Enzyme-Catalyzed Production of Potato Galactan-Oligosaccharides and Its Optimization by Response Surface Methodology

 ,
,  ,
,
Abstract
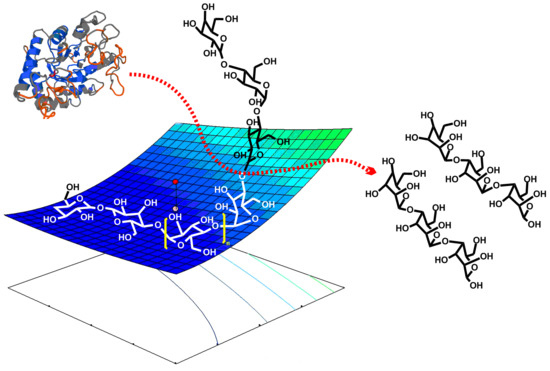
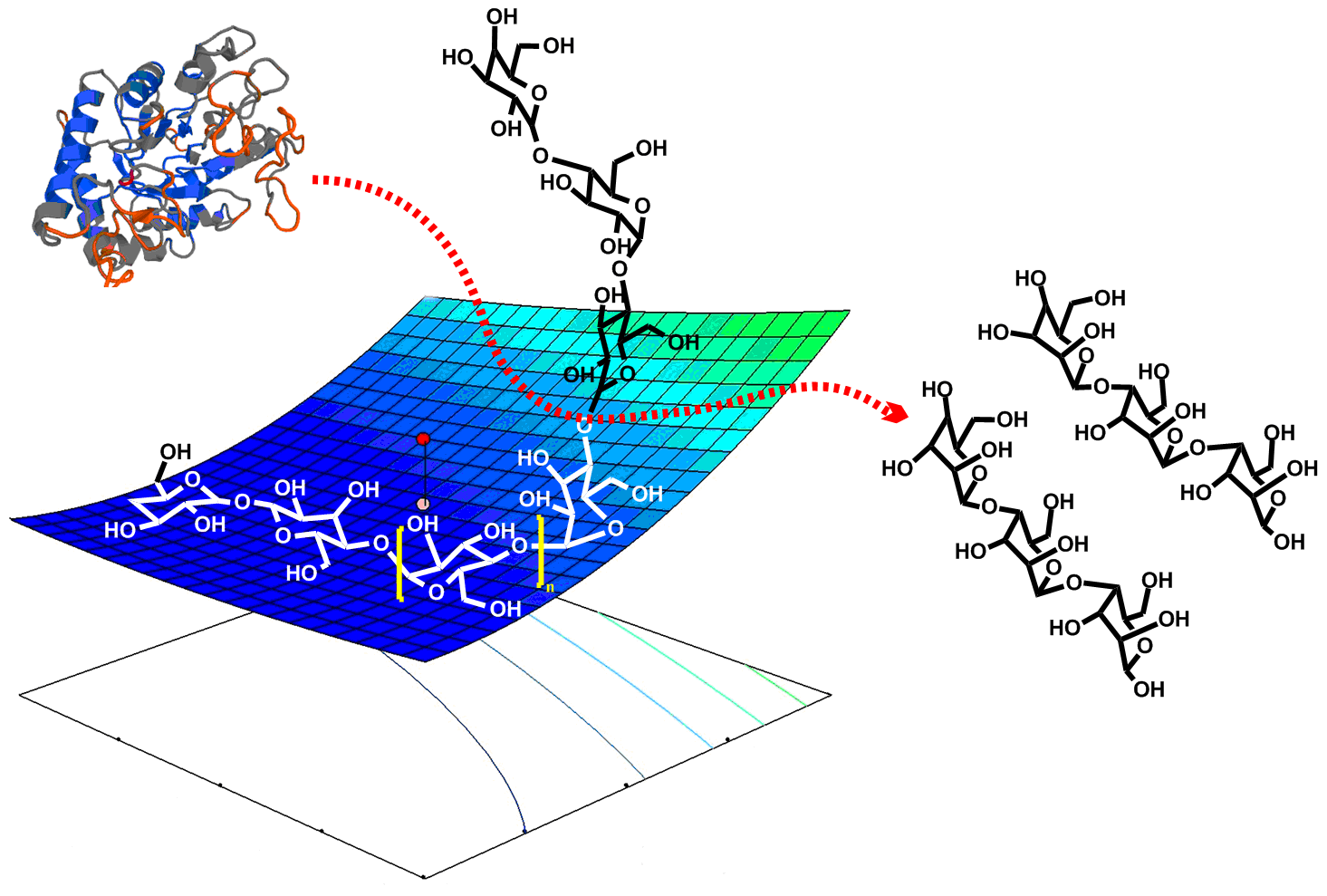
:
1. Introduction
2. Materials and Methods
2.1. Materials
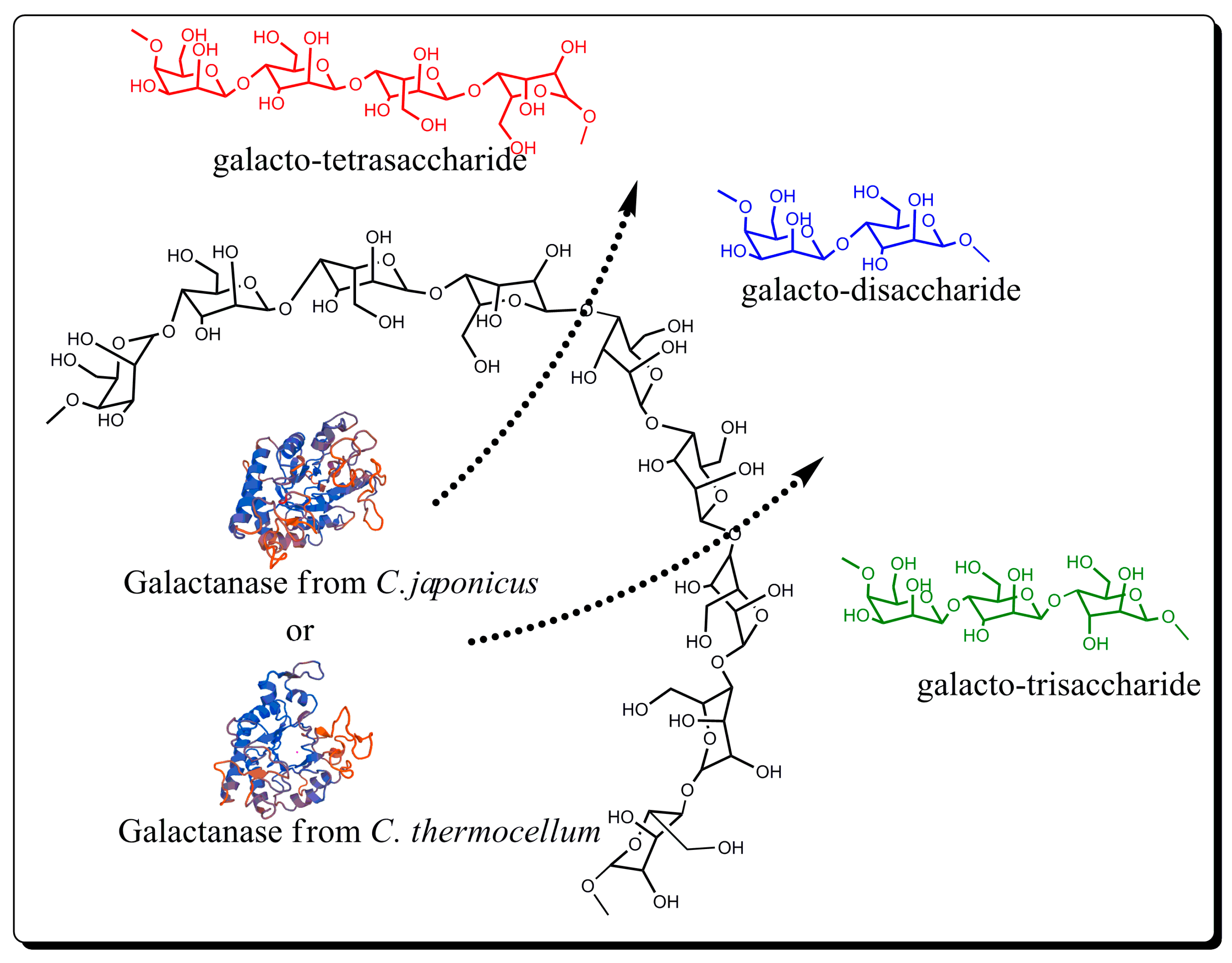
2.2. Enzymatic Hydrolysis of Potato Pectic Galactan
2.3. Size-Exclusion Chromatography
2.4. Experimental Design by RSM
2.5. Fourier Transform Infrared Spectroscopy (FT-IR)
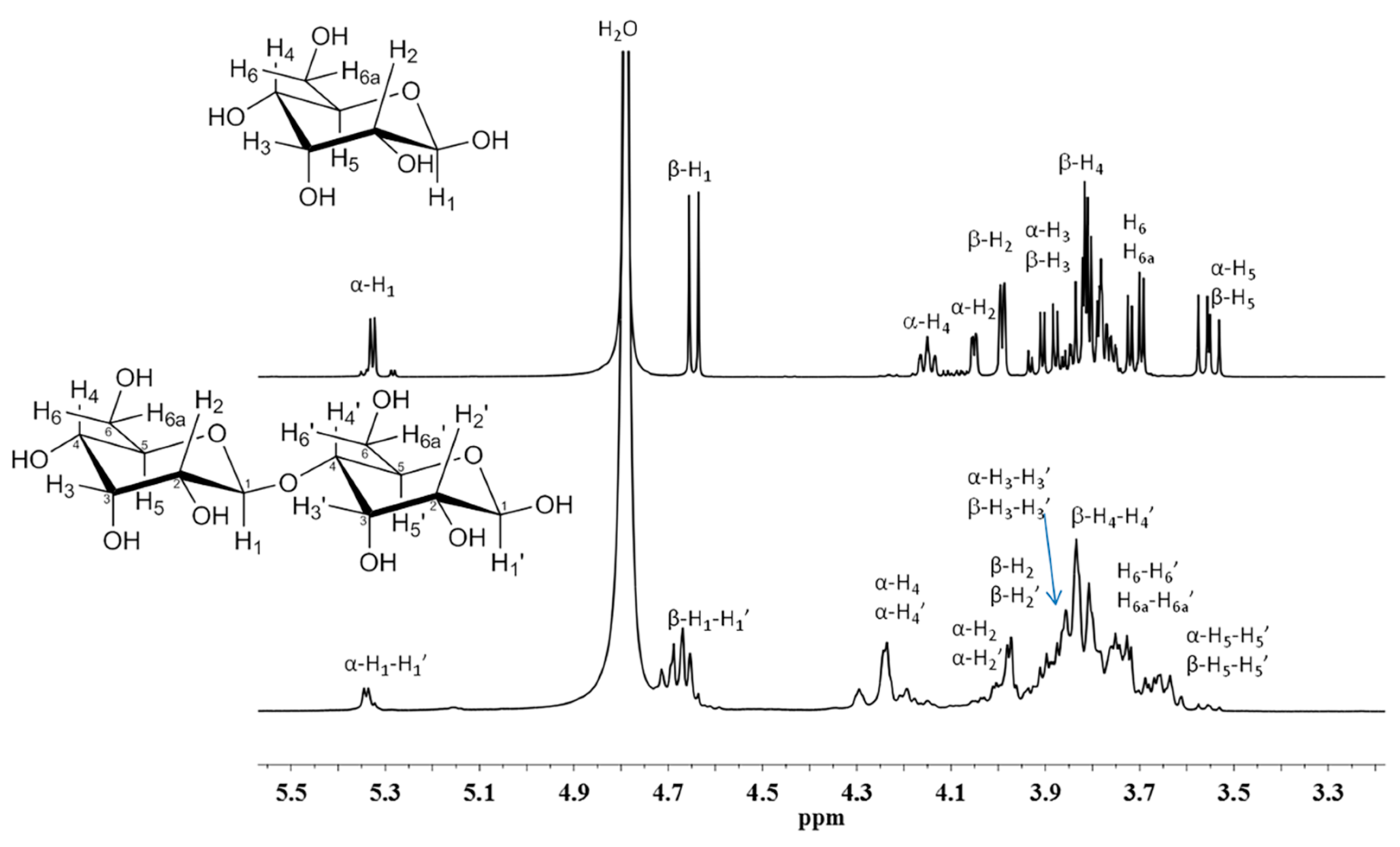
2.6. Nuclear Magnetic Resonance Spectroscopy (1H-NMR)
3. Results and Discussion
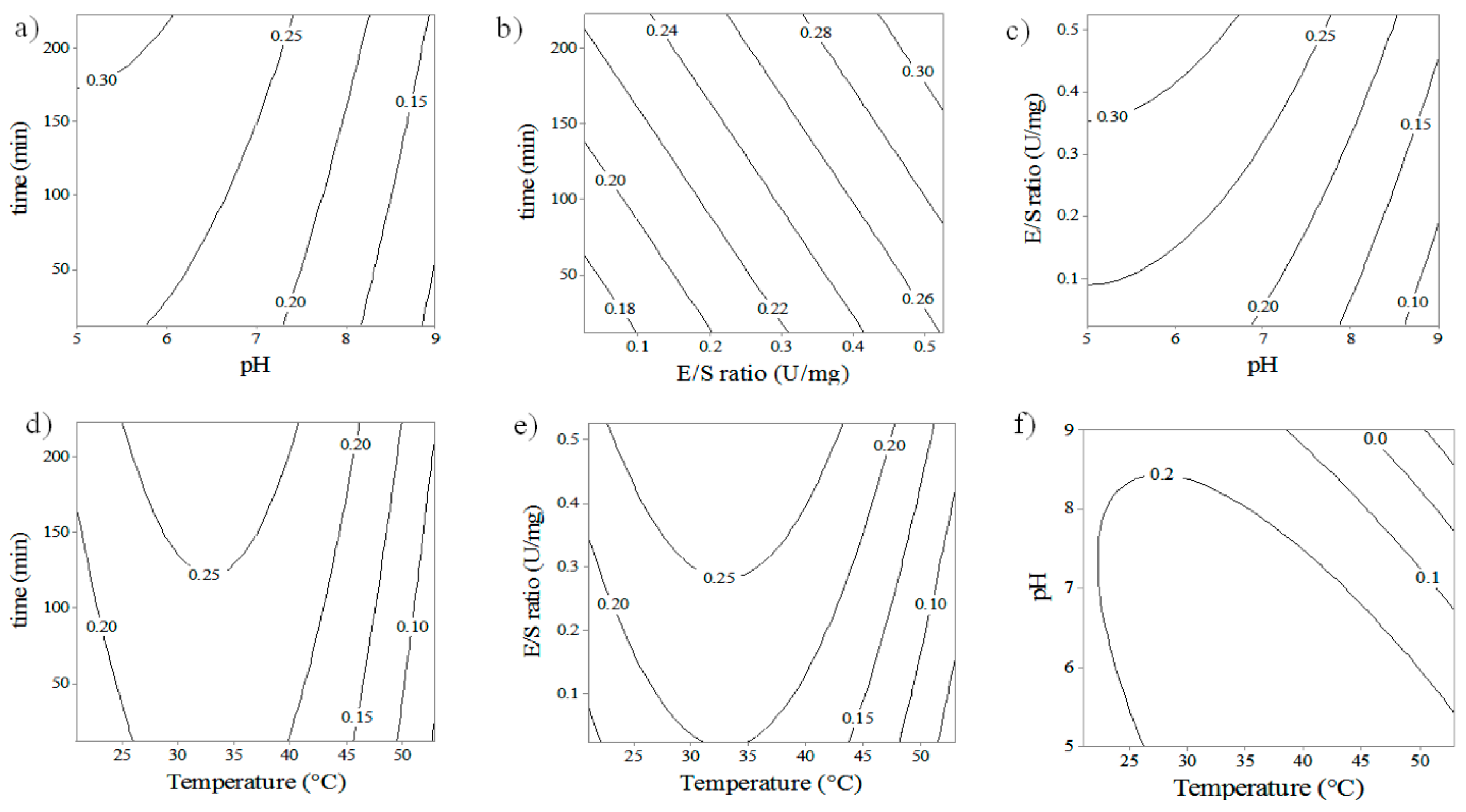
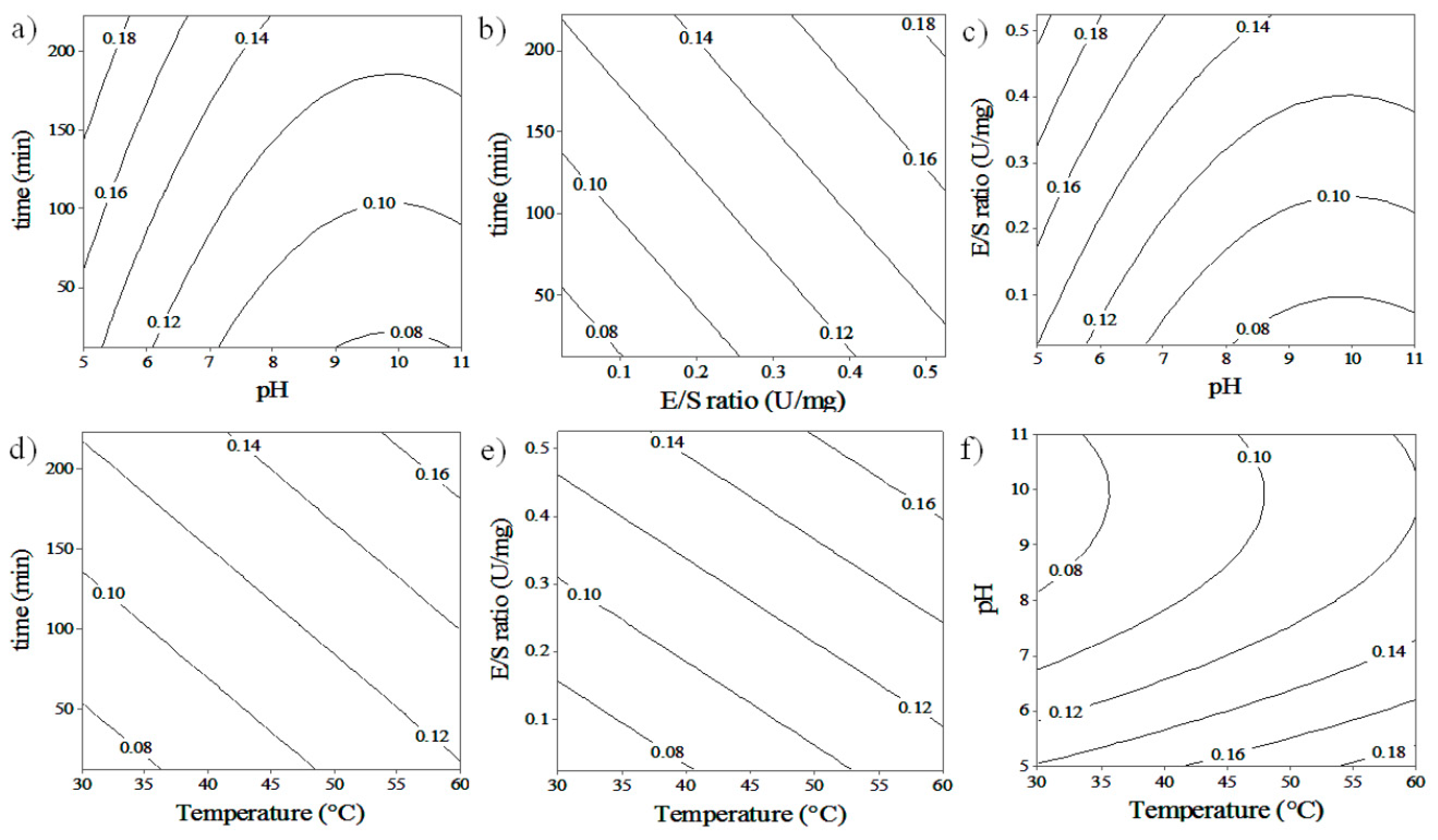
3.1. Response Surface Analysis
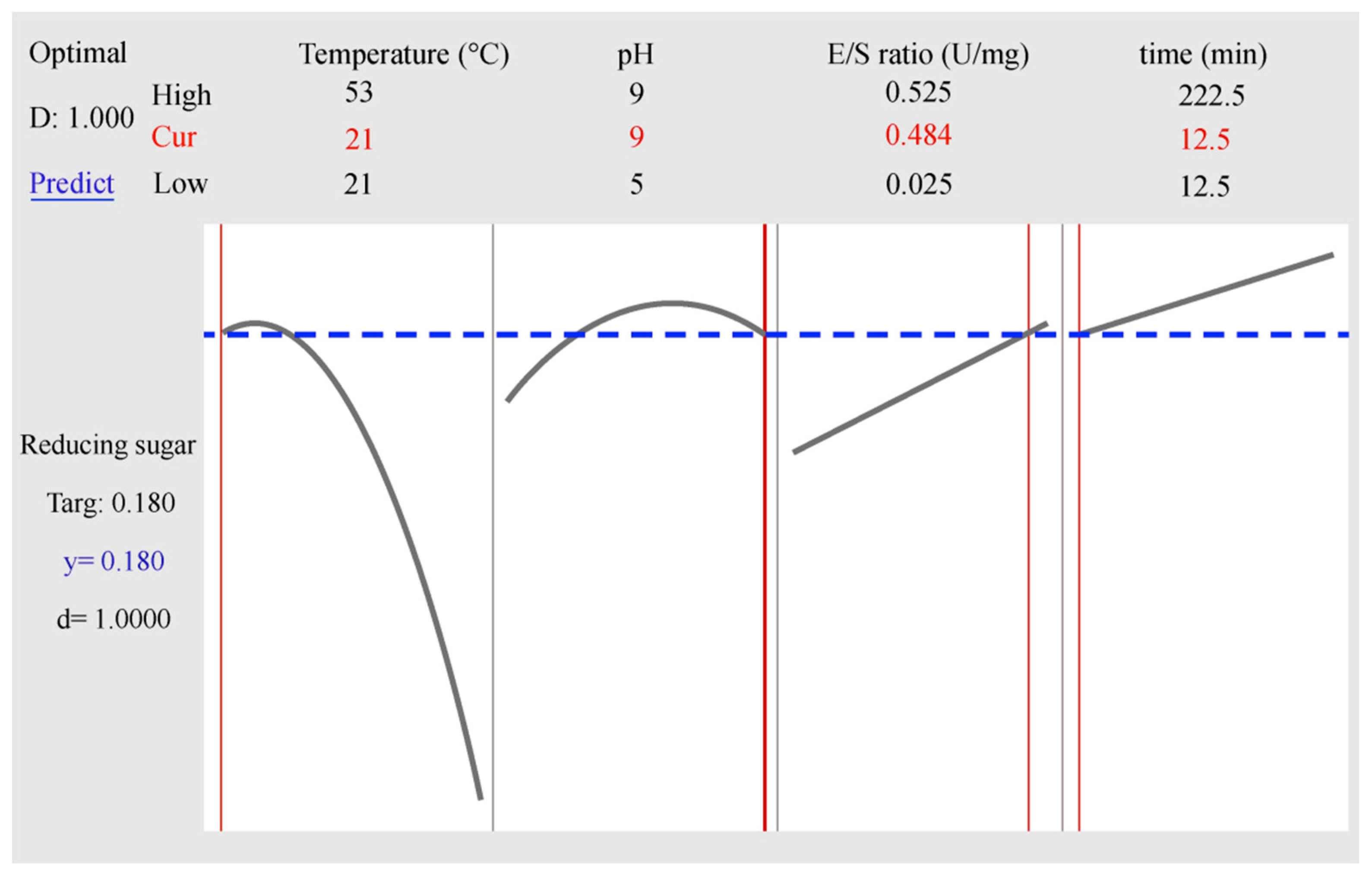
3.2. Confirmation and Validation Experiments
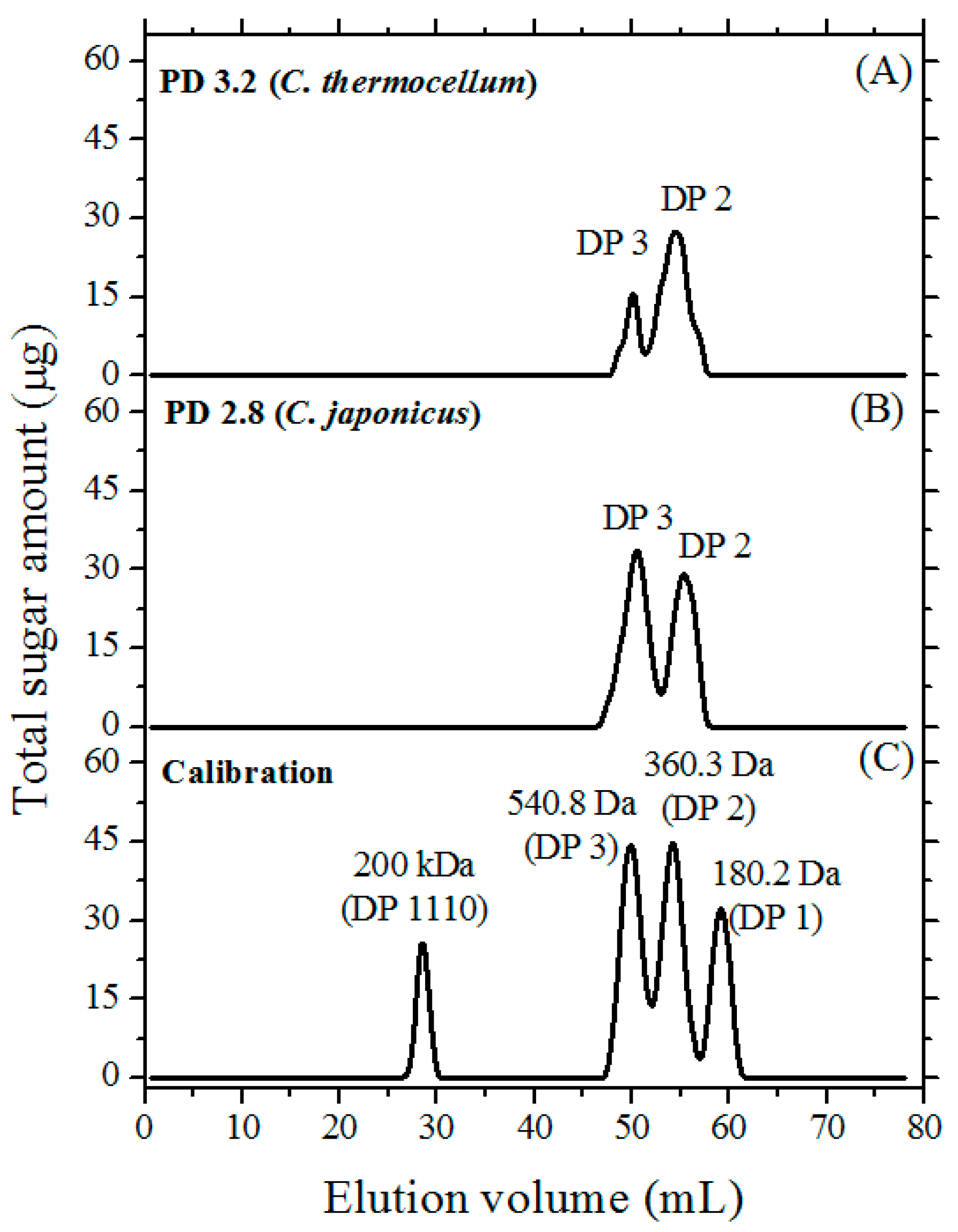
3.3. Size-Exclusion Chromatography (SEC)
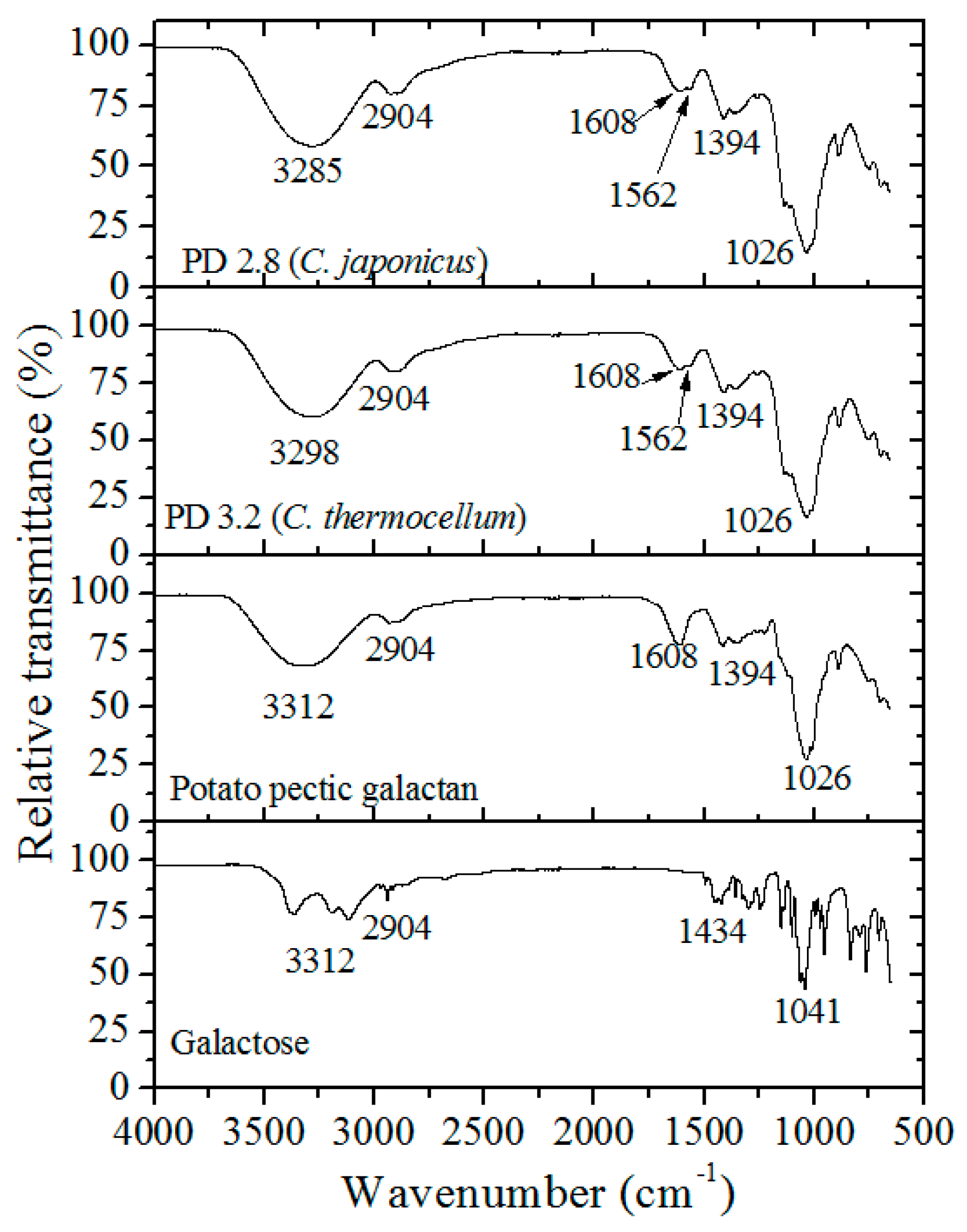
3.4. FT-IR and NMR Characterization
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2018, 67, 7–30. [Google Scholar] [CrossRef]
- Peer, D.; Karp, J.M.; Hong, S.; Farokhzad, O.C.; Margalit, R.; Langer, R. Nanocarriers as an emerging platform for cancer therapy. Nat. Nanotechnol. 2007, 2, 751–760. [Google Scholar] [CrossRef]
- Sinha, R.; Kim, G.J.; Nie, S. Nanotechnology in cancer therapeutics: Bioconjugated nanoparticles for drug delivery. Mol. Cancer Ther. 2006, 5, 1909–1917. [Google Scholar] [CrossRef]
- Morris, V.J.; Gromer, A.; Kirby, A.R.; Bongaerts, R.J.M.; Gunning, A.P. Using AFM and force spectroscopy to determine pectin structure and (bio) functionality. Food Hydrocoll. 2011, 25, 230–237. [Google Scholar] [CrossRef]
