
Generation of Skeletal Muscle Organoids from Human Pluripotent Stem Cells to Model Myogenesis and Muscle Regeneration

 ,
,  and
and
Abstract
:1. Introduction
2. Results
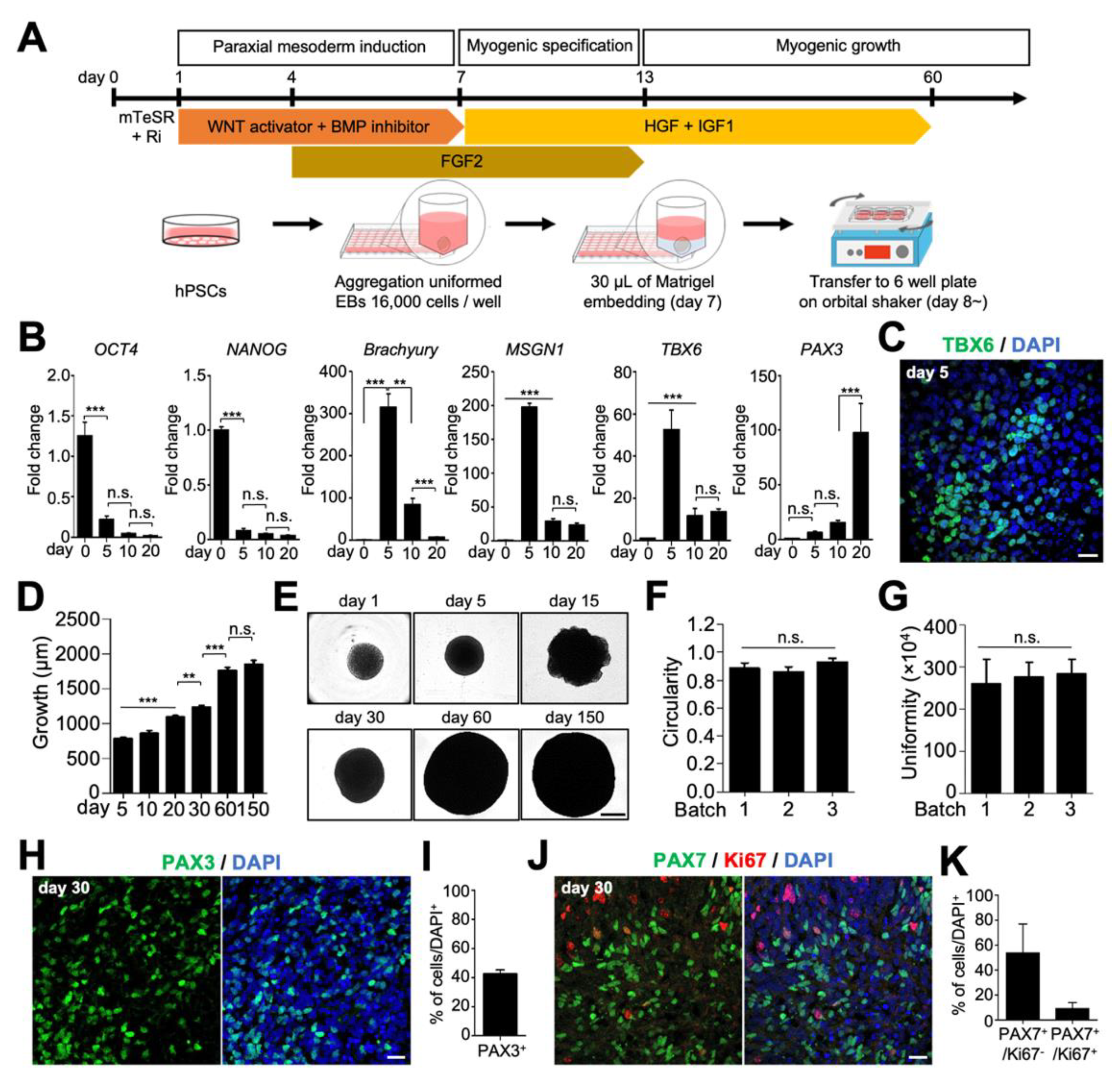
2.1. Generation and Characterization of hSkMOs from hPSCs
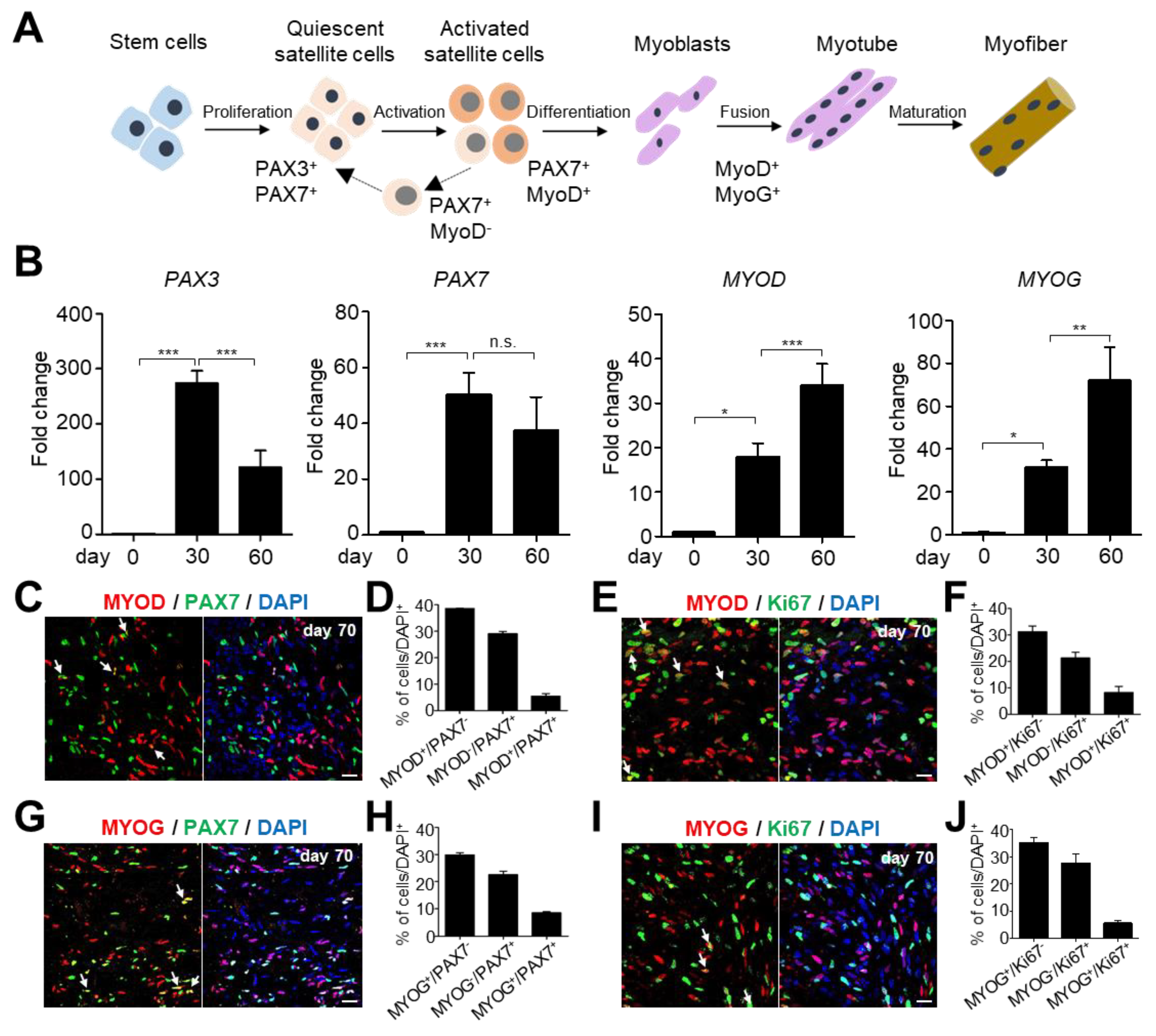
2.2. Identifying and Characterizing Different Types of Skeletal Muscle Stem/Progenitor Cells during Myogenesis in hSkMOs
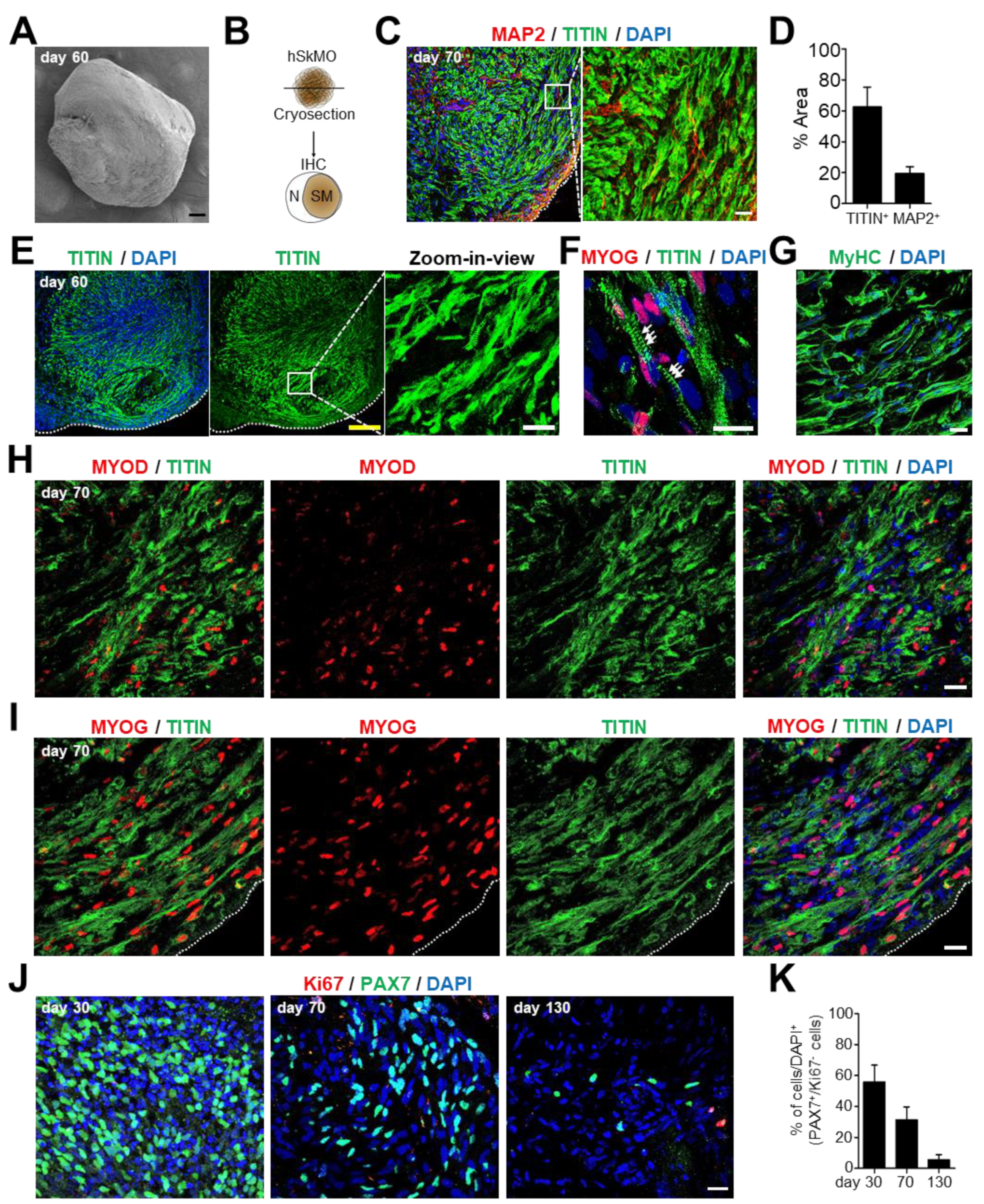
2.3. Maturation of Skeletal Muscle Cells in hSkMOs
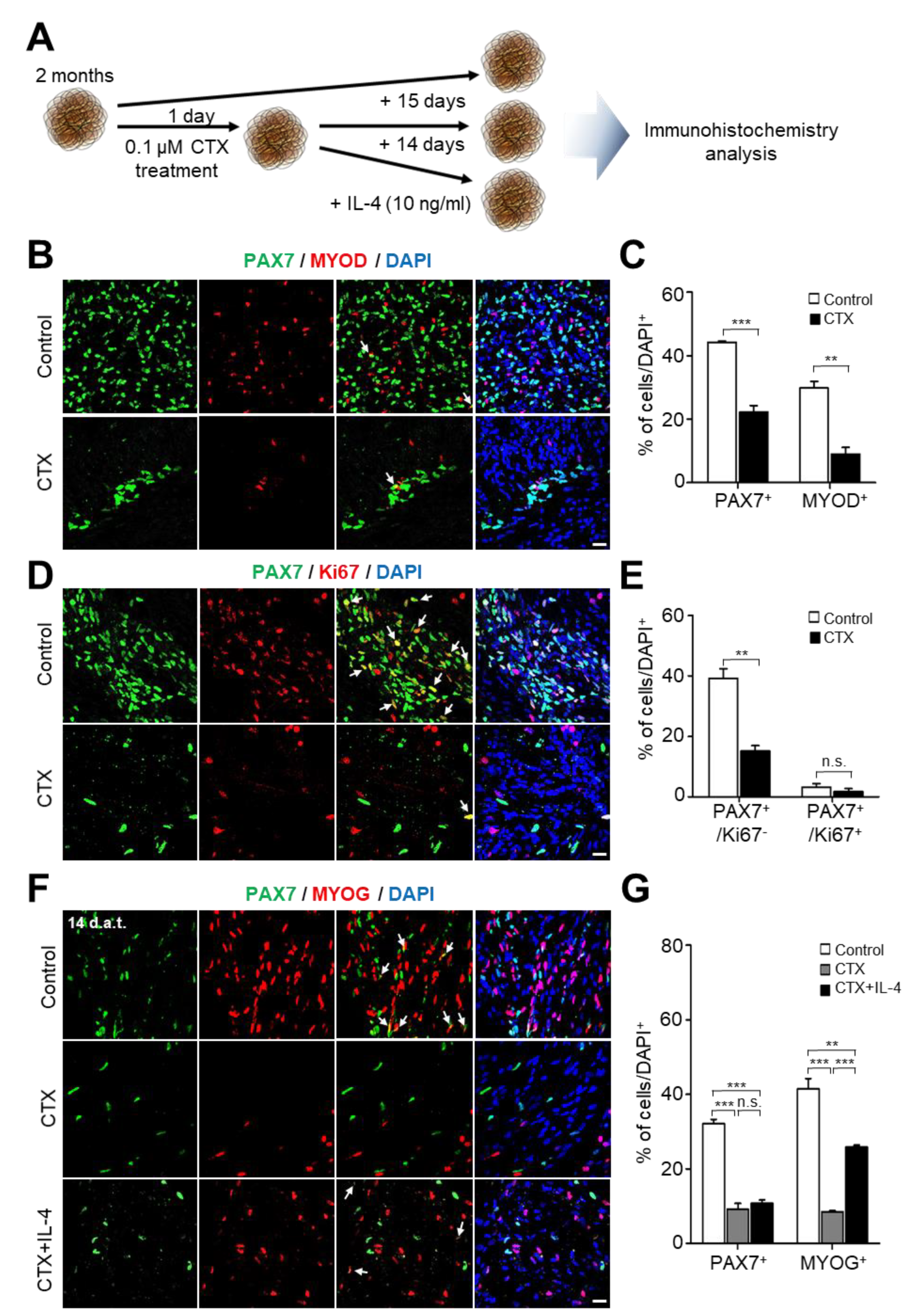
2.4. Toxin-Induced Muscular Degeneration and Regeneration Capacity of hSkMOs
3. Discussion
4. Materials and Methods
4.1. Culture of Human Pluripotent Stem Cells (hPSCs)
4.2. Generation of Human Skeletal Muscle Organoids
4.3. Immunofluorescence Analyses Using Cryosection
4.4. RNA Extraction, Brachyury Reverse Transcription, Quantitative Real-Time RT-PCR Analysis
4.5. Scanning Electron Microscopy (SEM)
4.6. Muscle Injury Procedures and Regeneration
4.7. Quantification and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamakawa, H.; Kusumoto, D.; Hashimoto, H.; Yuasa, S. Stem cell aging in skeletal muscle regeneration and disease. Int. J. Mol. Sci. 2020, 21, 1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimczak, A.; Kozlowska, U.; Kurpisz, M. Muscle stem/progenitor cells and mesenchymal stem cells of bone marrow origin for skeletal muscle regeneration in muscular dystrophies. Arch. Immunol. Ther. Exp. (Warsz.) 