Human mecC-Carrying MRSA: Clinical Implications and Risk Factors

 ,
,
Abstract
:1. Introduction
2. Detection of mecC-MRSA Isolates in Humans
2.1. Human Studies Related to mecC-MRSA
2.2. mecC-MRSA Human Case Reports
3. Risk Factors for mecC-MRSA Infection
3.1. Contact with Animals
3.2. mecC-MRSA Carriage in Humans
3.3. Patient Age
3.4. Underlying Chronic Disease
4. Characterization of mecC-MRSA Human Isolates
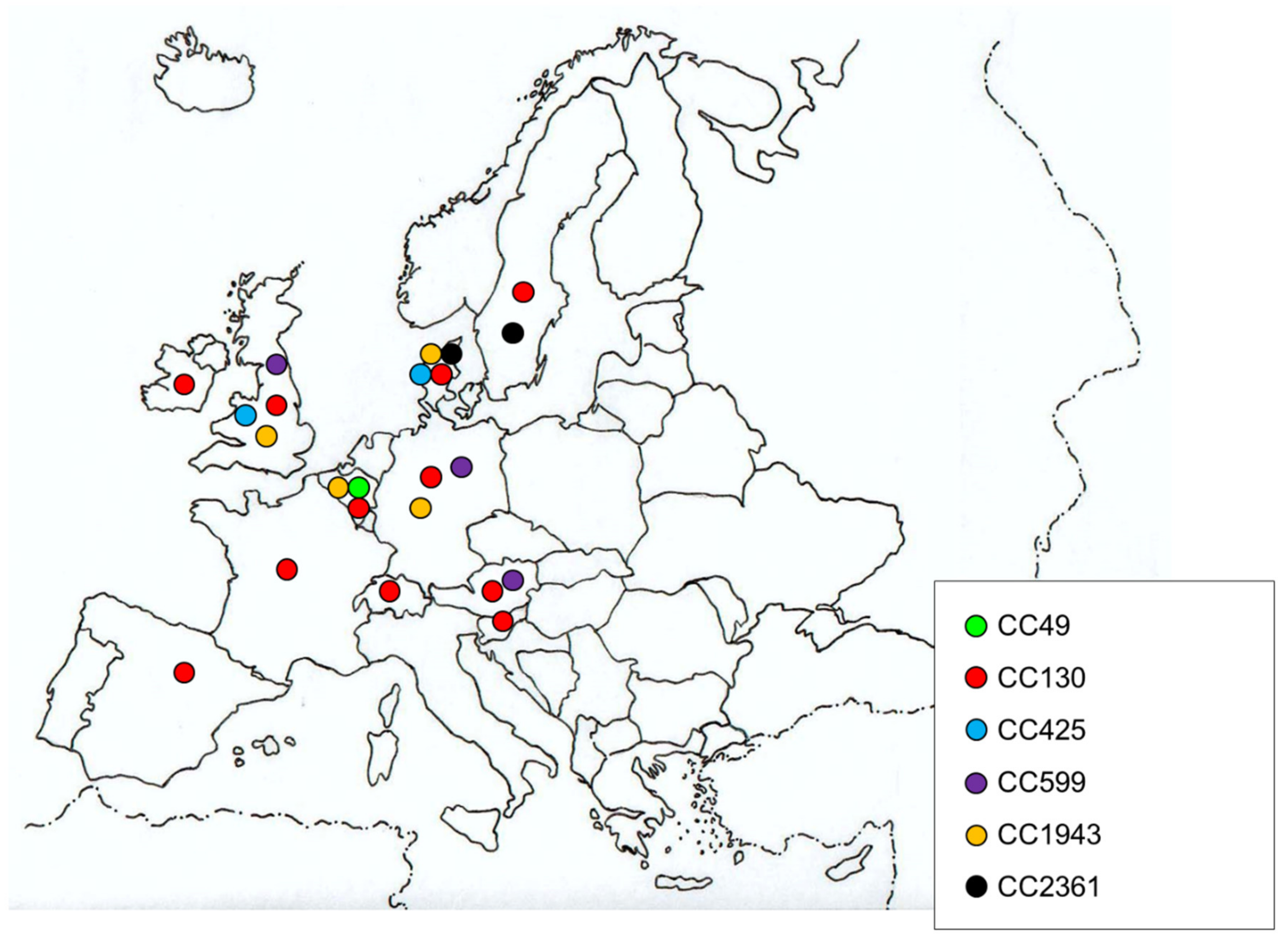
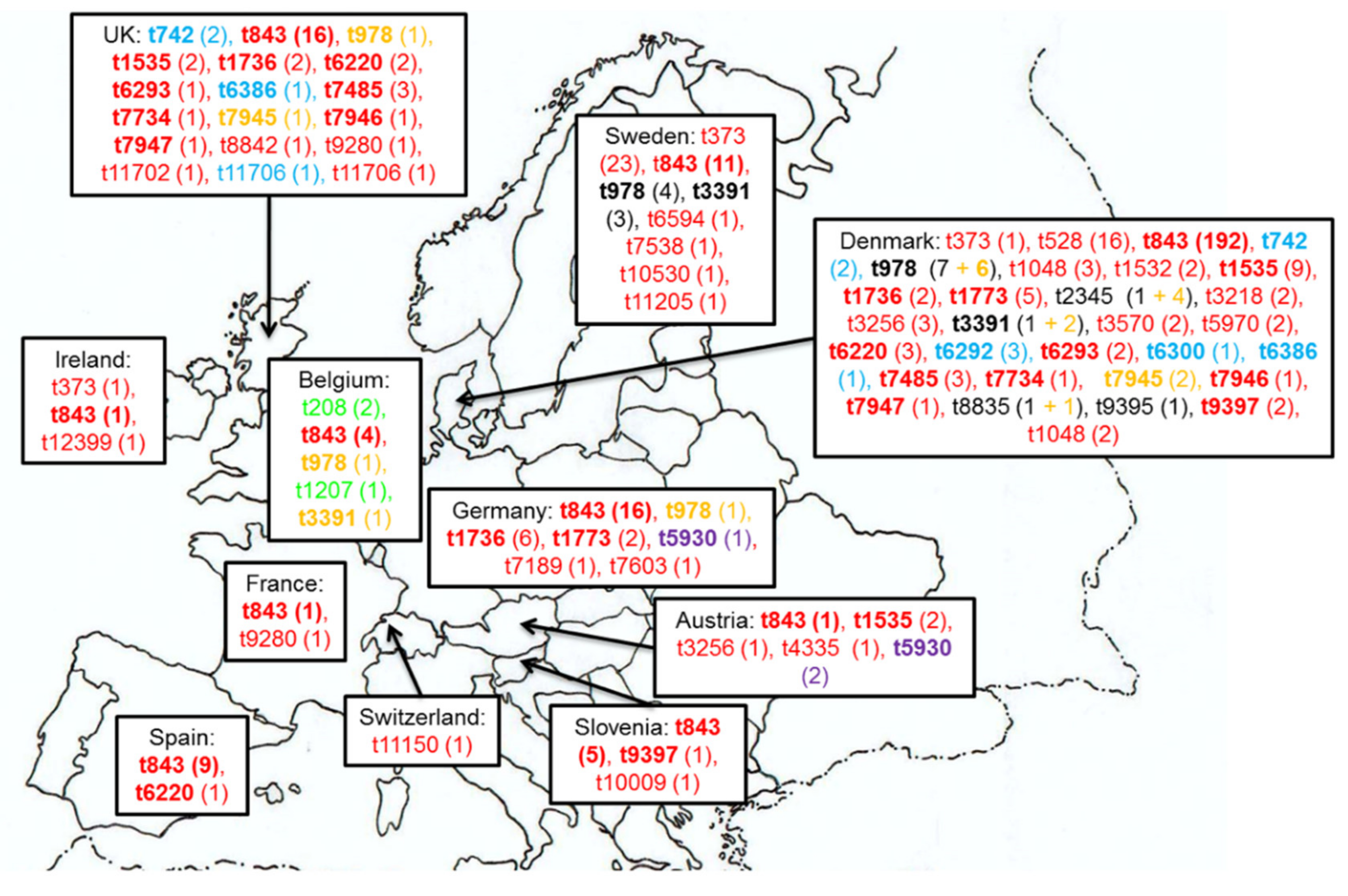
4.1. Clonal Lineages of mecC-MRSA of Human Origin
4.2. Treatment and Antimicrobial Resistance Profile of mecC-MRSA Human Isolates
4.3. Virulence of mecC-MRSA Human Isolates
5. mecC-MRSA Problem: What is its Origin? Is It an Emerging Problem?
