
Discovery of Phenolic Glycoside from Hyssopus cuspidatus Attenuates LPS-Induced Inflammatory Responses by Inhibition of iNOS and COX-2 Expression through Suppression of NF-κB Activation

 , ,
, , 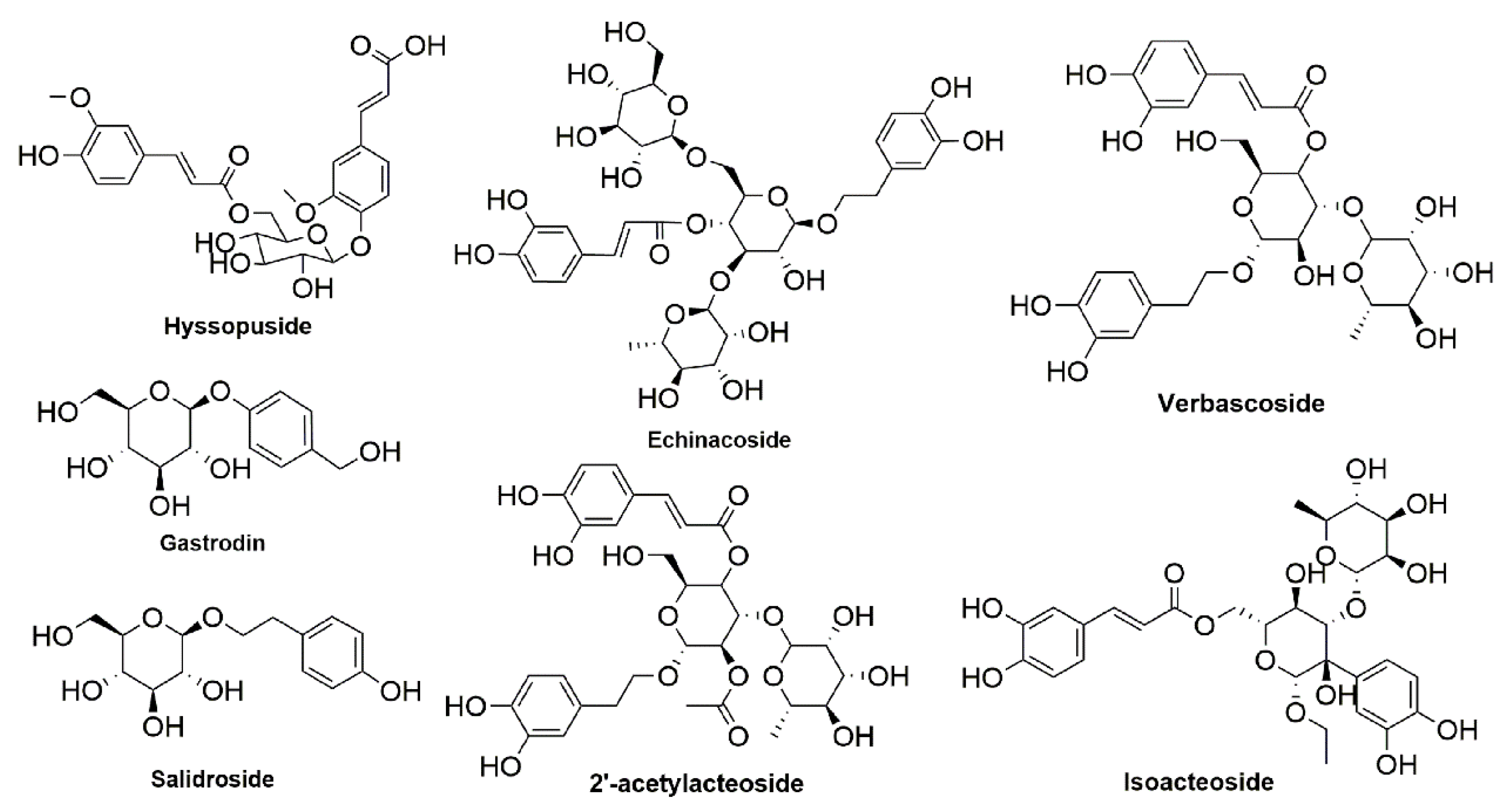{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
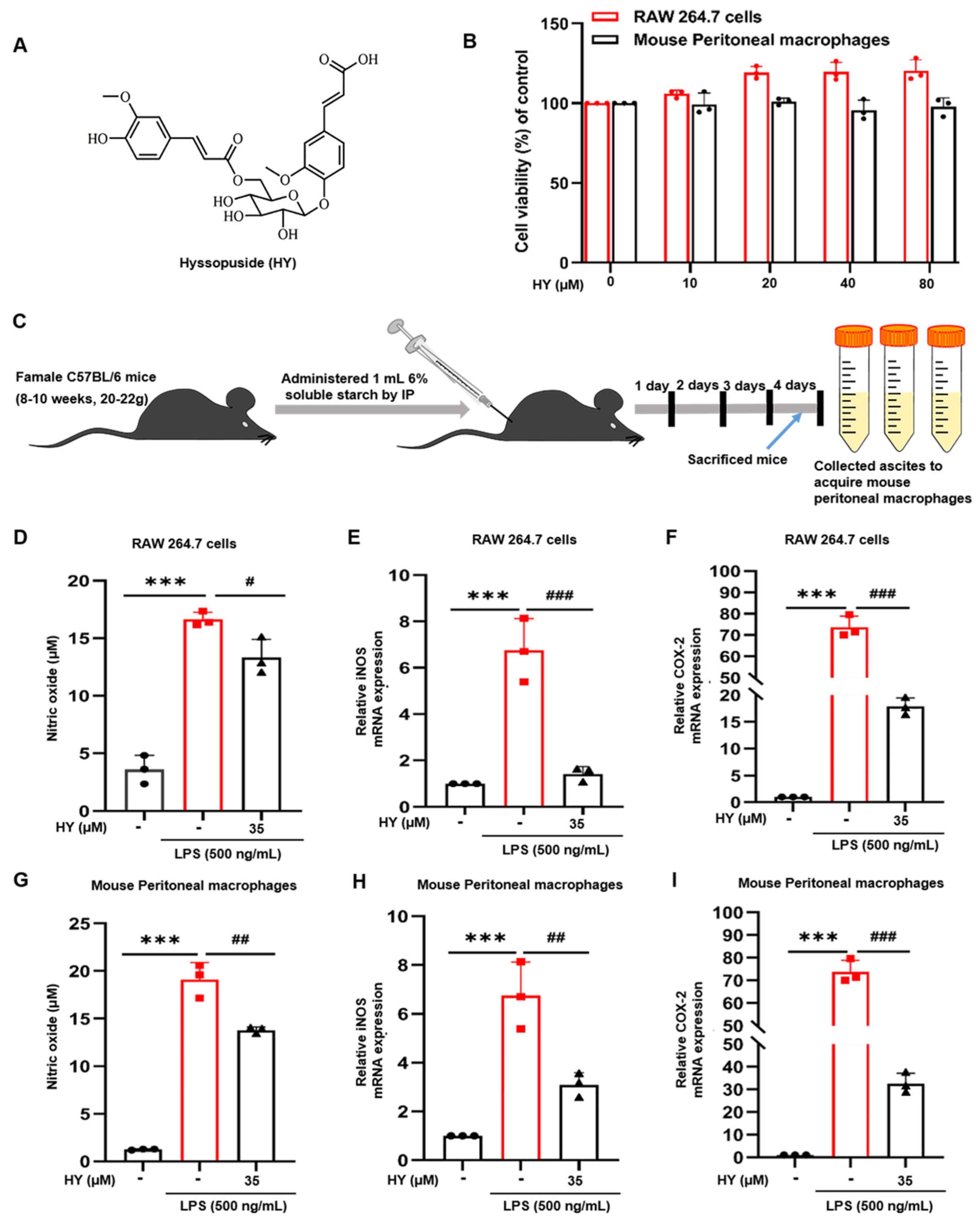
2.1. Effects of HY on the Viability of Macrophages
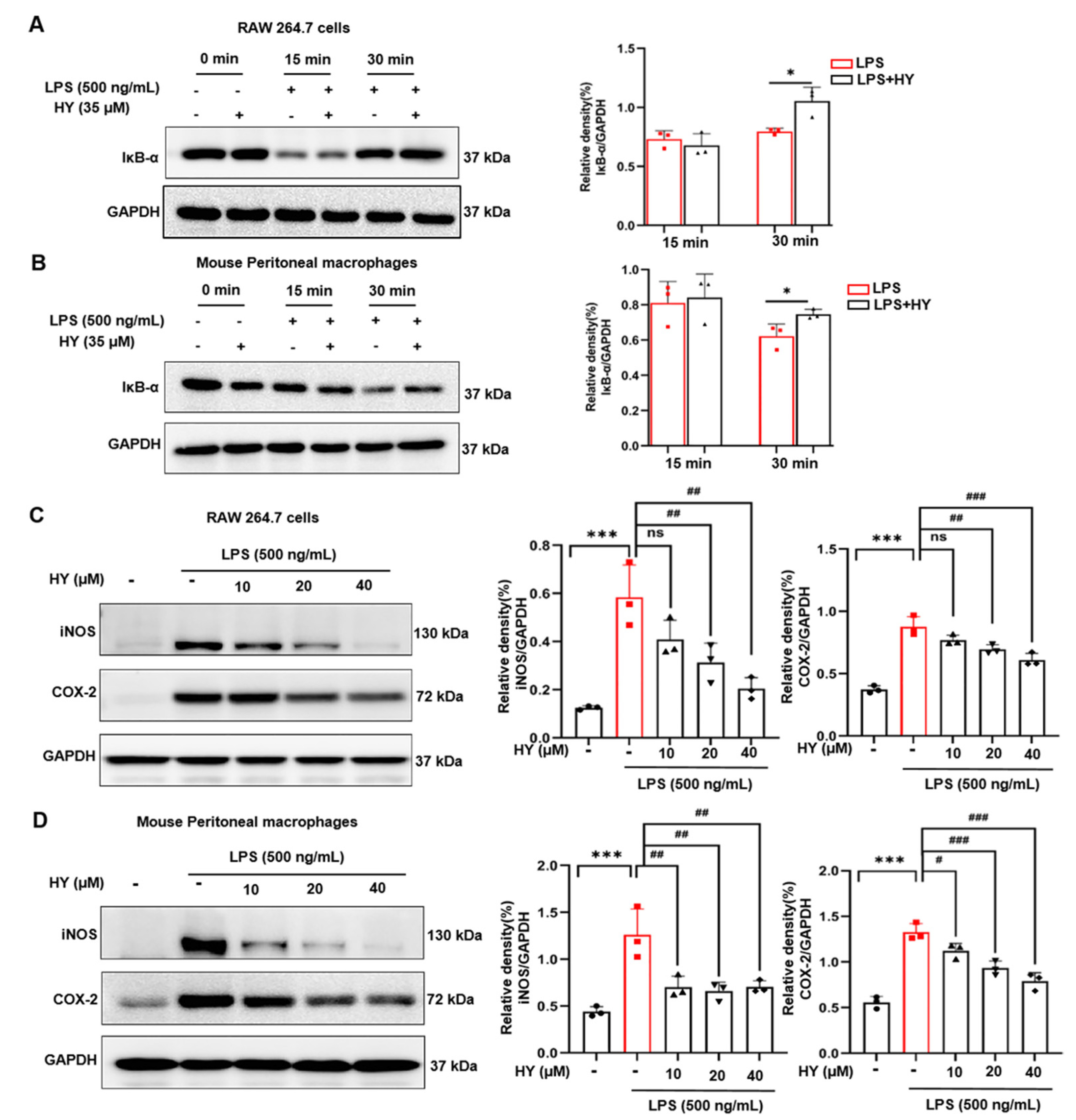
2.2. Effects of HY on Inflammatory Mediator Production in LPS-Induced Macrophages
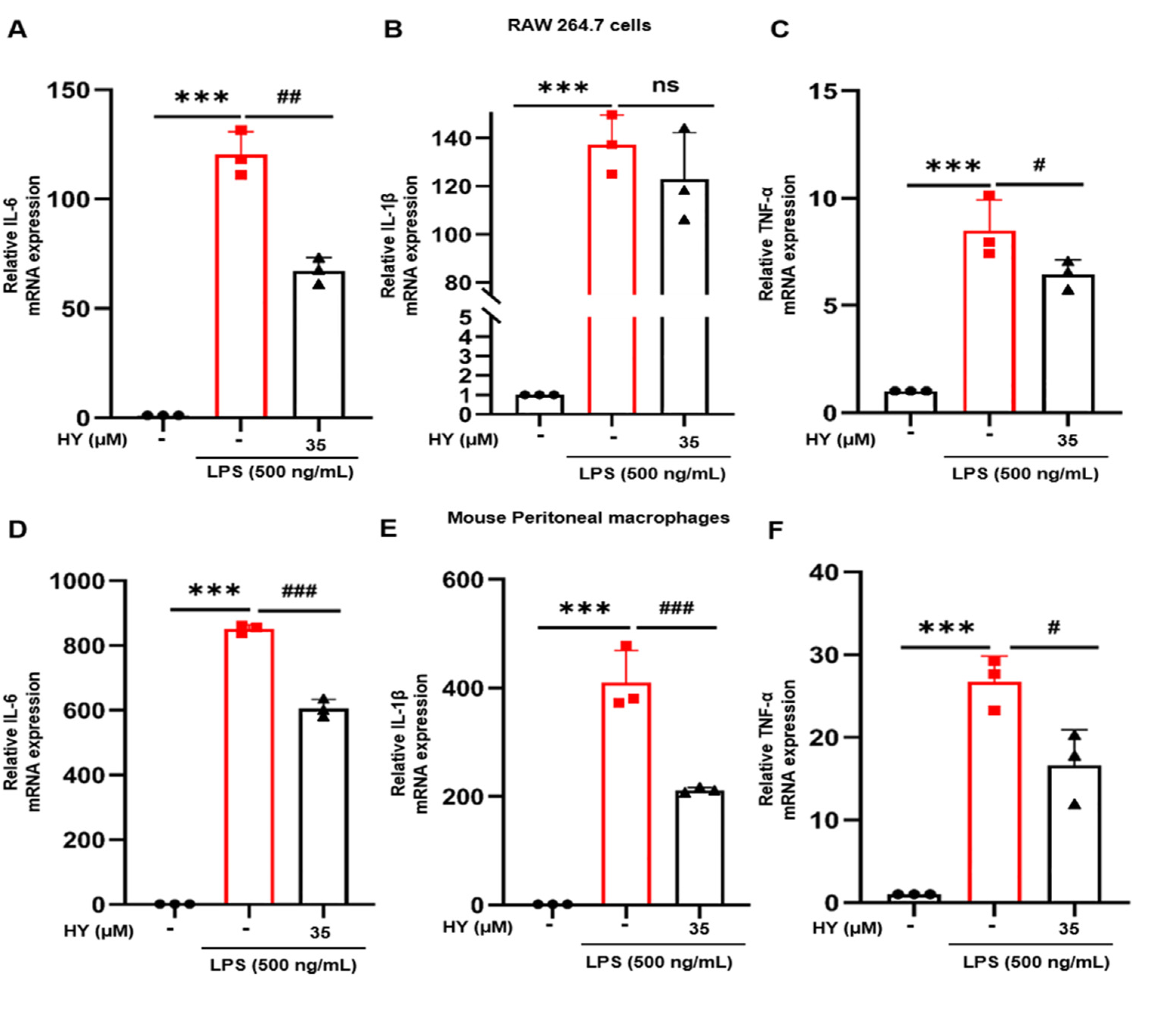
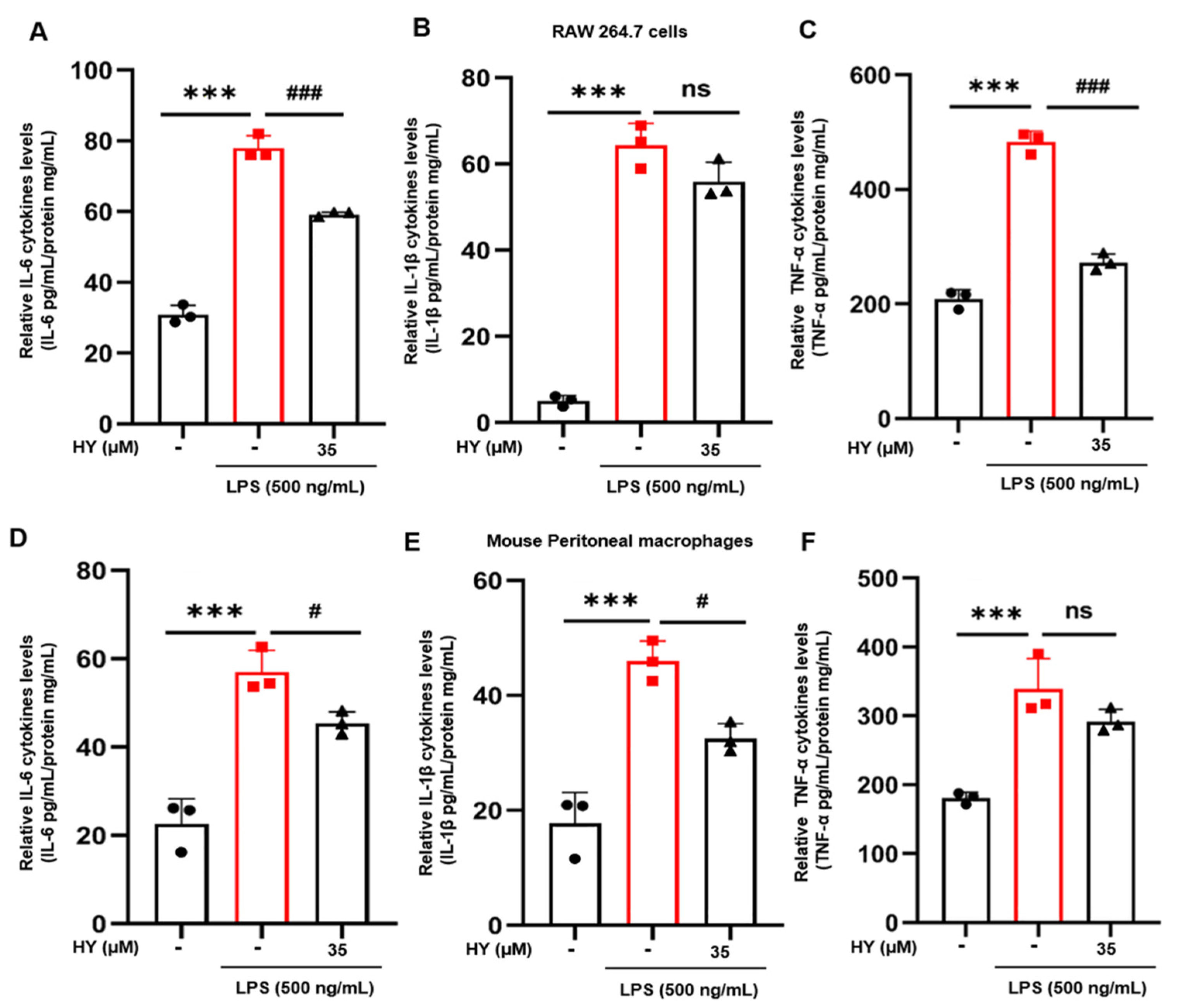
2.3. Effects of HY on Inflammatory Cytokine Expression and Production in LPS-Induced Macrophages
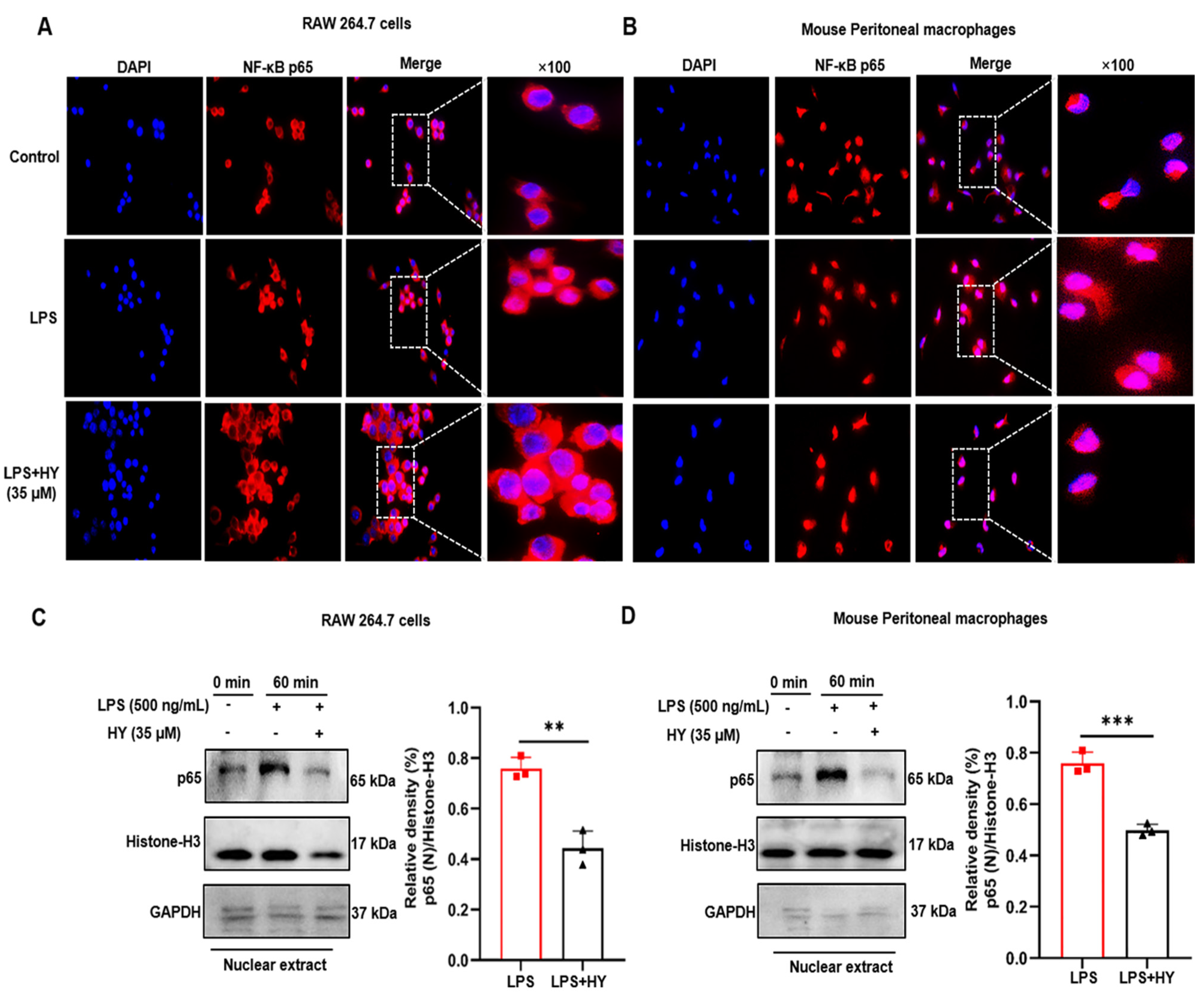
2.4. Effects of HY on Suppressing NF-κB Activation in LPS-Induced Macrophages
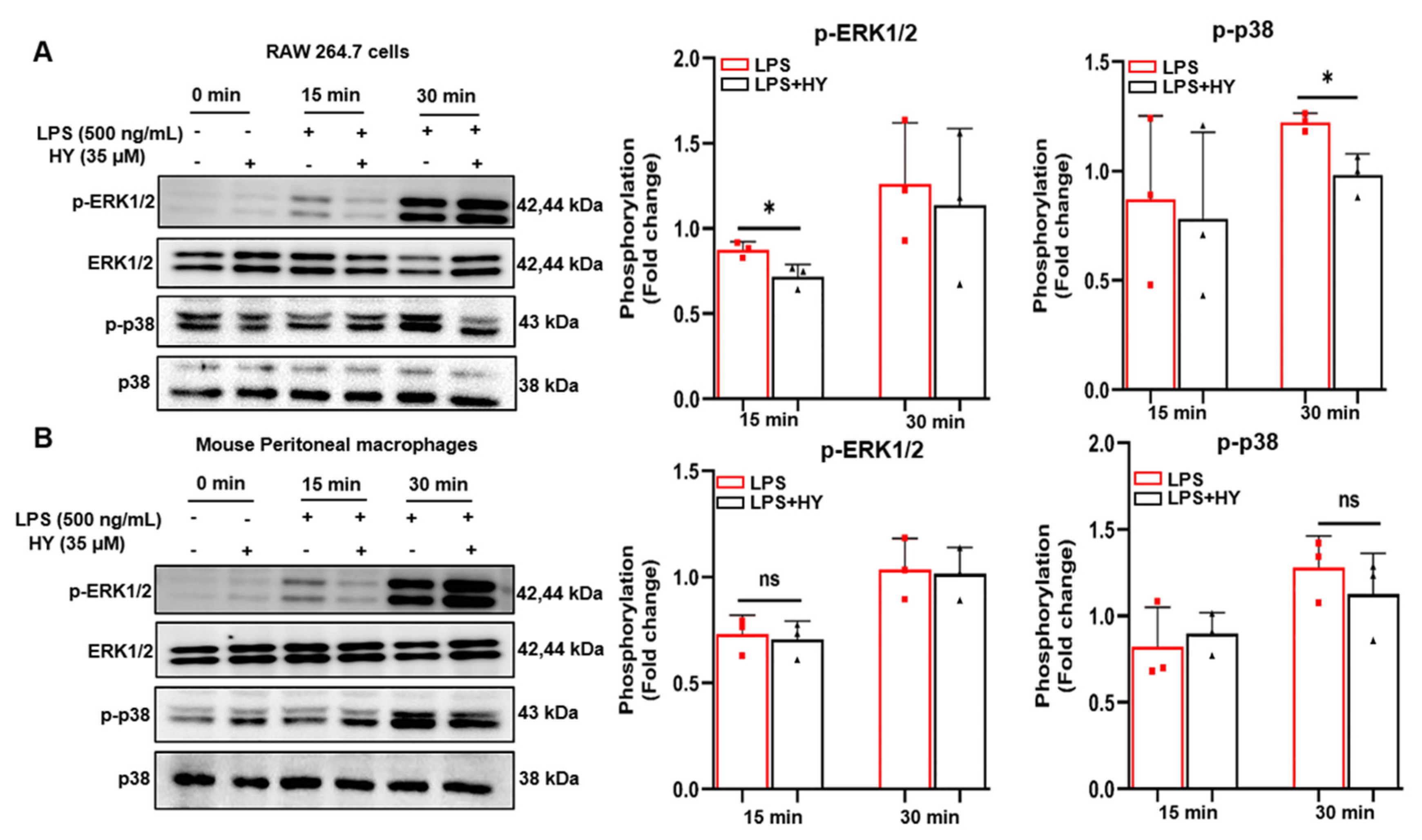
2.5. Effects of HY on MAPK Signaling Activation in LPS-Induced Macrophages
2.6. HY interacts with COX-2 to Activate NF-κB
3. Discussion
4. Materials and Methods
4.1. Plant Material and Prepared HY
4.2. Animals
4.3. Cell Culture and Treatment
4.4. Preparation and Culturing Primary Peritoneal Macrophages (PPMS)
4.5. Cell Viability Analysis
4.6. NO Production Assay
4.7. RNA Isolation, cDNA Synthesis, and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Total and Nuclear Protein Extraction and Western Blot Analysis
4.10. Immunofluorescence Assay
4.11. Molecular Modeling
