Changes in Energy Reserves and Gene Expression Elicited by Freezing and Supercooling in the Antarctic Midge, Belgica antarctica


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Temperature Treatments
2.3. Larval Survival after Cold Exposure
2.4. Cell Survival after Cold Exposure
2.5. Energy Store Analyses
2.6. Gene Expression Measurements
2.7. Statistical Analyses
3. Results
3.1. Larval Survival
3.2. Cell Survival
3.3. Energy Stores
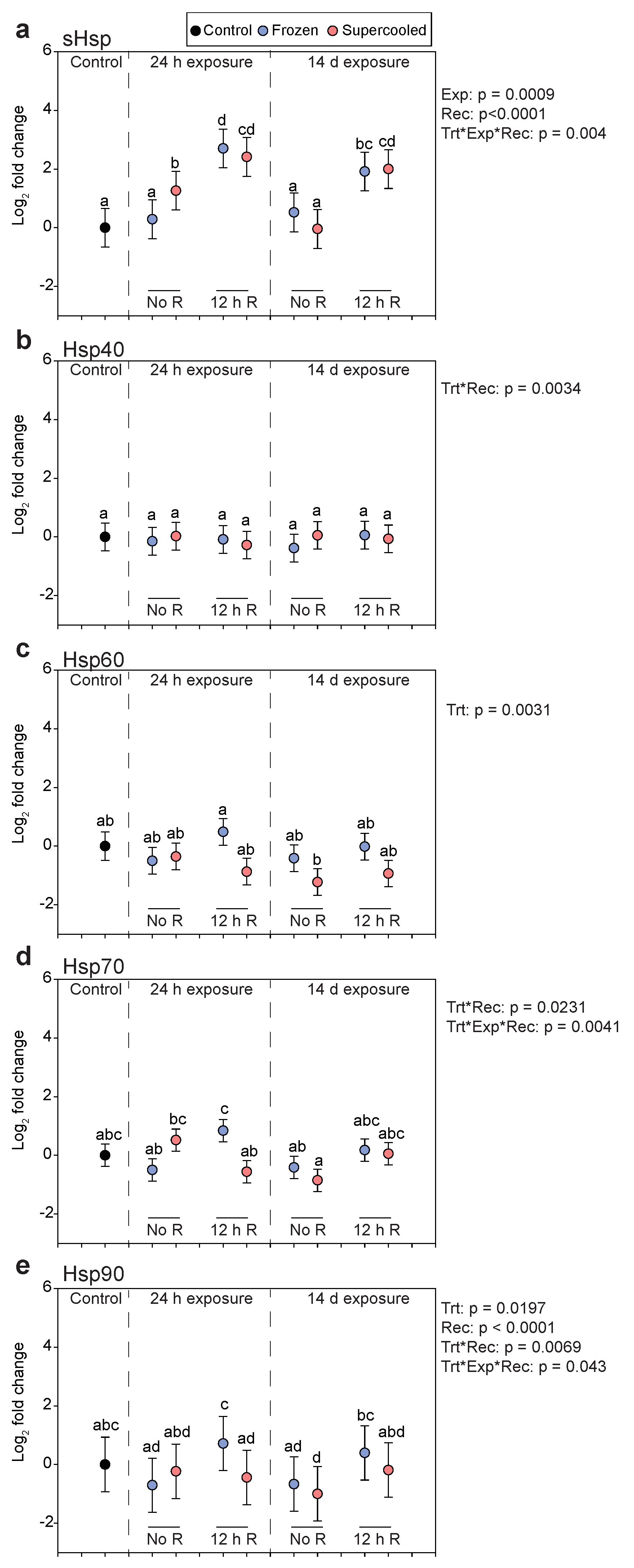
3.4. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Overgaard, J.; MacMillan, H.A. The integrative physiology of insect chill tolerance. Annu. Rev. Physiol. 2017, 79, 187–208. [Google Scholar] [CrossRef]
- Lee, R.E. Insect cold-hardiness—To freeze or not to freeze—How insects survive low temperatures. Bioscience 1989, 39, 308–313. [Google Scholar] [CrossRef]
- Lee, R.E. A primer on insect cold tolerance. In Low Temperature Biology of Insects; Denlinger, D.L., Lee, R.E., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 3–34. [Google Scholar]
- Sinclair, B.J.; Alvarado, L.E.C.; Ferguson, L.V. An invitation to measure insect cold tolerance: Methods, approaches, and workflow. J. Therm. Biol. 2015, 53, 180–197. [Google Scholar] [CrossRef] [Green Version]
- Toxopeus, J.; Sinclair, B.J. Mechanisms underlying insect freeze tolerance. Biol. Rev. 2018, 93, 1891–1914. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Chown, S.L. Climatic variability and hemispheric differences in insect cold tolerance: Support from southern Africa. Funct. Ecol. 2005, 19, 214–221. [Google Scholar] [CrossRef]
- Toxopeus, J.; Kostal, V.; Sinclair, B.J. Evidence for non-colligative function of small cryoprotectants in a freeze-tolerant insect. Proc. R. Soc. B 2019, 286, 20190050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toxopeus, J.; Marteaux, L.E.D.; Sinclair, B.J. How crickets become freeze tolerant: The transcriptomic underpinnings of acclimation in Gryllus veletis. Comp. Biochem. Physiol. Part D Genomics Proteom. 2019, 29, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Marteaux, L.E.D.; Hůla, P.; Koštál, V. Transcriptional analysis of insect extreme freeze tolerance. Proc. R. Soc. B Biol. Sci. 2019, 286, 20192019. [Google Scholar] [CrossRef] [Green Version]
- Teets, N.M.; Denlinger, D.L. Surviving in a frozen desert: Environmental stress physiology of terrestrial Antarctic arthropods. J. Exp. Biol. 2014, 217, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Shimada, K.; Ohyama, Y.; Pan, C.X. Cold-hardiness of the Antarctic winged midge Parochlus steinenii during the active season at King George Island. Polar Biol. 1991, 11, 311–314. [Google Scholar] [CrossRef]