6. Implications in Veterinary and Food Safety
7. Conclusions and Future Problems Associated with mecC
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cheng, M.; Antignac, A.; Kim, C.; Tomasz, A. Comparative study of the susceptibilities of major epidemic clones of methicillin-resistant Staphylococcus aureus to oxacillin and to the new broad-spectrum cephalosporin ceftobiprole. Antimicrob. Agents Chemother. 2008, 52, 2709–2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacqueline, C.; Caillon, J.; Mabecque, V.L.; Miègeville, A.F.; Ge, Y.; Biek, D.; Batard, E.; Potel, G. In vivo activity of a novel anti-methicillin-resistant Staphylococcus aureus cephalosporin, ceftaroline, against vancomycin-susceptible and -resistant Enterococcus faecalis strains in a rabbit endocarditis model: A comparative study with linezolid and vancomycin. Antimicrob. Agents Chemother. 2009, 53, 5300–5302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mediavilla, J.R.; Chen, L.; Mathema, B.; Kreiswirth, B.N. Global epidemiology of communityassociated methicillin resistant Staphylococcus aureus (CA-MRSA). Curr. Opin. Microbiol. 2012, 15, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.Y.; Lee, J.; Choi, E.H.; Lee, H.J. Changes in molecular epidemiology of communityassociated and health care-associated methicillin-resistant Staphylococcus aureus in Korean children. Diagn. Microbiol. Infect. Dis. 2012, 74, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Herron-Olson, L.; Fitzgerald, J.R.; Musser, J.M.; Kapur, V. Molecular correlates of host specialization in Staphylococcus aureus. PLoS ONE 2007, 2, e1120. [Google Scholar] [CrossRef] [PubMed]
- Aspiroz, C.; Lozano, C.; Vindel, A.; Lasarte, J.J.; Zarazaga, M.; Torres, C. Skin lesion caused by ST398 and ST1 MRSA, Spain. Emerg. Infect. Dis. 2010, 16, 157–159. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; Aspiroz, C.; Ezpeleta, A.I.; Gómez-Sanz, E.; Zarazaga, M.; Torres, C. Empyema caused by MRSA ST398 with atypical resistance profile, Spain. Emerg Infect. Dis. 2011, 17, 138–140. [Google Scholar] [CrossRef]
- Zarazaga, M.; Gómez, P.; Ceballos, S.; Torres, C. Molecular epidemiology of Staphylococcus aureus lineages in the animal-human interface. In Staphylococcus Aureus; Fetsch, A., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 189–214. [Google Scholar]
- Benito, D.; Lozano, C.; Rezusta, A.; Ferrer, I.; Vasquez, M.A.; Ceballos, S.; Zarazaga, M.; Revillo, M.J.; Torres, C. Characterization of tetracycline and methicillin resistant Staphylococcus aureus Strains in a Spanish hospital: Is livestock-contact a risk factor in infections caused by MRSA CC398? Int. J. Med. Microbiol. 2014, 304, 1226–1232. [Google Scholar] [CrossRef]
- Larsen, J.; Petersen, A.; Sørum, M.; Stegger, M.; van Alphen, L.; Valentiner-Branth, P.; Knudsen, L.K.; Larsen, L.S.; Feingold, B.; Price, L.B.; et al. Meticillin-resistant Staphylococcus aureus CC398 is an increasing cause of disease in people with no livestock contact in Denmark, 1999 to 2011. Euro Surveill. 2015, 20. [Google Scholar] [CrossRef] [Green Version]
- Lekkerkerk, W.; van Wamel, J.B.; Snijders, S.B.; Willems, R.J.; van Duijkeren, E.; Broens, E.M.; Wagenaar, J.A.; Lindsay, J.A.; Vos, M.C. What is the origin of livestock-associated methicillin-resistant Staphylococcus aureus Clonal Complex 398 isolates from humans without livestock contact? an epidemiological and genetic analysis. J. Clin. Microbiol. 2015, 53, 1836–1841. [Google Scholar] [CrossRef] [Green Version]
- Feltrin, F.; Alba, P.; Kraushaar, B.; Ianzano, A.; Argudín, M.A.; Di Matteo, P.; Porrero, M.C.; Aarestrup, F.M.; Butaye, P.; Franco, A.; et al. A Livestock-associated, multidrug-resistant, methicillin-resistant Staphylococcus aureus Clonal Complex 97 lineage spreading in dairy cattle and pigs in Italy. Appl. Environ. Microbiol. 2015, 82, 816–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowder, B.V.; Guinane, C.M.; Ben Zakour, N.L.; Weinert, L.A.; Conway-Morris, A.; Cartwright, R.A.; Simpson, A.J.; Rambaut, A.; Nübel, U.; Fitzgerald, J.R. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 19545–19550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Gavier-Widén, D.; Hotzel, H.; Peters, M.; Guenther, S.; Lazaris, A.; Loncaric, I.; Müller, E.; Reissig, A.; Ruppelt-Lorz, A.; et al. Diversity of Staphylococcus aureus isolates in European wildlife. PLoS ONE. 2016, 11, e0168433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Wang, X.; Fan, Y.; Peng, Y.; Li, L.; Li, S.; Huang, J.; Yao, Z.; Chen, S. Genotypic and phenotypic markers of livestock-associated methicillin-resistant Staphylococcus aureus CC9 in humans. Appl. Environ. Microbiol. 2016, 82, 3892–3899. [Google Scholar] [CrossRef] [Green Version]
- García-Álvarez, L.; Holden, M.T.G.; Lindsay, H.; Webb, C.R.; Brown, D.F.J.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Shore, A.C.; Deasy, E.C.; Slickers, P.; Brennan, G.; O’Connell, B.; Monecke, S.; Ehricht, R.; Coleman, D.C. Detection of staphylococcal cassette chromosome mec type XI carrying highly divergent mecA, mecI, mecR1, blaZ, and ccr genes in human clinical isolates of clonal complex 130 methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3765–3773. [Google Scholar] [CrossRef] [Green Version]
- Harrison, E.M.; Paterson, G.K.; Holden, M.T.G.; Larsen, J.; Stegger, M.; Larsen, A.R.; Petersen, A.; Skov, R.L.; Christensen, J.M.; Zeuthen, A.B.; et al. Whole genome sequencing identifies zoonotic transmission of MRSA isolates with the novel mecA homologue mecC. EMBO Mol. Med. 2013, 5, 509–515. [Google Scholar] [CrossRef]
