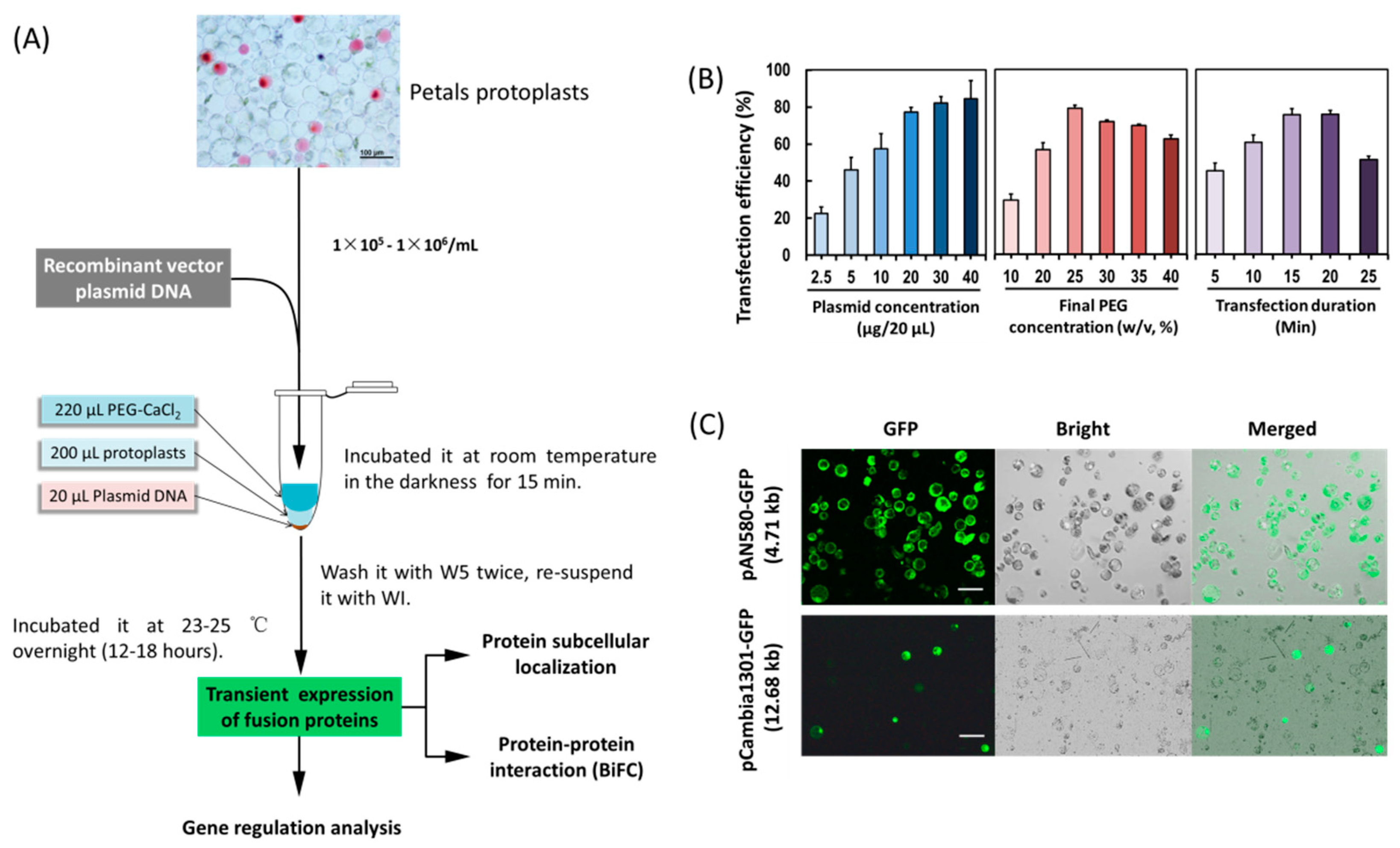
2.2. PEG-Mediated Transient Expression in Orchid Protoplasts
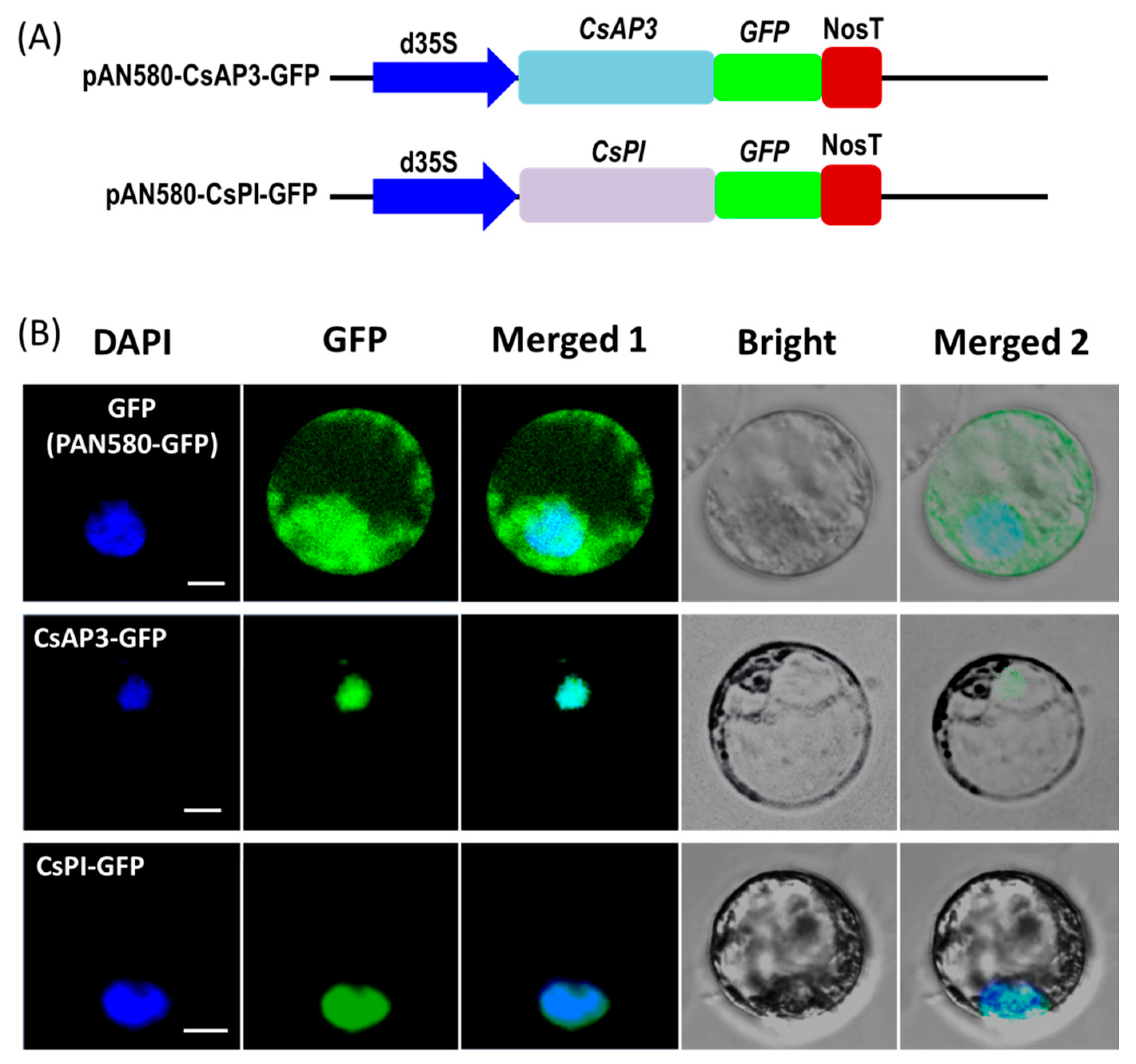
2.3. Protein Subcellular Localization
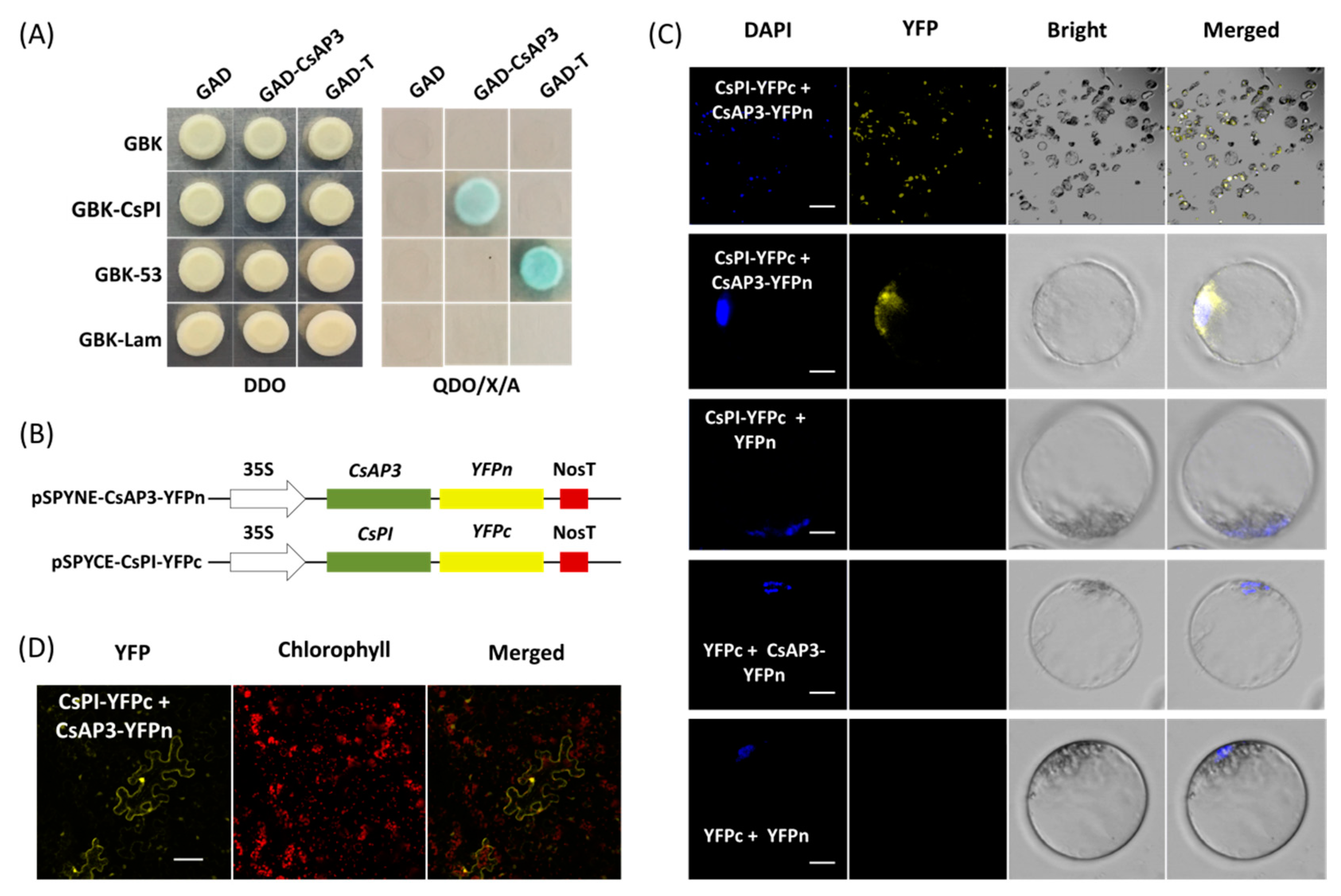
2.4. Protein–Protein Interaction Studies in Cymbidium Protoplasts
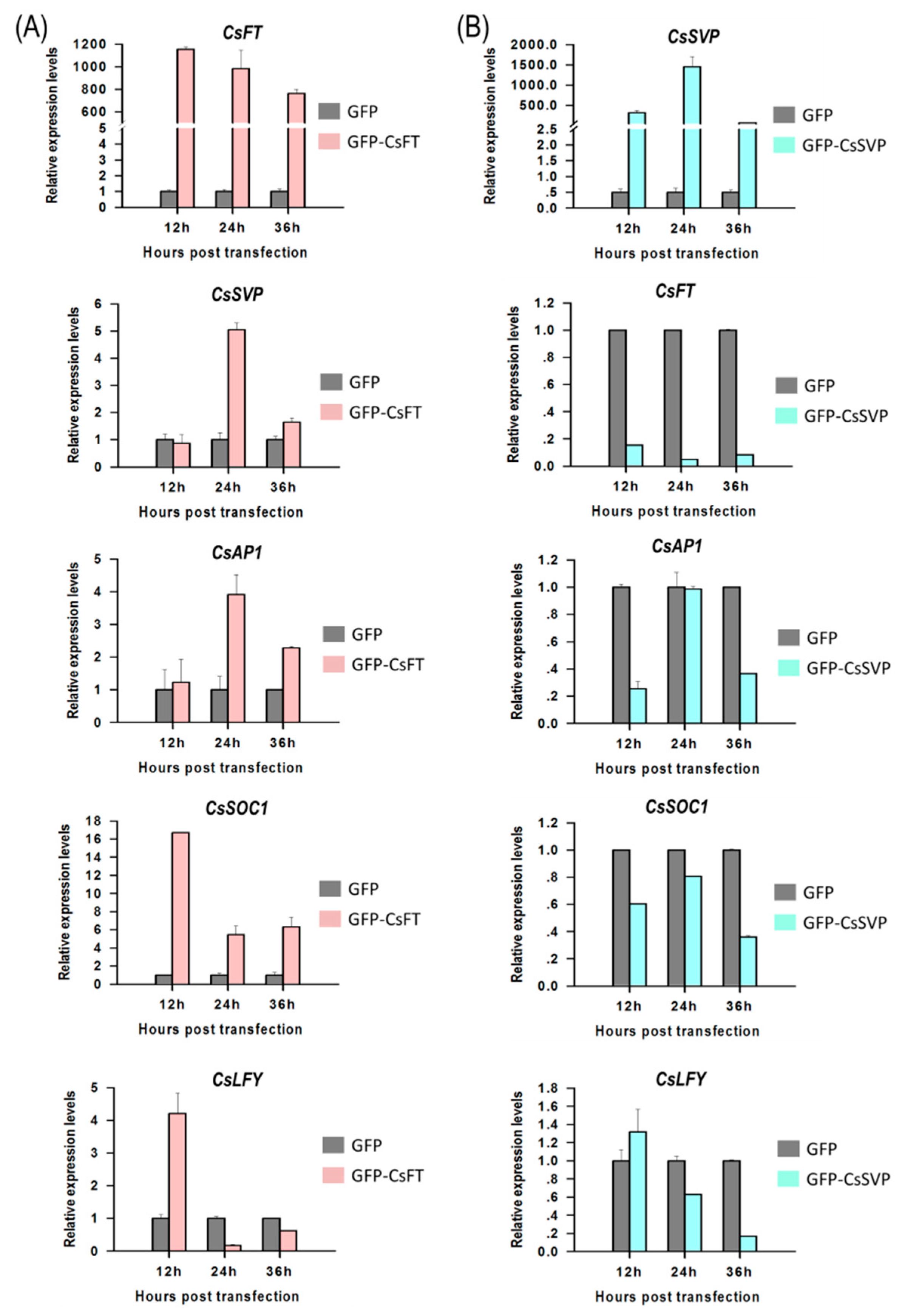
2.5. Gene Regulation Analysis in Cymbidium Protoplasts Using qRT-PCR
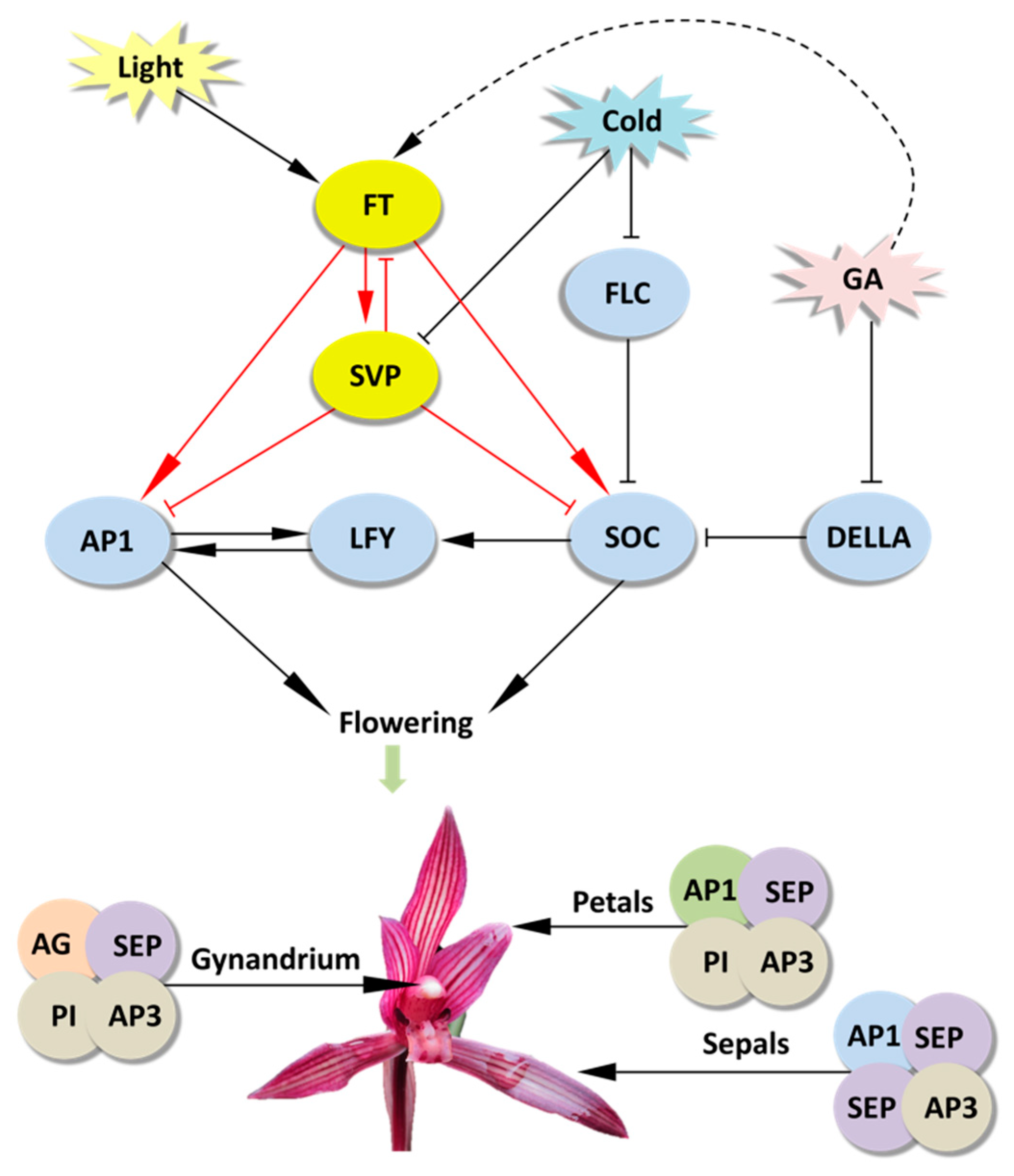
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Protoplast Isolation
4.3. PEG-Mediated Protoplast Transfection
4.4. Protein Subcellular Localization
4.5. Yeast Two-Hybrid Assay
4.6. BiFC Assay in Orchid Protoplasts and N. benthamiana Leaves
4.7. Gene Regulation Analysis by qRT-PCR
4.8. qRT-PCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PEG | Polyethylene glycol |
| PTES | Protoplast-based transient expression system |
| BiFC | Bimolecular fluorescence complementation |
| FW | Fresh weight |
| GFP | Green fluorescent protein |
| CDS | Coding sequences |
| DAPI | 4’-6’-diamidino-2-phenylindole |
| hpt | Hours post transfection |
| FT | FLOWERING LOCUS T |
| SVP | SHORT VEGETATIVE PHASE |
| CsSOC1 | SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 |
| Y2H | Yeast two-hybrid assay |
| Ca2+ | Calcium ion |
| FDA | Fluorescein diacetate |
| YFP | Yellow fluorescent protein |
References
- Hsiao, Y.Y.; Tsai, W.C.; Kuoh, C.S.; Huang, T.H.; Wang, H.C.; Wu, T.S.; Leu, Y.L.; Chen, W.H.; Chen, H.H. Comparison of transcripts in Phalaenopsis bellina and Phalaenopsis equestris (Orchidaceae) flowers to deduce monoterpene biosynthesis pathway. BMC Plant Biol. 2006, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, D.L.; Dixon, K.W. Orchids. Curr. Biol. 2008, 18, 325–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.M. Orchid viruses–A compendium. In Orchid Biology: Reviews and Perspectives, VIII; Kull, T., Arditti, J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 505–546. [Google Scholar]
- Shrestha, B.R.; Tokuhara, K.; Mii, M. Plant regeneration from cell suspension-derived protoplasts of Phalaenopsis. Plant Cell Rep. 2007, 26, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Yam, T.W.; Arditti, J. History of orchid propagation: A mirror of the history of biotechnology. Plant Biotechnol. Rep. 2009, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Liu, X.; Vanneste, K.; Proost, S.; Tsai, W.C.; Liu, K.W.; Chen, L.J.; He, Y.; Xu, Q.; Bian, C. The genome sequence of the orchid Phalaenopsis equestris. Nat. Genet. 