The Photoreceptor Components FaWC1 and FaWC2 of Fusarium asiaticum Cooperatively Regulate Light Responses but Play Independent Roles in Virulence Expression


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Sequence Analysis of Fawc1 and Fawc2 of F. Asiaticum
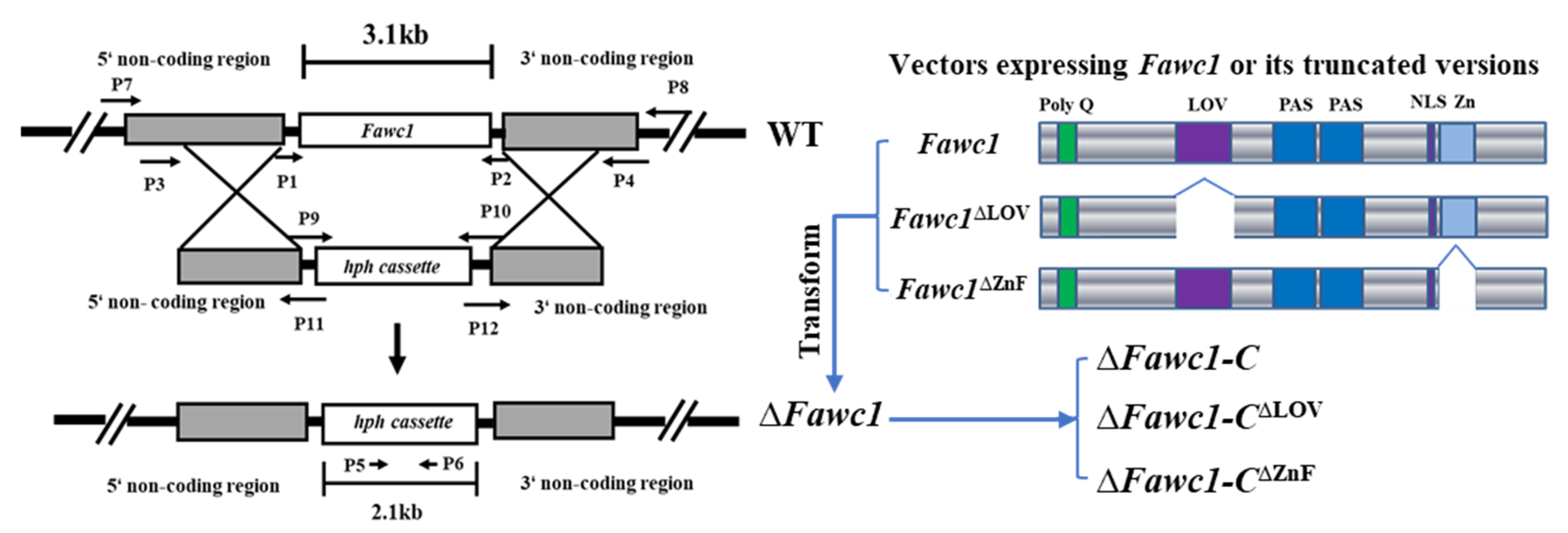
2.3. Generation of Mutants and Complementation Strains in F. asiaticum
2.4. Extraction of RNA and Quantitative RT-PCR Analysis
2.5. Growth and Development Phenotyping
2.6. Carotenoid Measurement Assay
2.7. UV Sensitivity Assay
2.8. Virulence Assay
2.9. Statistical Analysis
3. Results
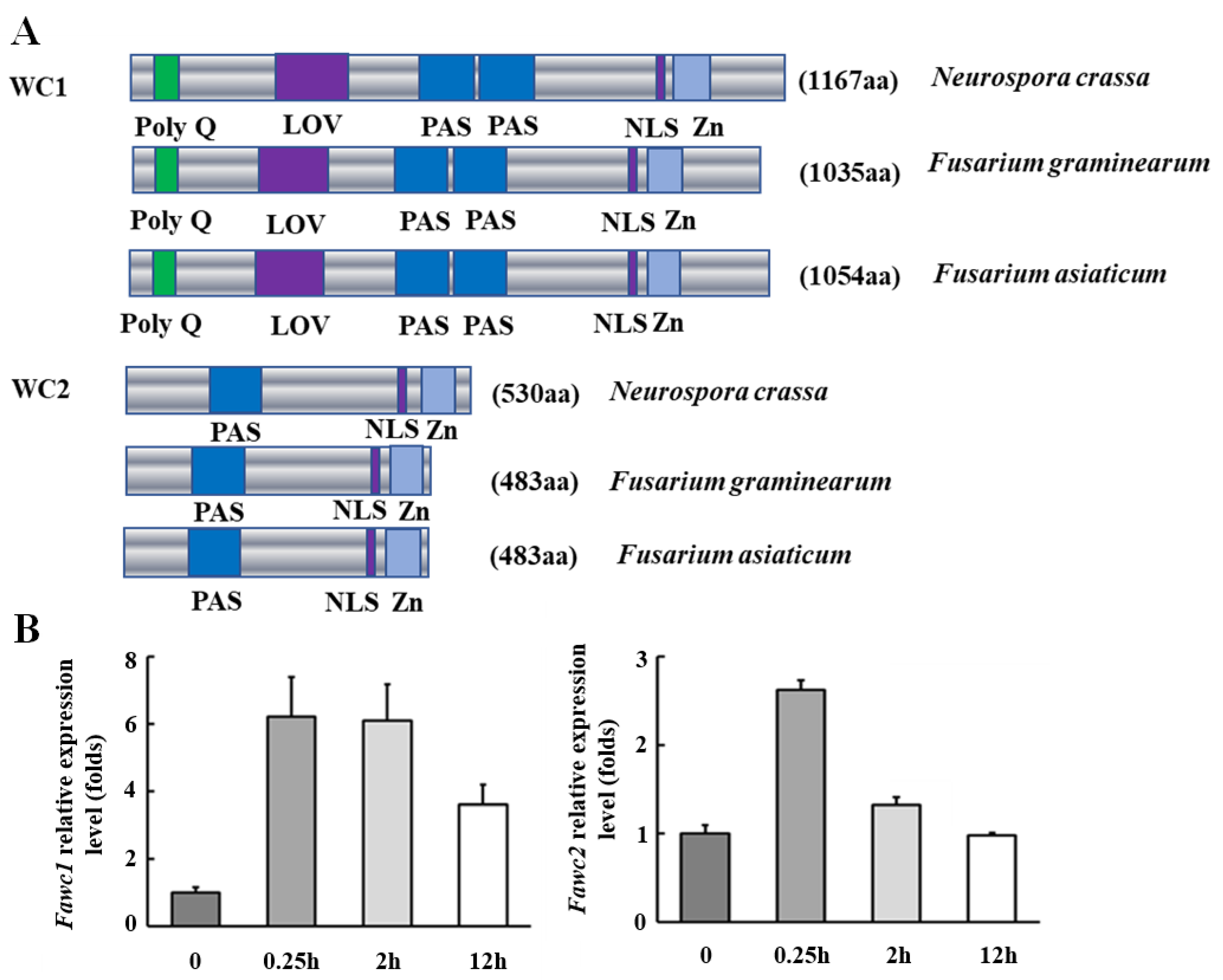
3.1. The Orthologs of WC-1 and WC-2 in F. asiaticum
3.2. Generation and Characterization of the ∆Fawc1 and ∆Fawc2 Mutants
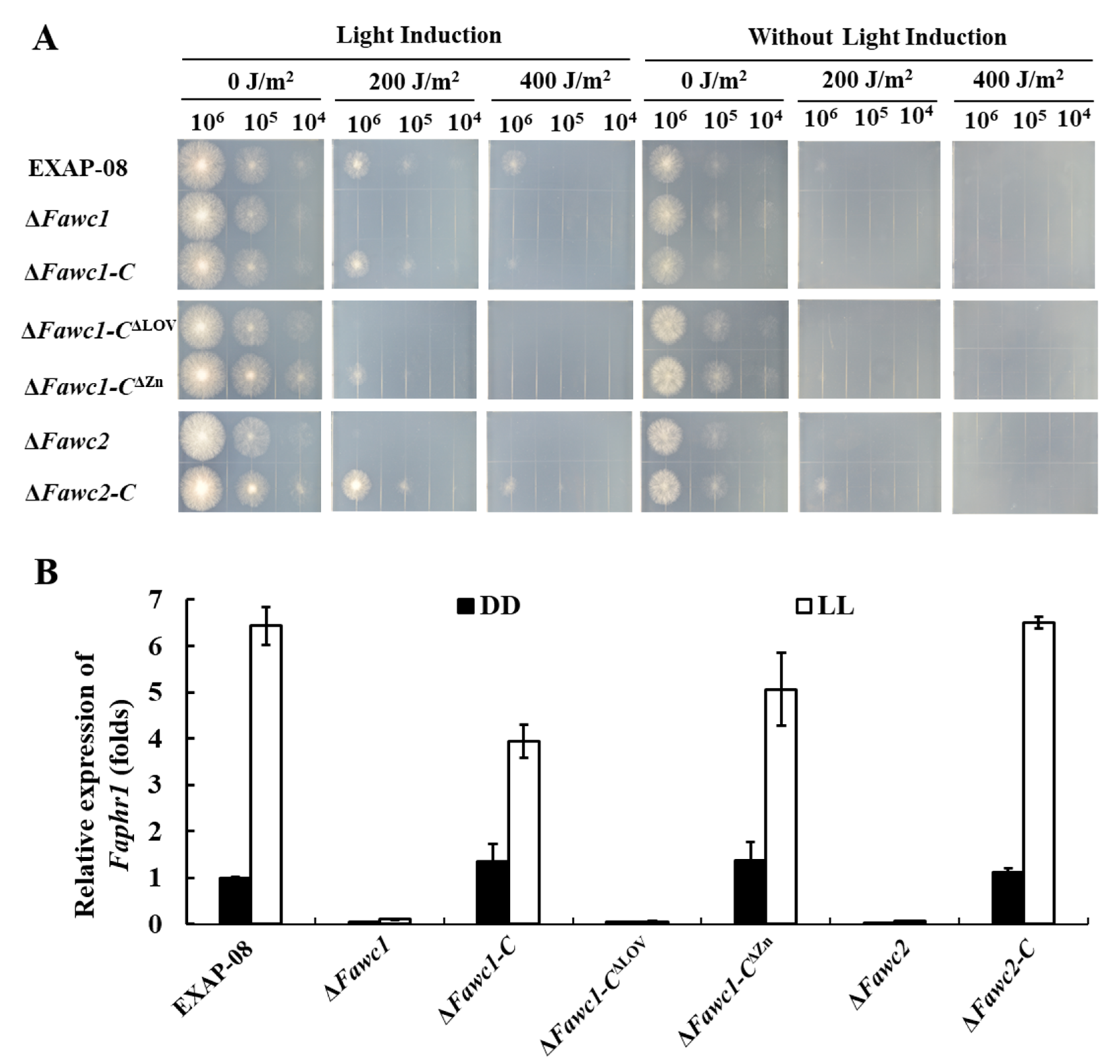
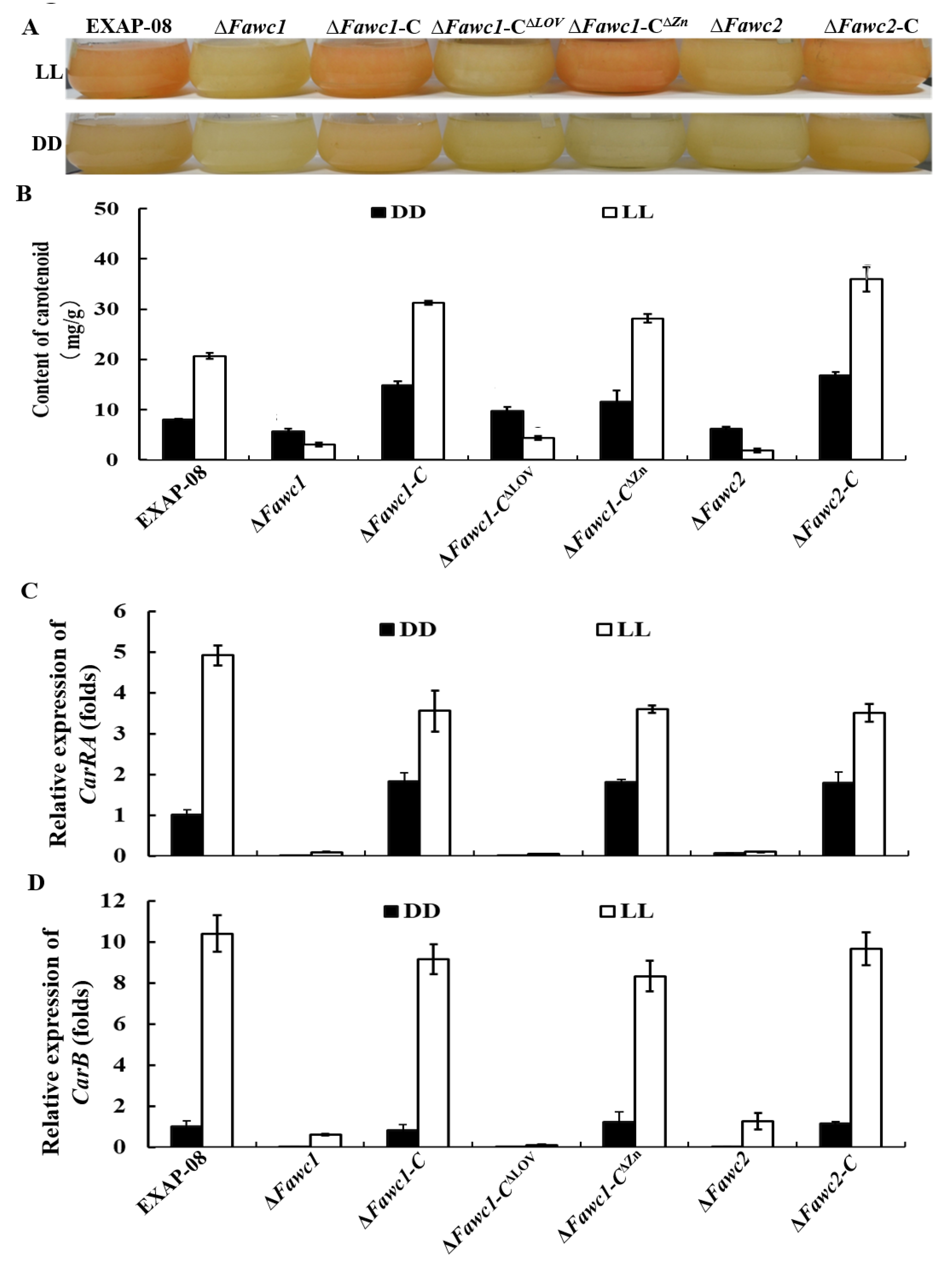
3.3. The Marker Responses to Light Signal Are Mediated by WCC and Dependent on LOV but not ZnF Domain of FaWC1 in F. asiaticum
3.4. Perithecia Maturation and Ascospore Development of F. asiaticum Are Regulated by WCC Photoreceptor
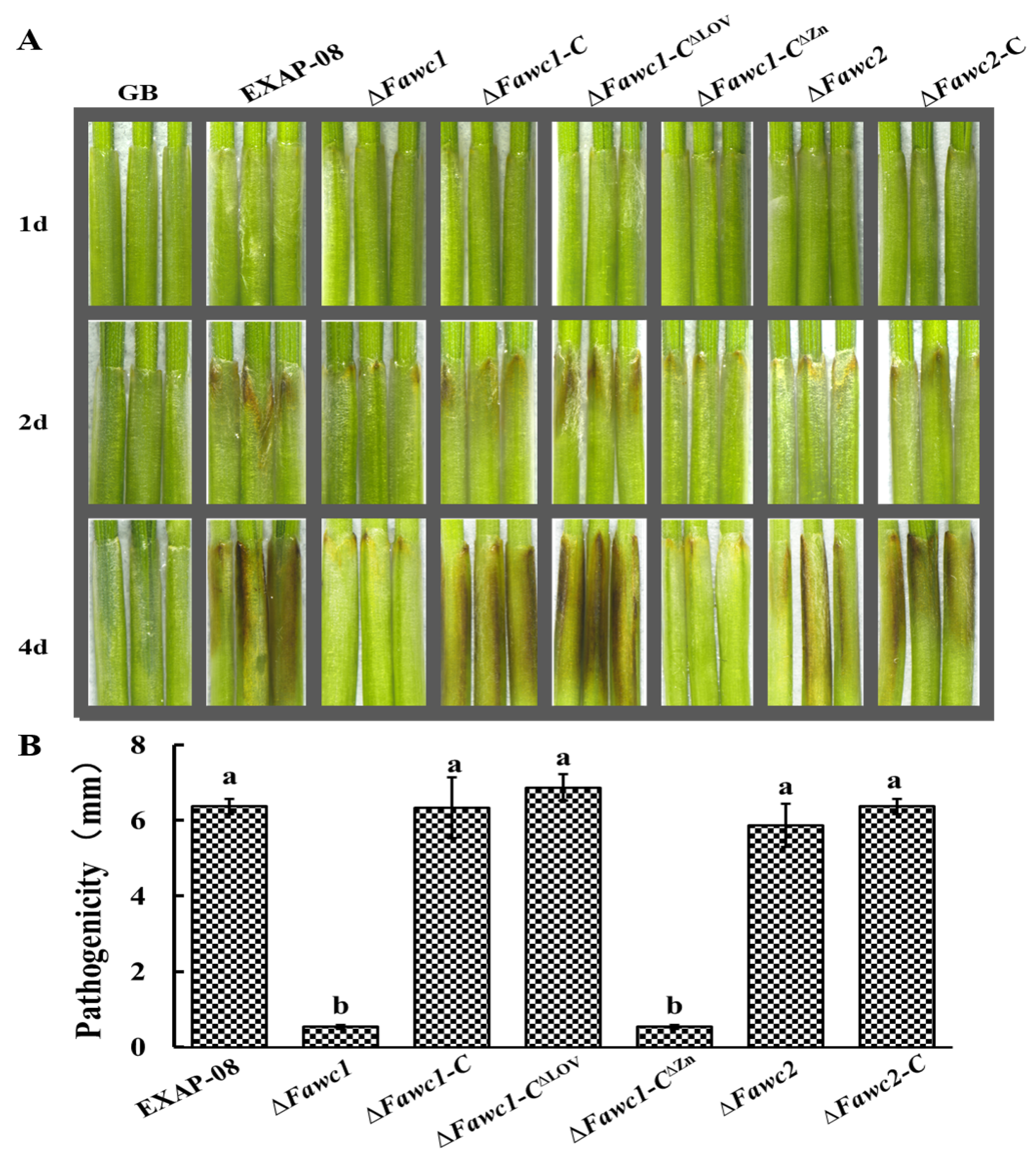
3.5. FaWC1 and FaWC2 Play Different Roles in Regulating Virulence Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cuomo, C.A.; Güldener, U.; Xu, J.R.; Trail, F.; Turgeon, B.G.; Pietro, A.D.; Walton, J.D.; Ma, L.J.; Baker, S.E.; Rep, M.; et al. The Fusarium graminearum genome reveals a link between localized polymorphism and pathogen specialization. Science 2007, 317, 1400–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Kistler, H.C.; Tacke, B.K.; Casper, H.H. Gene genealogies reveal global phylogeographic structure and reproductive isolation among lineages of Fusarium graminearum, the fungus causing wheat scab. PNAS 2000, 97, 7905–7910. