Effect of Rain-Shelter Cultivation of Vitis vinifera cv. Cabernet Gernischet on the Phenolic Profile of Berry Skins and the Incidence of Grape Diseases

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Rain Shelter on Incidence of Grape Diseases
2.2. Effect of Rain Shelter on Grape Berry Maturity
2.3. Effect of Rain Shelter on Phenolic Profile of Berry Skins

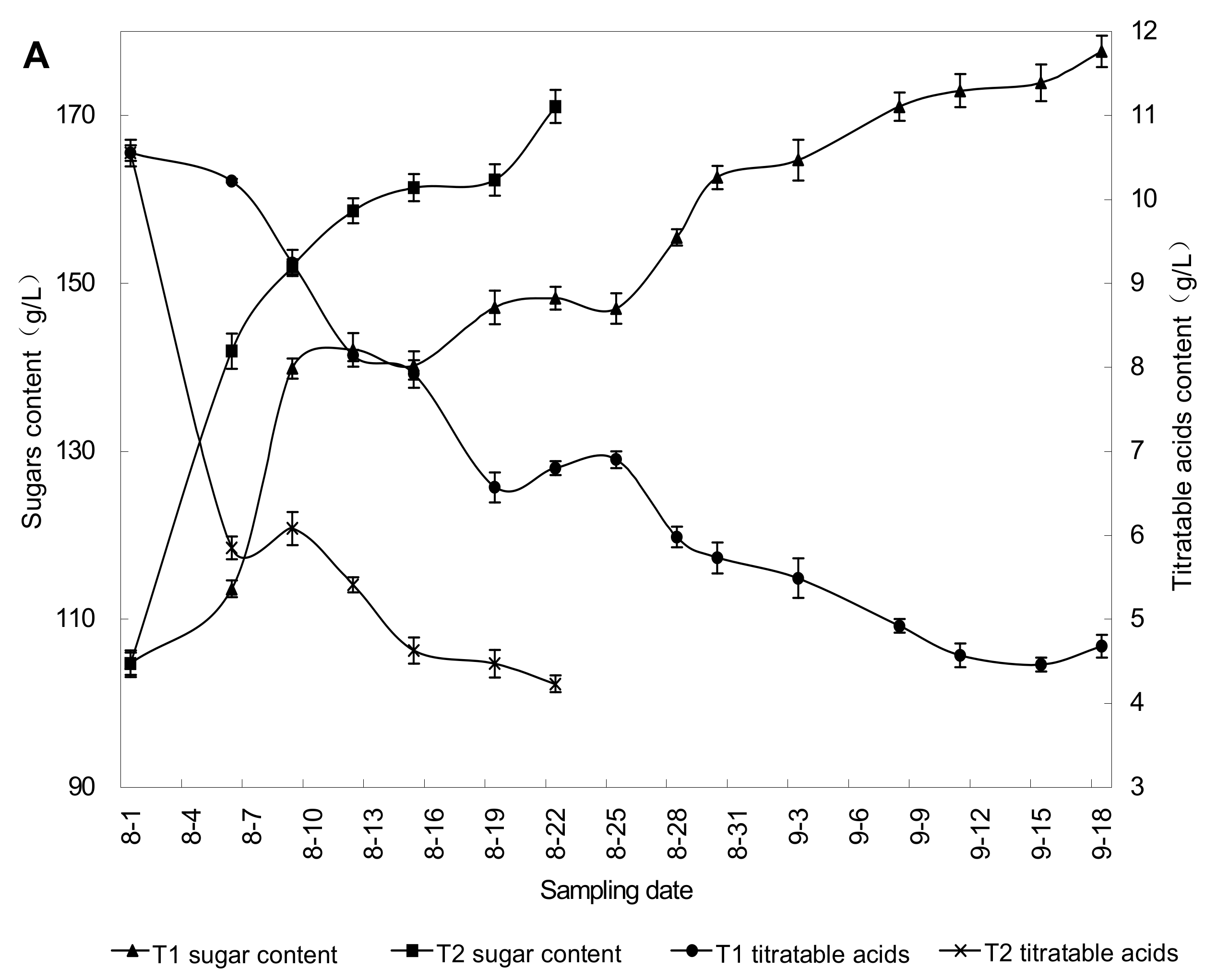{kind=link}
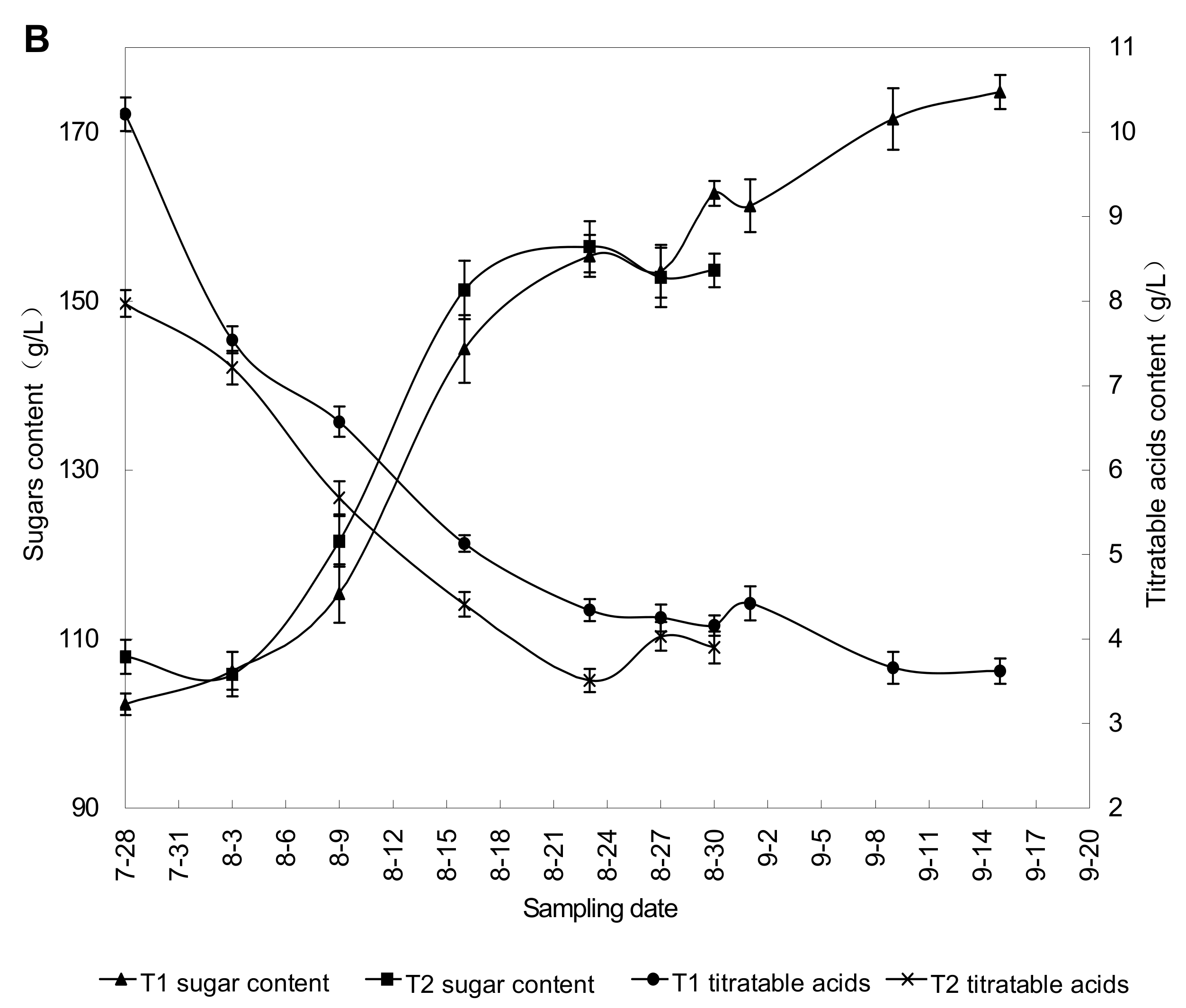{kind=link}
{kind=link}
| Phenolic compound | [M+]/[M-H]− (Frag. MS2 m/z) | Vintage 2010 | Vintage 2011 | ||
|---|---|---|---|---|---|
| T1 | T2 | T1 | T2 | ||
| Anthocyanins (mg ME/kg DW) | |||||
| Delphinidin-3-O-glucoside | 465 (303) | 686.03 ± 50.84 b | 1055.62 ± 85.37 a | 462.09 ± 40.27 b | 623.96 ± 59.64 a |
| Delphinidin-3-O-(6-O-acetyl)-glucoside | 507 (303) | 236.87 ± 20.67 b | 373.17 ± 15.84 a | nd | 152.18 ± 17.96 |
| Petunidin-3-O-glucoside | 479 (317) | 682.10 ± 73.49 b | 1126.87 ± 60.35 a | 523.99 ± 40.54 b | 664.54 ± 52.67 a |
| Petunidin-3-O-(6-O-acetyl)-glucoside | 521 (317) | 173.63 ± 21.44 b | 327.61 ± 33.72 a | 106.56 ± 17.33 b | 246.49 ± 20.67 a |
| Petunidin-3-O-(6-O-coumaroyl)-glucoside | 625 (317) | 336.26 ± 16.56 a | 52.27 ± 3.78 b | nd | nd |
| Cyanidin-3-O-glucoside | 449 (287) | 90.70 ± 7.45 a | 72.47 ± 8.30 b | 41.72 ± 5.34 a | 41.93 ± 3.33 a |
| Peonidin-3-O-glucoside | 463 (301) | 462.35 ± 12.84 a | 455.16 ± 20.35 a | 318.21 ± 25.84 a | 295.78 ± 16.47 ab |
| Peonidin-3-O-(6-O-acetyl)-glucoside | 505 (301) | 406.28 ± 51.35 b | 703.86 ± 47.84 a | 211.29 ± 14.67 a | 180.44 ± 15.45 b |