- Gunning, A.P.; Bongaerts, R.J.M.; Morris, V.J. Recognition of galactan components of pectin by galectin-3. FASEB J. 2009, 23, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, E.G.; Belshaw, N.J.; Waldron, K.W.; Morris, V.J. Pectin-an emerging new bioactive food polysaccharide. Trends Food Sci. Technol. 2012, 24, 64–73. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Functional oligosaccharides: Production, properties and applications. World J. Microbiol. Biotechnol. 2011, 27, 1119–1128. [Google Scholar] [CrossRef]
- Babbar, N.; Baldassarre, S.; Maesen, M.; Prandi, B.; Dejonghe, W.; Sforza, S.; Elst, K. Enzymatic production of pectic oligosaccharides from onion skins. Carbohydr. Polym. 2016, 146, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, Y.; Fukamori, T.; Raz, A. Galectin-3 and metastasis. Glycoconj. J. 2004, 19, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Subudhi, M.B.; Jain, A.; Jain, A.; Hurkat, P.; Shilpi, S.; Gulbake, A.; Jain, S.K. Eudragit S100 Coated Citrus Pectin Nanoparticles for Colon Targeting of 5-Fluorouracil. Materials 2015, 8, 832–849. [Google Scholar] [CrossRef] [Green Version]
- Tellez, S.R.; Garcia, F.L.; Vargas, B.A. Human Galectin-3 selective and high affinity inhibitors. Present state and future perspectives. Curr. Med. Chem. 2013, 20, 2979–2990. [Google Scholar] [CrossRef]
- Sundblad, V.; Croci, D.O.; Rabinovich, G.A. Regulated expression of galectin-3, a multifunctional glycan-binding protein, in haematopoietic and non-haematopoietic tissues. Histol. Histopathol. 2011, 26, 247–265. [Google Scholar] [CrossRef]
- Miller, M.C.; Zheng, Y.; Zhou, Y.; Tai, G.; Mayo, K.H. Galectin-3 binds selectively to the terminal, non-reducing end of β(1-4)-galactans, with overall affinity increasing with chain length. Glycobiology 2018, 29, 74–84. [Google Scholar] [CrossRef]
- Nakahara, S.; Oka, N.; Raz, A. On the role of galectin-3 in cancer apoptosis. Apoptosis 2005, 10, 267–275. [Google Scholar] [CrossRef]
