
The Roles of Direct Recognition by Animal Lectins in Antiviral Immunity and Viral Pathogenesis

Abstract
:1. Introduction

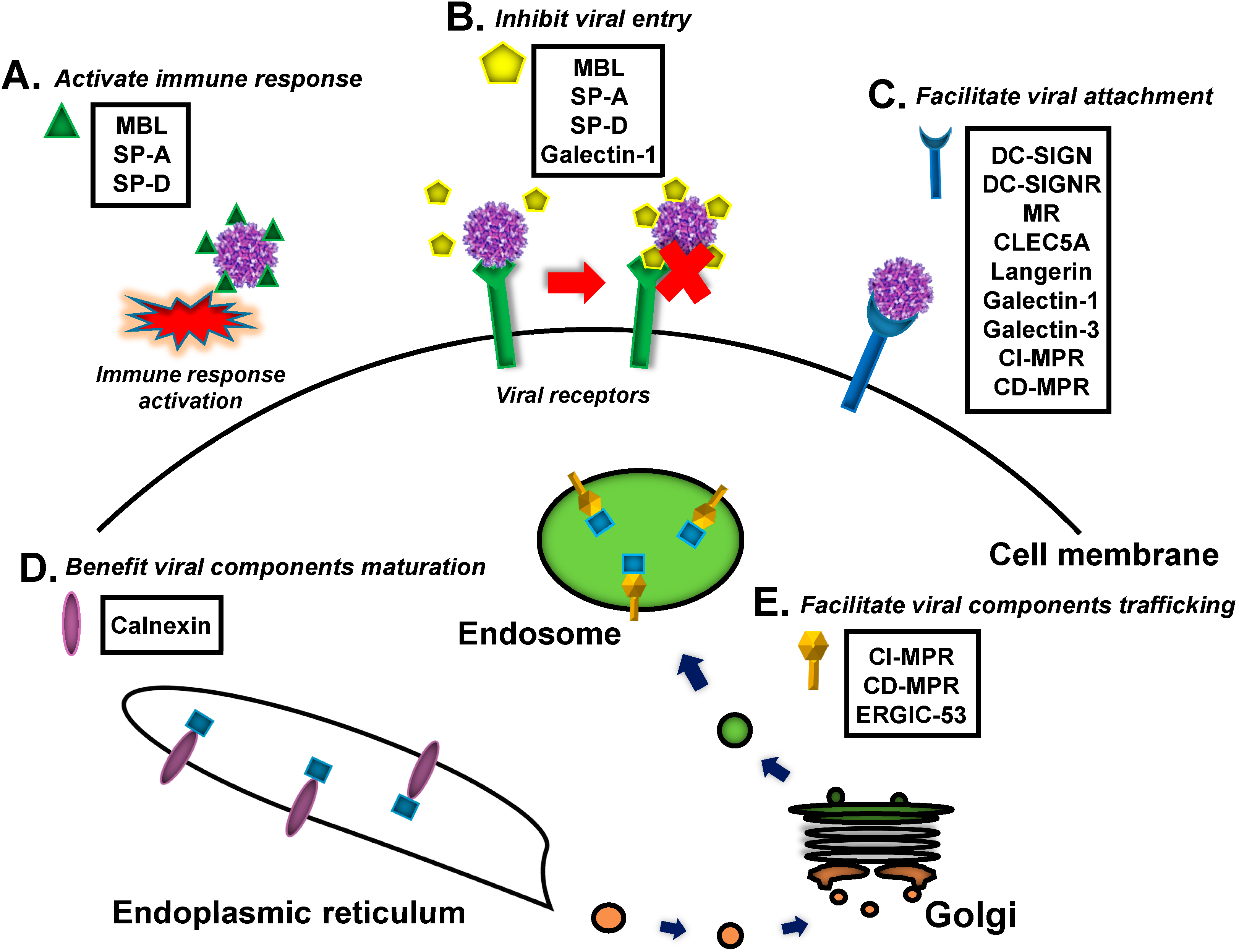{kind=link}
{kind=link}
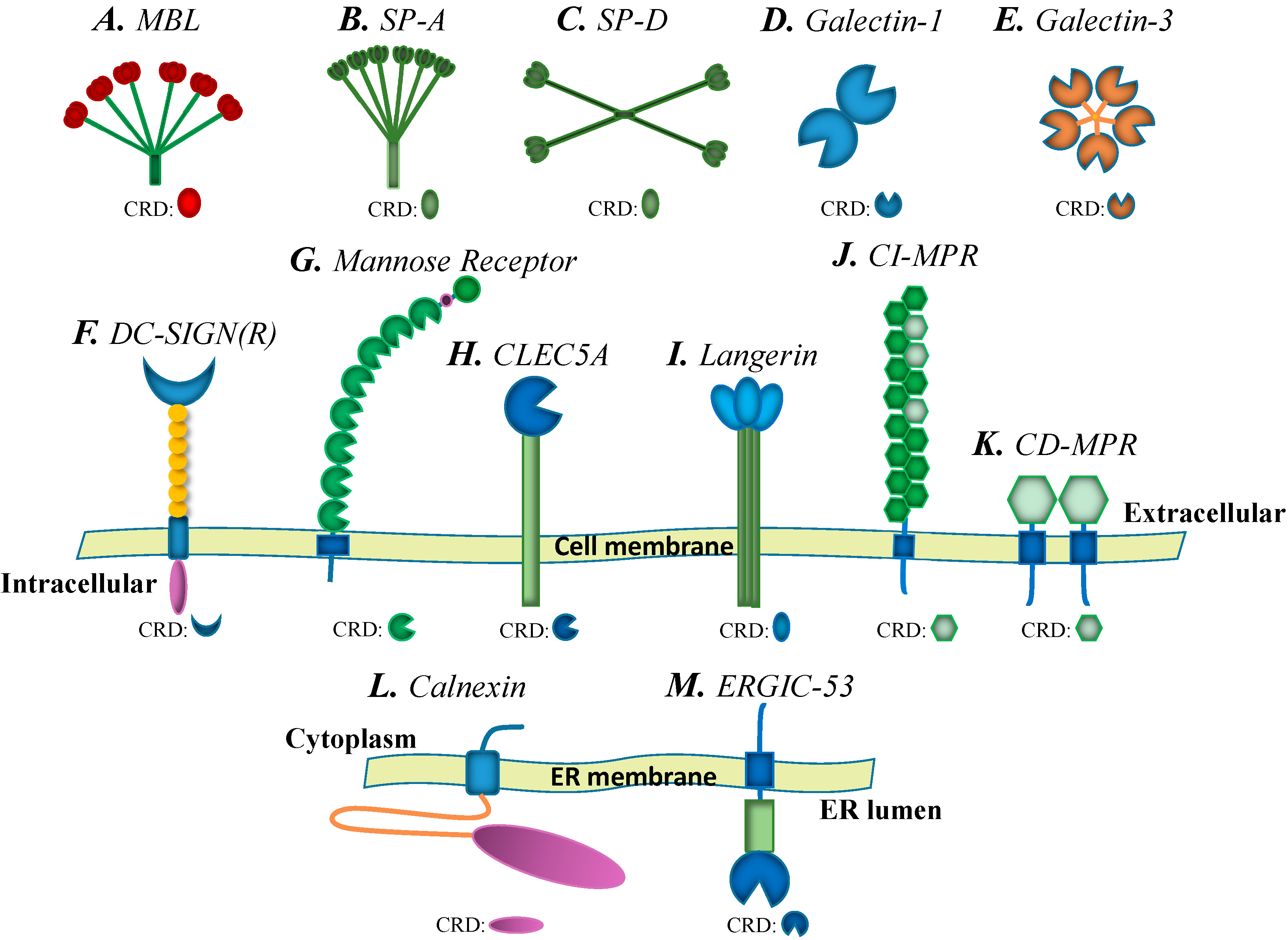{kind=link}
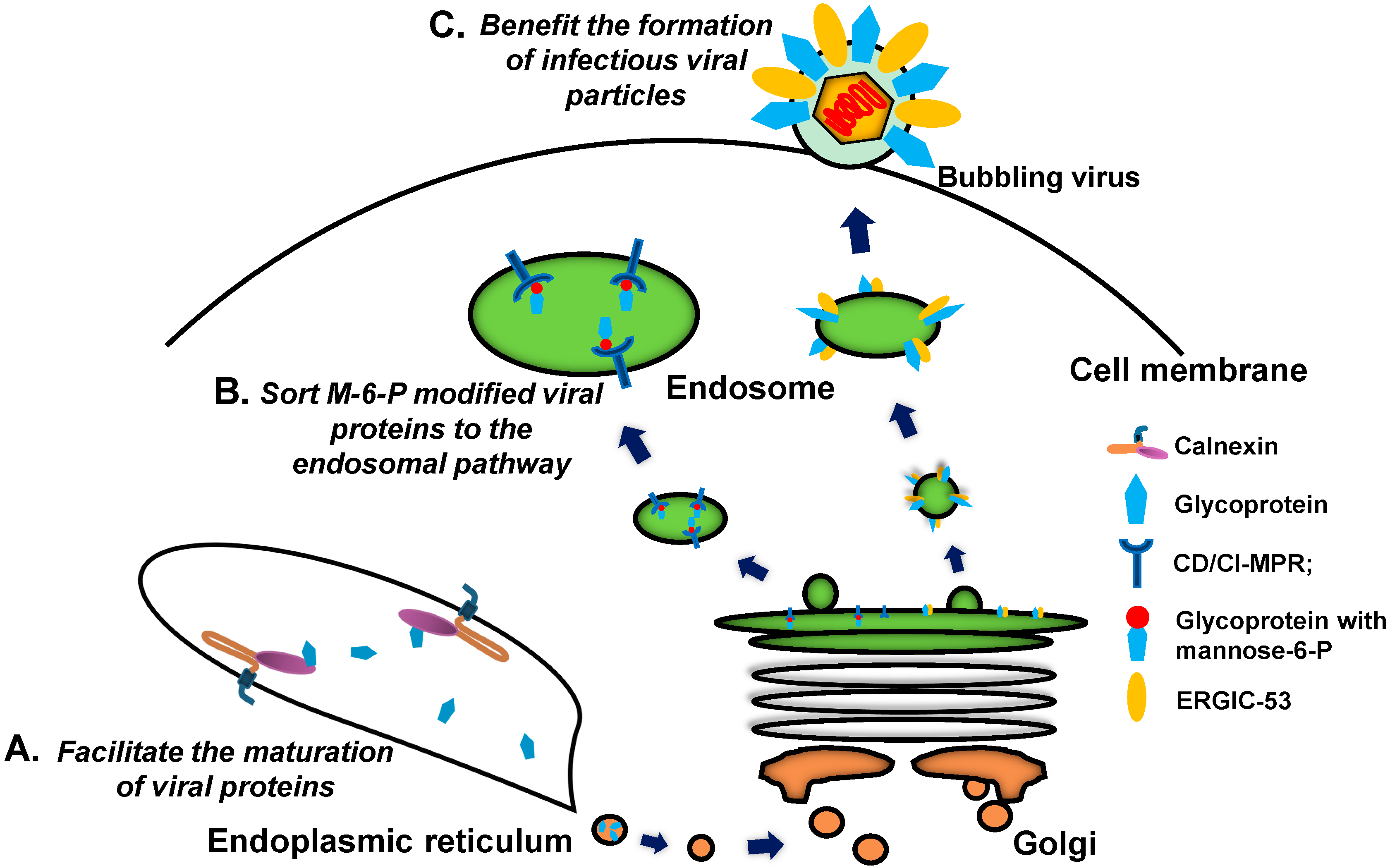{kind=link}
| Lectin | Saccharide Specificity | Core motif | Location | Direct Interaction to Viral Components |
|---|---|---|---|---|
| C-type lectin | Variable | C-type sequence motif | Extracellular, Cell membrane | Yes |
| Galectin | β-Galactosides | S-type sequence motif | Extracellular, Cytoplasm | Yes |
| Calnexin | Glc1Man9 | Calnexin sequence motif | ER | Yes |
| P-type lectin | Mannose-6-P, others | P-type sequence motif | Cell membrane, Endosome | Yes |
| L-type lectin | Variable | L-type sequence motif | ER, ERGIC, Golgi | Yes |
| I-type lectin | Sialic acid, variable | Ig-like domains | Cell membrane | No |
| M-type lectin | Man8 | M-type sequence motif | ER | No |
| F-type lectin | L-fucose | F-type sequence motif | Extracellular | No |
| R-type lectin | Variable | R-type sequence motif | Extracellular, Cell membrane | No |
| F-box lectin | GlcNAc2 | F-box sequence motif | Cytoplasm | No |
| Chitinase-like lectin | Chito-oligosaccharides | TIM (Triose-phosphate isomerase) barrel-like structure | Extracellular | No |
| Intelectin | Gal, glactofuranose, pentoses | Intelectin sequence motif | Extracellular, Cell membrane | No |

2. Viral Recognition by Mammalian Lectins
2.1. C-Type Lectins

2.1.1. Mannose-Binding Lectin (MBL)

2.1.2. Surfactant Proteins (SPs)
2.1.3. DC-SIGN, L-SIGN and the Mannose Receptor (MR)
2.1.4. C-Type Lectin Domain Family 5, Member A (CLEC5A)
2.1.5. Langerin
2.2. Galectins
2.3. Calnexin Family

2.4. P-Type Lectins/Mannose 6-Phosphate Receptors (MPRs)
2.5. L-Type Lectins
3. Arthropod Lectins and Viral Infections
4. Anti-Viral Drug and Vaccine Targeting of Lectins
| Species | Lectin Family | Name | Location | Virus | Target Protein | Function | Reference |
|---|---|---|---|---|---|---|---|
| Mammal | C-type lectins | MBL | Extracellular | HIV | gp120 | Inhibit viral infection | [21] |
| HBV | HBsAg | Inhibit viral infection | [22,23] | ||||
| HCV | Envelope glycoproteins | Inhibit viral infection | [24] | ||||
| WNV | E protein | Inhibit viral infection | [25] | ||||
| DENV | E protein | Inhibit viral infection | [25] | ||||
| Ebola virus | Envelope glycoproteins | Inhibit viral infection | [34] | ||||
| MARV | Envelope glycoproteins | Inhibit viral infection | [34] | ||||
| SP-A | Extracellular | IAV | HA and NA | Inhibit viral infection | [38] | ||
| RSV | F and G protein | Inhibit viral infection | [46,47] | ||||
| SP-D | Extracellular | IAV | HA | Inhibit viral infection | [39,40,41,42,43] | ||
| RSV | G protein | Inhibit viral infection | [48] | ||||
| DC-SIGN(R) | Transmembrane | HIV | gp120 | Facilitate viral infection | [28] | ||
| CMV | Envelope glycoproteins | Facilitate viral infection | [30] | ||||
| DENV | E protein | Facilitate viral infection | [32] | ||||
| WNV | E or prM protein | Facilitate viral infection | [52,53] | ||||
| MARV | GPs | Facilitate viral infection | [54] | ||||
| HCV | E2 protein | Facilitate viral infection | [57,58] | ||||
| Ebola virus | GP1 subunit | Facilitate viral infection | [29] | ||||
| SARS-CoV | S protein | Facilitate viral infection | [54,55,56] | ||||
| MR | Transmembrane | DENV | E protein | Facilitate viral infection | [61] | ||
| HBV | HBsAg | Facilitate viral infection | [62] | ||||
| IAV | HA or NA | Facilitate viral infection | [63,64] | ||||
| HIV | gp120 | Facilitate viral infection | [65] | ||||
| CLEC5A | Transmembrane | DENV | E protein | Facilitate viral infection | [67,68] | ||
| JEV | E protein | Facilitate viral infection | [69] | ||||
| Langerin | Transmembrane | HIV | gp120 | Facilitate viral infection | [71,72] | ||
| HIV | gp120 | Inhibit viral infection | [73] | ||||
| MV | F and H protein | Facilitate viral infection | [74] | ||||
| Mammal | Galectins | Galectin-1 | Secrete/Membrane | NiV | F proteins | Inhibit viral infection | [84] |
| IAV | Envelope glycoproteins | Inhibit viral infection | [85] | ||||
| HIV | gp120 | Facilitate viral infection | [86,87,88,89] | ||||
| Galectin-3 | Secrete/Membrane | HSV | Envelope glycoproteins | Facilitate viral infection | [90] | ||
| MVM | Capsid protein | Facilitate viral infection | [91] | ||||
| Calnexins | Calnexin/calreticulin | Intracellular | SeV | F and HN protein | Facilitate viral infection | [98] | |
| UUKV | G1/G2 protein | Facilitate viral infection | [99] | ||||
| SARS-CoV | S protein | Facilitate viral infection | [100] | ||||
| HBV | M protein | Facilitate viral infection | [101] | ||||
| HIV | gp160 and gp120 | Facilitate viral infection | [102,103] | ||||
| Rotavirus | NSP4 | Facilitate viral infection | [104,105,106] | ||||
| Hantavirus | Gn and Gc | Facilitate viral infection | [107] | ||||
| P-type lectins/MPRs | CI/CD-MPR | Transmembrane | HSV | gD | Facilitate viral infection | [109,110] | |
| VZV | Envelope glycoproteins | Facilitate viral infection | [111] | ||||