2015, 47, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.Q.; Xu, Q.; Bian, C.; Tsai, W.C.; Yeh, C.M.; Liu, K.W.; Yoshida, K.; Zhang, L.S.; Chang, S.B.; Chen, F. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 19029. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.Q.; Liu, K.W.; Li, Z.J.; Lohaus, R.; Hsiao, Y.Y.; Niu, S.C.; Wang, J.Y.; Lin, Y.C.; Xu, Q.; Chen, L.J. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Brisson, L.F.; Tenhaken, R.; Lamb, C. Function of oxidative cross-linking of cell wall structural proteins in plant disease resistance. Plant Cell 1994, 6, 1703–1712. [Google Scholar] [CrossRef] [Green Version]
- Kolewe, M.E.; Gaurav, V.; Roberts, S.C. Pharmaceutically active natural product synthesis and supply via plant cell culture technology. Mol. Pharm. 2008, 5, 243–256. [Google Scholar] [CrossRef]
- Pereira, S.P.; Fernandes, M.A.; Martins, J.D.; Santos, M.S.; Moreno, A.J.; Vicente, J.A.; Videirae, R.A.; Jurado, A.S. Toxicity assessment of the herbicide metolachlor comparative effects on bacterial and mitochondrial model systems. Toxicol. In Vitro 2009, 23, 1585–1590. [Google Scholar] [CrossRef]
- Petersson, S.V.; Lindén, P.; Moritz, T.; Ljung, K. Cell-type specific metabolic profiling of Arabidopsis thaliana protoplasts as a tool for plant systems biology. Metabolomics 2015, 11, 1679–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denyer, T.; Ma, X.; Klesen, S.; Scacchi, E.; Nieselt, K.; Timmermans, M.C. Spatiotemporal developmental trajectories in the Arabidopsis root revealed using high-throughput single-cell RNA sequencing. Dev. Cell 2019, 48, 840–852. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.R.; Anthony, P.; Power, J.B.; Lowe, K.C. Plant protoplast technology: Current status. Acta Physiol. Plant 2005, 27, 117–130. [Google Scholar] [CrossRef]
- Nagata, T.; Takebe, I. Plating of isolated tobacco mesophyll protoplasts on agar medium. Planta 1971, 99, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Menczel, L. Improved protoplast culture and plant regeneration from protoplast-derived callus in Arabidopsis thaliana. Z. Pflanzenphysiol. 1980, 96, 77–80. [Google Scholar]