- Becker, K.; Ballhausen, B.; Köck, R.; Kriegeskorte, A. Methicillin resistance in Staphylococcus isolates: The “mec alphabet” with specific consideration of mecC, a mec homolog associated with zoonotic S. aureus lineages. Int. J. Med. Microbiol. 2014, 304, 794–804. [Google Scholar] [CrossRef]
- Becker, K.; van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kaspar, U.; Peters, G. Plasmid-Encoded Transferable mecB-Mediated Methicillin Resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Aires-de-Sousa, M. Methicillin-resistant Staphylococcus aureus among animals: Current overview. Clin. Microbiol. Infect. 2017, 23, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; González-Barrio, D.; Benito, D.; García, J.T.; Viñuela, J.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. Detection of methicillin-resistant Staphylococcus aureus (MRSA) carrying the mecC gene in wild small mammals in Spain. J. Antimicrobial Chemother. 2014, 69, 2061–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ripa, L.; Alcalá, L.; Simón, C.; Gómez, P.; Mama, O.M.; Rezusta, A.; Zarazaga, M.; Torres, C. Diversity of Staphylococcus aureus clones in wild mammals in Aragon, Spain, with detection of MRSA ST130-mecC in wild rabbits. J. Appl. Microbiol. 2019, 127, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.; Stegger, M.; Heltberg, O.; Christensen, J.; Zeuthen, A.; Knudsen, L.K.; Urth, T.; Sorum, M.; Schouls, L.; Larsen, J.; et al. Epidemiology of methicillin-resistant Staphylococcus aureus carrying the novel mecC gene in Denmark corroborates a zoonotic reservoir with transmission to humans. Clin. Microbiol Infect. 2013, 19, E16–E22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegger, M.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecA(LGA251). Clin. Microbiol. Infect. 2012, 18, 395–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnevey, P.M.; Shore, A.C.; Brennan, G.I.; Sullivan, D.J.; Ehricht, R.; Monecke, S.; Coleman, D.C. Extensive genetic diversity identified among sporadic methicillin-resistant Staphylococcus aureus isolates recovered in Irish hospitals between 2000 and 2012. Antimicrob. Agents Chemother. 2014, 58, 1907–1917. [Google Scholar] [CrossRef] [Green Version]
- Monecke, S.; Jatzwauk, L.; Müller, E.; Nitschke, H.; Pfohl, K.; Slickers, P.; Reissig, A.; Ruppelt-Lorz, A.; Ehricht, R. Diversity of SCCmec elements in Staphylococcus aureus as observed in South-Eastern Germany. PLoS ONE. 2016, 11, e0162654. [Google Scholar] [CrossRef]
- Zarfel, G.; Luxner, J.; Folli, B.; Leitner, E.; Feierl, G.; Kittinger, C.; Grisold, A. Increase of genetic diversity and clonal replacement of epidemic methicillin-resistant Staphylococcus aureus strains in South-East Austria. FEMS Microbiol Lett. 2016, 363, fnw137. [Google Scholar] [CrossRef] [Green Version]
- Deplano, A.; Vandendriessche, S.; Nonhoff, C.; Denis, O. Genetic diversity among methicillin-resistant Staphylococcus aureus isolates carrying the mecC gene in Belgium. J. Antimicrob. Chemother. 2014, 69, 1457–1460. [Google Scholar] [CrossRef] [Green Version]
- Kriegeskorte, A.; Ballhausen, B.; Idelevich, E.A.; Köck, R.; Friedrich, A.W.; Karch, H.; Peters, G.; Becker, K. Human MRSA isolates with novel genetic homolog, Germany. Emerg. Infect. Dis. 2012, 18, 1016–1018. [Google Scholar] [CrossRef]
- Sabat, A.J.; Koksal, M.; Akkerboom, V.; Monecke, S.; Kriegeskorte, A.; Hendrix, R.; Ehricht, R.; Köck, R.; Becker, K.; Friedrich, A.W. Detection of new methicillin-resistant Staphylococcus aureus strains that carry a novel genetic homologue and important virulence determinants. J. Clin. Microbiol. 2012, 50, 3374–3377. [Google Scholar] [CrossRef] [Green Version]
- Schaumburg, F.; Köck, R.; Mellmann, A.; Richter, L.; Hasenberg, F.; Kriegeskorte, A.; Friedrich, A.W.; Gatermann, S.; Peters, G.; von Eiff, C.; et al. Population dynamics among methicillin-resistant Staphylococcus aureus isolates in Germany during a 6-year period. J. Clin. Microbiol. 2012, 50, 3186–3192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuny, C.; Layer, F.; Strommenger, B.; Witte, W. Rare occurrence of methicillin-resistant Staphylococcus aureus CC130 with a novel mecA homologue in humans in Germany. PLoS ONE. 2011, 6, e24360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartwright, E.J.P.; Paterson, G.K.; Raven, K.E.; Harrison, E.M.; Gouliouris, T.; Kearns, A.; Pichon, B.; Edwards, G.; Skov, R.L.; Larsen, A.; et al. Use of Vitek 2 antimicrobial susceptibility profile to identify mecC in methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2013, 51, 2732–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersdorf, S.; Herma, M.; Rosenblatt, M.; Layer, F.; Henrich, B. A Novel Staphylococcal Cassette Chromosome mec Type XI primer for detection of mecC-harboring methicillin-resistant Staphylococcus aureus directly from screening specimens. J. Clin. Microbiol. 2015, 53, 3938–3941. [Google Scholar] [CrossRef] [Green Version]
- Dermota, U.; Zdovc, I.; Strumbelj, I.; Grmek-Kosnik, I.; Ribic, H.; Rupnik, M.; Golob, M.; Zajc, U.; Bes, M.; Laurent, F.; et al. Detection of methicillin-resistant Staphylococcus aureus carrying the mecC gene in, human samples in Slovenia. Epidemiol. Infect. 2015, 143, 1105–1108. [Google Scholar] [CrossRef] [Green Version]
