Phenolics and Plant Allelopathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

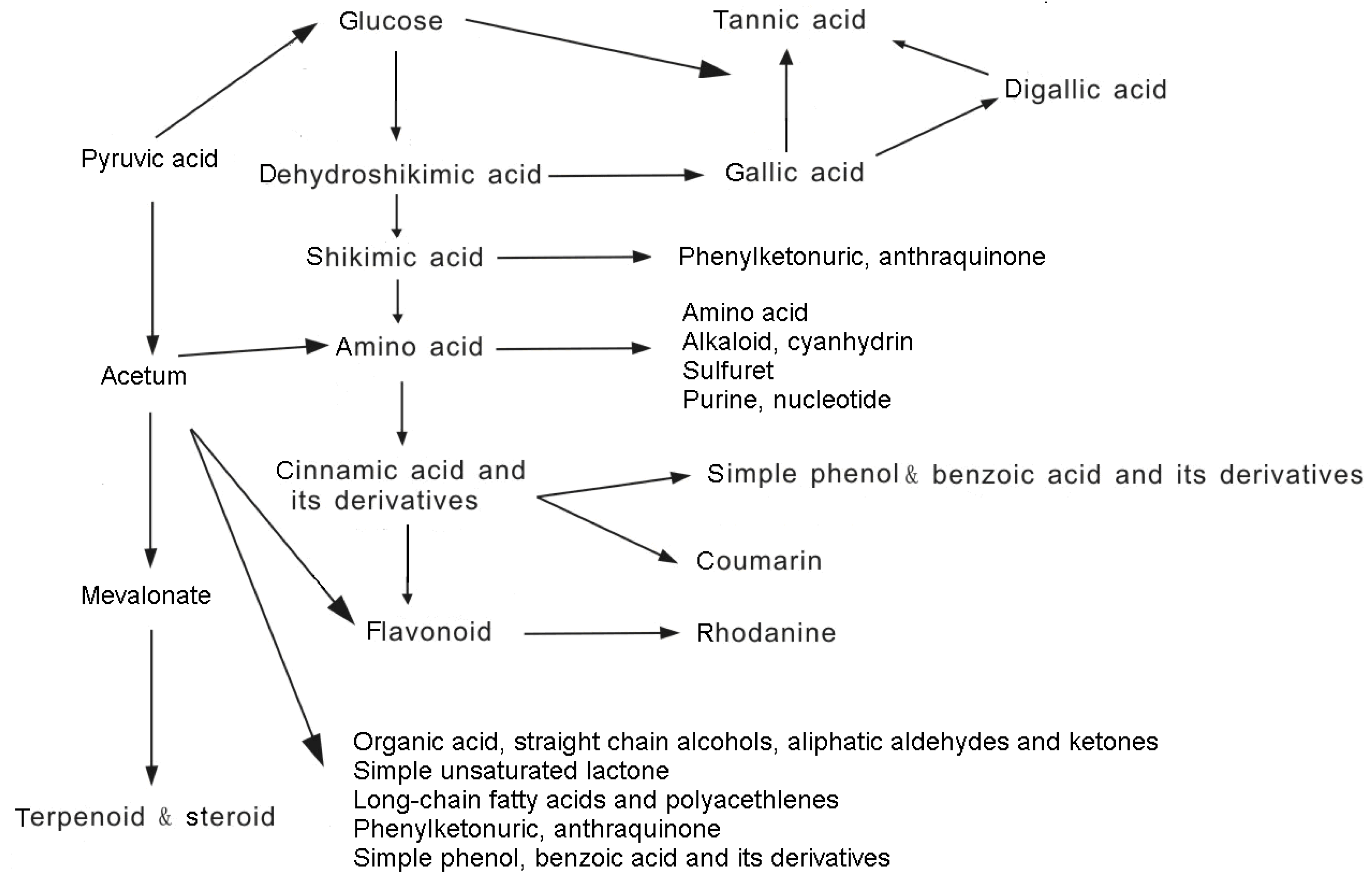
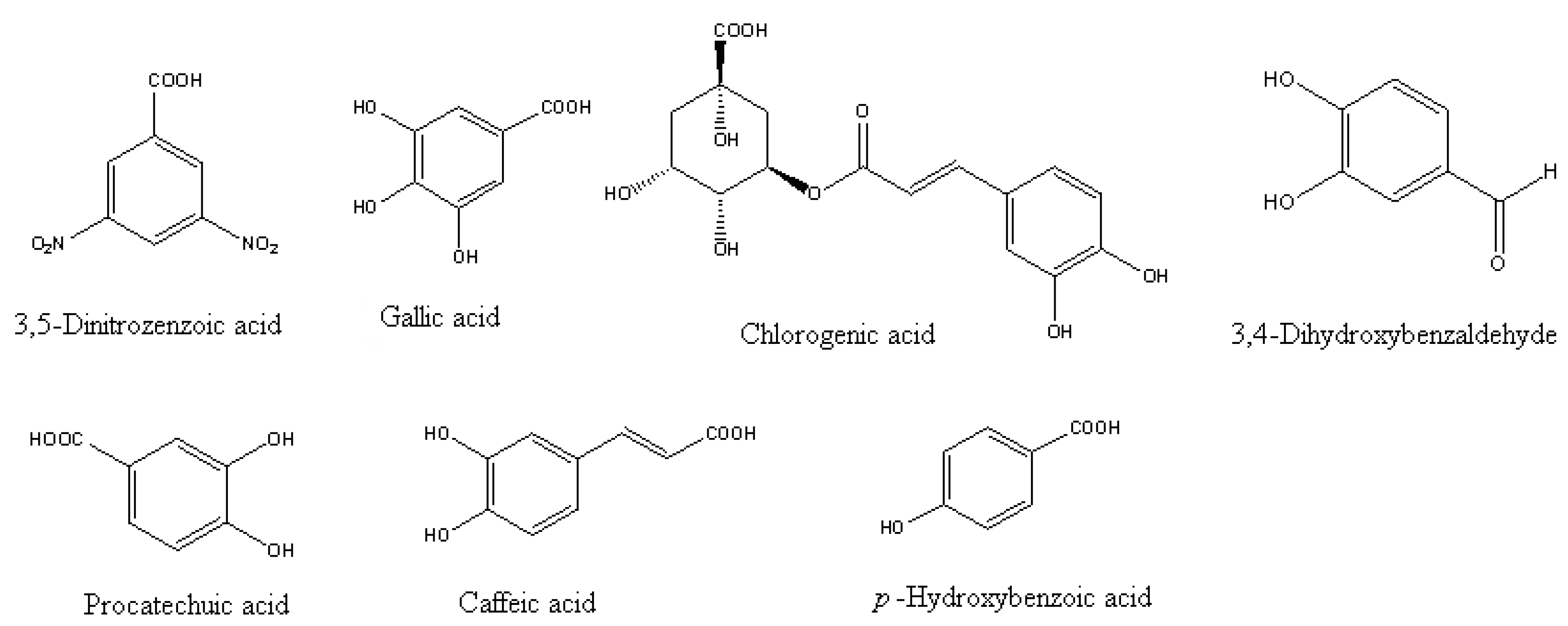
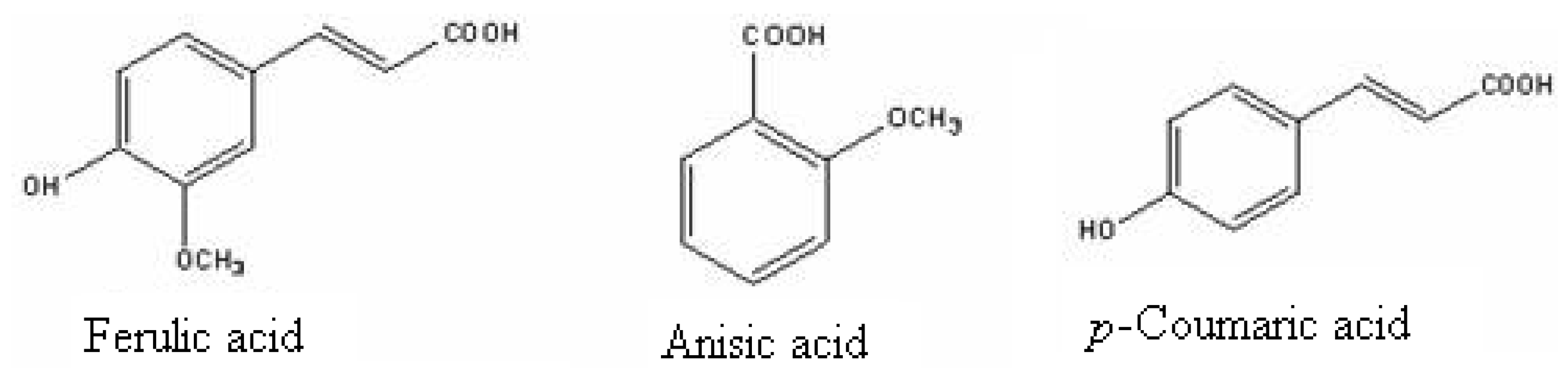
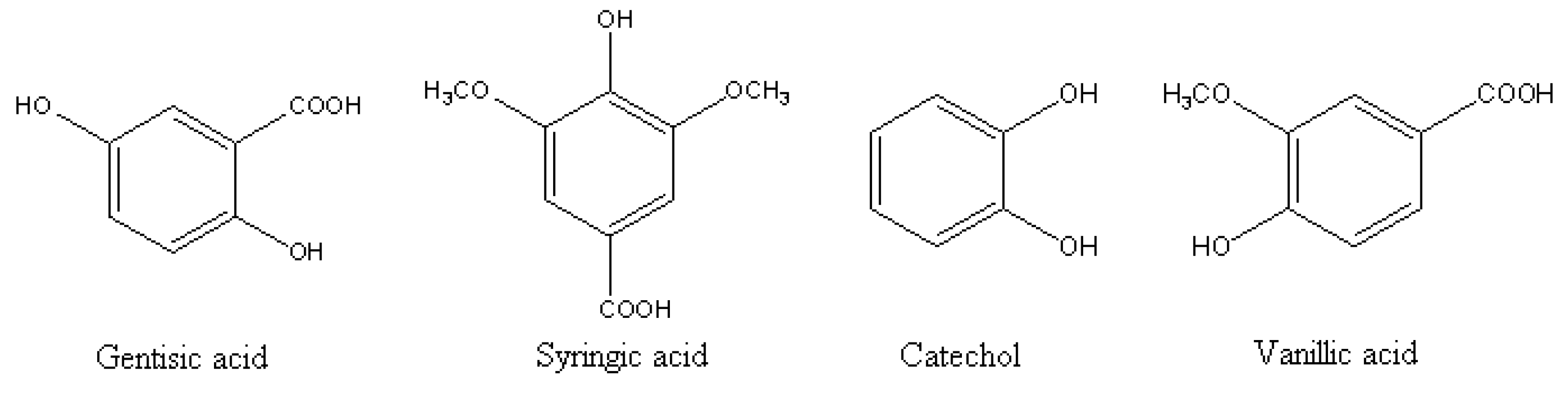
2. Origin and Allelopathy Effects of Phenolics



3. Analysis Methods for Phenolic Allelochemicals
3.1. Extration Methods of Total Phenolics with Solvents
3.2. Separation and Detection Methods of Total Phenolics
3.2.1. Liquid-liquid Extraction
3.2.2. Chromatography
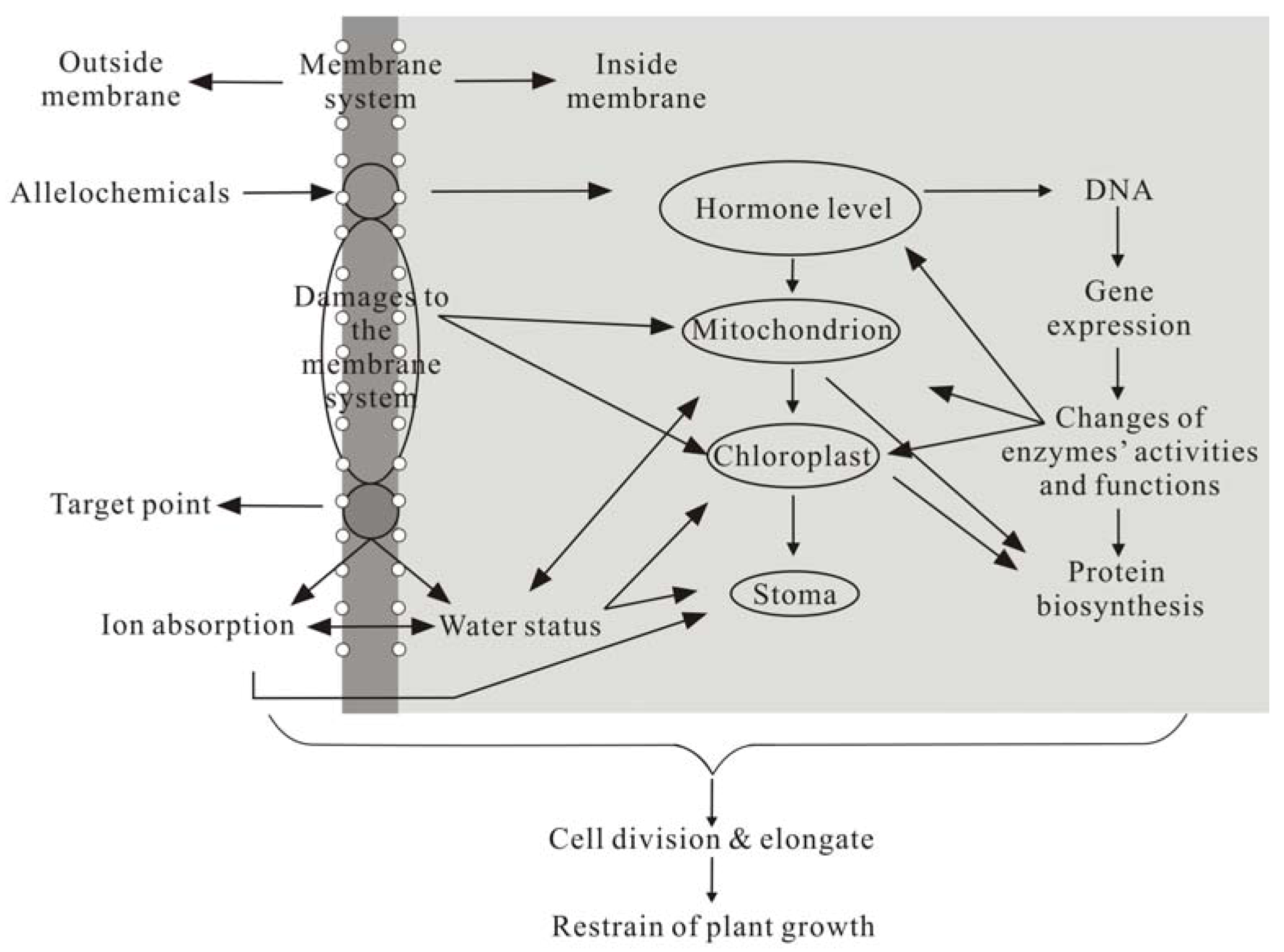
4. Allelopathy Mechanisms of Phenolics
4.1. Changes in Membrane Permeability and Inhibition of Plant Nutrient Uptake

4.2. Inhibition of Cell Division, Elongation, and Submicroscopic Structure

4.3. Effects on Plant Photosynthesis and Respiration
4.4. Effects on Various Enzyme Function and Activities

4.5. Effects on Synthesis of Plant Endogenous Hormones
4.6. Effects on Protein Synthesis

5. The Forefront of Plant Allelopathy and Our Work
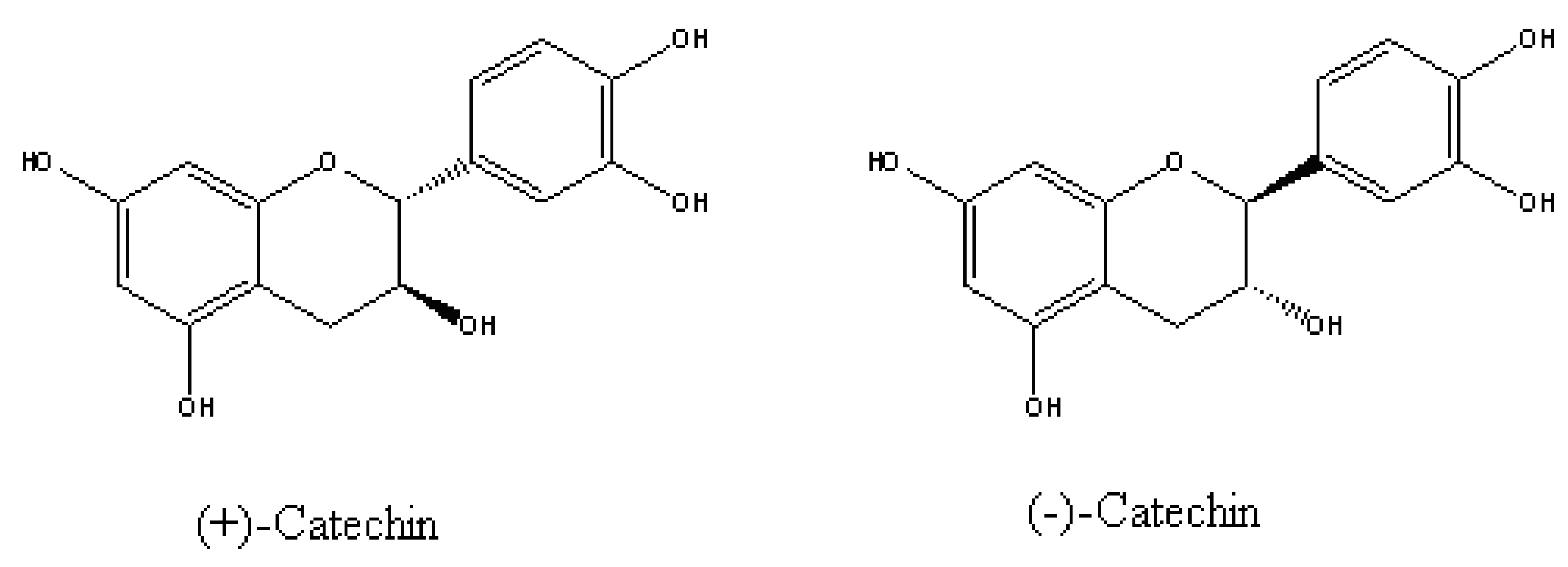
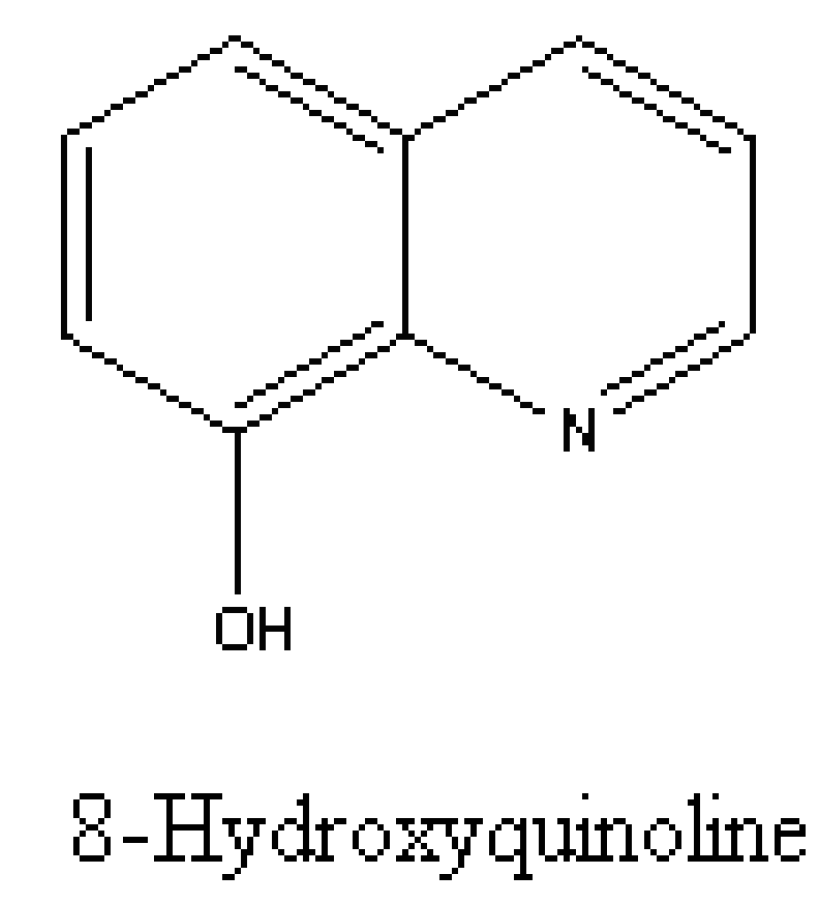
5.1. Are (±)-Catechin and 8-Hydroxyquinoline Putative Allelochemicals Underlying Knapweed’s Invasion Success?


5.2. Several International Allelopathy Research Centers
5.3. Our Work on Schrenk Spruce

6. Conclusions
Acknowledgements
- Sample Availability: Not available.
References and Notes
- Rice, E.L. Allelopathy; Academic Press Inc: Norman, Oklahoma, 1984. [Google Scholar]
- Weston, L.A.; Duke, S.O. Weed and crop allelopathy. Plant Sci. 2003, 22, 367–389. [Google Scholar]
- Zeng, R.S.; Mallik, A.U.; Luo, S.M. Allelopathy in Sustainable Agriculture and Forestry; Springer Science+Business Media, LLC: New York, NY, USA, 2008. [Google Scholar]
- Molisch, H. Der Einfluss einer Pflanze auf die andere-Allelopathie; Fischer: Jena, Austria, 1937. [Google Scholar]
- Singh, H.P.; Kohli, R.K.; Batish, D.R. Allelopathy in agroecosystems: An overview. J. Crop Prod. 2001, 4, 1–41. [Google Scholar]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic plant invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Ridenour, W.M.; Callaway, R.M. The relative importance of allelopathy in interference: the effects of an invasive weed on a native bunchgrass. Oecologia 2001, 126, 444–450. [Google Scholar] [CrossRef]
- Haribal, M.; Enwick, J.A.A. Isovitexin 6″″-O-β-D-glucopyranoside: a feeding deterrent to Pieris napi oleracea from Alliaria petiolata. Phytochemistry 1998, 47, 1237–1240. [Google Scholar]