- Kanai, R.; Edwards, G.E. Separation of mesophyll protoplasts and bundle sheath cells from maize leaves for photosynthetic studies. Plant Physiol. 1973, 51, 1133–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toriyama, K.; Hinata, K. Cell suspension and protoplast culture in rice. Plant Sci. 1985, 41, 179–183. [Google Scholar] [CrossRef]
- Song, J.; Sorensen, E.L.; Liang, G.H. Direct embryogenesis from single mesophyll protoplasts in alfalfa (Medicago sativa L.). Plant Cell Rep. 1990, 9, 21–25. [Google Scholar] [CrossRef]
- Mazarei, M.; Al-Ahmad, H.; Rudis, M.R.; Stewart, C.N., Jr. Protoplast isolation and transient gene expression in switchgrass, Panicum virgatum L. Biotechnol. J. 2008, 3, 354–359. [Google Scholar] [CrossRef]
- Masani, M.Y.A.; Noll, G.A.; Parveez, G.K.A.; Sambanthamurthi, R.; Prüfer, D. Efficient transformation of oil palm protoplasts by PEG-mediated transfection and DNA microinjection. PLoS ONE 2014, 9, e96831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.D.; Wang, L.J.; He, C.Z.; Luo, H.L. An efficient transient mesophyll protoplast system for investigation of the innate immunity responses in the rubber tree (Hevea brasiliensis). Plant Cell Tissue Organ Cult. 2016, 126, 281–290. [Google Scholar] [CrossRef]
- Nanjareddy, K.; Arthikala, M.K.; Blanco, L.; Arellano, E.S.; Lara, M. Protoplast isolation, transient transformation of leaf mesophyll protoplasts and improved Agrobacterium-mediated leaf disc infiltration of Phaseolus vulgaris: Tools for rapid gene expression analysis. BMC Biotechnol. 2016, 16, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.M.; Meng, D.; McGrouther, K.; Zhang, J.H.; Cheng, L.L. Efficient isolation of Magnolia protoplasts and the application to subcellular localization of MdeHSF1. Plant Methods 2017, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Kameya, T.; Ichihashi, S. Plant regeneration from protoplasts derived from callus of Phalaenopsis. Plant Tissue Cult. Lett. 1993, 10, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Kanchanapoom, K.; Jantaro, S.; Rakchad, D. Isolation and fusion of protoplasts from mesophyll cells of Dendrobium pompadour. Sci. Asia 2001, 27, 29–34. [Google Scholar] [CrossRef]