| Peonidin-3-O-(6-O-coumaroyl)-glucoside | 609 (301) | 50.14 ± 6.84 | nd | nd | nd |
| Malvidin-3-O-glucoside | 493 (331) | 5964.82 ± 231.42 b | 8975.48 ± 476.53 a | 6206.61 ± 366.67 b | 7444.34 ± 425.56 a |
| Malvidin-3-O-(6-O-acetyl)-glucoside | 535 (331) | 2937.81 ± 118.48 b | 5380.51 ± 234.84 a | 3174.80 ± 102.43 b | 3729.39 ± 229.33 a |
| Malvidin-3-O-(6-O-coumaroyl)-glucoside | 639 (331) | 5340.91 ± 346.76 b | 9299.04 ± 513.90 a | 4677.49 ± 194.33 a | 3800.57 ± 260.65 b |
| Malvidin-3-O-(6-O-caffeoyl)-glucoside | 655 (331) | nd | 673.65 ± 41.14 | 308.05 ± 37.24 b | 395.37 ± 29.56 a |
| Sum of anthocyanins | 17385.36 ± 1556.34 b | 28524.33 ± 3011.30 a | 16167.67 ± 837.67 b | 17575.00 ± 964.35 a | |
| Flavan-3-ols (mg CE/kg DW) | |||||
| (+)-Catechin | 289 (245, 205, 179) | 59.40 ± 4.89 b | 86.17 ± 10.84 a | 61.63 ± 6.31 a | 68.72 ± 4.54 a |
| (-)-Epicatechin | 289 (245, 205, 179) | 32.87 ± 3.92 b | 59.23 ± 4.76 a | 33.77 ± 2.67 b | 69.37 ± 5.23 a |
| Gallocatechin | 305 (179) | 61.32 ± 3.87 a | 35.39 ± 4.12 b | 15.61 ± 1.34 b | 17.41 ± 0.84 a |
| dimer(epi)gallocatechin-(epi)catechin | 593 (425) | 0.53 ± 0.08 b | 0.75 ± 0.10a | tr | 0.50 ± 0.10 |
| Procyanidin dimmer | 577 (425, 407, 289) | 217.77 ± 16.45 a | 225.56 ± 10.89 a | 117.62 ± 12.89 a | 125.35 ± 14.36 a |
| Procyanidin trimer | 865 (713, 577, 289) | 209.83 ± 12.35 b | 241.15 ± 16.45 a | nd | 142.09 ± 10.69 |
| Sum of flavan-3-ols | 581.72 ± 30.67 b | 648.23 ± 40.56 a | 228.61 ± 15.82 b | 423.44 ± 24.45 a | |
| Flavonols (mg QE/kg DW) | |||||
| isorhamnetin-3-O-glucoside | 477 (315) | 506.931 ± 36.48 a | 533.86 ± 41.22 a | nd | nd |