- Wang, Q.; Ruan, X.; Li, Z.H.; Pan, C.D. Autotoxicity of plants and research of coniferous forest autotoxicity. Sci. Sil. Sin. 2006, 43, 134–142. [Google Scholar]
- Capasso, R. The chemistry, biotechnology and ecotoxicology of the polyphenols naturally occurring in vegetable wastes. Curr. Topics Phytochem. 1997, 1, 145–156. [Google Scholar]
- Capasso, R.; Cristinzio, G.; Evidente, A.; Scognamiglio, F. Isolation, spectroscopy and selective phytotoxic effects of polyphenols from vegetable waste waters. Phytochemistry 1992, 31, 4125–4128. [Google Scholar] [CrossRef]
- Hartmann, T. Diversity and variability of plant secondary metabolism: A mechanistic view. Entomol. Gen. Appl. 1996, 80, 177–188. [Google Scholar] [CrossRef]
- Franz, H. Secondary Metabolites as Plant Traits: Current Assessment and Future Perspectives. Plant Sci. 2002, 21, 273–322. [Google Scholar]
- Santana, C.M.; Ferrera, Z.S.; Padrón, M.E.T.; Rodríguez, J.J.S. Methodologies for the extraction of phenolic compounds from environmental samples: New Approaches. Molecules 2009, 14, 298–320. [Google Scholar] [CrossRef]
- Chou, C.H.; Leu, L.L. Allelopathic substances and interactions of Delonix regia (BOJ) RAF. J. Chem. Ecol. 1992, 18, 2285–2303. [Google Scholar] [CrossRef]
- Chou, C.H.; Lin, H.J. Autointoxication mechanism of Oryza sativ I. Phytotoxic effects of decomposing rice residues in soil. J. Chem. Ecol. 1976, 2, 353–367. [Google Scholar] [CrossRef]
- Chou, C.H.; Muller, C.H. Allelopathic mechanisms of Arctostaphylos glandulosa var. zacaensis. Am. Midl. Nat. 1972, 88, 34–347. [Google Scholar]
- Chou, C.H.; Lee, Y.Y. Allelopathic dominance of Miscanthus transmorrisonensis in an alpine grassland community in Taiwan. J. Chem. Ecol. 1991, 17, 2267–2281. [Google Scholar] [CrossRef]
- Tang, C.S. Continuous trapping techniques for the study of allelochemicals from higher plants. In The Science of Allelopathy; Putnam, A.R., Tang, C.S., Eds.; Wiley: New York, NY, 1986; pp. 113–131. [Google Scholar]
- Waller, G.R. Atlelochemicals: Role in Agriculture and Forestry. In ACS Symposium Series 330; American Chemical Society: Washington, DC, USA, 1987; p. 606. [Google Scholar]
- Batish, D.R.; Kaur, S.; Singh, H.P.; Kohli, R.K. Role of root-mediated interactions in phytotoxic interference of Ageratum conyzoides with rice (Oryza sativa). Flora 2008, 204, 388–395. [Google Scholar]