| Intracellular | HSV | gD | Facilitate viral infection | [110] | |||
| VZV | gB, gE, gH and gI | Facilitate viral infection | [111] | ||||
| L-type lectins | ERGIC-53 | Intracellular | Arenavirus, Hantavirus, Coronavirus, Filovirus, Orthomyxovirus | GPs | Facilitate viral infection | [114] | |
| Arthropod | C-type lectins | mosGCTLs | Extracellular | WNV | E protein | Facilitate viral infection | [127] |
| DENV | E protein | Facilitate viral infection | [122] | ||||
| MjsvCL | Extracellular | WSSV | Protein 28 | Facilitate viral infection | [128] |
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lakhtin, V.M. Lectins for investigation of proteins and carbohydrates. In Reviews of Science and Technique, Series Biotechnology; Klyosov, A.A., Ed.; VINITI: Moscow, Russia, 1987; Volume 2, p. 290. [Google Scholar]
- Sharon, N.; Lis, H. Molecular structure. In Lectins, 2nd ed.; Wong, C.H., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; Volume 1, p. 450. [Google Scholar]
- Gupta, G.S.; Gupta, A.; Gupta, R.K. Lectins: An Overview. In Animal Lectins: Forms, Functions and Clinical Applications; Gupta, G.S., Ed.; Springer: New York, NY, USA, 2012; Volume 1, p. 1108. [Google Scholar]
- Gabius, H.J. Animal lectins. Eur. J. Biochem. 1997, 243, 543–576. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, D.C. Animal lectins: A historical introduction and overview. Biochim. Biophys. Acta 2002, 1572, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K. Demonstration of carbohydrate recognition activity in diverse proteins which share a common primary structure motif. Biochem. Soc. Trans. 1989, 17, 13–15. [Google Scholar] [PubMed]
- Drickamer, K. Two distinct classes of carbohydrate-recognition domains in animal lectins. J. Biol. Chem. 1988, 263, 9557–9560. [Google Scholar] [PubMed]
- Drickamer, K.; Dordal, M.S.; Reynolds, L. Mannose-binding proteins isolated from rat liver contain carbohydrate-recognition domains linked to collagenous tails. Complete primary structures and homology with pulmonary surfactant apoprotein. J. Biol. Chem. 1986, 261, 6878–6887. [Google Scholar] [PubMed]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef] [PubMed]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Iobst, S.T.; Drickamer, K. Binding of sugar ligands to Ca(2+)-dependent animal lectins. II. Generation of high-affinity galactose binding by site-directed mutagenesis. J. Biol. Chem. 1994, 269, 15512–15519. [Google Scholar] [PubMed]
- Haurum, J.S.; Thiel, S.; Haagsman, H.P.; Laursen, S.B.; Larsen, B.; Jensenius, J.C. Studies on the carbohydrate-binding characteristics of human pulmonary surfactant-associated protein A and comparison with two other collectins: Mannan-binding protein and conglutinin. Biochem. J. 1993, 293, 873–878. [Google Scholar] [PubMed]
- Haagsman, H.P.; Hawgood, S.; Sargeant, T.; Buckley, D.; White, R.T.; Drickamer, K.; Benson, B.J. The major lung surfactant protein, SP 28–36, is a calciumdependent, carbohydrate-binding protein. J. Biol. Chem. 1987, 262, 13877–13880. [Google Scholar] [PubMed]
- Clemmensen, I. Interaction of tetranectin with sulphated polysaccharides and trypan blue. Scand. J. Clin. Lab. Investig. 1989, 49, 719–725. [Google Scholar] [CrossRef]
- Elgavish, S.; Shaanan, B. Lectin-carbohydrate interactions: Different folds, common recognition principles. Trends Biochem. Sci. 1997, 22, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Kolatkar, A.R.; Leung, A.K.; Isecke, R.; Brossmer, R.; Drickamer, K.; Weis, W.I. Mechanism of N-acetylgalactosamine binding to a C-type animal lectin carbohydrate-recognition domain. J. Biol. Chem. 1998, 273, 19502–19508. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, H.; Torgersen, D.; Drickamer, K.; Weis, W.I. Mechanism of pH-dependent N-acetylgalactosamine binding by a functional mimic of the hepatocyte asialoglycoprotein receptor. J. Biol. Chem. 2000, 275, 35176–35184. [Google Scholar] [CrossRef] [PubMed]
- Sheriff, S.; Chang, C.Y.; Ezekowitz, R.A. Human mannose-binding protein carbohydrate recognition domain trimerizes through a triple alpha-helical coiled-coil. Nat. Struct. Biol. 1994, 1, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Holmskov, U.; Thiel, S.; Jensenius, J.C. Collections and ficolins: Humoral lectins of the innate immune defense. Annu. Rev. Immunol. 2003, 21, 547–578. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.B.; Wallis, R. Two mechanisms for mannose-binding protein modulation of the activity of its associated serine proteases. J. Biol. Chem. 2004, 279, 260058–26065. [Google Scholar]