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| COX-2 | Cyclooxygenase-2 |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DAPI | 4′,6-Diamidino-2-phenylindole4′,6-diamidino-2-phenylindole |
| ELISA | Enzyme-linked immunosorbent assay |
| ERK1/2 | Extracellular signal-regulated kinase 1/2 |
| FBS | Fetal bovine serum |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HY | Hyssopuside |
| IκB | Inhibitor of NF-κB |
| IL-6 | Interleukin-6 |
| IL-1β | Interleukin-1β |
| iNOS | Inducible nitric oxide synthase |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| MTT | 3-(4,5-Dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide |
| NF-κB/p65 | Nuclear factor kappa-B |
| NO | Nitric oxide |
| RT-qPCR | Real-time quantitative polymerase chain reaction |
| TNF-α | Tumor necrosis factor-α |
References
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Pierce, G.F. Macrophages: Important Physiologic and Pathologic Sources of Polypeptide Growth Factors. Am. J. Respir. Cell Mol. Biol. 1990, 2, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Wu, C.; Zhao, W.; Zhang, X.; Chen, X. Neocryptotanshinone inhibits lipopolysaccharide-induced inflammation in RAW264.7 macrophages by suppression of NF-κB and iNOS signaling pathways. Acta Pharm. Sin. B 2015, 5, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wu, Y.; Yao, X.; Fang, H.; Yu, C.; Bao, Y.; Wu, Y.; Song, Z.; Sun, Y.; Zheng, L. Ginkgolide A Ameliorates LPS-Induced Inflammatory Responses In Vitro and In Vivo. Int. J. Mol. Sci. 2017, 18, 794. [Google Scholar] [CrossRef] [Green Version]