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Ward, T.J.; Geiser, D.M.; Kistler, H.C.; Aoki, T. Genealogical concordance between the mating type locus and seven other nuclear genes supports formal recognition of nine phylogenetically distinct species within the Fusarium graminearum clade. Fungal Genet. Biol. 2004, 41, 600–623. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Ward, T.J.; Aberra, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjørnstad, Å.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef]
- Starkey, D.E.; Ward, T.J.; Aoki, T.; Gale, L.R.; Kistler, H.C.; Geiser, D.M.; Suga, H.; Tóth, B.; Varga, J.; O’Donnell, K. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity. Fungal Genet. Biol. 2007, 44, 1191–1204. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Gagkaeva, T.; Ward, T.J.; Aoki, T.; Kistler, H.C.; O’Donnell, K. A novel Asian clade within the Fusarium graminearum species complex includes a newly discovered cereal head blight pathogen from the Russian Far East. Mycologia 2009, 101, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Sarver, B.A.J.; Ward, T.J.; Gale, L.R.; Broz, K.; Kistler, H.C.; Aoki, T.; Nicholson, P.; Carter, J.; O’Donnell, K. Novel Fusarium head blight pathogens from Nepal and Louisiana revealed by multilocus genealogical concordance. Fungal Genet. Biol. 2011, 48, 1096–1107. [Google Scholar] [CrossRef]
- Zhang, H.; Van der Lee, T.; Waalwijk, C.; Chen, W.; Xu, J.; Xu, J.; Zhang, Y.; Feng, J. Population analysis of the Fusarium graminearum species complex from wheat in China show a shift to more aggressive isolates. PLoS ONE 2012, 7, e31722. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ma, H.; Zhou, Y.; Xing, J.; Chen, J.; Yu, G.; Sun, X.; Wang, L. Identification and genetic division of Fusarium graminearum and Fusarium asiaticum by species-specific SCAR markers. J. Phytopathol. 2014, 162, 81–88. [Google Scholar] [CrossRef]
- Zhu, P.; Wu, L.; Liu, L.; Huang, L.; Wang, Y.; Tang, W.; Wu, L. Fusarium asiaticum: An Emerging Pathogen Jeopardizing Postharvest Asparagus Spears. J. Phytopathol. 2013, 161, 696–703. [Google Scholar] [CrossRef]