| isorhamnetin-3-O-galactoside | 477 (315) | 12.23 ± 1.45b | 25.72 ± 1.76a | nd | tr |
| Flavonols (mg QE/kg DW) | |||||
| quercetin-3-O-hexoside | 463 (301) | nd | 13.45 ± 0.97 | 32.82 ± 2.34 | nd |
| quercetin-3-O-glucuronide | 477 (301) | 147.67 ± 13.33 a | 136.18 ± 14.52 a | 86.05 ± 6.67 a | 119.15 ± 10.48 a |
| quercetin-3-O-galactoside | 463 (301) | 333.57 ± 29.35 a | 319.57 ± 20.67 a | 204.88 ± 19.59 b | 329.10 ± 18.84 a |
| quercetin-3-O-rutinoside | 609 (301) | 254.88 ± 13.33 b | 286.73 ± 19.56 a | 91.99 ± 11.48 b | 166.04 ± 14.57a |
| dihydroquercetin-3′-O-rhamnoside | 449 (303) | 207.60 ± 15.27 a | 125.78 ± 10.44 b | 84.63 ± 6.42 a | 60.49 ± 10.76 b |
| laricitrin-3-O-galactoside | 493 (331) | 92.11 ± 16.46 a | 47.99 ± 6.79 b | 23.27 ± 3.46 a | 23.90 ± 2.32 a |
| laricitrin-3-O-glucoside | 493 (331) | 164.39 ± 28.54 b | 241.15 ± 30.21 a | 72.54 ± 6.48 b | 100.59 ± 14.39 a |
| myricetin-3-O-galactoside | 479 (317) | 253.73 ± 11.25 b | 286.31 ± 13.47 a | 238.61 ± 20.36 a | 224.28 ± 17.54 a |
| kaempferol-3-O-galactoside | 447 (285) | 76.22 ± 7.26 b | 114.28 ± 10.44 a | 197.26 ± 26.48 b | 337.65 ± 36.48 a |
| Sum of flavonols | 2049.31 ± 94.48 b | 2131.01 ± 106.25 a | 1032.04 ± 84.29 b | 1361.19 ± 116.84 a | |
| Stilbenes (mg RE/kg DW) | |||||
| trans-resveratrol | 227 (185, 159) | 7.72 ± 0.76 b | 11.63 ± 1.27 a | 4.36 ± 0.48 b | 8.69 ± 1.45 a |
| trans-Piceid | 389 (227) | 5.64 ± 1.49 a | 7.12 ± 1.74 a | 4.21 ± 0.85 b | 7.96 ± 1.44 a |
| Sum of stilbenes | 13.36 ± 1.54 b | 18.75 ± 1.63 a | 8.57 ± 1.01 b | 16.65 ± 2.46 a | |