- Karin, M.; Ben-neriah, Y. Phosporilation meets the ubiquitination: The Control of NF- kapp B Activity. Annu. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2001, 480–481, 243–268. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Chellappan, D.K.; Yap, W.S.; Bt Ahmad Suhaimi, N.A.; Gupta, G.; Dua, K. Current therapies and targets for type 2 diabetes mellitus. Panminerva Med. 2018, 60, 117–131. [Google Scholar] [CrossRef]
- Furukawa, M.; Makino, M.; Ohkoshi, E.; Uchiyama, T.; Fujimoto, Y. Terpenoids and phenethyl glucosides from Hyssopus cuspidatus (Labiatae). Phytochemistry 2011, 72, 2244–2252. [Google Scholar] [CrossRef]
- Yuan, F.; Liu, R.; Hu, M.; Rong, X.; Bai, L.; Xu, L.; Mao, Y.; Hasimu, H.; Sun, Y.; He, J. JAX2, an ethanol extract of Hyssopus cuspidatus Boriss, can prevent bronchial asthma by inhibiting MAPK/NF-κB inflammatory signaling. Phytomedicine 2019, 57, 305–314. [Google Scholar] [CrossRef]
- Shomirzoeva, O.; Li, J.; Numonov, S.; Atolikshoeva, S.; Aisa, H.A. Chemical components of Hyssopus cuspidatus Boriss.: Isolation and identification, characterization by HPLC-DAD-ESI-HRMS/MS, antioxidant activity and antimicrobial activity. Nat. Prod. Res. 2020, 34, 534–540. [Google Scholar] [CrossRef]