- Ying, H.; Ji, X.; Hart, M.L.; Gupta, K.; Saifuddin, M.; Zariffard, M.R.; Spear, G.T. Interaction of mannose-binding lectin with HIV-1 is sufficient for virus opsonization but not neutralization. AIDS Res. Hum. Retrovir. 2004, 20, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Filho, R.M.; Carmo, R.F.; Catsman, C.; Souza, C.; Silva, A.; Moura, P.; Tenorio, A.L.; Vasconcelos, L.R.; Cavalcanti Mdo, S.; Pereira, L.M. High frequency of variant alleles of the mannose-binding lectin 2 (MBL2) gene are associated with patients infected by hepatitis B virus. Viral Immunol. 2010, 23, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Chong, W.P.; To, Y.F.; Ip, W.K.; Yuen, M.F.; Poon, T.P.; Wong, W.H.; Lai, C.L.; Lau, Y.L.; Curtis, B.M. Mannose-binding lectin in chronic hepatitis B virus infection. Hepatology 2005, 42, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Koutsounaki, E.; Goulielmos, G.N.; Koulentaki, M.; Choulaki, C.; Kouroumalis, E.; Galanakis, E. Mannose-binding lectin MBL2 gene polymorphisms and outcome of hepatitis C virus-infected patients. J. Clin. Immunol. 2008, 28, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.; Lin, T.Y.; Beasley, D.W.; Stover, C.M.; Schwaeble, W.J.; Pierson, T.C.; Diamond, M.S. Direct complement restriction of flavivirus infection requires glycan recognition by mannose-binding lectin. Cell Host Microbe 2010, 8, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Ezekowitz, R.A.; Kuhlman, M.; Groopman, J.E.; Byrn, R.A. A human serum mannose-binding protein inhibits in vitro infection by the human immunodeficiency virus. J. Exp. Med. 1989, 169, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.L.; Saifuddin, M.; Spear, G.T. Glycosylation inhibitors and neuraminidase enhance human immunodeficiency virus type 1 binding and neutralization by mannose-binding lectin. J. Gen. Virol. 2003, 84, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances transinfection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.P.; Lasala, F.; Carrillo, J.; Muniz, O.; Corbi, A.L.; Delgado, R. C-type lectins DC-SIGN and L-SIGN mediate cellular entry by Ebola virus in cis and in trans. J. Virol. 2002, 76, 6841–6844. [Google Scholar] [CrossRef] [PubMed]
- Halary, F.; Amara, A.; Lortat-Jacob, H.; Messerle, M.; Delaunay, T.; Houlès, C.; Fieschi, F.; Arenzana-Seisdedos, F.; Moreau, J.F.; Déchanet-Merville, J. Human cytomegalovirus binding to DC-SIGN is required for dendritic cell infection and target cell trans-infection. Immunity 2002, 17, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Klimstra, W.B.; Nangle, E.M.; Smith, M.S.; Yurochko, A.D.; Ryman, K.D. DC-SIGN and L-SIGN can act as attachment receptors for alphaviruses and distinguish between mosquito cell- and mammalian cell-derived viruses. J. Virol. 2003, 77, 12022–12032. [Google Scholar] [CrossRef] [PubMed]
- Tassaneetrithep, B.; Burgess, T.H.; Granelli-Piperno, A.; Trumpfheller, C.; Finke, J.; Sun, W.; Eller, M.A.; Pattanapanyasat, K.; Sarasombath, S.; Birx, D.L. DC-SIGN (CD209) mediates dengue virus infection of human dendritic cells. J. Exp. Med. 2003, 197, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Spear, G.T.; Zariffard, M.R.; Xin, J.; Saifuddin, M. Inhibition of DC-SIGN-mediated trans infection of T cells by mannose-binding lectin. Immunology 2003, 110, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Olinger, G.G.; Aris, S.; Chen, Y.; Gewurz, H.; Spear, G.T. Mannose-binding lectin binds to Ebola and Marburg envelope glycoproteins, resulting in blocking of virus interaction with DC-SIGN and complement-mediated virus neutralization. J. Gen. Virol. 2005, 86, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.; Kliem, A.; Tornoe, I.; Skjodt, K.; Koch, C.; Holmskov, U. Localization of lung surfactant protein D on mucosal surfaces in human tissues. J. Immunol. 2000, 164, 5866–5870. [Google Scholar] [CrossRef] [PubMed]
- Voorhout, W.F.; Veenendaal, T.; Kuroki, Y.; Ogasawara, Y.; van Golde, L.M.G.; Geuze, H.J. Immunocytochemical localization of surfactant protein D (SP-D) in type II cells, Clara cells, and alveolar macrophages of rat lung. J. Histochem. Cytochem. 1992, 40, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Herias, M.V.; Hogenkamp, A.; van Asten, A.J.; Tersteeg, M.H.; van Eijk, M.; Haagsman, H.P. Expression sites of the collectin SP-D suggest its importance in first line host defence: Power of combining in situ hybridisation, RT-PCR and immunohistochemistry. Mol. Immunol. 2007, 44, 3324–3332. [Google Scholar] [CrossRef] [PubMed]
- Hartshorn, K.L.; White, M.R.; Shepherd, V.L.; Reid, K.B.M.; Jensenius, J.C.; Crouch, E.C. Mechanisms of anti-influenza activity of surfactant proteins A and D: Comparison with serum collectins. Am. J. Physiol. 1997, 273, L1156–L1166. [Google Scholar] [PubMed]
- Hillaire, M.L.; Haagsman, H.P.; Osterhaus, A.D.; Rimmelzwaan, G.F.; van Eijk, M. Pulmonary surfactant protein D in first-line innate defence against influenza A virusinfections. J. Innate Immun. 2013, 5, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Brown-Augsburger, P.; Hartshorn, K.L.; Chang, D.; Rust, K.; Fliszar, C.; Welgus, H.G.; Crouch, E.C. Site-directed mutagenesis of Cys-15 and Cys-20 of pulmonary surfactant protein D. Expression of a trimeric protein with altered anti-viral properties. J. Biol. Chem. 1996, 271, 13724–13730. [Google Scholar] [CrossRef] [PubMed]