| Hydroxybenzoic acids (mg GAE/kg DW) | |||||
| hexose ester of vanillic acid | 329 (167) | 54.66 ± 8.48 b | 80.86 ± 9.38 a | 31.05 ± 1.56 a | 31.94 ± 3.00 a |
| Syringic acid | 197 (153) | 91.83 ± 11.23 a | 103.24 ± 16.19 a | 105.40 ± 20.58 a | 126.07 ± 29.34 a |
| hexose ester of protocatechuic acid | 315 (153) | 4.79 ± 0.52 a | 3.55 ± 0.38 a | 0.28 ± 0.05 | nd |
| Sum of hydroxybenzoic acids | 150.02 ± 10.38 b | 188.87 ± 21.67 a | 136.74 ± 18.49 a | 158.01 ± 20.21 a | |
| Hydroxycinnamic acids (mg CAE/kg DW) | |||||
| hexose ester of ferulic acid | 355 (193) | 2.16 ± 0.28 b | 4.41 ± 0.33 a | 3.13 ± 0.97 a | 4.08 ± 0.89 a |
| Sum of hydroxycinnamic acids | 2.16 ± 0.28 b | 4.41 ± 0.33 a | 3.13 ± 0.97 a | 4.08 ± 0.89 a | |
| Sum of flavonoids | 20016.39 ± 1996.38 ab | 31303.57 ± 2910.86 a | 17428.32 ± 1489.65 b | 19359.63 ± 1480.46 a | |
| Sum of nonflavonoids | 165.53 ± 17.43 b | 212.02 ± 10.67 a | 148.45 ± 20.37 ab | 178.74 ± 16.33 a | |
| Sum of phenolics | 20181.92 ±2053.76 ab | 31515.59 ± 3219.34 a | 17576.77 ± 1524.63 b | 19538..37 ± 1805.33 a | |
3. Experimental
3.1. Plant Materials
3.2. Chemicals and Standards
3.3. Determination of Some Basic Physicochemical Parameters
3.4. Diseases Investigation and Microclimate Evaluation
3.5. Extraction of Phenolic Compounds
3.6. HPLC-DAD/ESI-MS Analysis of Phenolic Compounds
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References
- OIV. Statistical report on World vitiviniculture 2012. Available online: http://www.oiv.int/oiv/files/0%20-%20Actualites/EN/Report.pdf (accessed on 1 January 2012).