- Steiner, B.; Kurz, H.; Lemmens, M.; Buerstmayr, H. Differential gene expression of related wheat lines with contrasting levels of head blight resistance after Fusarium graminearum inoculation. Theor. Appl. Genet. 2009, 118, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, N.; Pumphrey, M.O.; Liu, S.X.; Zhang, X.F.; Tiwari1, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A.; et al. Wheat Fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight. Nat. Genetics. 2016, 48, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; Van der Does, H.C.; Borkovich, B.A.; Coleman, J.J.; Daboussi, M.J.; Pietro, A.D.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.; et al. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Jia, L.J.; Zhang, Y.; Jiang, G.; Li, X.; Zhang, D.; Tang, W.H. In planta stage-specific fungal gene profiling elucidates the molecular strategies of Fusarium graminearum growing inside wheat coleoptiles. Plant Cell. 2012, 24, 5159–5176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.D.; Saparno, A.; Blackwell, B.; Anoop, V.; Gleddie, S.; Tinker, N.A.; Harriset, L.J. Proteomic analyses of Fusarium graminearum grown under mycotoxin-inducing conditions. Proteomics 2008, 8, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Z.; Van der Lee, T.; Chen, W.Q.; Xu, J.; Xu, J.S.; Yang, L.; Yu, D.; Waalwijk, C.; Feng, J. Population genetic analyses of Fusarium asiaticum populations from barley suggest a recent shift favoring 3ADON producers in southern China. Phytopathology 2010, 100, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Bahn, Y.S.; Xue, C.; Idnurm, A.; Rutherford, J.C.; Heitman, J.; Cardenas, M.E. Sensing the environment: Lessons from fungi. Nat. Rev. Microbiol. 2007, 5, 57–69. [Google Scholar] [CrossRef]