- Hartshorn, K.L.; White, M.R.; Voelker, D.R.; Coburn, J.; Zaner, K.; Crouch, E.C. Mechanism of binding of surfactant protein D to influenza A viruses: Importance of binding to haemagglutinin to antiviral activity. Biochem. J. 2000, 351, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Hillaire, M.L.; van Eijk, M.; van Trierum, S.E.; van Riel, D.; Saelens, X.; Romijn, R.A.; Hemrika, W.; Fouchier, R.A.; Kuiken, T.; Osterhaus, A.D.; et al. Assessment of the antiviral properties of recombinant porcine SP-D against various influenza A viruses in vitro. PLoS One 2011, 6, e25005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tecle, T.; White, M.R.; Crouch, E.C.; Hartshorn, K.L. Inhibition of influenza viral neuraminidase activity by collectins. Arch. Virol. 2007, 152, 1731–1742. [Google Scholar] [CrossRef] [PubMed]
- Hartshorn, K.L.; Crouch, E.C.; White, M.R.; Eggleton, P.; Tauber, A.I.; Chang, D.; Sastry, K. Evidence for a protective role of pulmonary surfactant protein D (SP-D) against influenza A viruses. J. Clin. Investig. 1994, 94, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.Z.; Griffin, G.L.; Senior, R.M.; Longmore, W.J.; Moxley, M.A. Recombinant SP-D carbohydrate recognition domain is a chemoattractant for human neutrophils. Am. J. Physiol. 1999, 276, L131–L136. [Google Scholar] [PubMed]
- Ghildyal, R.; Hartley, C.; Varrasso, A.; Meanger, J.; Voelker, D.R.; Anders, E.M.; Mills, J. Surfactant protein A binds to the fusion glycoprotein of respiratory syncytial virus and neutralizes virion infectivity. J. Infect. Dis. 1999, 180, 2009–2013. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.E.; Pedigo, H.; Johnson, T.R.; Shepherd, V.L. Surfactant protein A enhances uptake of respiratory syncytial virus by monocytes and U937 macrophages. Am. J. Respir. Cell Mol. Biol. 2000, 23, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Griese, M. Respiratory syncytial virus and pulmonary surfactant. Viral Immunol. 2002, 15, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Figdor, C.G.; van Kooyk, Y.; Adema, G.J. C-type lectin receptors on dendritic cells and Langerhans cells. Nat. Rev. Immunol. 2002, 2, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Van Kooyk, Y.; Geijtenbeek, T.B. DC-SIGN: Escape mechanism for pathogens. Nat. Rev. Immunol. 2003, 3, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Lozach, P.Y.; Burleigh, L.; Staropoli, I.; Amara, A. The C type lectins DC-SIGN and L-SIGN: Receptors for viral glycoproteins. Methods Mol. Biol. 2007, 379, 51–68. [Google Scholar] [PubMed]
- Davis, C.W.; Nguyen, H.Y.; Hanna, S.L.; Sánchez, M.D.; Doms, R.W.; Pierson, T.C. West Nile virus discriminates between DC-SIGN and DC-SIGNR for cellular attachment and infection. J. Virol. 2006, 80, 1290–1301. [Google Scholar] [CrossRef] [PubMed]
- Martina, B.E.; Koraka, P.; van den Doel, P.; Rimmelzwaan, G.F.; Haagmans, B.L.; Osterhaus, A.D. DC-SIGN enhances infection of cells with glycosylated West Nile virus in vitro and virus replication in human dendritic cells induces production of IFN-alpha and TNF-alpha. Virus Res. 2008, 135, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Marzi, A.; Gramberg, T.; Simmons, G.; Möller, P.; Rennekamp, A.J.; Krumbiegel, M.; Geier, M.; Eisemann, J.; Turza, N.; Saunier, B.; et al. DC-SIGN and DC-SIGNR interact with the glycoprotein of Marburg virus and the S protein of severe acute respiratory syndrome coronavirus. J. Virol. 2004, 78, 12090–12095. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y.; Huang, Y.; Ganesh, L.; Leung, K.; Kong, W.P.; Schwartz, O.; Subbarao, K.; Nabel, G.J. pH-Dependent entry of severe acute respiratory syndrome coronavirus is mediated by the spike glycoprotein and enhanced by dendritic cell transfer through DC-SIGN. J. Virol. 2004, 78, 5642–5650. [Google Scholar] [CrossRef] [PubMed]
- Jeffers, S.A.; Tusell, S.M.; Gillim-Ross, L.; Hemmila, E.M.; Achenbach, J.E.; Babcock, G.J.; Thomas, W.D.; Thackray, L.B.; Young, M.D.; Mason, R.J.; et al. CD209L (L-SIGN) is a receptor for severe acute respiratory syndrome coronavirus. Proc. Natl. Acad. Sci. USA 2004, 101, 15748–15753. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.P.; Durso, R.J.; Arrigale, R.R.; Donovan, G.P.; Maddon, P.J.; Dragic, T.; Olson, W.C. L-SIGN (CD 209L) is a liver-specific capture receptor for hepatitis C virus. Proc. Natl. Acad. Sci. USA 2003, 100, 4498–4503. [Google Scholar] [CrossRef] [PubMed]
- Pöhlmann, S.; Zhang, J.; Baribaud, F.; Chen, Z.; Leslie, G.J.; Lin, G.; Granelli-Piperno, A.; Doms, R.W.; Rice, C.M.; McKeating, J.A. Hepatitis C virus glycoproteins interact with DC-SIGN and DC-SIGNR. J. Virol. 2003, 77, 4070–4080. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Sanchez, E.; Altmeyer, R.; Amara, A.; Schwartz, O.; Fieschi, F.; Virelizier, J.L.; Arenzana-Seisdedos, F.; Desprès, P. Dendritic-cell-specific ICAM3-grabbing non-integrin is essential for the productive infection of human dendritic cells by mosquito-cell-derived dengue viruses. EMBO Rep. 2003, 4, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Lozach, P.Y.; Burleigh, L.; Staropoli, I.; Navarro-Sanchez, E.; Harriague, J.; Virelizier, J.L.; Rey, F.A.; Desprès, P.; Arenzana-Seisdedos, F.; Amara, A. Dendritic cell-specific intercellular adhesion molecule 3-grabbing non-integrin (DC-SIGN)-mediated enhancement of Dengue virus infection is independent of DC-SIGN internalization signals. J. Biol. Chem. 2005, 280, 23698–23708. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.L.; de Wet, B.J.M.; Martinez-Pomares, L.; Radcliffe, C.M.; Dwek, R.A.; Rudd, P.M.; Gordon, S. The mannose receptor mediates dengue virus infection of macrophages. PLoS Pathog. 2008, 4, e17. [Google Scholar] [CrossRef] [PubMed]