- Keskitalo, M.; Pehu, E.; Simon, J.E. Variation in volatile compounds from tansy (Tanacetum vulgare L.) related to genetic and morphological differences of genotypes. Biochem. Syst. Ecol. 2001, 29, 267–285. [Google Scholar] [CrossRef]
- Mliki, A.; Jardak, R.; Reustle, G.M.; Ghorbel, A. Isolation and culture of leaf protoplasts from Tunisian grapes. OENO One 2003, 37, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.M.; Ahn, N.Y.; Son, B.H.; Kim, C.Y.; Han, C.D.; Kim, G.D.; Gal, S.W.; Wan, S.; Lee, S.H. Efficient transient expression and transformation of PEG-mediated gene uptake into mesophyll protoplasts of pepper (Capsicum annuum L.). Plant Cell Tissue Organ Cult. 2007, 88, 225–232. [Google Scholar] [CrossRef]
- Priyadarshan, S.V.G.N.; Hu, B.Y.; Li, W.M.; Ali, H.; Jia, H.F.; Zhao, L.H.; Ojolo, S.P.; Azam, S.M.; Xiong, J.J.; Yan, M. Simple protoplast isolation system for gene expression and protein interaction studies in pineapple (Ananas comosus L.). Plant Methods 2018, 14, 95. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Ohnishi, T.; Ishida, H.; Tomida, K.; Sakai, M.; Hara, M.; Watanabe, N. Functional characterization of aromatic amino acid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts. J. Plant Physiol. 2012, 169, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.W.; Wong, S.M.; Loh, C.S.; Goh, C.J. Synergism in replication of cymbidium mosaic potexvirus (CymMV) and odontoglossum ringspot tobamovirus (ORSV) RNA in orchid protoplasts. Arch. Virol. 1998, 143, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Chen, J.C.; Fang, S.C. A protoplast transient expression system to enable molecular, cellular, and functional studies in Phalaenopsis orchids. Front. Plant Sci. 2018, 9, 843. [Google Scholar] [CrossRef] [PubMed]
- Rezazadeh, R.; Niedz, R.P. Protoplast isolation and plant regeneration of guava (Psidium guajava L.) using experiments in mixture-amount design. Plant Cell Tissue Organ Cult. 2015, 122, 585–604. [Google Scholar] [CrossRef]
- Capesius, I.; Meyer, Y. Isolation of nuclei from protoplasts of orchids. Cytobiologie 1977, 15, 485–490. [Google Scholar]
- Teo, C.K.H.; Neumann, K.H. Gewinnung, Kultur and fusion von Orchideenprotoplasten. Orchidee 1978, 29, 90–92. [Google Scholar]
- Teo, C.K.H.; Neumann, K.H. The culture of protoplasts isolated from Renantanda Rosalind Cheok. Orchid Rev. 1978, 86, 156–158. [Google Scholar]