- Mahall, B.E.; Callaway, R.M. Root communication mechanisms and intercommunity distributions of 2 mojave desert shrubs. Ecology 1992, 73, 2145–2151. [Google Scholar] [CrossRef]
- Ghafar, A.; Saleem, B.; Qureshi, M.J. Allelopathic effects of sunflower on germination and seedling growth of wheat. Pak. Pak. J. Biol. Sci. 2000, 3, 1301–1302. [Google Scholar] [CrossRef]
- Sasikumar, K.; Vijayalakshmi, C.; Parthiban, K.T. Allelopathic effects of Eucalyptus on blackgram (Phaseolus mungo L.). Allelopathy J. 2002, 9, 205–214. [Google Scholar]
- Willis, R.J. The historical bases of the concept of allelopathy. J. Hist. Biol. 1985, 18, 71–102. [Google Scholar] [CrossRef]
- Hostettmann, K.; Marston, A.; Hostettmann, M. Preparative Chromatography Techniques: Application in Natural Product Isolation; Springer: Berlin, Germany, 1997. [Google Scholar]
- Djurdjevic, L.; Mitrovic, M.; Pavlovic, P. Methodology of allelopathy research: 2. Forest ecosystems. Allelopathy J. 2007, 20, 1–24. [Google Scholar]
- Putnam, A.R.; Tang, C.S. The science of Allelopathy; Wiley: New York, NY, USA, 1986; pp. 1–19. [Google Scholar]
- Schmidt, S.K. Ecological implications of the destruction of juglone (5-hydroxy-1,4-naphthoquinone) by soil bacteria. J. Chem. Ecol. 1990, 16, 3547–3549. [Google Scholar] [CrossRef]
- Wu, H.; Haig, T.; Pratley, J.; Lemerle, D.; An, M. Allelochemicals in wheat (Triticum aestivum L.): Production and exudation of 2,4-dihydroxy-7-methoxy-1,4- benzoxazin-3-one. J. Chem. Ecol. 2001, 27, 1691–1700. [Google Scholar] [CrossRef]
- Inderjit. Plant phenolics in allelopathy. Bot. Rev. 1996, 62, 186–202. [CrossRef]
- Hadacek, F. Secondary metabolites as plant traits: Current assessment and future perspectives. Crit. Rev. Plant Sci. 2002, 21, 273–322. [Google Scholar] [CrossRef]
- Liebler, D.C.; Burr, J.A.; Philips, L.; Ham, A.J.L. Gas chromatography-mass spectrometry analysis of vitamin E and its oxidation products. Anal. Biochem. 1996, 263, 27–34. [Google Scholar]
- Durant, A.A.; Fente, C.A.; Franco, C.M.; Vazquez, B.I.; Cepeda, A. Gas-chromatography-tandem mass spectrometry determination of 17 α-ethinylestradiol residue in the hair of cattle, application to treated animals. J. Agric. Food Chem. 2002, 50, 436–440. [Google Scholar] [CrossRef]