- García-Garrote, F.; Cercenado, E.; Marín, M.; Bal, M.; Trincado, P.; Corredoira, J.; Ballesteros, C.; Pita, J.; Alonso, P.; Vindel, A. Methicillin-resistant Staphylococcus aureus carrying the mecC gene: Emergence in Spain and report of a fatal case of bacteraemia. J. Antimicrob. Chemother. 2014, 69, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Kerschner, H.; Harrison, E.M.; Hartl, R.; Holmes, M.A.; Apfalter, P. First report of mecC MRSA in human samples from Austria: Molecular characteristics and clinical data. New Microbes New Infect. 2014, 3, 4–9. [Google Scholar] [CrossRef] [Green Version]
- Petersen, A.; Medina, A.; Rhod Larsen, A. Ability of the GENSPEED(®) MRSA test kit to detect the novel mecA homologue mecC in Staphylococcus aureus. APMIS. 2015, 123, 478–481. [Google Scholar] [CrossRef]
- Paterson, G.K.; Morgan, F.J.E.; Harrison, E.M.; Cartwright, E.J.P.; Török, M.E.; Zadoks, R.N.; Parkhill, J.; Peacock, S.J.; Holmes, M.A. Prevalence and characterization of human mecC methicillin-resistant Staphylococcus aureus isolates in England. J. Antimicrob. Chemother. 2014, 69, 907–910. [Google Scholar] [CrossRef] [Green Version]
- Harrison, E.M.; Coll, F.; Toleman, M.S.; Blane, B.; Brown, N.B.; Török, M.E.; Parkhill, J.; Peacock, S.J. Genomic surveillance reveals low prevalence of livestock-associated methicillin-resistant Staphylococcus aureus in the East of England. Sci. Rep. 2017, 7, 7406. [Google Scholar] [CrossRef] [Green Version]
- Paterson, G.K. Low prevalence of livestock-associated methicillin-resistant Staphylococcus aureus clonal complex 398 and mecC MRSA among human isolates in North-West England. J. Appl. Microbiol. 2020, 128, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Ciesielczuk, H.; Xenophontos, M.; Lambourne, J. Methicillin-resistant Staphylococcus aureus harboring mecC still eludes us in East London, United Kingdom. J. Clin. Microbiol. 2019, 57, e00020-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriegeskorte, A.; Idelevich, E.A.; Schlattmann, A.; Layer, F.; Strommenger, B.; Denis, O.; Paterson, G.K.; Holmes, M.A.; Werner, G.; Becker, K. Comparison of different phenotypic approaches to screen and detect mecC-harboring methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2017, 56, e00826-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basset, P.; Prod’hom, G.; Senn, L.; Greub, G.; Blanc, D.S. Very low prevalence of meticillin-resistant Staphylococcus aureus carrying the mecC Gene in Western Switzerland. J. Hosp. Infect. 2013, 83, 257–259. [Google Scholar] [CrossRef] [Green Version]
- Dekker, D.; Wolters, M.; Mertens, E.; Boahen, K.G.; Krumkamp, R.; Eibach, D.; Schwarz, N.G.; Adu-Sarkodie, Y.; Rohde, H.; Christner, M.; et al. Antibiotic resistance and clonal diversity of invasive Staphylococcus aureus in the rural Ashanti Region, Ghana. BMC Infect. Dis. 2016, 16, 720. [Google Scholar] [CrossRef] [Green Version]
- Kılıç, A.; Doğan, E.; Kaya, S.; Baysallar, M. Investigation of the presence of mecC and Panton-Valentine leukocidin genes in Staphylococcus aureus strains isolated from clinical specimens during seven years period. Mikrobiyol. Bul. 2015, 49, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Vandendriessche, S.; Vanderhaeghen, W.; Valente Soares, F.; Hallin, M.; Catry, B.; Hermans, K.; Butaye, P.; Haesebrouck, F.; Struelens, M.J.; Denis, O. Prevalence, risk factors and genetic diversity of methicillin-resistant Staphylococcus aureus carried by humans and animals across livestock production sectors. J. Antimicrob. Chemother. 2013, 68, 1510–1516. [Google Scholar] [CrossRef] [Green Version]
- Laub, K.; Tóthpál, A.; Kardos, S.; Dobay, O. Epidemiology and antibiotic sensitivity of Staphylococcus aureus nasal carriage in children in Hungary. Acta Microbiol. Immunol. Hung. 2017, 64, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, A.; Crawford, K.; Mende, K.; Murray, C.K.; Lloyd, B.; Ellis, M.; Tribble, D.R.; Weintrob, A.C. Evaluation for a novel methicillin resistance (mecC) homologue in methicillin-resistant Staphylococcus aureus isolates obtained from injured military personnel. J. Clin. Microbiol. 2013, 51, 3073–3075. [Google Scholar] [CrossRef] [Green Version]
- Ludden, C.; Brennan, G.; Morris, D.; Austin, B.; O’Connell, B.; Cormican, M. Characterization of methicillin-resistant Staphylococcus aureus from residents and the environment in a long-term care facility. Epidemiol. Infect. 2015, 143, 2985–2988. [Google Scholar] [CrossRef] [Green Version]
- Paterson, G.K.; Harrison, E.M.; Craven, E.F.; Petersen, A.; Larsen, A.R.; Ellington, M.J.; Török, M.E.; Peacock, S.J.; Parkhill, J.; Zadoks, R.N.; et al. Incidence and characterisation of methicillin-resistant Staphylococcus aureus (MRSA) from nasal colonisation in participants attending a cattle veterinary conference in the UK. PLoS ONE. 2013, 8, e68463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aqel, A.A.; Alzoubi, H.M.; Vickers, A.; Pichon, B.; Kearns, A.M. Molecular epidemiology of nasal isolates of methicillin-resistant Staphylococcus aureus from Jordan. J. Infect. Public Health. 2015, 8, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, K.; Schaumburg, F.; Fegeler, C.; Friedrich, A.W.; Köck, R. Prevalence of Multiresistant Microorganisms PMM Study. Staphylococcus aureus from the German general population is highly diverse. Int. J. Med. Microbiol. 2017, 307, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Dodémont, M.; Argudín, M.A.; Willekens, J.; Vanderhelst, E.; Pierard, D.; Miendje Deyi, V.Y.; Hanssens, L.; Franckx, H.; Schelstraete, P.; Leroux-Roels, I.; et al. Emergence of livestock-associated MRSA isolated from cystic fibrosis patients: Result of a Belgian national survey. J. Cyst. Fibros. 2019, 18, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Drougka, E.; Foka, A.; Koutinas, C.K.; Jelastopulu, E.; Giormezis, N.; Farmaki, O.; Sarrou, S.; Anastassiou, E.D.; Petinaki, E.; Spiliopoulou, I. Interspecies spread of Staphylococcus aureus clones among companion animals and human close contacts in a veterinary teaching hospital. A cross-sectional study in Greece. Prev. Vet. Med. 2016, 126, 190–198. [Google Scholar] [CrossRef]