- Tisch, D.; Schmoll, M. Light regulation of metabolic pathways in fungi. Appl. Microbiol. Biotechnol. 2010, 85, 1259–1277. [Google Scholar] [CrossRef] [Green Version]
- Heintzen, C.; Loros, J.J.; Dunlap, J.C. The PAS protein VIVID defines a clock-associated feedback loop that represses light input, modulates gating, and regulates clock resetting. Cell 2001, 104, 453–464. [Google Scholar] [CrossRef]
- Idnurm, A.; Crosson, S. The photobiology of microbial pathogenesis. PLoS Pathog. 2009, 5, e1000470. [Google Scholar] [CrossRef] [Green Version]
- Ballario, P.; Vittorioso, P.; Magrelli, A.; Talora, C.; Cabibbo, A.; Macino, G. White collar-1, a central regulator of blue light responses in Neurospora, is a zinc finger protein. EMBO J. 1996, 15, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Harding, R.W.; Melles, S. Genetic Analysis of Phototropism of Neurospora crassa Perithecial Beaks Using White Collar and Albino Mutants. Plant Physiol. 1983, 72, 996–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauter, F.R.; Russo, V.E. Blue light induction of conidiation-specific genes in Neurospora crassa. Nucleic Acids. Res. 1991, 19, 6883–6886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmoll, M.; Tian, C.; Sun, J.; Tisch, D.; Glass, N.L. Unravelling the molecular basis for light modulated cellulase gene expression - the role of photoreceptors in Neurospora crassa. BMC Genomics. 2012, 13, 127. [Google Scholar] [CrossRef] [Green Version]
- Tisch, D.; Schmoll, M. Targets of light signalling in Trichoderma reesei. BMC Genomics. 2013, 14, 657. [Google Scholar] [CrossRef] [Green Version]
- Scholthof, K.B.G. The disease triangle: Pathogens, the environment and society. Nat. Rev. Microbiol. 2007, 5, 152–156. [Google Scholar] [CrossRef]
- Kim, H.; Ridenour, J.B.; Dunkle, L.D.; Bluhm, B.H. Regulation of stomatal tropism and infection by light in Cercospora zeae-maydis: Evidence for coordinated host/pathogen responses to photoperiod? PLoS Pathog. 2011, 7, e1002113. [Google Scholar] [CrossRef]