- Van Pelt, C.D.; Haggarty, H.; Brenna, J.T. Quantitative subfemtomole analysis of α-tocopherol and deuterated isotopomers in plasma using tabletop GC/MS/MS. Anal. Chem. 1998, 70, 4369–4375. [Google Scholar] [CrossRef]
- Seal, A.N.; Pratley, J.E.; Haig, T; An, M. Identification and quantitation of compounds in a series of allelopathic and non-allelopathic rice root exudates. J. Chem. Ecol. 2004, 30, 1647–1662. [Google Scholar] [CrossRef]
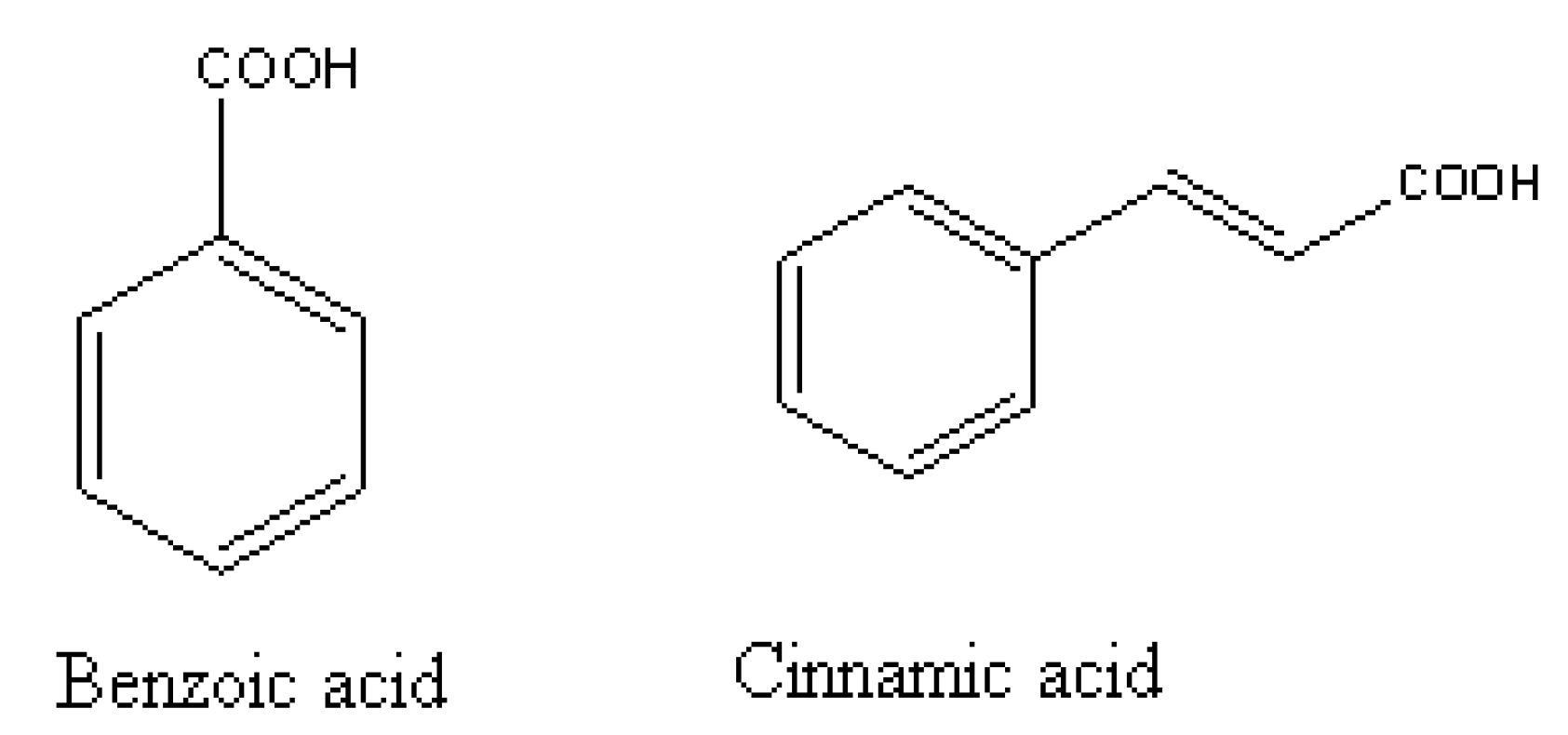
- Politycka, B. Free and glucosylated phenolics, phenol-beta-glucosyltransferase activity and membrane perability in cucumber roots affected by derivatives of cinnamic and benzoic acid. Acta Physiol. Plantarum. 1997, 19, 311–317. [Google Scholar] [CrossRef]
- Cruz, O.R.; Anaya, A.L.; Hernandez-Bautista, B.E. Effects of allelochemical stress produced by sicyosdeppei on seedling root ultrastructure of Phaseolous valgaris and Cucubita ficifolia. J. Chem. Ecol. 1998, 24, 2039–2057. [Google Scholar] [CrossRef]
- Li, H.H.; Inoue, M.; Nishimura, H; Mizutani, J.; Tsuzuki, E. Interaction of trans-cinnamic acid, its related phenolic allelochemicals, and abscisic-acid in seedling growth and seed-germination of lettuce. J. Chem. Ecol. 1993, 19, 1775–1787. [Google Scholar] [CrossRef]
- Patterson, D.T. Effects of allelopathic chemicals on growth and physiological response of soybean(Glycine max). Weed Sci. 1981, 29, 53–58. [Google Scholar]
- Yu, J.Q.; Ye, S.F.; Zhang, M.F.; Hu, W.H. Effects of root exudates and aqueous root extracts of cucumber (Cucumis sativus) and allelochemicals, on photosynthesis and antioxidant enzymes in cucumber. Biochem. Syst. Ecol. 2003, 31, 129–139. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy; Academic Press: New York, NY, USA, 1974; p. 353. [Google Scholar]
- Politycka, B. Phenolics and the activities of phenylalanine ammonia-1yase, phenol-beta-glucosyltransferase and beta-glucosidase in cucumber roots as affected by phenolic allelochemicals. Acta Physiol. Plantarum. 1998, 20, 405–410. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kaur, S.; Kohli, R.K.; Yadav, S.S. Caffeic acid affects early growth, and morphogenetic response of hypocotyl cuttings of mung bean (Phaseolus aureus). J. Plant Physiol. 2008, 165, 297–305. [Google Scholar] [CrossRef]
- Devi, S.R. Effects of ferulic acid on growth and hydrolytic enzyme activities of germinating maize seeds. J. Chem. Ecol. 1992, 18, 1981–1990. [Google Scholar]