- Arslan, H.; Pfaff, A.; Lu, Y.; Stepanek, P.; Müller, A.H.E. Stimuli-responsive spherical brushes based on D-galactopyranose and 2-(dimethylamino)-ethyl methacrylate. Macromol. Biosci. 2014, 14, 81–91. [Google Scholar] [CrossRef]
- Diaz, J.V.; Anthon, G.E.; Barrett, D.M. Nonenzymatic degradation of citrus pectin and pectate during prolonged heating: Effects of pH temperature, and degree of methyl esterification. J. Agric. Food Chem. 2007, 55, 5131–5136. [Google Scholar] [CrossRef]
- Michalak, M.; Thomassen, L.V.; Roytio, H.; Ouwehand, A.C.; Meyer, A.S.; Mikkelsen, J.D. Expression and characterization of an endo-1,4-β-galactanase from Emericella nidulans in pichia pastoris for enzymatic design of potentially prebiotic oligosaccharides from potato galactans. Enzyme Microb. Technol. 2012, 50, 121–129. [Google Scholar] [CrossRef]
- Tabachnikov, O.; Shoham, Y. Functional characterization of the galactan utilization system of Geobacillus stearothermophilus. FEBS J. 2013, 280, 950–964. [Google Scholar] [CrossRef]
- Yapo, B.M.; Lerouge, P.; Thibault, J.F.; Ralet, M.C. Pectins from citrus peel cell walls contain homogalacturonans homogeneous with respect to molar mass, rhamnogalacturonan I and rhamnogalacturonan II. Carbohydr. Polym. 2007, 69, 426–435. [Google Scholar] [CrossRef]
- Torpenholt, S.; Le, N.; Christensen, U.; Jahn, M.; Withers, S.; Rahbek, O.; Borchert, T.V.; Poulsen, J.C.; Lo, L. Activity of three β-1,4-galactanases on small chromogenic substrates. Carbohydr. Res. 2010, 346, 2028–2033. [Google Scholar] [CrossRef]
- Hinz, S.W.A.; Pastink, M.I.; van den Broek, L.A.M.; Vincken, J.P.; Voragen, A.G. Bifidobacterium longum endogalactanase liberates galactotriose from type I galactans. Appl. Environ. Microbiol. 2005, 71, 5501–5510. [Google Scholar] [CrossRef]