- Lee, R.E.; Baust, J.G. Adaptive potential of terrestrial invertebrates: Maritime Antarctica. Antarct. JUS 1982, 16, 175–176. [Google Scholar]
- Lee, R.E.; Denlinger, D.L. Stress tolerance in a polyextremophile: The southernmost insect. Can. J. Zool. 2015, 93, 679–686. [Google Scholar] [CrossRef]
- Worland, M.R. Eretmoptera murphyi: Pre-adapted to survive a colder climate. Physiol. Entomol. 2010, 35, 140–147. [Google Scholar] [CrossRef]
- Everatt, M.J.; Worland, M.R.; Bale, J.S.; Convey, P.; Hayward, S.A.L. Pre-adapted to the maritime Antarctic?—Rapid cold hardening of the midge, Eretmoptera murphyi. J. Insect Physiol. 2012, 58, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E.; Elnitsky, M.A.; Rinehart, J.P.; Hayward, S.A.L.; Sandro, L.H.; Denlinger, D.L. Rapid cold-hardening increases the freezing tolerance of the Antarctic midge Belgica antarctica. J. Exp. Biol. 2006, 209, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Kawarasaki, Y.; Teets, N.M.; Denlinger, D.L.; Lee, R.E. The protective effect of rapid cold-hardening develops more quickly in frozen versus supercooled larvae of the Antarctic midge, Belgica antarctica. J. Exp. Biol. 2013, 216, 3937–3945. [Google Scholar] [CrossRef] [Green Version]
- Kawarasaki, Y.; Teets, N.M.; Denlinger, D.L.; Lee, R.E. Alternative overwintering strategies in an Antarctic midge: Freezing vs. cryoprotective dehydration. Funct. Ecol. 2014, 28, 933–943. [Google Scholar] [CrossRef]
- Elnitsky, M.A.; Hayward, S.A.L.; Rinehart, J.P.; Denlinger, D.L.; Lee, R.E. Cryoprotective dehydration and the resistance to inoculative freezing in the Antarctic midge, Belgica antarctica. J. Exp. Biol. 2008, 211, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Kawarasaki, Y.; Teets, N.M.; Denlinger, D.L.; Lee, R.E. Wet hibernacula promote inoculative freezing and limit the potential for cryoprotective dehydration in the Antarctic midge, Belgica antarctica. Polar Biol. 2014, 37, 753–761. [Google Scholar] [CrossRef]
- Teets, N.M.; Kawarasaki, Y.; Lee, R.E.; Denlinger, D.L. Survival and energetic costs of repeated cold exposure in the Antarctic midge, Belgica antarctica: A comparison between frozen and supercooled larvae. J. Exp. Biol. 2011, 214, 806–814. [Google Scholar] [CrossRef] [Green Version]