- Khentry, Y.; Paradornuvat, A.; Tantiwiwat, S.; Phansiri, S.; Thaveechai, N. Protoplast isolation and culture of Dendrobium Sonia “Bom 17”. Kasetsart J. 2006, 40, 361–369. [Google Scholar]
- Pindel, A. Optimization of isolation conditions of Cymbidium protoplasts. Folia Hortic. 2007, 19, 79–88. [Google Scholar]
- Li, J.L.; Liao, X.Z.; Zhou, S.S.; Liu, S.; Jiang, L.; Wang, G. Efficient protoplast isolation and transient gene expression system for Phalaenopsis hybrid cultivar ‘Ruili Beauty’. In Vitro Cell. Dev. Biol. Plant 2018, 54, 87–93. [Google Scholar] [CrossRef]
- Wu, F.H.; Shen, S.C.; Lee, L.Y.; Lee, S.H.; Chan, M.T.; Lin, C.S. Tape-Arabidopsis Sandwich-a simpler Arabidopsis protoplast isolation method. Plant Methods 2009, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.M.; Yao, D.M.; Lin, F.; Jiang, M.Y. PEG-mediated transient gene expression and silencing system in maize mesophyll protoplasts: A valuable tool for signal transduction study in maize. Acta Physiol. Plant 2014, 36, 1271–1281. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, J.B.; Duan, S.; Ao, Y.; Dai, J.R.; Liu, J.; Wang, P.; Li, Y.G.; Liu, B.; Wang, H.B. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods 2011, 7, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Z.; Liu, Q.; Geng, X.S.; Li, K.M.; Luo, L.J.; Liu, J.P. Highly efficient mesophyll protoplast isolation and PEG-mediated transient gene expression for rapid and large-scale gene characterization in cassava (Manihot esculenta Crantz). BMC Biotechnol. 2017, 17, 29. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Tao, L.; Zeng, L.; Vega-Sanchez, M.E.; Umemura, K.; Wang, G.L. A highly efficient transient protoplast system for analyzing defence gene expression and protein–protein interactions in rice. Mol. Plant Pathol. 2006, 7, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Müller, B.; Sheen, J. Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis. Nature 2008, 453, 1094–1097. [Google Scholar] [CrossRef] [Green Version]
- Zulkarnain, Z.; Tapingkae, T.; Taji, A. Applications of in vitro techniques in plant breeding. In Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools; Springer: Cham, Switzerland, 2015; pp. 293–328. [Google Scholar]
- Page, M.T.; Parry, M.A.; Carmo-Silva, E. A high-throughput transient expression system for rice. Plant Cell Environ. 2019. [Google Scholar] [CrossRef]