- Seo, S.; Karboune, S.; Yaylayan, V.; Hocine, L.L. Glycation of lysosyme with galactose, galactooligosaccharides and potato galactano through the Maillard reaction and optimization of the production of prebiotic glycoproteins. Process Biochem. 2012, 47, 297–304. [Google Scholar] [CrossRef]
- Courtin, C.M.; Van den Broeck, H.; Delcour, J.A. Determination of reducing end sugar residues in oligo- and polysaccharides by gas-liquid chromatography. J. Chromatogr. 2000, 866, 97–104. [Google Scholar] [CrossRef]
- Zhang, Y.H.P.; Lynd, L.R. Determination of the number-average degree of polymerization of cellodextrins and cellulose with application to enzymatic hydrolysis. Biomacromolecules 2005, 6, 1510–1515. [Google Scholar] [CrossRef]
- Brummer, Y.; Cui, S.W. Understanding carbohydrate analysis. In Food Carbohydrates: Chemistry, Physical Properties and Applications; Cui, S.W., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 74–104. [Google Scholar]
- Bach, E.; Schollmeyer, E. An ultraviolet-spectrophotometric method wih 2-cyanoacetamide for the determination of the enzymatic degradation of reducing polysaccharides. Anal. Biochem. 1992, 203, 335–339. [Google Scholar] [CrossRef]
- Cui, P.; Dou, T.Y.; Sun, Y.P.; Li, S.Y.; Feng, L.; Zou, W.; Wang, P.; Hao, D.C.; Ge, G.B.; Yang, L. Efficient enzymatic preparation of esculentoside B following condition optimization by response surface methodology. J. Mol. Catal. B Enzym. 2016, 130, 25–31. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, B.; Chen, B.; Jing, L.; Zhu, Z.; Kazemi, K. Modeling and optimization of Newfoundland shrimp waste hydrolysis for microbial growth using response surface methodology and artificial neural networks. Mar. Pollut. Bull. 2016, 109, 245–252. [Google Scholar] [CrossRef]
- Astray, G.; Gullon, B.; Labidi, J.; Gullon, P. Comparison between developed models using response surface methodology (RSM) and artificial neural networks (ANNs) with the purpose to optimize oligosaccharide mixtures production from sugar beet pulp. Ind. Crop. Prod. 2016, 92, 290–299. [Google Scholar] [CrossRef]