- Op den Brouw, M.L.; Binda, R.S.; Geijtenbeek, T.B.; Janssen, H.L.; Woltman, A.M. The mannose receptor acts as hepatitis B virus surface antigen receptor mediating interaction with intrahepaticdendritic cells. Virology 2009, 393, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Reading, P.C.; Miller, J.L.; Anders, E.M. Involvement of themannose receptor in infection of macrophages by influenza virus. J. Virol. 2000, 74, 5190–5197. [Google Scholar] [CrossRef] [PubMed]
- Upham, J.P.; Pickett, D.; Irimura, T.; Anders, E.M. Reading PC Macrophage receptors for influenza a virus: Role of the macrophage galactose-type lectin and mannose receptorin viral entry. J. Virol. 2010, 84, 3730–3737. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Bernhard, O.K.; Turville, S.G.; Harman, A.N.; Wilkinson, J.; Cunningham, A.L. Oligomerization of the macrophage mannose receptor enhances gp120-mediated binding of HIV-1. J. Biol. Chem. 2009, 284, 11027–11038. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L.; Corliss, B.C.; Wu, J.; Leong, C.; Phillips, J.H. Immunoreceptor DAP12 bearing a tyrosine-based activation motif is involved in activating NK cells. Nature 1998, 391, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.T.; Lin, Y.L.; Huang, M.T.; Wu, M.F.; Cheng, S.C.; Lei, H.Y.; Lee, C.K.; Chiou, T.W.; Wong, C.H.; Hsieh, S.L. CLEC5A is critical for dengue-virus-induced lethal disease. Nature 2008, 453, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.F.; Chen, S.T.; Yang, A.H.; Lin, W.W.; Lin, Y.L.; Chen, N.J.; Tsai, I.S.; Li, L.; Hsieh, S.L. CLEC5A is critical for dengue virus-induced inflammasome activation in human macrophages. Blood 2013, 121, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.T.; Liu, R.S.; Wu, M.F.; Lin, Y.L.; Chen, S.Y.; Tan, D.T.; Chou, T.Y.; Tsai, I.S.; Li, L.; Hsieh, S.L. CLEC5A Regulates Japanese encephalitis virus-induced neuroinflammation and lethality. PLoS Pathog. 2012, 8, e1002655. [Google Scholar] [CrossRef] [PubMed]
- Valladeau, J.; Ravel, O.; Dezutter-Dambuyant, C.; Moore, K.; Kleijmeer, M.; Liu, Y.; Duvert-Frances, V.; Vincent, C.; Schmitt, D.; Davoust, J.; et al. Langerin, a novel C-type lectin specific to Langerhans cells, is an endocytic receptor that induces the formation of Birbeck granules. Immunity 2000, 12, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Turville, S.; Wilkinson, J.; Cameron, P.; Dable, J.; Cunningham, A.L. The role of dendritic cell C-type lectin receptors in HIV pathogenesis. J. Leukoc. Biol. 2003, 74, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Turville, S.G.; Cameron, P.U.; Handley, A.; Lin, G.; Pöhlmann, S.; Doms, R.W.; Cunningham, A.L. Diversity of receptors binding HIV on dendritic cell subsets. Nat. Immunol. 2002, 3, 975–983. [Google Scholar] [CrossRef] [PubMed]
- De Witte, L.; Nabatov, A.; Pion, M.; Fluitsma, D.; De Jong, M.A.; de Gruijl, T.; Piguet, V.; van Kooyk, Y.; Geijtenbeek, T.B. Langerin is a natural barrier to HIV-1 transmission by Langerhans cells. Nat. Med. 2007, 13, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Van der Vlist, M.; de Witte, L.; de Vries, R.D.; Litjens, M.; de Jong, M.A.; Fluitsma, D.; de Swart, R.L.; Geijtenbeek, T.B. Human Langerhans cells capture measles virus through Langerin and present viral antigens to CD4+ T cells but are incapable of cross-presentation. Eur. J. Immunol. 2011, 41, 2619–2631. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-type lectin superfamily in the immune system. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; van Vliet, S.J.; Engering, A.; ’t Hart, B.A.; van Kooyk, Y. Self-and nonself-recognition by C-type lectins on dendritic cells. Annu. Rev. Immunol. 2004, 22, 33–54. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Drickamer, K. Trimeric structure of a C-type mannose-binding protein. Structure 1994, 2, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Barondes, S.H.; Castronovo, V.; Cooper, D.N.; Cummings, R.D.; Drickamer, K.; Feizi, T.; Gitt, M.A.; Hirabayashi, J.; Hughes, C.; Kasai Hpohlmann, K.; et al. Galectins: A family of animal beta-galactosidebinding lectins. Cell 1994, 76, 597–598. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.C. Secretion of the galectin family of mammalian carbohydrate-binding proteins. BBA-Gen. Subj. 1999, 1473, 172–185. [Google Scholar] [CrossRef]