- He, H.Q.; Lin, W.X. Studies on allelopathic physiobiochemical characteristics of rice. Chin. J. Eco-Agric. 2001, 9, 56–57. [Google Scholar]
- Zeng, R.S.; Luo, S.M.; Shi, Y.H. Physiological and biochemical mechanism of allelopathy of secalonic acid on higher plants. Agron. J. 2001, 93, 72–79. [Google Scholar] [CrossRef]
- Leslie, C.A. Romani R.J. Inhibition of ethylene biosynthesis by salicylic acid. Plant Physiol. 1998, 88, 833–837. [Google Scholar]
- Ni, H.W. Present status and prospect of crop allelopathy in China. In Rice Allelopathy; Kim, K.U., Shin, D.H., Eds.; Kyunpook National University: Kyungpook, Korea, 2000; pp. 41–48. [Google Scholar]
- Einhellig, F.A.; Galindo, J.C.G.; Molinillo, J.M.G.; Cutler, H.G. Mode of allelochemical action of phenolic compounds. In Allelopathy: Chemistry and Mode of Action of Allelochemicals; CRC Press: Boca Raton, FL, USA, 2004; pp. 217–238. [Google Scholar]
- Watson, A.K.; Renney, A.J. The biology of Canadian weeds. 6. Centaurea diffusa and Centaurea maculosa. Can. J. Plant Sci. 1974, 54, 687–701. [Google Scholar] [CrossRef]
- Tyser, R.W.; Key, C.H. Spotted knapweed in natural area fescue grasslands: An ecological assessment. Northwest Sci. 1988, 62, 981–987. [Google Scholar]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436. [Google Scholar] [CrossRef]
- Bais, H.P.; Walker, T.S.; Stermitz, F.R.; Hufbauer, R.A.; Vivanco, J.M. Enantiomeric-dependent phytotoxic and antimicrobial activity of (±)-catechin. A rhizosecreted racemic mixture from spotted knapweed. Plant Physiol. 2002, 128, 1173–1179. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Perry, L.G.; Thelen, G.C.; Ridenour, W.M.; Weir, T.L.; Callaway, R.M.; Paschke, M.W.; Vivanco, J.M. Dual role for an allelochemical (±)-catechin from Centaurea maculosa root exudates regulates conspecific seedling establishment. J. Ecol. 2005, 93, 1126–1135. [Google Scholar] [CrossRef]
- Thelen, G.C.; Vivanco, J.M.; Newingham, B.; Good, W.; Bais, H.P.; Landers, P.; Caesar, A.; Callaway, R.M. Insect herbivory stimulates allelopathic exudation by an invasive plant and the suppression of natives. Ecol. Lett. 2005, 8, 209–217. [Google Scholar] [CrossRef]
- Weir, T.L.; Bais, H.P.; Stull, V.J.; Callaway, R.M.; Thelen, G.C.; Ridenour, W.M.; Bhamidi, S.; Stermitz, F. R.; Vivanco, J. M. Oxalate contributes to the resistance of Gaillardia grandiflora and Lupinus sericeus to a phytotoxin produced by Centaurea maculosa. Planta 2006, 223, 785–795. [Google Scholar] [CrossRef]