2018, 66, 341–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olguín, H.C.; Pisconti, A. Marking the tempo for myogenesis: Pax7 and the regulation of muscle stem cell fate decisions. J. Cell. Mol. Med. 2012, 16, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493. [Google Scholar] [CrossRef]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [Green Version]
- Jo, J.; Xiao, Y.; Sun, A.X.; Cukuroglu, E.; Tran, H.-D.; Göke, J.; Tan, Z.Y.; Saw, T.Y.; Tan, C.-P.; Lokman, H. Midbrain-like organoids from human pluripotent stem cells contain functional dopaminergic and neuromelanin-producing neurons. Cell Stem Cell 2016, 19, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Hor, J.-H.; Ng, S.-Y. Generating ventral spinal organoids from human induced pluripotent stem cells. Methods Cell Biol. 2020, 159, 257–277. [Google Scholar] [CrossRef]
- Cotovio, J.P.; Fernandes, T.G. Production of human pluripotent stem cell-derived hepatic cell lineages and liver organoids: Current status and potential applications. Bioengineering 2020, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Cruz, N.M.; Freedman, B.S. Differentiation of human kidney organoids from pluripotent stem cells. Methods Cell Biol. 2019, 153, 133–150. [Google Scholar] [CrossRef]
- Koike, H.; Iwasawa, K.; Ouchi, R.; Maezawa, M.; Kimura, M.; Kodaka, A.; Nishii, S.; Thompson, W.L.; Takebe, T. Engineering human hepato-biliary-pancreatic organoids from pluripotent stem cells. Nat. Protoc. 2021, 16, 919–936. [Google Scholar] [CrossRef]