- Kim, S.; Singh, P.; Park, K.; Park, S.; Friedman, A.; Zheng, T.; Lee, Y.H.; Lee, K. Genetic and molecular characterization of a blue light photoreceptor MGWC-1 in Magnaporth oryzae. Fungal Genet. Biol. 2011, 48, 400–407. [Google Scholar] [CrossRef]
- Canessa, P.; Schumacher, J.; Hevia, M.A.; Tudzynski, P.; Larronodo, L.F. Assessing the effects of light on differentiation and virulence of the plant pathogen Botrytis cinerea: Characterization of the White Collar Complex. PLoS ONE 2013, 8, e84223. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, H.K.; Lee, S.; Yun, S.H. The white collar complex is involved in sexual development of Fusarium graminearum. PLoS ONE 2015, 10, e0120293. [Google Scholar] [CrossRef]
- Xu, Y.B.; Li, H.P.; Zhang, J.B.; Song, B.; Chen, F.F.; Duan, X.J.; Xu, H.Q.; Liao, Y.C. Disruption of the chitin synthase gene CHS1 from Fusarium asiaticum results in an altered structure of cell walls and reduced virulence. Fungal Genet. Biol. 2010, 47, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. Fusarium laboratory workshops-A recent history. Mycotoxin Res. 2006, 22, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; Maclean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crosthwaite, S.K.; Dunlap, J.C.; Loros, J.J. Neurospora wc-1 and wc-2: Transcription, photoresponses, and the origins of circadian rhythmicity. Science 1997, 276, 763–769. [Google Scholar] [CrossRef]
- Thoma, F. Light and dark in chromatin repair: Repair of UV- induced DNA lesions by photolyase and nucleotide excision repair. EMBO J. 1991, 18, 6585–6598. [Google Scholar] [CrossRef] [Green Version]
- Corrochano, L.M. Light in the Fungal World: From Photoreception to Gene Transcription and Beyond. Annu. Rev. Genet. 2019, 53, 1–22. [Google Scholar] [CrossRef]