- Weir, T.L.; Bais, H.P.; Vivanco, J.M. Intraspecific and interspecific interactions mediated by a phytotoxin, (−)-catechin, secreted by the roots of Centaurea maculosa (spotted knapweed). J. Chem. Ecol. 2003, 29, 2379–2394. [Google Scholar] [CrossRef]
- Perry, L.G.; Johnson, C.; Alford, É.R.; Vivanco, J.M.; Paschke, M.W. Screening of grassland plants for restoration after spotted knapweed invasion. Restor. Ecol. 2005, 13, 725–735. [Google Scholar] [CrossRef]
- Thorpe, A. Biochemical effects of Centaurea maculosa on soil nutrient cycles and plant communities; PhD Dissertation, University of Montana: Missoula, MT, USA, 2006. [Google Scholar]
- Blair, A.C.; Hanson, B.D.; Brunk, G.R.; Marrs, R.A.; Westra, P.; Nissen, S.J.; Hufbauer, R.A. New techniques and findings in the study of a candidate allelochemical implicated in invasion success. Ecol. Lett. 2005, 8, 1039–1047. [Google Scholar] [CrossRef]
- Blair, A.C.; Nissen, S.J.; Brunk, G.R.; Hufbauer, R.A. A lack of evidence for an ecological role of the putative allelochemical (±)-catechin in Centaurea maculosa invasion process. J. Chem. Ecol. 2006, 32, 2327–2331. [Google Scholar] [CrossRef]
- Duke, S.O.; Blaie, A.C.; Dayan, F.E.; Johnson, R.D.; Meepagala, K.M.; Cook, D.; Bajsa, J. Is (-)-cathchin a novel weapon of Spooted Knapweed (Centaurea stoebe)? J. Chem. Ecol. 2009, 35, 141–153. [Google Scholar] [CrossRef]
- Tharayil, N.; Bhowmik, P.C.; Xing, B. Bioavailability of allelochemicals as affected by companion compounds in oil matrices. J. Agric. Food Chem. 2008, 56, 3706–3713. [Google Scholar] [CrossRef]
- Blair, A.C.; Weston, L.A.; Nissen, S.J.; Brunk, G.R.; Hufbauer, R. The importance of analytical techniques in allelopathy studies with the reported allelochemical catechin as an example. Biol. Invasions 2009, 11, 325–332. [Google Scholar] [CrossRef]
- Furubayashi, A.; Hiradate, S.; Fujii, Y. Role of catechol structure in the adsorption and transformation reactions of L-DOPA in soils. J. Chem. Ecol. 2007, 33, 239–250. [Google Scholar] [CrossRef]
- Inderjit; Pollock, J.; Callaway, R.M.; Hoben, W. Phytotoxic effects of (±)-catechin in vitro, in soil, and in the field. Plos One 2008, 3, 1–11. [Google Scholar]
- Vivanco, J.M.; Bais, H.P.; Stermitz, F.R.; Thelen, G.C.; Callaway, R.M. Biogeographical variation in community response to root allelochemistry: Novel weapons and exotic invasion. Ecol. Lett. 2004, 7, 285–292. [Google Scholar] [CrossRef]