- Chal, J.; Al Tanoury, Z.; Hestin, M.; Gobert, B.; Aivio, S.; Hick, A.; Cherrier, T.; Nesmith, A.P.; Parker, K.K.; Pourquié, O. Generation of human muscle fibers and satellite-like cells from human pluripotent stem cells in vitro. Nat. Protoc. 2016, 11, 1833–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, H.; Fujiwara, W.; Gonzalez, K.; Jan, M.; Liebscher, S.; Van Handel, B.; Schenke-Layland, K.; Pyle, A.D. In vivo human somitogenesis guides somite development from hPSCs. Cell Rep. 2017, 18, 1573–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hicks, M.R.; Hiserodt, J.; Paras, K.; Fujiwara, W.; Eskin, A.; Jan, M.; Xi, H.; Young, C.S.; Evseenko, D.; Nelson, S.F. ERBB3 and NGFR mark a distinct skeletal muscle progenitor cell in human development and hPSCs. Nat. Cell Biol. 2018, 20, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.-M.F.; Fischer, C.; Urzi, A.; Vidal, R.; Kunz, S.; Ruffault, P.-L.; Kabuss, L.; Hube, I.; Gazzerro, E.; Birchmeier, C. Self-organizing 3D human trunk neuromuscular organoids. Cell Stem Cell 2020, 26, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Maffioletti, S.M.; Sarcar, S.; Henderson, A.B.; Mannhardt, I.; Pinton, L.; Moyle, L.A.; Steele-Stallard, H.; Cappellari, O.; Wells, K.E.; Ferrari, G. Three-dimensional human iPSC-derived artificial skeletal muscles model muscular dystrophies and enable multilineage tissue engineering. Cell Rep. 2018, 23, 899–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, J.; Yang, L.; Tran, H.D.; Yu, W.; Sun, A.X.; Chang, Y.Y.; Jung, B.C.; Lee, S.J.; Saw, T.Y.; Xiao, B. Lewy body–like inclusions in human midbrain organoids carrying glucocerebrosidase and α-synuclein mutations. Ann. Neurol. 2021, 90, 490–505. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [Green Version]