- Nijhuis, R.H.; van Maarseveen, N.M.; van Hannen, E.J.; van Zwet, A.A.; Mascini, E.M. A rapid and high-throughput screening approach for methicillin-resistant Staphylococcus aureus based on the combination of two different real-time PCR assays. J. Clin. Microbiol. 2014, 52, 2861–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, K.; Ahmad, N.; Dryden, M.; Cortes, N.; Marsh, P.; Sitjar, A.; Wyllie, S.; Bourne, S.; Hemming, J.; Jeppesen, C.; et al. Oxacillin-susceptible methicillin-resistant Staphylococcus aureus (OS-MRSA), a hidden resistant mechanism among clinically significant isolates in the Wessex region/UK. Infection 2014, 42, 843–847. [Google Scholar] [CrossRef]
- Khairalla, A.S.; Wasfi, R.; Ashour, H.M. Carriage frequency, phenotypic, and genotypic characteristics of methicillin-resistant Staphylococcus aureus isolated from dental health-care personnel, patients, and environment. Sci. Rep. 2017, 7, 7390. [Google Scholar] [CrossRef]
- Ho, C.M.; Lin, C.Y.; Ho, M.W.; Lin, H.C.; Chen, C.J.; Lin, L.C.; Lu, J.J. Methicillin-resistant Staphylococcus aureus isolates with SCCmec type V and spa types t437 or t1081 associated to discordant susceptibility results between oxacillin and cefoxitin, Central Taiwan. Diagn. Microbiol. Infect. Dis. 2016, 86, 405–411. [Google Scholar] [CrossRef]
- van Duijkeren, E.; Hengeveld, P.; Zomer, T.P.; Landman, F.; Bosch, T.; Haenen, A.; van de Giessen, A. Transmission of MRSA between humans and animals on duck and turkey farms. J. Antimicrob. Chemother. 2016, 71, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Cikman, A.; Aydin, M.; Gulhan, B.; Karakecili, F.; Kurtoglu, M.G.; Yuksekkaya, S.; Parlak, M.; Gultepe, B.S.; Cicek, A.C.; Bilman, F.B.; et al. Absence of the mecC gene in methicillin-resistant Staphylococcus aureus isolated from various clinical samples: The first multi-centered study in Turkey. Multicenter Study. J. Infect. Public Health. 2019, 12, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Gomez, P.; Lozano, C.; González-Barrio, D.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. High prevalence of methicillin-resistant Staphylococcus aureus (MRSA) carrying the mecC gene in a semi-extensive red deer (Cervus elaphus hispanicus) farm in Southern Spain. Vet. Microbiology. 2015, 177, 326–331. [Google Scholar] [CrossRef]
- Szymanek-Majchrzak, K.; Kosiński, J.; Żak, K.; Sułek, K.; Młynarczyk, A.; Młynarczyk, G. Prevalence of methicillin resistant and mupirocin-resistant Staphylococcus aureus strains among medical students of Medical University in Warsaw. Przegl Epidemiol. 2019, 73, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Rödel, J.; Bohnert, J.A.; Stoll, S.; Wassill, L.; Edel, B.; Karrasch, M.; Löffler, B.; Pfister, W. Evaluation of loop-mediated isothermal amplification for the rapid identification of bacteria and resistance determinants in positive blood cultures. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Horner, C.; Utsi, L.; Coole, L.; Denton, M. Epidemiology and microbiological characterization of clinical isolates of Staphylococcus aureus in a single healthcare region of the UK, 2015. Epidemiol. Infect. 2017, 145, 386–396. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.R.; Estrada, J.; Ybarra, J.; Fierer, J. Comparison of the BD MAX MRSA XT to the Cepheid™ Xpert® MRSA assay for the molecular detection of methicillin-resistant Staphylococcus aureus from nasal swabs. Diagn. Microbiol. Infect. Dis. 2017, 87, 308–310. [Google Scholar] [CrossRef]
- Venugopal, N.; Mitra, S.; Tewari, R.; Ganaie, F.; Shome, R.; Rahman, H.; Shome, B.R. Molecular detection and typing of methicillin-resistant Staphylococcus aureus and methicillin-resistant coagulase-negative staphylococci isolated from cattle, animal handlers, and their environment from Karnataka, Southern Province of India. Vet. World. 2019, 12, 1760–1768. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, S.; Aspiroz, C.; Ruiz-Ripa, l.; Azcona-Gutierrez, J.M.; López-Cerero, L.; López-Calleja, A.I.; Álvarez, L.; Gomáriz, M.; Fernández, M.; Torres, C.; et al. Multicenter study of clinical non-β-lactam-antibiotic susceptible MRSA strains: Genetic lineages and Panton-Valentine leukocidin (PVL) production. Enferm. Infecc. Microbiol. Clin. 2019, 37, 509–513. [Google Scholar] [CrossRef]
- Papadopoulos, P.; Angelidis, A.S.; Papadopoulos, T.; Kotzamanidis, C.; Zdragas, A.; Papa, A.; Filioussis, G.; Sergelidis, D. Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) in bulk tank milk, livestock and dairy-farm personnel in north-central and north-eastern Greece: Prevalence, characterization and genetic relatedness. Food Microbiol. 2019, 84, 103249. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Larsen, J.; van Wijk, R.E.; Jones, O.R.; Bjørneboe Berg, T.; Angen, O.; Rhod Larsen, A. European hedgehogs (Erinaceus europaeus) as a natural reservoir of methicillin-resistant Staphylococcus aureus carrying mecC in Denmark. PLoS ONE. 2019, 14, e0222031. [Google Scholar] [CrossRef]