- Liu, X.; Wang, G.; Lv, C.; Song, Z.; Bao, Y.; Wang, S.; Huang, Y. Phytochemistry Letters Two new phenolic acids and one new phenolic glycoside from Hyssopus cuspidatus Boriss and their anti-inflammatory activities. Phytochem. Lett. 2021, 46, 15–20. [Google Scholar] [CrossRef]
- Hankittichai, P.; Buacheen, P.; Pitchakarn, P.; Takuathung, M.N.; Wikan, N.; Smith, D.R.; Potikanond, S.; Nimlamool, W. Artocarpus lakoocha Extract Inhibits LPS-Induced Inflammatory Response in RAW 264.7 Macrophage cells. Int. J. Mol. Sci. 2020, 21, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farha, M.A.; Brown, E.D. Strategies for target identification of antimicrobial natural products. Nat. Prod. Rep. 2016, 33, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Megeed, R.M.; El Newary, S.A.; Kadry, M.O.; Ghanem, H.Z.; El-Shesheny, R.A.; Said-Al Ahl, H.A.H.; Abdel-Hamid, A.H.Z. Hyssopus officinalis exerts hypoglycemic effects on streptozotocin-induced diabetic rats via modulating GSK-3β, C-fos, NF-κB, ABCA1 and ABGA1 gene expression. J. Diabetes Metab. Disord. 2020, 19, 483–491. [Google Scholar] [CrossRef]
- Vlase, L.; Benedec, D.; Hanganu, D.; Damian, G.; Csillag, I.; Sevastre, B.; Mot, A.C.; Silaghi-Dumitrescu, R.; Tilea, I. Evaluation of antioxidant and antimicrobial activities and phenolic profile for Hyssopus officinalis, Ocimum basilicum and Teucrium chamaedrys. Molecules 2014, 19, 5490–5507. [Google Scholar] [CrossRef]