- González, D.I.; López, M.M.J.; Morales, G.; Segura, Y. Optimization of the synthesis of high galacto-oligosaccharides (GOS) from lactose with β-galactosidase from Kluyveromyces lactis. Int. Dairy J. 2016, 61, 211–219. [Google Scholar] [CrossRef]
- Qi, B.; Chen, X.; Shen, F.; Su, Y.; Wan, Y. Optimization of enzymatic hydrolysis of wheat straw pretreated by alkaline peroxide using response surface methodology. Ind. Eng. Chem. Res. 2009, 48, 7346–7353. [Google Scholar] [CrossRef]
- Zhong, K.; Wang, Q. Optimization of ultrasonic extraction of polysaccharides from dried longan pulp using response surface methodology. Carbohydr. Polym. 2010, 80, 19–25. [Google Scholar] [CrossRef]
- Das, B.; Roy, A.P.; Bhattachrjee, S.; Chakraborty, S.; Bhattacharjee, C. Lactose hydrolysis by β-galactosidase enzyme: Optimization using response surface methodology. Ecotoxicol. Environ. Saf. 2015, 121, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Somero, G.N. Temperature adaptation of enzymes: Biological optimization through structure-function compromises. Ann. Rev. Ecol. Syst. 1978, 9, 1–29. [Google Scholar] [CrossRef]
- Tavares, F.; Da Silva, E.A.; Pinzan, F.; Canevesi, R.S.; Milinsk, M.C.; Scheufele, F.B.; Borba, C.E. Hydrolysis of crambe oil by enzymatic catalysis: An evaluation of the operational conditions. Biocatal. Biotransform. 2018, 37, 1–14. [Google Scholar] [CrossRef]
- Daniel, R.M.; Danson, M.J. Temperature and catalytic activity of enzymes: A fresh understanding. FEBS Lett. 2013, 587, 2738–2743. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.M.C.; Dantas-Santos, N.; Gomes, D.L.; Costa, L.S.; Cordeiro, S.L.; Costa, M.S.S.P.; Silva, N.B.; Freitas, M.L.; Scortecci, K.C.; Leite, E.L.; et al. Biological activities of the sulfated polysaccharide from the vascular plant Halodule wrightii. Braz. J. Pharmacogn. 2012, 22, 94–101. [Google Scholar] [CrossRef]