- He, P.C. Ampeliology, 1st ed.; China Agriculture Press: Beijing, China, 1999; pp. 54–76. [Google Scholar]
- Morinaga, K. Grape Production in Japan. In Grape Production in the Asia-Pacific Region, 1st ed.; Papademetriou, M.K., Dent, F.J., Eds.; Regional Office for Asia and the Pacific, Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 2001; Available online: http://www.fao.org/docrep/003/x6897e/x6897e07.htm#6.%20GRAPE%20PRODUCTION%20IN%20JAPAN/ (accessed on 21 December 2012).
- Li, S.H. Grape Production in China. In Grape Production in the Asia-Pacific Region, 1st ed.; Papademetriou, M.K., Dent, F.J., Eds.; Regional Office for Asia and the Pacific, Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 2001; Available online: http://www.fao.org/docrep/003/x6897e/x6897e05.htm#4.%20GRAPE%20PRODUCTION%20IN%20CHINA/ (accessed on 21 December 2012).
- Sun, Q.B.; Wang, L.; Yu, F.F.; Sun, J.; Lu, L.J. Research Advances in grape rain-shelter cultivation and its matching technique. J. Anhui Agric. Sci. 2006, 34, 4560–4561. [Google Scholar]
- Detoni, A.M.; Clemente, E.; Fornari, C. Productivity and quality of grape “Cabernet Sauvignon” produced in organic sistem under plastic covering. Revieta Brasileira de fruticultura 2007, 29, 530–534. [Google Scholar] [CrossRef]
- Wang, X.J.; Xu, D.X.; Wang, X.Q.; Huang, W.D. The comparative study of rain-shelter on fruit quality of Cabernet Sauvignon. Chinese Agric. Sci. Bull. 2011, 27, 114–118. [Google Scholar]
- Garrido, J.; Borges, F. Wine and grape polyphenols––A chemical perspective. Food Res. Int. 2011, 44, 3134–3148. [Google Scholar] [CrossRef]
- Kennedy, J.A. Grape and wine phenolics: Observations and recent findings. Ciencia e Investigación Agraria 2008, 35, 107–120. [Google Scholar] [CrossRef]
- Cynkar, W.; Cozzolino, D.; Dambergs, R.G. The effect of sample storage and homogenisation techniques on the chemical composition and near infrared spectra of white grapes. Food Res. Int. 2009, 6, 100–110. [Google Scholar] [CrossRef]
- Elías, O.S.; Álvaro, P.N.; Remigio, L.S.; Fernando, Z.M.; Jorge, M.R.S.; Olga, L. Comparative study of the phenolic composition of seeds and skins from Carménère and Cabernet Sauvignon grape varieties (Vitis vinifera L.) during ripening. J. Agric. Food Chem. 2010, 58, 3591–3599. [Google Scholar]
- Meng, J.F.; Fang, Y.L.; Qin, M.Y.; Zhuang, X.F.; Zhang, Z.W. Varietal differences among the phenolic profiles and antioxidant properties of four cultivars of spine grape (Vitis davidii Foex) in Chongyi County (China). Food Chem. 2012, 134, 2049–2056. [Google Scholar] [CrossRef] [PubMed]
- Montealegre, R.R.; Peces, R.R.; Vozmediano, J.L.C.; Gascueña, J.M.; Romero, E.G. Phenolic compounds in skins and seeds of ten grape Vitis vinifera varieties grown in a warm climate. J. Food Compost. Anal. 2006, 19, 687–693. [Google Scholar] [CrossRef]
- Puertolas, E.; Hernandez-Orte, P.; Sladana, G.; Alvarez, I.; Raso, J. Improvement of winemaking process using pulsed electric fields at pilot-plant scale. Evolution of chromatic parameters and phenolic content of Cabernet Sauvignon red wines. Food Res. Int. 2010, 43, 761–766. [Google Scholar] [CrossRef]
- Meng, J.F.; Xu, T.F.; Qin, M.Y.; Zhuang, X.F.; Fang, Y.L.; Zhang, Z.W. Phenolic characterization of young wines made from spine grape (Vitis davidii Foex) grown in Chongyi County (China). Food Res. Int. 2012, 49, 664–671. [Google Scholar] [CrossRef]