- Sato, S.; Burdett, I.; Hughes, R.C. Secretion of the baby hamster kidney 30-kDa galactose-binding lectin from polarized and nonpolarized cells: A pathway independent of the endoplasmic reticulum-Golgi complex. Exp. Cell. Res. 1993, 207, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Mehul, B.; Hughes, R.C. Plasma membrane targetting, vesicular budding and release of galectin 3 from the cytoplasm of mammalian cells during secretion. J. Cell Sci. 1997, 110, 1169–1178. [Google Scholar] [PubMed]
- Liu, F.T.; Rabinovich, G.A. Galectins: Regulators of acute and chronic inflammation. Ann. N. Y. Acad. Sci. 2010, 1183, 158–182. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.-Y.; Rabinovich, G.A.; Liu, F.-T. Galectins: Structure, function and therapeutic potential. Expert Rev. Mol. Med. 2008, 10, e17. [Google Scholar] [CrossRef] [PubMed]
- Garner, O.B.; Aguilar, H.C.; Fulcher, J.A.; Levroney, E.L.; Harrison, R.; Wright, L.; Robinson, L.R.; Aspericueta, V.; Panico, M.; Haslam, S.M.; et al. Endothelial galectin-1 binds to specific glycans on nipah virus fusion protein and inhibits maturation, mobility, and function to block syncytia formation. PLoS Pathog. 2010, 6, e1000993. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.L.; Chen, Y.H.; Wang, S.W.; Huang, Y.J.; Leu, C.H.; Yeh, N.C.; Chu, C.Y.; Lin, C.C.; Shieh, G.S.; Chen, Y.L.; et al. Galectin-1 binds to influenza virus and ameliorates influenza virus pathogenesis. J. Virol. 2011, 85, 10010–10020. [Google Scholar] [CrossRef] [PubMed]
- Mercier, S.; St-Pierre, C.; Pelletier, I.; Ouellet, M.; Tremblay, M.J.; Sato, S. Galectin-1 promotes HIV-1 infectivity in macrophages through stabilization of viral adsorption. Virology 2008, 371, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, M.; Mercier, S.; Pelletier, I.; Bounou, S.; Roy, J.; Hirabayashi, J.; Sato, S.; Tremblay, M.J. Galectin-1 acts as a soluble host factor that promotes HIV-1 infectivity through stabilization of virus attachment to host cells. J. Immunol. 2005, 174, 4120–4126. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, C.; Ouellet, M.; Tremblay, M.J.; Sato, S. Galectin-1 and HIV-1 infection. Methods Enzymol. 2010, 480, 267–294. [Google Scholar] [PubMed]
- St-Pierre, C.; Manya, H.; Ouellet, M.; Clark, G.F.; Endo, T.; Tremblay, M.J.; Sato, S. Host-soluble galectin-1 promotes HIV-1 replication through a direct interaction with glycans of viral gp120 andhost CD4. J. Virol. 2011, 85, 11742–11751. [Google Scholar] [CrossRef] [PubMed]
- Woodward, A.M.; Mauris, J.; Argüeso, P. Binding of transmembrane mucins to galectin-3 limits herpesvirus 1 infection of human corneal keratinocytes. J. Virol. 2013, 87, 5841–5847. [Google Scholar] [CrossRef] [PubMed]
- Garcin, P.; Cohen, S.; Terpstra, S.; Kelly, I.; Foster, L.J.; Panté, N. Proteomic analysis identifies a novel function for galectin-3 in the cell entry of parvovirus. J. Proteomics 2013, 79, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Parodi, A.J. Role of N-oligosaccharides endoplasmic reticulum processing reactions in glycoprotein folding and degradation. Biochem. J. 2000, 348, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, E.S.; Helenius, A. Lectins as chaperones in glycoprotein folding. Curr. Opin. Struct. Biol. 1998, 8, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Ou, W.J.; Cameron, P.H.; Thomas, D.Y.; Bergeron, J.J. Association of folding intermediates of glycoproteins with calnexin during protein maturation. Nature 1993, 364, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.; Braakman, I.; Helenius, A. Role of N-linked oligosaccharide recognition, glucose trimming, and calnexin in glycoprotein folding and quality control. Proc. Natl. Acad. Sci. USA 1994, 91, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.R.; Ora, A.; Nguyen Van, P.; Helenius, A. Transient, lectin-like association of calreticulin with folding intermediates of cellular and viral glycoproteins. Mol. Biol. Cell 1995, 6, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Cannon, K.S.; Hebert, D.N.; Helenius, A. Glycan-dependent and -independent association of vesicular stomatitis virus G protein with calnexin. J. Biol. Chem. 1996, 271, 14280–14284. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Yamashita, T.; Sato, H.; Taira, H. Kinetics of interactions of sendai virus envelope glycoproteins, F and HN, with endoplasmic reticulum-residentmolecular chaperones, BiP, calnexin, and calreticulin. J. Biochem. 1999, 126, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Veijola, J.; Pettersson, R.F. Transient association of calnexin and calreticulin with newly synthesized G1 and G2 glycoproteins of uukuniemivirus (family Bunyaviridae). J. Virol. 1999, 73, 6123–6127. [Google Scholar] [PubMed]
- Fukushi, M.; Yoshinaka, Y.; Matsuoka, Y.; Hatakeyama, S.; Ishizaka, Y.; Kirikae, T.; Sasazuki, T.; Miyoshi-Akiyama, T. Monitoring of S protein maturation in the endoplasmic reticulum by calnexin is important for the infectivity of severe acute respiratory syndrome coronavirus. J. Virol. 2012, 86, 11745–11753. [Google Scholar] [CrossRef] [PubMed]
- Werr, M.; Prange, R. Role for calnexin and N-linked glycosylation in the assembly and secretion of hepatitis B virus middle envelopeprotein particles. J. Virol. 1998, 72, 778–782. [Google Scholar] [PubMed]