- Kelley, J.L.; Peyton, J.T.; Fiston-Lavier, A.S.; Teets, N.M.; Yee, M.C.; Johnston, J.S.; Bustamante, C.D.; Lee, R.E.; Denlinger, D.L. Compact genome of the Antarctic midge is likely an adaptation to an extreme environment. Nat. Commun. 2014, 5, 4611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teets, N.M.; Kawarasaki, Y.; Potts, L.J.; Philip, B.N.; Gantz, J.D.; Denlinger, D.L.; Lee, R.E. Rapid cold hardening protects against sublethal freezing injury in an Antarctic insect. J. Exp. Biol. 2019, 222, jeb206011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, S.-X.; Lee, R.E. Detecting freeze injury and seasonal cold-hardening of cells and tissues in the gall fly larvae, Eurosta solidaginis (Diptera: Tephritidae) using fluorescent vital dyes. J. Insect Physiol. 2003, 49, 999–1004. [Google Scholar] [CrossRef]
- Coast, G.M.; Krasnoff, S.B. Fluid secretion by single isolated Malpighian tubules of the house cricket, Acheta domesticus, and their response to diuretic hormone. Physiol. Entomol. 1988, 13, 381–391. [Google Scholar] [CrossRef]
- Teets, N.M.; Kawarasaki, Y.; Lee, R.E.; Denlinger, D.L. Expression of genes involved in energy mobilization and osmoprotectant synthesis during thermal and dehydration stress in the Antarctic midge, Belgica antarctica. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2013, 183, 189–201. [Google Scholar] [CrossRef]
- Teets, N.M.; Kawarasaki, Y.; Lee, R.E.; Denlinger, D.L. Energetic consequences of repeated and prolonged dehydration in the Antarctic midge, Belgica antarctica. J. Insect Physiol. 2012, 58, 498–505. [Google Scholar] [CrossRef]
- Teets, N.M.; Peyton, J.T.; Colinet, H.; Renault, D.; Kelley, J.L.; Kawarasaki, Y.; Lee, R.E.; Denlinger, D.L. Gene expression changes governing extreme dehydration tolerance in an Antarctic insect. Proc. Natl. Acad. Sci. USA 2012, 109, 20744–20749. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. Roy. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Spacht, D.E.; Gantz, J.D.; Lee, R.E., Jr.; Denlinger, D.L. Onset of seasonal metabolic depression in the Antarctic midge Belgica antarctica appears to be independent of environmental cues. Physiol. Entomol. 2019. [Google Scholar] [CrossRef]
- Marshall, K.E.; Sinclair, B.J. Repeated freezing induces a trade-off between cryoprotection and egg production in the goldenrod gall fly, Eurosta solidaginis. J. Exp. Biol. 2018, 221, jeb177956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, B.J.; Chown, S.L. Deleterious effects of repeated cold exposure in a freeze-tolerant sub-Antarctic caterpillar. J. Exp. Biol. 2005, 208, 869–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, K.B.; Storey, J.M. Biochemistry of cryoprotectants. In Insects at Low Temperature; Lee, R.E., Denlinger, D.L., Eds.; Chapman and Hall: New York, NY, USA, 1991; pp. 64–93. [Google Scholar]
- Baust, J.G.; Lee, R.E. Environmental “homeothermy” in an Antarctic insect. Antarct. JUS 1981, 15, 170–172. [Google Scholar]