- Hofer, M.; Lutolf, M.P. Engineering organoids. Nat. Rev. Mater. 2021, 6, 402–420. [Google Scholar] [CrossRef]
- Lagha, M.; Sato, T.; Bajard, L.; Daubas, P.; Esner, M.; Montarras, D.; Relaix, F.; Buckingham, M. Regulation of skeletal muscle stem cell behavior by Pax3 and Pax7. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2008; pp. 307–315. [Google Scholar]
- Mukund, K.; Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdiscip Rev. Syst. Biol. Med. 2020, 12, e1462. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Akasaki, Y.; Sato, K.; Ouchi, N.; Izumiya, Y.; Walsh, K. Insulin-like 6 is induced by muscle injury and functions as a regenerative factor. J. Biol. Chem. 2010, 285, 36060–36069. [Google Scholar] [CrossRef] [Green Version]
- Langone, F.; Cannata, S.; Fuoco, C.; Lettieri Barbato, D.; Testa, S.; Nardozza, A.P.; Ciriolo, M.R.; Castagnoli, L.; Gargioli, C.; Cesareni, G. Metformin protects skeletal muscle from cardiotoxin induced degeneration. PLoS ONE 2014, 9, e114018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costamagna, D.; Duelen, R.; Penna, F.; Neumann, D.; Costelli, P.; Sampaolesi, M. Interleukin-4 administration improves muscle function, adult myogenesis, and lifespan of colon carcinoma-bearing mice. J. Cachexia Sarcopenia Muscle 2020, 11, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almada, A.E.; Wagers, A.J. Molecular circuitry of stem cell fate in skeletal muscle regeneration, ageing and disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 