- Morroni, G.; Brenciani, A.; Brescini, L.; Fioriti, S.; Simoni, S.; Pocognoli, A.; Mingoia, M.; Giovanetti, E.; Barchiesi, F.; Giacometti, A.; et al. High rate of ceftobiprole resistance among clinical methicillin-resistant Staphylococcus aureus isolates from a hospital in central Italy. Antimicrob. Agents Chemother. 2018, 62, e01663-18. [Google Scholar] [CrossRef] [Green Version]
- Hefzy, E.M.; Hassan, G.M.; El Reheem, F.A. Detection of Panton-Valentine Leukocidin-positive methicillin-resistant Staphylococcus aureus nasal carriage among Egyptian health care workers. Surg. Infect. (Larchmt). 2016, 17, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.; Rakotozandrindrainy, R.; Al-Emran, H.; Dekker, D.; Hahn, A.; Jaeger, A.; Poppert, S.; Frickmann, H.; Hagen, R.M.; Micheel, V.; et al. Prevalence of nasal colonisation by methicillin-sensitive and methicillin-resistant Staphylococcus aureus among healthcare workers and students in Madagascar. B.M.C. Infect. Dis. 2016, 16, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, R.; Ramalheira, E.; Afreixo, V.; Gago, B. Methicillin-resistant Staphylococcus aureus carrying the new mecC gene--a meta-analysis. Diagn. Microbiol. Infect. Dis. 2016, 84, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, A.K.; Gustafsson, E.; Petersson, A.C.; Melander, E. Methicillin-resistant Staphylococcus aureus with mecC: A description of 45 human cases in southern Sweden. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Laurent, F.; Chardon, H.; Haenni, M.; Bes, M.; Reverdy, M.E.; Madec, J.V.; Lagier, E.; Vandenesch, F.; Tristan, A. MRSA harboring mecA variant gene mecC, France. Emerg. Infect. Dis. 2012, 18, 1465–1467. [Google Scholar] [CrossRef] [PubMed]
- Barraud, O.; Laurent, F.; François, B.; Bes, M.; Vignon, P.; Ploy, M.C. Severe human bone infection due to methicillin-resistant Staphylococcus aureus carrying the novel mecC variant. J. Antimicrob. Chemother. 2013, 68, 2949–2950. [Google Scholar] [CrossRef] [Green Version]
- Cano García, M.E.; Monteagudo Cimiano, I.; Mellado Encinas, P.; Ortega Álvarez, C. Methicillin-resistant Staphylococcus aureus carrying the mecC gene in a patient with a wound infection. Enferm. Infecc. Microbiol. Clin. 2015, 33, 287–288. [Google Scholar] [CrossRef]
- Romero-Gómez, M.P.; Mora-Rillo, M.; Lázaro-Perona, F.; Gómez-Gil, M.R.; Mingorance, J. Bacteraemia due to meticillin-resistant Staphylococcus aureus carrying the mecC gene in a patient with urothelial carcinoma. J. Med. Microbiol. 2013, 62, 1914–1916. [Google Scholar] [CrossRef] [Green Version]
- Benito, D.; Gómez, P.; Aspiroz, C.; Zarazaga, M.; Lozano, C.; Torres, C. Molecular characterization of Staphylococcus aureus isolated from humans related to a livestock farm in Spain, with detection of MRSA-CC130 carrying mecC gene: A zoonotic case? Enferm. Infecc. Microbiol. Clin. 2016, 34, 280–285. [Google Scholar] [CrossRef]
- Worthing, K.A.; Coombs, G.W.; Pang, S.; Abraham, S.; Saputra, S.; Trott, D.J.; Jordan, D.; Wong, H.S.; Abraham, R.J.; Norris, J.M. Isolation of mecC MRSA in Australia. J. Antimicrob. Chemother. 2016, 71, 2348–2349. [Google Scholar] [CrossRef] [Green Version]
- Aklilu, E.; Ying, C.H. First mecC and mecA positive livestock-associated Methicillin resistant Staphylococcus aureus (mecC MRSA/LA-MRSA) from dairy cattle in Malaysia. Microorganisms. 2020, 8, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakr, A.; Brégeon, F.; Mège, J.L.; Rolain, J.M.; Blin, O. Staphylococcus aureus nasal colonization: An update on mechanisms, epidemiology, risk factors, and subsequent infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef]
- Brown, A.F.; Leech, J.M.; Rogers, T.R.; McLoughlin, R.M. Staphylococcus aureus colonization: Modulation of host immune response and impact on human vaccine design. Front. Immunol. 2014, 4, 507. [Google Scholar] [CrossRef] [PubMed]
- Asadollahi, P.; Farahani, N.N.; Mirzaii, N.; Sajjad Khoramrooz, S.S.; van Belkum, A.; Asadollahi, K.; Dadashi, M.; Darban-Sarokhalil, D. Distribution of the most prevalent spa types among clinical isolates of Methicillin-Resistant and -Susceptible Staphylococcus aureus around the World: A Review. Front. Microbiol. 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmekki, M.; Mammeri, H.; Hamdad, F.; Rousseau, F.; Canarelli, B.; Biendo, M. Comparison of Xpert MRSA/SA Nasal and MRSA/SA ELITe MGB assays for detection of the mecA gene with susceptibility testing methods for determination of methicillin resistance in Staphylococcus aureus isolates. J. Clin. Microbiol. 2013, 51, 3183–3191. [Google Scholar] [CrossRef] [Green Version]
- Stegger, M.; Lindsay, J.A.; Sørum, M.; Gould, K.A.; Skov, R. Genetic diversity in CC398 methicillin-resistant Staphylococcus aureus isolates of different geographical origin. Clin. Microbiol. Infect. 2010, 16, 1017–1019. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, J.; Espinosa-Gongora, C.; Stamphøj, I.; Rhod Larsen, A.; Guardabassi, L. Carriage frequency, diversity and methicillin resistance of Staphylococcus aureus in Danish small ruminants. Vet. Microbiol. 2013, 163, 110–115. [Google Scholar] [CrossRef]
- Paterson, G.K.; Larsen, A.R.; Robb, A.; Edwards, G.E.; Pennycott, T.W.; Foster, G.; Mot, D.; Hermans, K.; Baert, K.; Peacock, S.J.; et al. The newly described mecA homologue, mecA(LGA251), is present in methicillin-resistant Staphylococcus aureus isolates from a diverse range of host species. J. Antimicrob. Chemother. 2012, 67, 2809–2813. [Google Scholar] [CrossRef]