- Norton, A.P.; Blair, A.C.; Hardin, J.G.; Nissen, S.J.; Brunk, G.R. Herbivory and novel weapons: No evidence for enhanced competitive ability or allelopathy induction of Centaurea diffusa by biological controls. Biol. Invasions 2008, 10, 79–88. [Google Scholar] [CrossRef]
- Romagni, J.G.; Allen, S.N.; Dayan, F.E. Allelopathic effects of volatile cineoles on two weedy plant species. J. Chem. Ecol. 2000, 26, 303–313. [Google Scholar] [CrossRef]
- Duke, S.O.; Dayan, F.E.; Hernández, A.; Duke, M.V.; Abbas, H.K. Natural products as leads for new herbicide modes of action. In In Proceedings Brighton Crop Protection Conference---Weeds, Brighton, UK, British Crop Protection Council, Brighton, UK; 17-20 November 1997; pp. 579–586. [Google Scholar]
- Rimando, A.M.; Olofsdotter, M.; Dayan, F.E.; Duke, S.O. Searching for Rice Allelochemicals. Agron. J. 1999, 93, 16–20. [Google Scholar]
- Nakano, H.; Fujii, Y.; Suzuki, T.; Yamada, K.; Kosemura, S.S.; Yamamura, S.; Suzuki, T.; Hasegawa, K. A growth-inhibitory substance exuded from freeze-dried mesquite (Prosopis juliflora (Sw.) DC.) leaves. Plant Growth Regul. 2001, 33, 165–168. [Google Scholar] [CrossRef]
- Iqbal, Z.; Hiradate, S.; Noda, A.; Fujii, Y. Allelopathic activity of buckwheat: Isolzation and characterization of phenolics. Weed Sci. 2003, 51, 657–662. [Google Scholar] [CrossRef]
- Shiraishi, S.; Watanabe, I.; Kuno, K.; Fujii, Y. Evaluation of the allelopathic activity of five Oxalidaceae cover plants and the demonstration of potent weed suppression by Oxalis species. Weed Biol. Manag. 2005, 5, 128–136. [Google Scholar] [CrossRef]
- Parvez, M.M.; Tomita-Yokotani, K.; Fujii, Y.; Konishi, T.; Iwashina, T. Effects of quercetin and its seven derivatives on the growth of Arabidopsis thaliana and Neurospora crassa. Biochem. Syst. Ecol. 2004, 32, 631–635. [Google Scholar] [CrossRef]
- Inderjit; Callaway, R.M. Experimantal designs for the study of allelopathy. Plant Soil. 2003, 256, 1–11. [Google Scholar] [CrossRef]
- Inderjit; Kaur, M.; Foy, C.L. On the significance of field studies in allelopathy. Weed Technol. 2001, 15, 792–797. [Google Scholar] [CrossRef]
- Ai Hamdi, B.; Inderjit; Olofsdotter, M.; Streibig, J.C. Laboratory bioassay for phytotoxicity: An example from wheat straw. Agron J. 2001, 93, 43–48. [Google Scholar]
