Structure Characterization of Honey-Processed Astragalus Polysaccharides and Its Anti-Inflammatory Activity In Vitro

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Determination of the Molecular Weight of HAPS by HPGFC-ELSD
2.2. Determining the Composition of Monosaccharide
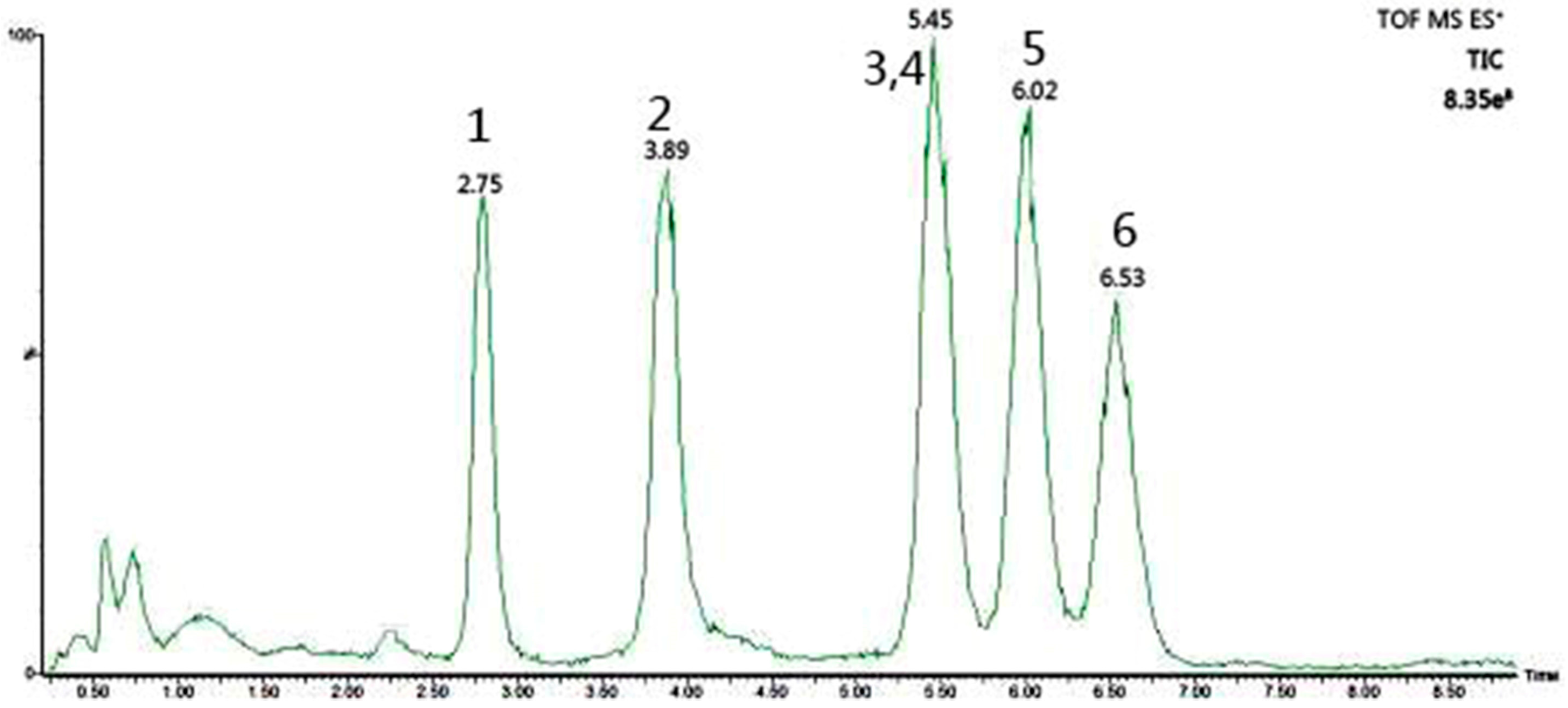
2.2.1. Monosaccharide Composition Determination