- Ma, X.; Ma, X.; Ma, Z.; Wang, J.; Sun, Z.; Yu, W.; Li, F.; Ding, J. Effect of Hyssopus officinalis L. on inhibiting airway inflammation and immune regulation in a chronic asthmatic mouse model. Exp. Ther. Med. 2014, 8, 1371–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, H.; Miyazaki, H.; Asakawa, C.; Amano, M.; Yoshihara, T.; Mizutani, J. Isolation of α-glusosidase inhibitors from hyssop (Hyssopus officinalis). Phytochemistry 2004, 65, 91–97. [Google Scholar] [CrossRef]
- Chen, G.W.; Chen, T.Y.; Yang, P.M. Differential effect of herbal tea extracts on free fatty acids-, ethanol- and acetaminophen-induced hepatotoxicity in FL83B hepatocytes. Drug Chem. Toxicol. 2019, 9, 1–6. [Google Scholar] [CrossRef]
- Gaieski, D.F.; Edwards, J.M.; Kallan, M.J.; Carr, B.G. Benchmarking the incidence and mortality of severe sepsis in the united states. Crit. Care Med. 2013, 41, 1167–1174. [Google Scholar] [CrossRef]
- Goulopoulou, S.; McCarthy, C.G.; Clinton Webb, R. Toll-like receptors in the vascular system: Sensing the dangers within. Pharmacol. Rev. 2016, 68, 142–167. [Google Scholar] [CrossRef] [PubMed]
- Takakura, A.; Zandi-Nejad, K. Lactate-induced activation of HCA2 improves survival in mice with sepsis. FASEB J. 2019, 33, 7625–7634. [Google Scholar] [CrossRef] [PubMed]
- Arima, H.; Motoyama, K.; Matsukawa, A.; Nishimoto, Y.; Hirayama, F.; Uekama, K. Inhibitory effects of dimethylacetyl-β-cyclodextrin on lipopolysaccharide-induced macrophage activation and endotoxin shock in mice. Biochem. Pharmacol. 2005, 70, 1506–1517. [Google Scholar] [CrossRef]