- Weinert, L.A.; Welch, J.J.; Suchard, M.A.; Lemey, P.; Rambaut, A.; Fitzgerald, J.R. Molecular dating of human-to-bovid host jumps by Staphylococcus aureus reveals an association with the spread of domestication. Biol. Lett. 2012, 8, 829–832. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ripa, L.; Gómez, P.; Alonso, C.A.; Camacho, M.C.; de la Puente, J.; Fernández-Fernández, R.; Ramiro, Y.; Quevedo, M.A.; Blanco, J.M.; Zarazaga, M.; et al. Detection of MRSA of Lineages CC130-mecC and CC398-mecA and Staphylococcus delphini-lnu(A) in Magpies and Cinereous Vultures in Spain. Microb. Ecol. 2019, 78, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Ford, A. mecC-harboring methicillin-resistant Staphylococcus aureus: Hiding in plain sight. J. Clin. Microbiol. 2017, 56, e01549-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenni, M.; Châtre, P.; Dupieux, C.; Métayer, V.; Maillard, K.; Bes, M.; Madec, J.Y.; Laurent, F. mecC-positive MRSA in horses. J. Antimicrob. Chemother. 2015, 70, 3401–3402. [Google Scholar] [CrossRef] [Green Version]
- Medhus, A.; Slettemeås, J.S.; Marstein, L.; Larssen, K.W.; Sunde, M. Methicillin-resistant Staphylococcus aureus with the novel mecC gene variant isolated from a cat suffering from chronic conjunctivitis. J. Antimicrob. Chemother. 2013, 68, 968–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenni, M.; Châtre, P.; Tasse, J.; Nowak, N.; Bes, M.; Madec, J.Y.; Laurent, F. Geographical clustering of mecC-positive Staphylococcus aureus from bovine mastitis in France. J. Antimicrob. Chemother. 2014, 69, 2292–2293. [Google Scholar] [CrossRef] [Green Version]
- Gindonis, V.; Taponen, S.; Myllyniemi, A.L.; Pyörälä, S.; Nykäsenoja, S.; Salmenlinna, S.; Lindholm, L.; Rantala, M. Occurrence and characterization of methicillin-resistant staphylococci from bovine mastitis milk samples in Finland. Acta Vet. Scand. 2013, 55, 61. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Reference | Country | Sampling Date | Prevalence: mecC Positive Isolates/S. aureus or Methicillin Resistant S. aureus (MRSA) (%) | Type of Sample/Infection (Number of Isolates) | Clonal Complex: Sequence Type 2 (Number of Isolates) | IEC 3 (Number of Isolates) | Non-beta lactam Resistance (Number of Isolates) 4 | Possible Relationship with Animals |
|---|---|---|---|---|---|---|---|---|
| [18,24] | Denmark | 1960–2011 | 112 (0.21%)/53746 MRSA | Wound (37), skin (26), blood (8), post-operative wound (5), urine (4), eye/ear (2), impetigo (2), unknown (28) | CC130 (98)/CC2361 (14): ST2173, ST2174 | Negative (2) | Q (NOR) (1), S (111) | Most were from rural areas (106): 4 with contact with animals |
| [16] | United Kingdom (UK) and Denmark | 1975–2011 | 51 (0.04%)/approximately 120500 S. aureus | Screen swab (10), Skin and soft tissue infections (7), nose (5), wound (5), blood (4), skin (4), nose/mouth (2), ear (1), eye/ear (1), finger (1), fluid (1), hand (1), PEG site(1), sputum (1), toe (1), unknown (6) | CC130: ST130 (18), ST1245 (3), ST1764 (3), ST1945 (3), ST1526 (1), ST1944 (1), n/d (17)/CC1943: ST1943 (1), ST1946 (1)/CC425: ST425 (3) | - | S (51) | - |
| [25] | Denmark | 1975–011 | 127/-: in routine testing 12 (5.91%)/203 MRSA | - | CC130 (107): ST130, ST1245, ST1526, ST1945/CC1943 (14): ST1943, ST1946, ST2173, ST2174/CC425 (6): ST425 | - | - | - |
| [26] | Ireland | 2000–2012 | 1 (1.14%)/88 MRSA | - | CC130 (1) | Negative (1) | Q (NOR) (1) | Patient lived on a Farm |
| [27] | Germany | 2000–2016 | 2 (0.16%)/1277 MRSA | - | CC130 (2) | - | - | - |
| [28] | Austria | 2002–2012 | 1 (0.31%)/327 MRSA | - | CC130 (1) | - | - | - |
| [29] | Belgium | 2003–2012 | 9 (0.18%)/4869 S. aureus | Screen swab (4), urine (2), wound (2), sputum (1) | CC130 (4)/CC49 (3)/CC1943 (2) | - | S (9) | Most were from a rural area with a high density of cattle farms |
| [30,31] | Germany and The Netherlands | 2004–2011 | 16/− | nasal swab (11), wound (2), joint aspirate (1), mouth swab (1), sputum (1) | CC130 (14)/CC1943: ST2361 (1)/CC599: ST599 (1) | Negative (16) | S (1) | - |
| [32] | Germany | 2004–2005 2010-2011 | 2 (0.06%)/3207 MRSA | Screen swab (1), sputum (1) | - | - | - | - |
| [33] | Germany | 2006–2011 | 11 (0.09%)/12691 MRSA | Wound (8), dermatitis (1), nasal swab (1), nosocomial pneumonia (1) | CC130 (11) | Negative (11) | Q [CIP (1), MFL (1)] (2), S (9) | Veterinarian (1) |
| [34,35] | UK | 2006–2012 | 2/− | Screen swab (2) | CC130 (2) | - | - | - |
| [36] | Slovenia | 2006–2013 | 6 (1.52%)/395 MRSA | Wound (4), Screen swab (2) | CC130: ST130 (6) | Negative (6) | S (6) | Most were from rural areas |
| [37] | Spain | 2008–2013 | 2 (0.04%)/5505 S. aureus | Joint fluid (1), wound (1) | CC130: ST1945 (2) | - | S (2) | -. |
| [38] | Austria | 2009-2013 | 6 (2%)/301 S. aureus | blood (2), screen swab (2), wound (2) | CC130: ST130 (3), new SLV (1)/ CC599: ST599 (2) | - | S (6) | Contact with pet rabbit (1), unknown (5) |
| [39] | Denmark | 2010-2011 | 6 (6.32%)/95 MRSA | - | - | - | - | - |
| [40] | England | 2011-2012 | 9 (0.45%)/2010 MRSA | Screen swab (6), wound (2), leg ulcer (1) | CC130: ST130 (2), ST1245(4), ST2573 (1), ST2574 (1)/CC425: ST425 (1) | - | L (CLI) (1) -M (ERY) (1), S (8) | - |
| [41] | UK | 2012-2013 | 12 (0.53%)/2282 MRSA | Screen swab (9), SSTI (3) | CC130: ST1245 (6), ST130 (2), ST1945 (1), ST2574 (1)/CC425: ST425 (1)/CC1943: ST1943 (1) | Negative (11)/type E (1) | M (ERY) (1), S (11) | - |
| [42] | England | 2015 | 1 (0.08%)/1242 MRSA | Screen swab (1) | CC130: ST130 (1) | Negative (1) | S (1) | - |
| [43] | England | 2018-2019 | 1 (0.7%)/142 S. aureus | - | - | - | - | - |
| [44] | Germany, UK, Belgium | - | 80/- | - | - | - | - | - |