- Lee, J.R.; Raymond, B.; Bracegirdle, T.J.; Chades, I.; Fuller, R.A.; Shaw, J.D.; Terauds, A. Climate change drives expansion of Antarctic ice-free habitat. Nature 2017, 547, 49. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Dillon, M.E. Recent geographic convergence in diurnal and annual temperature cycling flattens global thermal profiles. Nat. Clim. Chang. 2014, 4, 988–992. [Google Scholar] [CrossRef]
- Dillon, M.E.; Woods, H.A.; Wang, G.; Fey, S.B.; Vasseur, D.A.; Telemeco, R.S.; Marshall, K.; Pincebourde, S. Life in the frequency domain: The biological impacts of changes in climate variability at multiple time scales. Integr. Comp. Biol. 2016, 56, 14–30. [Google Scholar] [CrossRef]
- Allegrucci, G.; Carchini, G.; Convey, P.; Sbordoni, V. Evolutionary geographic relationships among orthocladine chironomid midges from maritime Antarctic and sub-Antarctic islands. Biol. J. Linn. Soc. 2012, 106, 258–274. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession | Forward Primer | Reverse Primer |
|---|---|---|---|
| rpl19 | JX462670 | 5′-ACATCCACAAGCGTAAGGCTGAGA-3′ | 5′-TTCTTGTTTCTTGGTGGCGATGCG-3′ |
| sHsp | GAAK01009816 | 5′-GACACCCTTATCAGACGACTAC-3′ | 5′-CTTCTCGTTCTTCGTGCTTTG-3′ |
| Hsp40 | GAAK01004380 | 5′-ACTCTGACCGGAGAAGTGATA-3′ | 5′-CTCGCTTTGTTGGCTCTTTG-3′ |
| Hsp60 | GAAK01010161 | 5′-GTTGCAGGGAGTTGACATAC-3′ | 5′-GGCAACAGTTACACCATCTT-3′ |
| Hsp70 | GAAK01011953 | 5′-CTGCTTTGGCTTACGGTTTG-3′ | 5′-CCTTCGTCGATGGTCAAGATAG-3′ |
| Hsp90 | GAAK01011429 | 5′-CCGGTGGTAGCTTTATCATCTC-3′ | 5′-GGTAACGATGGCCTTGATCTTA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teets, N.M.; Dalrymple, E.G.; Hillis, M.H.; Gantz, J.D.; Spacht, D.E.; Lee, R.E., Jr.; Denlinger, D.L. Changes in Energy Reserves and Gene Expression Elicited by Freezing and Supercooling in the Antarctic Midge, Belgica antarctica. Insects 2020, 11, 18. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11010018
Teets NM, Dalrymple EG, Hillis MH, Gantz JD, Spacht DE, Lee RE Jr., Denlinger DL. Changes in Energy Reserves and Gene Expression Elicited by Freezing and Supercooling in the Antarctic Midge, Belgica antarctica. Insects. 2020; 11(1):18. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11010018
Chicago/Turabian StyleTeets, Nicholas M., Emma G. Dalrymple, Maya H. Hillis, J. D. Gantz, Drew E. Spacht, Richard E. Lee, Jr., and David L. Denlinger. 2020. "Changes in Energy Reserves and Gene Expression Elicited by Freezing and Supercooling in the Antarctic Midge, Belgica antarctica" Insects 11, no. 1: 18. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11010018