- Amaro-Leal, Â.; Shvachiy, L.; Pinto, R.; Geraldes, V.; Rocha, I.; Mota-Filipe, H. Therapeutic effects of IkB kinase inhibitor during systemic inflammation. Int. Immunopharmacol. 2020, 84, 106509. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, A.; Xie, K.; Yu, Y. Dietary supplementation with high fiber alleviates oxidative stress and inflammatory responses caused by severe sepsis in mice without altering microbiome diversity. Front. Physiol. 2019, 9, 1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, A.J.; Ae, R.K.; Hae, Y.C.; Jae, S.C. In vitro antioxidant activity of some selected Prunus species in Korea. Arch. Pharm. Res. 2002, 25, 865–872. [Google Scholar]
- Yang, G.; Ham, I.; Choi, H.Y. Anti-inflammatory effect of prunetin via the suppression of NF-κB pathway. Food Chem. Toxicol. 2013, 58, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Yeung, Y.T.; Aziz, F.; Guerrero-Castilla, A.; Arguelles, S. Signaling Pathways in Inflammation and Anti-inflammatory Therapies. Curr. Pharm. Des. 2018, 24, 1449–1484. [Google Scholar] [CrossRef] [PubMed]
- Picon-Ruiz, M.; Morata-Tarifa, C.; Valle-Goffin, J.J.; Friedman, E.R.; Slingerland, J.M. Obesity and adverse breast cancer risk and outcome: Mechanistic insights and strategies for intervention. CA Cancer J. Clin. 2017, 67, 378–397. [Google Scholar] [CrossRef]