| Reference | Country | Sampling Date | Type of Samples 1 | Number of Samples or (S. aureus or MRSA) Isolates |
|---|---|---|---|---|
| [45] | Switzerland | 2005–2012 | Clinical/screening | 1695 S. aureus isolates |
| [46] | Ghana | 2007–2012 | Clinical | 9834 blood samples |
| [47] | Turkey | 2007–2014 | Clinical | 1700 S. aureus isolates |
| [48] | Belgium | 2009–2011 | Screening | 149 farmers and family members (41 MRSA isolates) |
| [49] | Hungary | 2009–2011 | Screening | 878 children |
| [50] | United States | 2009–2011 | Clinical/screening | 364 S. aureus isolates (102 MRSA isolates) |
| [51] | Ireland | 2011 | Screening | 64 residents |
| [52] | UK | 2011 | Screening | 307 cattle veterinarians |
| [53] | Jordan | 2011–2012 | Screening | 716 humans (56 MRSA isolates) |
| [54] | Germany | 2011–2013 | Screening | 1878 non-hospitalized adults |
| [55] | Belgium | 2012–2013 | Clinical | 510 cystic fibrosis patients |
| [56] | Greece | 2012–2013 | Screening | 18 veterinary personnel |
| [57] | The Netherlands | 2012–2013 | Clinical/screening | 13,387 samples |
| [58] | UK | 2012–2013 | Clinical | 500 S. aureus isolates |
| [59] | Egypt | 2013 | Clinical/screening | 1300 dental patients |
| [60] | Taiwan | 2013–2014 | Clinical/screening | 3717 S. aureus isolates |
| [61] | Turkey | 2013–2014 | Screening | 7 MRSA isolates |
| [62] | Turkey | 2013–2016 | Clinical/screening | 494 MRSA isolates |
| [63] | Spain | 2014 | Screening | 15 humans in contact with animals |
| [64] | Poland | 2014–2016 | Screening | 955 students (only one MRSA isolate) |
| [65] | Germany | 2015 | Clinical | 140 Gram-positive isolates |
| [66] | UK | 2015 | Clinical | 520 S. aureus isolates |
| [67] | United States | 2015 | Screening | 479 patients |
| [68] | India | 2015–2017 | Screening | 32 animal handlers |
| [69] | Spain | 2016 | Clinical/screening | 45 non-beta-lactam susceptible MRSA isolates |
| [70] | Greece | 2016–2017 | Screening | 68 farmers |
| [71] | Denmark | 2017 | Screening | 16 workers at wildlife rehabilitation centres |
| [72] | Italy | 2017–2018 | Clinical/screening | 102 MRSA isolates |
| [73] | Egypt | - | Screening | 223 health care personnel |
| [74] | Madagascar | - | Screening | 1548 students and healthcare workers |
| Reference 1 | Country | Sampling Date | Number of Described Case Reports | Year-Old Patient | Type of Sample/Infection 2 | Clonal Complex: Sequence Type 3 | IEC 4 | Non-Beta Lactam Resistance 5 | Possible Relationship with Animals |
|---|---|---|---|---|---|---|---|---|---|
| [76] | Sweden | 2005–2014 | 45 | Median age (range) 60 (2–86) | Wound, sputum, nasopharynx | CC130/CC2361 | - | L-M (1 isolate), S (44 isolates) | Most were from a rural area: farmer (1), patients lived on farms (4) |
| [77] | France | 2007 | 1 | 67 | Fluid of lesion heel | CC130: ST1945 | - | - | - |
| [37] | Spain | 2008–2013 | 7 | 3, 50, 63, 64, 76, 80, 85 | Blood, joint fluid, nasal screen swab, urine, wound | CC130: ST130, ST1945 | - | S | No epidemiological data were available except for one patient who did not have any contact with animals |
| [78] | France | 2010 | 1 | 48 | Blood, ear fluid, retrosternal abscess | CC130 | Negative | S | Contact with cows |
| [17] | Ireland | 2010 | 2 | 64, 85 | - | CC130: ST130, ST1764 | Negative | S (but detection of tet efflux) | - |
| [45] | Switzerland | 2011 | 1 | 59 | Groin, nose, and throat screen swab | CC130: ST130 | - | - | Contact with a cat |
| [79] | Spain | 2012 | 1 | 46 | Skin lesion swab | CC130 | - | S | Patient lived in rural area with high density of livestock animals |
| [36] | Slovenia | 2013 | 1 | 86 | Nose and skin screen swabs | CC130: ST130 | Negative | S | Patient lived on a farm and had contact with pigs, cats and dogs |
| [80] | Spain | 2013 | 1 | 76 | Blood | CC130: ST1945 | - | S | No contact with livestock |
| [81] | Spain | 2013–2014 | 1 | 34 | Superficial skin lesion swab | CC130: ST130 | Negative | S | Contact with livestock animals |
| Type of Sample/Infection | Number of Isolates 2 | References |
|---|---|---|
| Screen swab 1 | 54 | [16,29,30,31,32,33,34,36,38,40,41,42,45,76] |
| Skin lesion/dermatitis/impetigo wound/post-operative wound/skin and soft tissue infections | 158 | 16,24,29,30,33,36-38,40,41,76,79,81 |
| Blood | 16 | [16,24,37,38,80] |
| Urine | 7 | [24,29,37] |
| Nosocomial pneumonia/sputum/ Tracheal aspirate | 7 | [29,30,32,33,76] |
| Nose | 5 | [16] |
| Eye/ear | 3 | [16,24] |
| Fluid of heel/joint fluid | 3 | [30,37,77] |
| Mouth/Nose | 2 | [16] |
| Ear | 1 | [16] |
| Finger | 1 | [16] |
| Fluid | 1 | [16] |
| Hand | 1 | [16] |
| Percutaneous endoscopic gastrostomy site | 1 | [16] |
| Retrosternal abscess | 1 | [78] |
| Toe | 1 | [16] |
| Unknown | 34 | [16,24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano, C.; Fernández-Fernández, R.; Ruiz-Ripa, L.; Gómez, P.; Zarazaga, M.; Torres, C. Human mecC-Carrying MRSA: Clinical Implications and Risk Factors. Microorganisms 2020, 8, 1615. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101615
Lozano C, Fernández-Fernández R, Ruiz-Ripa L, Gómez P, Zarazaga M, Torres C. Human mecC-Carrying MRSA: Clinical Implications and Risk Factors. Microorganisms. 2020; 8(10):1615. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101615
Chicago/Turabian StyleLozano, Carmen, Rosa Fernández-Fernández, Laura Ruiz-Ripa, Paula Gómez, Myriam Zarazaga, and Carmen Torres. 2020. "Human mecC-Carrying MRSA: Clinical Implications and Risk Factors" Microorganisms 8, no. 10: 1615. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101615