267–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horsley, V.; Jansen, K.M.; Mills, S.T.; Pavlath, G.K. IL-4 acts as a myoblast recruitment factor during mammalian muscle growth. Cell 2003, 113, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Khodabukus, A.; Rao, L.; Vandusen, K.; Abutaleb, N.; Bursac, N. Engineered skeletal muscles for disease modeling and drug discovery. Biomaterials 2019, 221, 119416. [Google Scholar] [CrossRef]
- Cho, S.; Jang, J. Recent trends in biofabrication technologies for studying skeletal muscle tissue-related diseases. Front. Bioeng. Biotechnol. 2021, 9, 782333. [Google Scholar] [CrossRef]
- Mavrommatis, L.; Jeong, H.-W.; Gomez-Giro, G.; Stehling, M.; Kienitz, M.-C.; Psathaki, O.E.; Bixel, G.; Morosan-Puopolo, G.; Gerovska, D.; Arauzo-Bravo, M.J. Human skeletal muscle organoids model fetal myogenesis and sustain uncommitted PAX7 myogenic progenitors. bioRxiv 2020, preprint. [Google Scholar] [CrossRef]
- Guardiola, O.; Andolfi, G.; Tirone, M.; Iavarone, F.; Brunelli, S.; Minchiotti, G. Induction of acute skeletal muscle regeneration by cardiotoxin injection. JoVE (J. Vis. Exp.) 2017, 119, e54515. [Google Scholar] [CrossRef]
- Williams, H.F.; Mellows, B.A.; Mitchell, R.; Sfyri, P.; Layfield, H.J.; Salamah, M.; Vaiyapuri, R.; Collins-Hooper, H.; Bicknell, A.B.; Matsakas, A. Mechanisms underpinning the permanent muscle damage induced by snake venom metalloprotease. PLoS Negl. Trop. Dis. 2019, 13, e0007041. [Google Scholar] [CrossRef] [Green Version]