- Avalos, J.; Pardo-Medina, J.; Parra-Rivero, O.; Ruger-Herreros, M.; Rodríguez-Ortiz, R.; Hornero-Méndez, D.; Limón, M.C. Carotenoid Biosynthesis in Fusarium. J. Fungi. 2017, 3, 39. [Google Scholar] [CrossRef] [Green Version]
- Prado, M.M.; Prado-Cabrero, A.; Fernández-Martín, R.; Avalos, J. A gene of the opsin family in the carotenoid gene cluster of Fusarium fujikuroi. Curr. Genet. 2004, 46, 47–58. [Google Scholar] [CrossRef]
- Tschanz, A.T.; Horst, R.K.; Nelson, P.E. The Effect of Environment on Sexual Reproduction of Gibberella Zeae. Mycologia. 1976, 68, 327–340. [Google Scholar] [CrossRef]
- Kim, H.; Son, H.; Lee, Y.W. Effects of light on secondary metabolism and fungal development of Fusarium graminearum. J. Appl. Microbiol. 2014, 116, 380–389. [Google Scholar] [CrossRef]
- Chen, C.H.; Ringelberg, C.S.; Gross, R.H.; Dunlap, J.C.; Loros, J.J. Genome-wide analysis of light-inducible responses reveals hierarchical light signalling in Neurospora. EMBO J. 2009, 28, 1029–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Yang, F.; Smith, K.M.; Peterson, M.; Dekhang, M.; Zhang, Y.; Zucker, J.; Bredeweg, E.L.; Mallappa, C.; Zhou, X.; et al. Genome-wide characterization of light-regulated genes in Neurospora crassa. Genes 2014, 4, 1731–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Lee, T.; Zhang, H.; van Diepeningenc, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. 2015, 32, 453–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backhouse, D. Global distribution of Fusarium graminearum, F. asiaticum and F. boothii from wheat in relation to climate. Eur. J. Plant Pathol. 2014, 139, 161–173. [Google Scholar] [CrossRef]
- Fuller, K.K.; Loros, J.J.; Dunlap, J.C. Fungal photobiology: Visible light as a signal for stress, space and time. Curr. Genet. 2015, 61, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Fischer, R. Light sensing and responses in fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef]