- Gladstones, J.S. Climate and Australian viticulture. In Viticulture, 2nd ed.; Dry, P.R., Coombe, B.G., Eds.; Winetitles Pty Ltd: Ashford, Australia, 2004; Volume 1, pp. 90–118. [Google Scholar]
- Considine, J.A.; Kriedemann, P.E. Fruit splitting in grapes: Determination of the critical turgor pressure. Aust. J. Agric. Res. 1972, 23, 17–24. [Google Scholar] [CrossRef]
- Chavarria, G.; Cardoso, L.S.; Bergamaschi, H.; dos Santos, H.P.; Mandelli, F.; Marodin, G.A.B. Microclimate of vineyards under protected cultivation. Ciência Rural 2009, 39, 2029–2034. [Google Scholar] [CrossRef]
- Cardoso, L.S.; Bergamaschi, H.; Comiran, F.; Chavarria, G.; Marodin, G.A.B.; Dalmago, G.A.; dos Santos, H.P.; Mandelli, F. Micrometeorological alterations in vineyards by using plastic covering. Pesq. agropec. bras. 2008, 43, 441–444. [Google Scholar] [CrossRef]
- Rana, G.; Katerji, N.; Introna, M.; Hammami, A. Microclimate and plant water relationship of the “overhead” table grape vineyard managed with three different covering techniques. Sci. Hortic. 2004, 102, 105–120. [Google Scholar] [CrossRef]
- Chavarria, G.; dos Santos, H.P.; Zanus, M.C.; Marodin, G.A.B.; Zorzan, C. Plastic cover use and its influences on physical-chemical characteristics in must and wine. Rev. Bras. Frutic. 2011, 33, 809–815. [Google Scholar] [CrossRef]
- Bergqvist, J.; Dokoozlian, N.; Ebisuda, N. Sunlight exposure and temperature effects on berry growth and composition of Cabernet Sauvignon and Grenache in the central San Joaquin valley of California. Am. J. Enol. Vitic. 2001, 52, 1–7. [Google Scholar]
- Gawel, R. Red wine astringency: A review. Aust. J. Grape Wine Res. 1998, 4, 74–95. [Google Scholar] [CrossRef]
- Hufnagel, J.C.; Hofmann, T. Orosensory-directed identification of astringent mouthfeel and bitter-tasting compounds in red wine. J. Agric. Food Chem. 2008, 56, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Obreque-Slier, E.; Peña-Neira, Á.; López-Solís, R.; Zamora-Marín, F.; Ricardo-da Silva, J.M.; Laureano, O. omparative study of the phenolic composition of seeds and skins from Carménère and Cabernet Sauvignon grape varieties (Vitis vinifera L.) during ripening. J. Agric. Food Chem. 2010, 58, 3591–3599. [Google Scholar] [CrossRef] [PubMed]
- Ferrandino, A.; Guidoni, S. Anthocyanins, xavonols and hydroxycinnamates: An attempt to use them to discriminate Vitis vinifera L. cv “Barbera” clones. Eur. Food Res. Technol. 2010, 230, 417–427. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, Y.; Lu, J. Phenolic contents and compositions in skins of red Wine grape cultivars among various genetic backgrounds and originations. Int. J. Mol. Sci. 2012, 13, 3492–3510. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.-M.; He, J.-J.; Bi, H.-Q.; Cui, X.-Y.; Duan, C.-Q. Phenolic Compound Profiles in Berry Skins from Nine Red Wine Grape Cultivars in Northwest China. Molecules 2009, 14, 4922–4935. [Google Scholar] [CrossRef] [PubMed]
- González-Manzano, S.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Extraction of flavan-3-ols from grape seed and skin into wine using simulated maceration. Anal. Chim. Acta 2004, 513, 283–289. [Google Scholar] [CrossRef]