- Otteken, A.; Moss, B. Calreticulin interacts with newly synthesized human immunodeficiency virus type 1 envelope glycoprotein, suggesting a chaperone function similar to that of calnexin. J. Biol. Chem. 1996, 271, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bergeron, J.J.; Luo, L.; Ou, W.J.; Thomas, D.Y.; Kang, C.Y. Effects of inefficient cleavage of the signal sequence of HIV-1 gp 120 on its association with calnexin, folding, and intracellular transport. Proc. Natl. Acad. Sci. USA 1996, 93, 9606–9611. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Ball, J.M.; Zeng, C.Q.; Estes, M.K. The rotavirus nonstructural glycoprotein NSP4 possesses membrane destabilization activity. J. Virol. 1996, 70, 6973–6981. [Google Scholar] [PubMed]
- Newton, K.; Meyer, J.C.; Bellamy, A.R.; Taylor, J.A. Rotavirus nonstructural glycoprotein NSP4 alters plasma membrane permeability in mammalian cells. J. Virol. 1997, 71, 9458–9465. [Google Scholar] [PubMed]
- Mirazimi, A.; Nilsson, M.; Svensson, L. The molecular chaperone calnexin interacts with the NSP4 enterotoxin of rotavirus in vivo and in vitro. J. Virol. 1998, 72, 8705–8709. [Google Scholar] [PubMed]
- Shi, X.; Elliott, R.M. Analysis of N-linked glycosylation of Hantaan virus glycoproteins and the role of oligosaccharide side chains in protein folding and intracellular trafficking. J. Virol. 2004, 78, 5414–5422. [Google Scholar] [CrossRef] [PubMed]
- Hoflack, B.; Kornfeld, S. Lysosomal enzyme binding to mouse P388D1 macrophage membranes lacking the 215-kDa mannose 6-phosphate receptor: Evidence for the existence of a second mannose 6-phosphate receptor. Proc. Natl. Acad. Sci. USA 1985, 82, 4428–4432. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, C.R.; Burke, R.L.; Kornfeld, S.; Gregory, W.; Dingwell, K.S.; Masiarz, F.; Johnson, D.C. Herpes simplex virus glycoprotein D acquires mannose 6-phosphate residues and binds to mannose 6-phosphate receptors. J. Biol. Chem. 1994, 269, 17067–17074. [Google Scholar] [PubMed]
- Brunetti, C.R.; Burke, R.L.; Hoflack, B.; Ludwig, T.; Dingwell, K.S.; Johnson, D.C. Role of mannose-6-phosphate receptors in herpes simplex virus entry into cells and cell-to-cell transmission. J. Virol. 1995, 69, 3517–3528. [Google Scholar] [PubMed]
- Chen, J.J.; Zhu, Z.L.; Gershon, A.A.; Gershon, M.D. Mannose 6-phosphate receptor dependence of varicella zoster virus infection in vitro and in the epidermis during Varicella and Zoster. Cell 2004, 119, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, K.; Simons, K. A putative novel class of animal lectins in the secretory pathway homologous to leguminous lectins. Cell 1994, 77, 625–626. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Sato, K.; Kanoh, A.; Yamashita, K.; Yamada, Y.; Igarashi, N.; Kato, R.; Nakano, A.; Wakatsuki, S. Structures of the carbohydrate recognition domain of Ca2+-independent cargo receptors Emp46p and Emp47p. J. Biol. Chem. 2006, 281, 10410–10419. [Google Scholar] [CrossRef] [PubMed]
- Klaus, J.P.; Eisenhauer, P.; Russo, J.; Mason, A.B.; Do, D.; King, B.; Taatjes, D.; Cornillez-Ty, C.; Boyson, J.E.; Thali, M.; et al. The intracellular cargo receptor ERGIC-53 is required for the production of infectious arenavirus, coronavirus, and filovirus particles. Cell Host Microbe 2013, 14, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.Z.; Wang, L.; Xu, S.; Zhang, X.W.; Zhao, X.F.; Vasta, G.R.; Wang, J.X. A galectin from the kuruma shrimp (Marsupenaeus japonicus) functions as an opsonin and promotes bacterial clearance from hemolymph. PLoS One 2014, 9, e91794. [Google Scholar] [CrossRef] [PubMed]
- Kurz, S.; Jin, C.; Hykollari, A.; Gregoric, D.; Giomarelli, B.; Vasta, G.R.; Wilson, I.B.; Paschinger, K. Hemocytes and plasma of the eastern oyster (Crassostrea virginica) display a diverse repertoire of sulfated and blood group A-modified N-glycans. J. Biol. Chem. 2013, 288, 24410–24428. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Ghosh, A.; Amin, M.N.; Giomarelli, B.; Shridhar, S.; Banerjee, A.; Fernández-Robledo, J.A.; Bianchet, M.A.; Wang, L.X.; Wilson, I.B.; et al. The galectin CvGal1 from the eastern oyster (Crassostrea virginica) binds to blood group A oligosaccharides on the hemocyte surface. J. Biol. Chem. 2013, 288, 24394–24409. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhang, Y.; Yu, F.; Yu, Z. A novel sialic acid binding lectin with anti-bacterial activity from the Hong Kong oyster (Crassostrea hongkongensis). Fish Shellfish Immunol. 2011, 31, 1247–1250. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yu, S.; Zhao, J.; Su, X.; Li, T. Cloning and characterization of a sialic acid binding lectins (SABL) from Manila clam. Fish Shellfish Immunol. 2011, 30, 1202–1206. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, T.; Erdmann, H.; Fleischer, B. Molecular interaction of Siglecs (sialic acid-binding Ig-like lectins) with sialylated ligands on Trypanosoma cruzi. Eur. J. Cell Biol. 2010, 89, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Wang, J.X. Diversity and multiple functions of lectins in shrimp immunity. Dev. Comp. Immunol. 