- Zhu, P.; Idnurm, A. The contribution of the White Collar complex to Cryptococcus neoformans virulence is independent of its light-sensing capabilities. Fungal Genet. Biol. 2018, 121, 56–64. [Google Scholar] [CrossRef]
- Berrocal-Tito, G.M.; Esquivel-Naranjo, E.U.; Horwitz, B.A.; Herrera-Estrella, A. Trichoderma atroviride PHR1, a fungal photolyase responsible for DNA repair, autoregulates its own photoinduction. Eukaryot Cell. 2007, 6, 1682–1692. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Roldán, M.C.; Garre, V.; Guarro, J.; Mariné, M.; Roncero, M.I.G. Role of the white collar 1 photoreceptor in carotenogenesis, UV resistance, hydrophobicity, and virulence of Fusarium oxysporum. Eukaryot Cell. 2008, 7, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Idnurm, A. The Uve1 endonuclease is regulated by the white collar complex to protect cryptococcus neoformans from UV damage. PLoS Genet. 2013, 9, e1003769. [Google Scholar] [CrossRef] [Green Version]
- Bayram, Ö.; Braus, G.H.; Fischer, R.; Rodriguez-Romero, J. Spotlight on Aspergillus nidulans photosensory systems. Fungal Genet. Biol. 2010, 47, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Pruβ, S.; Fetzner, R.; Seither, K.; Herr, A.; Pfeiffer, E.; Metzler, M.; Lawrence, C.B.; Fischer, R. Role of the Alternaria alternata blue-light receptor LreA (white-collar 1) in spore formation and secondary metabolism. Appl. Environ. Microbiol. 2014, 80, 2582–2591. [Google Scholar] [CrossRef] [Green Version]
- Bayram, Ö.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA Complex Coordinates Light Signal with Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Purschwitz, J.; Müller, S.; Kastner, C.; Schöser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and physical interaction of blue- and red-light sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trail, F.; Xu, H.; Loranger, R.; Gadoury, D. Physiological and environmental aspects of ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum). Mycologia. 