- Guibaud, G.; Tixier, N.; Bouju, A.; Baudu, M. Relation between extracellular polymers’ composition and its ability to complex Cd, Cu and Pb. Chemosphere 2003, 52, 1701–1710. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Symbols | Levels | ||||
|---|---|---|---|---|---|---|
| C. japonicus/C. thermocellum | ||||||
| –2 a | –1 | 0 | 1 | 2 a | ||
| T b | X1 | 21/30 | 29/40 | 37/50 | 45/60 | 53/70 |
| pH | X2 | 5/3 | 6/5 | 7/7 | 8/9 | 9/11 |
| E/S c | X3 | 0.025 | 0.15 | 0.275 | 0.4 | 0.525 |
| t d | X4 | 12.5 | 65 | 117.5 | 170 | 222.5 |
| Run | Coded Level of Variable | Reducing Sugar | DP f | |||||
|---|---|---|---|---|---|---|---|---|
| T a | pH | E/S b | T c | C.j. d | C.t. e | C.j. d | C.t. e | |
| 1 | −1 | −1 | −1 | −1 | 0.2048 | 0.1427 | 2.7 | 4.1 |
| 2 | 2 | 0 | 0 | 0 | 0.0383 | 0.1421 | 14.2 | 4.1 |
| 3 | 0 | 0 | 0 | 0 | 0.2131 | 0.1166 | 2.6 | 5.0 |
| 4 | 0 | 0 | 0 | 0 | 0.2670 | 0.1205 | 2.0 | 4.9 |
| 5 | 1 | −1 | −1 | −1 | 0.2509 | 0.1599 | 2.2 | 3.7 |
| 6 | 1 | 1 | −1 | −1 | 0.0522 | 0.0883 | 10.4 | 6.6 |
| 7 | 1 | −1 | −1 | 1 | 0.2359 | 0.1998 | 2.3 | 2.9 |
| 8 | −1 | −1 | 1 | −1 | 0.2392 | 0.146 | 2.3 | 4.0 |
| 9 | −1 | −1 | 1 | 1 | 0.2864 | 0.1743 | 1.9 | 3.4 |
| 10 | 1 | 1 | 1 | 1 | 0.1371 | 0.1349 | 4.0 | 4.3 |
| 11 | 0 | 2 | 0 | 0 | 0.1726 | 0.1132 | 3.2 | 5.2 |
| 12 | 0 | 0 | 2 | 0 | 0.2792 | 0.1682 | 2.0 | 3.5 |
| 13–16 | 0 | 0 | 0 | 0 | 0.2409 | 0.1427 | 2.3 | 4.1 |
| 17 | −1 | 1 | −1 | −1 | 0.1349 | 0.0644 | 4.0 | 9.1 |
| 18 | −1 | 1 | 1 | 1 | 0.2243 | 0.1277 | 2.4 | 4.6 |
| 19 | −1 | −1 | −1 | 1 | 0.2548 | 0.0816 | 2.1 | 7.2 |
| 20 | 0 | 0 | −2 | 0 | 0.1837 | 0.0788 | 3.0 | 7.4 |