- Yu, J.Q.; Matsui, Y. Effects of root exudates of cucumber (Cucumis sativus.) and allelochemicals on ion uptake by cucumber seedlings. J. Chem. Ecol. 1997, 23, 817–827. [Google Scholar] [CrossRef]
- Yu, J.Q.; Matsui, Y. Phytotoxic substances in root exudates of cucumber (Cucumis sativus L.). J. Chem. Ecol. 1994, 20, 21–31. [Google Scholar] [CrossRef]
- Ye, S.F.; Zhou, Y.H.; Sun, Y.; Zou, L.Y.; Yu, J.Q. Cinnamic acid causes oxidative stress in cucumber roots, and promotes incidence of Fusarium wilt. Environ. Exp. Bot. 2006, 56, 255–262. [Google Scholar] [CrossRef]
- Luo, X.; Pan, C.D.; Huang, M.M.; Liu, C.L. Autotoxicity of Picea schrenkiana litter aqueous extracts on seed germination and seedling growth. Xinjiang Agr. Sci. 2006, 43, 1–5. [Google Scholar]
- Wang, Q.; Ruan, X.; Pan, C.D;; Xu, N.Y.; Luo, X.; Huang, M.M. Need for sustainability policy—A case study of the National Forest Conservation Program (NFCP) in the western region of Tianshan Mountain, China. Forest. Chron. 2006, 82, 31–39. [Google Scholar]
- Appel, H.M. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol. 1993, 19, 1521–1552. [Google Scholar] [CrossRef]
- Blum, U. Allelopathic interactions involving phenolic acids. J. Nematol. 1996, 28, 259–267. [Google Scholar]
- Blum, U.; Dalton, B.R. Effects of ferulic acid, an allelopathic compound on leaf expansion of cucumber seedlings in nutrient culture. J. Chem. Ecol. 1985, 11, 279–301. [Google Scholar] [CrossRef]
- Blum, U.; Shafer, S.R. Microbial populations and phenolic acids in soil. Soil Biol. Biochem. 1988, 20, 793–800. [Google Scholar] [CrossRef]
- Blum, U. Effects of microbial utilization of phenolic acids and their phenolic acid breakdown products on allelopathic interactions. J. Chem. Ecol. 1998, 24, 685–708. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Z.-H.; Wang, Q.; Ruan, X.; Pan, C.-D.; Jiang, D.-A. Phenolics and Plant Allelopathy. Molecules 2010, 15, 8933-8952. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules15128933
Li Z-H, Wang Q, Ruan X, Pan C-D, Jiang D-A. Phenolics and Plant Allelopathy. Molecules. 2010; 15(12):8933-8952. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules15128933
Chicago/Turabian StyleLi, Zhao-Hui, Qiang Wang, Xiao Ruan, Cun-De Pan, and De-An Jiang. 2010. "Phenolics and Plant Allelopathy" Molecules 15, no. 12: 8933-8952. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules15128933