2002, 94, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhou, X.; Loros, J.J.; Dunlap, J.C. Alternative Use of DNA Binding Domains by the Neurospora White Collar Complex Dictates Circadian Regulation and Light Responses. Mol. Cell Biol. 2015, 36, 781–793. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Genotype | Reference |
|---|---|---|
| EXAP-08 | Wild type | [10] |
| ∆Fawc1 | Knockout mutant, Fawc1::Hyg | This study |
| ∆Fawc2 | Knockout mutant, Fawc2::Hyg | This study |
| ∆Fawc1-C | Fawc-1 complemented transformant of ΔFawc-1 | This study |
| ∆Fawc2-C | Fawc-2 complemented transformant of ΔFawc-2 | This study |
| ∆Fawc1-C∆LOV | LOV domain deletion mutant | This study |
| ∆Fawc1-C∆ZnF | ZnF domain deletion mutant | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Zhu, P.; Lu, Z.; Qu, Y.; Huang, L.; Zheng, N.; Wang, Y.; Nie, H.; Jiang, Y.; Xu, L. The Photoreceptor Components FaWC1 and FaWC2 of Fusarium asiaticum Cooperatively Regulate Light Responses but Play Independent Roles in Virulence Expression. Microorganisms 2020, 8, 365. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030365
Tang Y, Zhu P, Lu Z, Qu Y, Huang L, Zheng N, Wang Y, Nie H, Jiang Y, Xu L. The Photoreceptor Components FaWC1 and FaWC2 of Fusarium asiaticum Cooperatively Regulate Light Responses but Play Independent Roles in Virulence Expression. Microorganisms. 2020; 8(3):365. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030365
Chicago/Turabian StyleTang, Ying, Pinkuan Zhu, Zhengyu Lu, Yao Qu, Li Huang, Ni Zheng, Yiwen Wang, Haozhen Nie, Yina Jiang, and Ling Xu. 2020. "The Photoreceptor Components FaWC1 and FaWC2 of Fusarium asiaticum Cooperatively Regulate Light Responses but Play Independent Roles in Virulence Expression" Microorganisms 8, no. 3: 365. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030365