| 21 | 0 | 0 | 0 | 2 | 0.2537 | 0.1482 | 2.1 | 4.0 |
| 22 | 0 | 0 | 0 | −2 | 0.1993 | 0.101 | 2.7 | 5.8 |
| 23 | 1 | 1 | 1 | −1 | 0.1282 | 0.1299 | 4.2 | 4.5 |
| 24 | 0 | 0 | 0 | 0 | 0.2720 | 0.1610 | 2.0 | 3.6 |
| 25 | −1 | 1 | −1 | 1 | 0.1965 | 0.086 | 2.8 | 6.8 |
| 26 | 0 | −2 | 0 | 0 | 0.2415 | 0.0955 | 2.3 | 6.1 |
| 27 | 1 | 1 | −1 | 1 | 0.0866 | 0.1138 | 6.3 | 5.2 |
| 28 | −2 | 0 | 0 | 0 | 0.2342 | 0.1044 | 2.3 | 5.6 |
| 29 | 1 | −1 | 1 | −1 | 0.2737 | 0.1843 | 2.0 | 3.2 |
| 30 | 1 | −1 | 1 | 1 | 0.2964 | 0.232 | 1.8 | 2.5 |
| 31 | −1 | 1 | 1 | −1 | 0.2065 | 0.0822 | 2.6 | 7.1 |
| Source | Adj. Sum of Squares | Degree of Freedom | Adj. Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Model | 0.116164 | 7 | 0.016595 | 19.01 | 0.000 |
| X1-Temp | 0.019171 | 1 | 0.019171 | 21.97 | 0.000 |
| X2-pH | 0.042805 | 1 | 0.042805 | 49.05 | 0.000 |
| X3-E/S | 0.013356 | 1 | 0.013356 | 15.30 | 0.001 |
| X4-time | 0.004715 | 1 | 0.004715 | 5.40 | 0.029 |
| X12 | 0.022792 | 1 | 0.022792 | 26.12 | 0.000 |
| X22 | 0.003066 | 1 | 0.003066 | 3.51 | 0.074 |
| X1×2 | 0.011536 | 1 | 0.011536 | 13.22 | 0.001 |
| Error | 0.020073 | 23 | 0.000873 | ||
| Lack of fit | 0.017176 | 17 | 0.001010 | 2.09 | 0.184 |
| Pure error | 0.002897 | 6 | 0.000483 | ||
| Total | 0.136237 | 30 |
| Source | Adj. Sum of Squares | Degree of Freedom | Adj. Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Model | 0.035948 | 5 | 0.007190 | 42.32 | 0.000 |
| X1-Temp | 0.005213 | 1 | 0.005213 | 30.69 | 0.000 |
| X2-pH | 0.019945 | 1 | 0.019945 | 117.41 | 0.000 |
| X3-E/S | 0.006435 | 1 | 0.006435 | 37.88 | 0.000 |
| X4-time | 0.003940 | 1 | 0.003940 | 23.19 | 0.000 |
| X22 | 0.002132 | 1 | 0.002132 | 12.55 | 0.002 |
| Error | 0.003907 | 23 | 0.000170 | ||
| Lack of fit | 0.002279 | 17 | 0.000134 | 0.49 | 0.882 |
| Pure error | 0.001629 | 6 | 0.000271 | ||