- Bart, R.; Chern, M.; Park, C.J.; Bartley, L.; Ronald, P.C. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods 2006, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Zheng, X.L.; Qi, Y.P.; Zhang, D.W.; Cheng, Y.; Tang, A.T.; Voytas, D.F.; Zhang, Y. A single transcript CRISPR-Cas9 system for efficient genome editing in plants. Mol. Plant 2016, 9, 1088–1091. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Hsu, C.T.; Yang, L.H.; Lee, L.Y.; Fu, J.Y.; Cheng, Q.W.; Wu, F.H.; Hsiao, H.C.W.; Zhang, Y.S.; Zhang, R. Application of protoplast technology to CRISPR/Cas9 mutagenesis: From single-cell mutation detection to mutant plant regeneration. Plant Biotechnol. J. 2018, 16, 1295–1310. [Google Scholar] [CrossRef] [Green Version]
- Krens, F.A.; Molendijk, L.; Wullems, G.J.; Schilperoort, R.A. In vitro transformation of plant protoplasts with Ti-plasmid DNA. Nature 1982, 296, 72. [Google Scholar] [CrossRef]
- Hauptmann, R.M.; Ozias-Akins, P.; Vasil, V.; Tabaeizadeh, Z.; Rogers, S.G.; Horsch, R.B.; Vasil, I.K.; Fraley, R.T. Transient expression of electroporated DNA in monocot and dicotyledonous species. Plant Cell Rep. 1987, 6, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Hayashimoto, A.; Li, Z.; Murai, N. A polyethylene glycol-mediated protoplast transformation system for production of fertile transgenic rice plants. Plant Physiol. 1990, 93, 857–863. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Wang, Z.Y.; Cheng, J.T.; Zhao, W.C.; Li, X.; Wang, H.Y.; Zhang, Z.X.; Sui, X. An efficient cucumber (Cucumis sativus L.) protoplast isolation and transient expression system. Sci. Hortic. Amst. 2013, 150, 206–212. [Google Scholar] [CrossRef]
- Hong, S.Y.; Seo, P.J.; Cho, S.H.; Park, C.M. Preparation of leaf mesophyll protoplasts for transient gene expression in Brachypodium distachyon. J. Plant Biol. 2012, 55, 390–397. [Google Scholar] [CrossRef]
- Burris, K.P.; Dlugosz, E.M.; Collins, A.G.; Stewart, C.N.; Lenaghan, S.C. Development of a rapid, low-cost protoplast transfection system for switchgrass (Panicum virgatum L.). Plant Cell Rep. 2016, 35, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.L.; Li, Y.J.; Hu, Y.; Gao, Y.R.; Zang, X.W.; Ding, Q.; Wang, Y.J.; Wen, Y.Q. A highly efficient grapevine mesophyll protoplast system for transient gene expression and the study of disease resistance proteins. Plant Cell Tissue Organ Cult. 2016, 125, 43–57. [Google Scholar] [CrossRef]