- Cortell, J.M.; Kennedy, J.A. Effect of shading on accumulation of flavonoid compounds in (Vitis Vinifera L.) pinot noir fruit and extraction in a model system. J. Agric. Food Chem. 2006, 54, 8510–8520. [Google Scholar] [CrossRef] [PubMed]
- Price, S.F.; Breen, P.J.; Valladao, M.; Watson, B.T. Cluster sun exposure and quercetin in Pinot-noir grapes and wine. Am. J. Enol. Viticul. 1995, 46, 187–194. [Google Scholar]
- Bogs, J.; Downey, M.; Harvey, J.; Ashton, A.; Tanner, G.; Robinson, S. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Boulton, R. The copigmentation of anthocyanins and its role in the color of red wine: A critical review. Am. J. Enol. Viticul. 2001, 52, 67–87. [Google Scholar]
- Saiko, P.; Szakmary, A.; Jaeger, W.; Szekeres, T. Resveratrol and its analogs: Defense against cancer, coronary disease and neurodegenerative maladies or just a fad? Mutat. Res. 2008, 658, 68–94. [Google Scholar] [CrossRef] [PubMed]
- Vitrac, X.; Monti, J.P.; Vercauteren, J.; Deffieux, G.; Mérillon, J.M. Direct liquid chromatographic analysis of resveratrol derivatives and flavanonols in wines with absorbance and fluorescence detection. Anal. Chim. Acta 2002, 458, 103–110. [Google Scholar] [CrossRef]
- China National Standardization Administration. Ampeliology Analytical Methods of Wine and Fruit Wine, GB/T 15038; China Standards Press: Beijing, China, 2006; pp. 2–23. [Google Scholar]
- Li, Z.; Pan, Q.H.; Jin, Z.M.; Mu, L.; Duan, C.Q. Comparison on phenolic compounds in Vitis vinifera cv. Cabernet Sauvignon wines from five wine-growing regions in China. Food Chem. 2011, 125, 77–83. [Google Scholar] [CrossRef]
- Fanzone, M.; Zamora, F.; Jofré, V.; Assof, M.; Gómez-Cordovés, C.; Peña-Neira, Á. Phenolic characterisation of red wines from different grape varieties cultivated in Mendoza province (Argentina). J. Sci. Food Agric. 2011, 92, 704–718. [Google Scholar] [CrossRef] [PubMed]
- Han, F.-L.; Zhang, W.-N.; Pan, Q.-H.; Zheng, C.-R.; Chen, H.-Y.; Duan, C.-Q. Principal component regression analysis of the relation between cielab color and monomeric anthocyanins in young Cabernet Sauvignon wines. Molecules 2008, 13, 2859–2870. [Google Scholar] [CrossRef] [PubMed]
- Peña-Neira, A.; Cáceres, A.; Pastenes, C. Low molecular weight phenolic and anthocyanin composition of grape skins from cv. Syrah (Vitis vinifera L.) in the Maipo Valley (Chile): Effect of clusters thinning and vineyard yield. Food Sci. Technol. Int. 2007, 13, 153–158. [Google Scholar] [CrossRef]
- Villiers, D.; Vanhonacker, G.; Majek, P.; Sandra, P. Determination of anthocyanins in wine by direct injection liquid chromatography-diode array detection-mass spectrometry and classification of wines using discriminant analysis. J. Chromatogr. A 2004, 1054, 195–204. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, Y.; Deng, J.; Li, H.; Lu, J. Phenolic concentrations and antioxidant properties of wines made from North American grapes grown in China. Molecules 2012, 17, 3304–3323. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of grape berries are available from the authors. |



| Disease parameters | August 6 | August 22 | September 15 | |||
|---|---|---|---|---|---|---|
| T1 | T2 | T1 | T2 | T1 | T2 | |
| Leaves | ||||||
| Diseases incidence of leaves (%) | 11.03 ± 4.56 a | 8.13 ± 3.27 a | 33.98 ± 3.62 b | 64.36 ± 7.67 a | 34.19 ± 7.84 b | 87.93 ± 5.39 a |
| Defoliation rates (%) | — | — | — | — | 2.05 ± 0.57 b | 41.92 ± 6.81 a |