| Total | 0.039855 | 28 |
| Endo-1,4-β-galactanase | d * | T (°C) | pH | E/S (U/mg) | Time (min) | RSpred | TS (μmol) | RS (μmol) | DPcal |
|---|---|---|---|---|---|---|---|---|---|
| Cellvibrio japonicus | 1 | 51.8 | 5 | 0.508 | 77.5 | 0.27 | 0.478 | 0.254 ± 0.016 | 1.9 |
| 1 | 21 | 9 | 0.484 | 12.5 | 0.18 | 0.475 | 0.173 ± 0.007 | 2.8 | |
| 1 | 21 | 5 | 0.462 | 12.5 | 0.13 | 0.579 | 0.137 ± 0.007 | 4.2 | |
| Clostridium thermocellum | 1 | 60 | 5 | 0.525 | 148 | 0.23 | 0.576 | 0.274 ± 0.044 | 2.1 |
| 1 | 59.7 | 5 | 0.506 | 12.5 | 0.19 | 0.597 | 0.184 ± 0.006 | 3.2 | |
| 1 | 34.5 | 11 | 0.525 | 222.5 | 0.14 | 0.617 | 0.151 ± 0.011 | 4.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Ayón, M.A.; Licea-Claveríe, Á.; Valdez-Torres, J.B.; Picos-Corrales, L.A.; Vélez-de la Rocha, R.; Contreras-Esquivel, J.C.; Labavitch, J.M.; Sañudo-Barajas, J.A. Enzyme-Catalyzed Production of Potato Galactan-Oligosaccharides and Its Optimization by Response Surface Methodology. Materials 2019, 12, 1465. https://0-doi-org.brum.beds.ac.uk/10.3390/ma12091465
González-Ayón MA, Licea-Claveríe Á, Valdez-Torres JB, Picos-Corrales LA, Vélez-de la Rocha R, Contreras-Esquivel JC, Labavitch JM, Sañudo-Barajas JA. Enzyme-Catalyzed Production of Potato Galactan-Oligosaccharides and Its Optimization by Response Surface Methodology. Materials. 2019; 12(9):1465. https://0-doi-org.brum.beds.ac.uk/10.3390/ma12091465
Chicago/Turabian StyleGonzález-Ayón, Mirian Angelene, Ángel Licea-Claveríe, José Benigno Valdez-Torres, Lorenzo A. Picos-Corrales, Rosabel Vélez-de la Rocha, Juan Carlos Contreras-Esquivel, John M. Labavitch, and Josefa Adriana Sañudo-Barajas. 2019. "Enzyme-Catalyzed Production of Potato Galactan-Oligosaccharides and Its Optimization by Response Surface Methodology" Materials 12, no. 9: 1465. https://0-doi-org.brum.beds.ac.uk/10.3390/ma12091465