2013, 39, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, F.; Liu, J.; Xiao, X.; Zhang, S.; Qin, C.; Xiang, Y.; Wang, P.; Cheng, G. Transmission-blocking antibodies against mosquito C-type lectins for dengue prevention. PLoS Pathog. 2014, 10, e1003931. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, Y.; Li, F.; Liu, J.; Sun, Y.; Wu, J. Cloning and characterization of a mannose binding C-type lectin gene from salivary gland of Aedes albopictus. Parasite. Vector. 2014, 7, 337. [Google Scholar] [CrossRef]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.M.; Wides, R.; et al. The genome sequence of the malaria mosquito Anopheles gambiae. Science 2002, 298, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Yang, H.J.; Li, F.; Zhang, X.B.; Xu, X. Purification, characterization and cDNA cloning of a novel lipopolysaccharide-binding lectin from the shrimp Penaeus monodon. Dev. Comp. Immunol. 2006, 30, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Zhang, X.W.; Xu, W.T.; Zhao, X.F.; Wang, J.X. A novel C-type lectin (FcLec4) facilitates the clearance of Vibrio anguillarumin in vivo in Chinese white shrimp. Dev. Comp. Immunol. 2009, 33, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Cox, J.; Wang, P.; Krishnan, M.N.; Dai, J.; Qian, F.; Anderson, J.F.; Fikrig, E. A C-type lectin collaborates with a CD45 phosphatase homolog to facilitate West Nile virus infection of mosquitoes. Cell 2010, 142, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xu, Y.H.; Xu, J.D.; Zhao, X.F.; Wang, J.X. Collaboration between a soluble C-type lectin and calreticulin facilitates white spot syndrome virus infection in shrimp. J. Immunol. 2014, 193, 2106–20117. [Google Scholar] [CrossRef] [PubMed]
- Pustylnikov, S.; Sagar, D.; Jain, P.; Khan, Z.K. Targeting the C-type lectins-mediated host-pathogen interactions with dextran. J. Pharm. Pharm. Sci. 2014, 17, 371–392. [Google Scholar] [PubMed]
- Xiong, C.; O’Keefe, B.R.; Botos, I.; Wlodawer, A.; McMahon, J.B. Overexpression and purification of scytovirin, a potent, novel anti-HIV protein from the cultured cyanobacterium Scytonema varium. Protein Expr. Purif. 2006, 46, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Neyts, J.; Schols, D.; Hosoya, M.; Van Damme, E.; Peumans, W.; De Clercq, E. The mannose-specific plant lectins from Cymbidium hybrid and Epipactis helleborineand the (N-acetylglucosamine)n-specific plant lectin from Urtica dioicaare potent and selective inhibitors of human immunodeficiency virus and cytomegalovirus replication in vitro. Antivir. Res. 1992, 18, 191–207. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J.; Van Laethem, K.; Hatse, S.; Froeyen, M.; Peumans, W.; Van Damme, E.; Schols, D. Carbohydrate-binding agents cause deletions of highly conserved glycosylation sites in HIV gp120 A new therapeutic concept to hit the Achilles heel of HIV. J. Biol. Chem. 2005, 280, 41005–41014. [Google Scholar] [CrossRef] [PubMed]
- Alen, M.M.; de Burghgraeve, T.; Kaptein, S.J.; Balzarini, J.; Neyts, J.; Schols, D. Broad antiviral activity of carbohydrate-binding agents against the four serotypes of dengue virus in monocyte-derived dendritic cells. PLoS One 2011, 6, e21658. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, C.N.; Pantophlet, R.; Wormald, M.R.; Ollmann, S.E.; Stanfield, R.; Wilson, I.A.; Katinger, H.; Dwek, R.A.; Rudd, P.M.; Burton, D.R. The broadly neutralizing anti-human immunodeficiency virus type 1 antibody 2G12 recognizes a cluster of alpha 1→2 mannose residues on the outer face of gp120. J. Virol. 2002, 76, 7306–7321. [Google Scholar] [CrossRef] [PubMed]
- Calarese, D.A.; Scanlan, C.N.; Zwick, M.B.; Deechongkit, S.; Mimura, Y.; Kunert, R.; Zhu, P.; Wormald, M.R.; Stanfield, R.L.; Roux, K.H.; et al. Antibody domain exchange is an immunological solution to carbohydrate cluster recognition. Science 2003, 300, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Calarese, D.A.; Lee, H.K.; Huang, C.Y.; Best, M.D.; Astronomo, R.D.; Stanfield, R.L.; Katinger, H.; Burton, D.R.; Wong, C.H.; Wilson, I.A. Dissection of the carbohydrate specificity of the broadly neutralizing anti-HIV-1 antibody 2G12. Proc. Natl. Acad. Sci. USA 2005, 102, 13372–13377. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Liu, J.; Pang, X.; Liu, T.; Ning, Z.; Cheng, G. The Roles of Direct Recognition by Animal Lectins in Antiviral Immunity and Viral Pathogenesis. Molecules 2015, 20, 2272-2295. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules20022272
Liu Y, Liu J, Pang X, Liu T, Ning Z, Cheng G. The Roles of Direct Recognition by Animal Lectins in Antiviral Immunity and Viral Pathogenesis. Molecules. 2015; 20(2):2272-2295. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules20022272
Chicago/Turabian StyleLiu, Yang, Jianying Liu, Xiaojing Pang, Tao Liu, Zhijie Ning, and Gong Cheng. 2015. "The Roles of Direct Recognition by Animal Lectins in Antiviral Immunity and Viral Pathogenesis" Molecules 20, no. 2: 2272-2295. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules20022272