- Sheen, J. Signal transduction in maize and Arabidopsis mesophyll protoplasts. Plant Physiol. 2001, 127, 1466–1475. [Google Scholar] [CrossRef]
- Mathieu, Y.; Lapous, D.; Thomine, S.; Laurie´re, C.; Guern, J. Cytoplasmic acidification as an early phosphorylation-dependent response of tobacco cells to elicitors. Planta 1996, 199, 416–424. [Google Scholar] [CrossRef]
- Raikar, S.V.; Braun, R.H.; Bryant, C.; Conner, A.J.; Christey, M.C. Efficient isolation, culture and regeneration of Lotus corniculatus protoplasts. Plant Biotechnol. Rep. 2008, 2, 171. [Google Scholar] [CrossRef]
- Yao, L.P.; Liao, X.; Gan, Z.Z.; Peng, X.; Wang, P.; Li, S.J.; Li, T.H. Protoplast isolation and development of a transient expression system for sweet cherry (Prunus avium L.). Sci. Hortic. Amst. 2016, 209, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Wehner, N.; Hartmann, L.; Ehlert, A.; Böttner, S.; Oñate-Sánchez, L.; Dröge-Laser, W. High-throughput protoplast transactivation (PTA) system for the analysis of Arabidopsis transcription factor function. Plant J. 2011, 68, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Schultz, E.A.; Pickett, F.B.; Haughn, G.W. The FLO10 gene product regulates the expression domain of homeotic genes AP3 and PI in Arabidopsis flowers. Plant Cell 1991, 3, 1221–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature 2000, 405, 200. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.C.; Ao, X.; Liu, G.X.; Cheng, F.Y.; He, C.Y. Duplication and whorl-specific down-regulation of the obligate AP3–PI heterodimer genes explain the origin of Paeonia lactiflora plants with spontaneous corolla mutation. Plant Cell Physiol. 2017, 58, 411–425. [Google Scholar]
- Huang, W.T.; Fang, Z.M.; Zeng, S.J.; Zhang, J.X.; Wu, K.L.; Chen, Z.L.; Teixeira da Silva, J.A.; Duan, J. Molecular cloning and functional analysis of three FLOWERING LOCUS T (FT) homologous genes from Chinese Cymbidium. Int. J. Mol. Sci. 2012, 13, 11385–11398. [Google Scholar] [CrossRef] [Green Version]