- Joe, A.W.; Yi, L.; Natarajan, A.; Le Grand, F.; So, L.; Wang, J.; Rudnicki, M.A.; Rossi, F. Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nat. Cell Biol. 2010, 12, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Uezumi, A.; Fukada, S.-I.; Yamamoto, N.; Takeda, S.I.; Tsuchida, K. Mesenchymal progenitors distinct from satellite cells contribute to ectopic fat cell formation in skeletal muscle. Nat. Cell Biol. 2010, 12, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Lee, J.Y.; Park, C.-H.; Eum, J.H.; Jung, S.K.; Han, A.-R.; Seol, D.-W.; Lee, J.S.; Shin, H.S.; Im, J.H. Cryopreserved human oocytes and cord blood cells can produce somatic cell nuclear transfer-derived pluripotent stem cells with a homozygous HLA type. Stem Cell Rep. 2020, 15, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, J.E.; Lee, C.W.; Shim, S.H.; Lee, D.R. Rapid production and genetic stability of human mesenchymal progenitor cells derived from human somatic cell nuclear transfer-derived pluripotent stem cells. Int. J. Mol. Sci. 2021, 22, 9238. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody Name | Catalogue Number | Company | Host Species | Dilution |
|---|---|---|---|---|
| Ki67 | 550609 | BD | Mouse | 1:200 |
| Ki67 | ab15580 | Abcam | Rabbit | 1:200 |
| PAX3 | PAX3 | DSHB | Mouse | 1:100 |
| PAX7 | PAX7 | DSHB | Mouse | 1:200 |
| MAP2 | ab5392 | Abcam | Chicken | 1:2000 |
| MyHC | 5-6-s | DSHB | Mouse | 1:25 |
| MYOD | ab133627 | Abcam | Rabbit | 1:500 |
| MYOG | ab124800 | Abcam | Rabbit | 1:500 |
| TBX6 | AF4744 | R&D Systems | Goat | 1:50 |
| TITIN | 9D10 | DSHB | Mouse | 1:200 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | CAAGATCATCAGCAATGCCTCCTG | GCCTGCTTCACCACCTTCTTGA |