- Clark, A.R.; Dean, J.L.E.; Saklatvala, J. Post-transcriptional regulation of gene expression by mitogen-activated protein kinase p38. FEBS Lett. 2003, 546, 37–44. [Google Scholar] [CrossRef]
- Su, B.; Karin, M. Mitogen-activated protein kinase cascades and regulation of gene expression. Curr. Opin. Immunol. 1996, 8, 402–411. [Google Scholar] [CrossRef]
- Wang, J.; Gao, L.; Lee, Y.M.; Kalesh, K.A.; Lin, Q. Target identification of natural and traditional medicines with quantitative chemical proteomics approaches. Pharmacol. Ther. 2016, 162, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Park, K.I.; Kim, G.S.; An, S.J.; Chung, H.J.; Lee, W.S.; Kim, E.H.; Shin, S.C.; Jin, A.K.; Nagappan, A.; Lee, D.H. Regulation of Proinflammatory Mediators via NF-κB and p38 MAPK-Dependent Mechanisms in RAW 264.7 Macrophages by Polyphenol Components Isolated from Korea Lonicera japonica THUNB. Evid.-Based Complementary Altern. Med. 2012, 2012, 828521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Zhang, D.M.; Song, Z.B.; Hou, Y.Q.; Bao, Y.L.; Sun, L.G.; Yu, C.L.; Li, Y.X.; Johnson, R. Protumoral TSP50 Regulates Macrophage Activities and Polarization via Production of TNF-α and IL-1β, and Activation of the NF-κB Signaling Pathway. PLoS ONE 2015, 10, e0145095. [Google Scholar]
- Islam, M.T.; Zihad, S.; Rahman, M.S.; Sifat, N.; Rouf, R. Agathisflavone: Botanical sources, therapeutic promises, and molecular docking study. Int. Union Biochem. Mol. Biol. Life 2019, 71, 1192–1200. [Google Scholar] [CrossRef]