| Diseases index | 3.73 ± 0.58 a | 1.97 ± 0.34 b | 6.91 ± 0.84 b | 38.94 ± 2.38 a | 9.48 ± 2.84 b | 71.48 ± 6.88 a |
| Fruits | ||||||
| Diseases incidence of cluster (%) | 23.33 ± 2.14 b | 43.33 ± 3.47 a | 58.33 ± 6.12 b | 85.00 ± 6.33 a | 66.66 ± 3.27 b | 90.27 ± 5.56 a |
| Diseases incidence of berry (%) | 2.07 ± 0.46 a | 1.93 ± 0.32 a | 9.50 ± 1.64 b | 49.43 ± 4.26 a | 10.50 ± 1.38 b | 67.98 ± 4.84 a |
| Diseases index | 5.48 ± 0.46 a | 7.62 ± 0.94 a | 18.81 ± 2.56 b | 61.67 ± 4.55 a | 22.14 ± 1.89 b | 84.43 ± 3.22 a |
| Disease parameters | August 9 | August 23 | September 15 | |||
|---|---|---|---|---|---|---|
| T1 | T2 | T1 | T2 | T1 | T2 | |
| Leaves | ||||||
| Diseases incidence of leaves (%) | 54.22 ± 3.29 b | 85.30 ± 7.49 a | 51.10 ± 6.38 b | 87.29 ± 4.56 a | 25.42 ± 4.56 b | 92.04 ± 6.39 a |
| Defoliation rates (%) | 1.35 ± 0.56 a | 2.68 ± 0.71 a | 4.39 ± 0.32 b | 15.60 ± 1.26 a | 8.12 ± 0.47 b | 25.95 ± 1.67 a |
| Diseases index | 8.13 ± 0.63 b | 13.55 ± 1.56 a | 7.97 ± 0.78 b | 19.74 ± 2.56 a | 4.41 ± 0.46 b | 25.82 ± 2.25 a |
| Fruits | ||||||
| Diseases incidence of cluster (%) | 16.67 ± 1.22 b | 28.33 ± 3.24 a | 21.67 ± 2.18 b | 35.00 ± 3.46 a | 53.33 ± 6.25 b | 84.00 ± 5.21 a |
| Diseases incidence of berry (%) | 0.65 ± 0.10 b | 1.70 ± 0.21 a | 1.05 ± 0.12 b | 4.75 ± 0.64 a | 3.97 ± 0.43 b | 35.00 ± 4.29 a |
| Diseases index | 2.62 ± 0.27 b | 5.71 ± 0.67 a | 2.86 ± 0.22 b | 9.29 ± 1.38 a | 14.05 ± 1.22 b | 78.00 ± 6.27 a |
| Microclimatic variables | Vintage 2010 | Vintage 2011 | ||
|---|---|---|---|---|
| T | C | T | C | |
| Average temperature in the canopy (°C) | 27.62 | 26.49 | 26.97 | 25.64 |
| Relative humidity in the canopy (%) | 70.12 | 68.26 | 69.45 | 68.00 |
| Illumination intensity in the canopy (×105 lx) | 0.046 | 0.130 | 0.057 | 0.169 |
| Parameter | April | May | June | July | August | September |
|---|---|---|---|---|---|---|
| Vintage 2010 | ||||||
| Average temperature (°C) | 13.2 | 19.9 | 25.4 | 27.2 | 24.7 | 21.3 |
| Rainfall (mm) | 41.0 | 42.7 | 23.8 | 77.0 | 145.1 | 85.9 |
| Sunshine hours | 170.0 | 150.5 | 219.6 | 158.0 | 145.4 | 152.1 |
| Vintage 2011 | ||||||
| Average temperature (°C) | 16.5 | 19.4 | 25.5 | 26.5 | 24.1 | 18.3 |
| Rainfall (mm) | 15.2 | 83.5 | 25.7 | 50.3 | 87.2 | 321.1 |
| Sunshine hours | 224.6 | 197.5 | 200.5 | 183.1 | 189.1 | 94.7 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Meng, J.-F.; Ning, P.-F.; Xu, T.-F.; Zhang, Z.-W. Effect of Rain-Shelter Cultivation of Vitis vinifera cv. Cabernet Gernischet on the Phenolic Profile of Berry Skins and the Incidence of Grape Diseases. Molecules 2013, 18, 381-397. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules18010381
Meng J-F, Ning P-F, Xu T-F, Zhang Z-W. Effect of Rain-Shelter Cultivation of Vitis vinifera cv. Cabernet Gernischet on the Phenolic Profile of Berry Skins and the Incidence of Grape Diseases. Molecules. 2013; 18(1):381-397. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules18010381
Chicago/Turabian StyleMeng, Jiang-Fei, Peng-Fei Ning, Teng-Fei Xu, and Zhen-Wen Zhang. 2013. "Effect of Rain-Shelter Cultivation of Vitis vinifera cv. Cabernet Gernischet on the Phenolic Profile of Berry Skins and the Incidence of Grape Diseases" Molecules 18, no. 1: 381-397. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules18010381