| OCT4 | GTGGAGGAAGCTGACAACAA | ATTCTCCAGGTTGCCTCTCA |
| NANOG | TTTGTGGGCCTGAAGAAAACT | AGGGCTGTCCTGAATAAGCAG |
| Brachyury | TTCATAGCGGTGACTGCTTATCA | CACCCCCATTGGGAGTACC |
| MSGN1 | CTGCACACCCTCCGGAATT | CTCTGCCGCGGTTAAGGAG |
| TBX6 | CATCCACGAGAATTGTACCCG | AGCAATCCAGTTTAGGGGTGT |
| PAX3 | AGCTCGGCGGTGTTTTTATCA | CTGCACAGGATCTTGGAGACG |
| PAX7 | CGTGCTCAGAATCAAGTTCG | GTCAGGTTCCGACTCCACAT |
| MYOD | ACTTTCTGGAGCCCTCCTGGCA | TTTGTTGCACTACACAGCATG |
| MYOG | GCCAACCCAGGGGATCAT | CCCGGCTTGGAAGACAATCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, M.-K.; Bang, J.S.; Lee, J.E.; Tran, H.-D.; Park, G.; Lee, D.R.; Jo, J. Generation of Skeletal Muscle Organoids from Human Pluripotent Stem Cells to Model Myogenesis and Muscle Regeneration. Int. J. Mol. Sci. 2022, 23, 5108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095108
Shin M-K, Bang JS, Lee JE, Tran H-D, Park G, Lee DR, Jo J. Generation of Skeletal Muscle Organoids from Human Pluripotent Stem Cells to Model Myogenesis and Muscle Regeneration. International Journal of Molecular Sciences. 2022; 23(9):5108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095108
Chicago/Turabian StyleShin, Min-Kyoung, Jin Seok Bang, Jeoung Eun Lee, Hoang-Dai Tran, Genehong Park, Dong Ryul Lee, and Junghyun Jo. 2022. "Generation of Skeletal Muscle Organoids from Human Pluripotent Stem Cells to Model Myogenesis and Muscle Regeneration" International Journal of Molecular Sciences 23, no. 9: 5108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095108