- Taidi, L.; Maurady, A.; Britel, M.R. Molecular docking study and molecular dynamic simulation of human cyclooxygenase-2 (COX-2) with selected eutypoids. J. Biomol. Struct. Dyn. 2020, 15, 1–16. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Su, J.; Wang, G.; Zheng, L.; Wang, G.; Sun, Y.; Bao, Y.; Wang, S.; Huang, Y. Discovery of Phenolic Glycoside from Hyssopus cuspidatus Attenuates LPS-Induced Inflammatory Responses by Inhibition of iNOS and COX-2 Expression through Suppression of NF-κB Activation. Int. J. Mol. Sci. 2021, 22, 12128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212128
Liu X, Su J, Wang G, Zheng L, Wang G, Sun Y, Bao Y, Wang S, Huang Y. Discovery of Phenolic Glycoside from Hyssopus cuspidatus Attenuates LPS-Induced Inflammatory Responses by Inhibition of iNOS and COX-2 Expression through Suppression of NF-κB Activation. International Journal of Molecular Sciences. 2021; 22(22):12128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212128
Chicago/Turabian StyleLiu, Xingyu, Jie Su, Geng Wang, Lihua Zheng, Guannan Wang, Ying Sun, Yongli Bao, Shuyue Wang, and Yanxin Huang. 2021. "Discovery of Phenolic Glycoside from Hyssopus cuspidatus Attenuates LPS-Induced Inflammatory Responses by Inhibition of iNOS and COX-2 Expression through Suppression of NF-κB Activation" International Journal of Molecular Sciences 22, no. 22: 12128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212128