- Mondragón Palomino, M. Perspectives on MADS-box expression during orchid flower evolution and development. Front. Plant Sci. 2013, 4, 377. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.M.; Tong, C.G.; Jang, S. Current progress in orchid flowering/flower development research. Plant Signal. Behav. 2017, 12, e1322245. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.X.; Zhu, G.F.; Wei, Y.L.; Gao, J.; Liang, G.; Peng, L.Y.; Lu, C.Q.; Jin, J.P. Low-temperature-induced changes in the transcriptome reveal a major role of CgSVP genes in regulating flowering of Cymbidium goeringii. BMC Genom. 2019, 20, 53. [Google Scholar] [CrossRef] [Green Version]
- Negrutiu, I.; de Brouwer, D.; Watts, J.W.; Sidorov, V.I.; Dirks, R.; Jacobs, M. Fusion of plant protoplasts: A study using auxotrophic mutants of Nicotiana plumbaginifolia, Viviani. Theor. Appl. Genet. 1986, 72, 279–286. [Google Scholar] [CrossRef]
- Negrutiu, I.; Shillito, R.; Potrykus, I.; Biasini, G.; Sala, F. Hybrid genes in the analysis of transformation conditions. Plant Mol. Biol. 1987, 8, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, R.; Liu, S.C.; Karthikeyan, A.; Wang, T.; Niu, H.P.; Yin, J.L.; Yang, Y.H.; Wang, L.Q.; Yang, Q.H.; Zhi, H.J. Fine-mapping and identification of a novel locus Rsc15 underlying soybean resistance to Soybean mosaic virus. Theor. Appl. Genet. 2017, 130, 2395–2410. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, R.; Gao, J.; Lu, C.; Wei, Y.; Jin, J.; Wong, S.-M.; Zhu, G.; Yang, F. Highly Efficient Protoplast Isolation and Transient Expression System for Functional Characterization of Flowering Related Genes in Cymbidium Orchids. Int. J. Mol. Sci. 2020, 21, 2264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072264
Ren R, Gao J, Lu C, Wei Y, Jin J, Wong S-M, Zhu G, Yang F. Highly Efficient Protoplast Isolation and Transient Expression System for Functional Characterization of Flowering Related Genes in Cymbidium Orchids. International Journal of Molecular Sciences. 2020; 21(7):2264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072264
Chicago/Turabian StyleRen, Rui, Jie Gao, Chuqiao Lu, Yonglu Wei, Jianpeng Jin, Sek-Man Wong, Genfa Zhu, and Fengxi Yang. 2020. "Highly Efficient Protoplast Isolation and Transient Expression System for Functional Characterization of Flowering Related Genes in Cymbidium Orchids" International Journal of Molecular Sciences 21, no. 7: 2264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072264