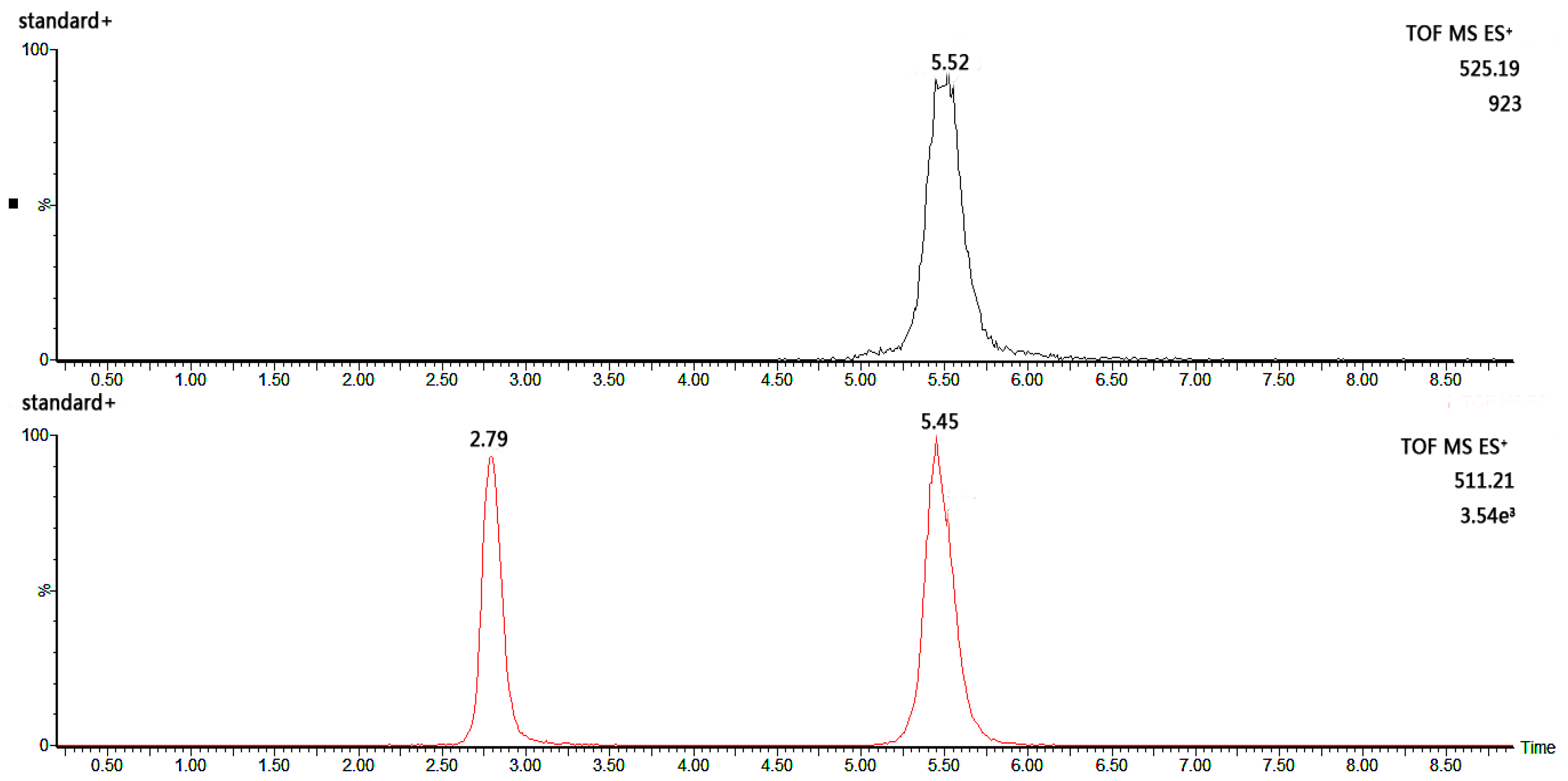
2.2.2. Method Validation for Monosaccharide Composition Testing
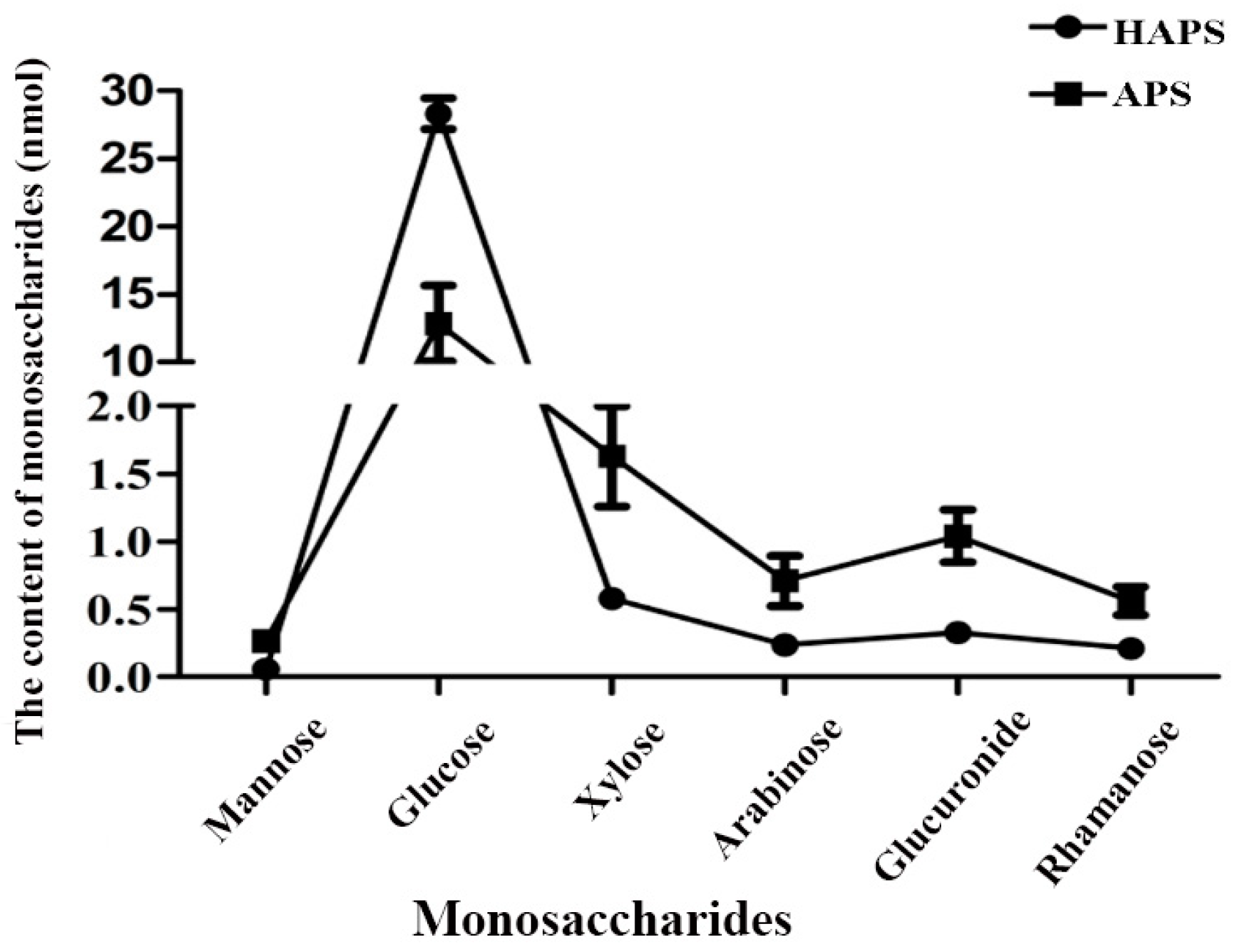
2.2.3. Monosaccharide Composition of HAPS
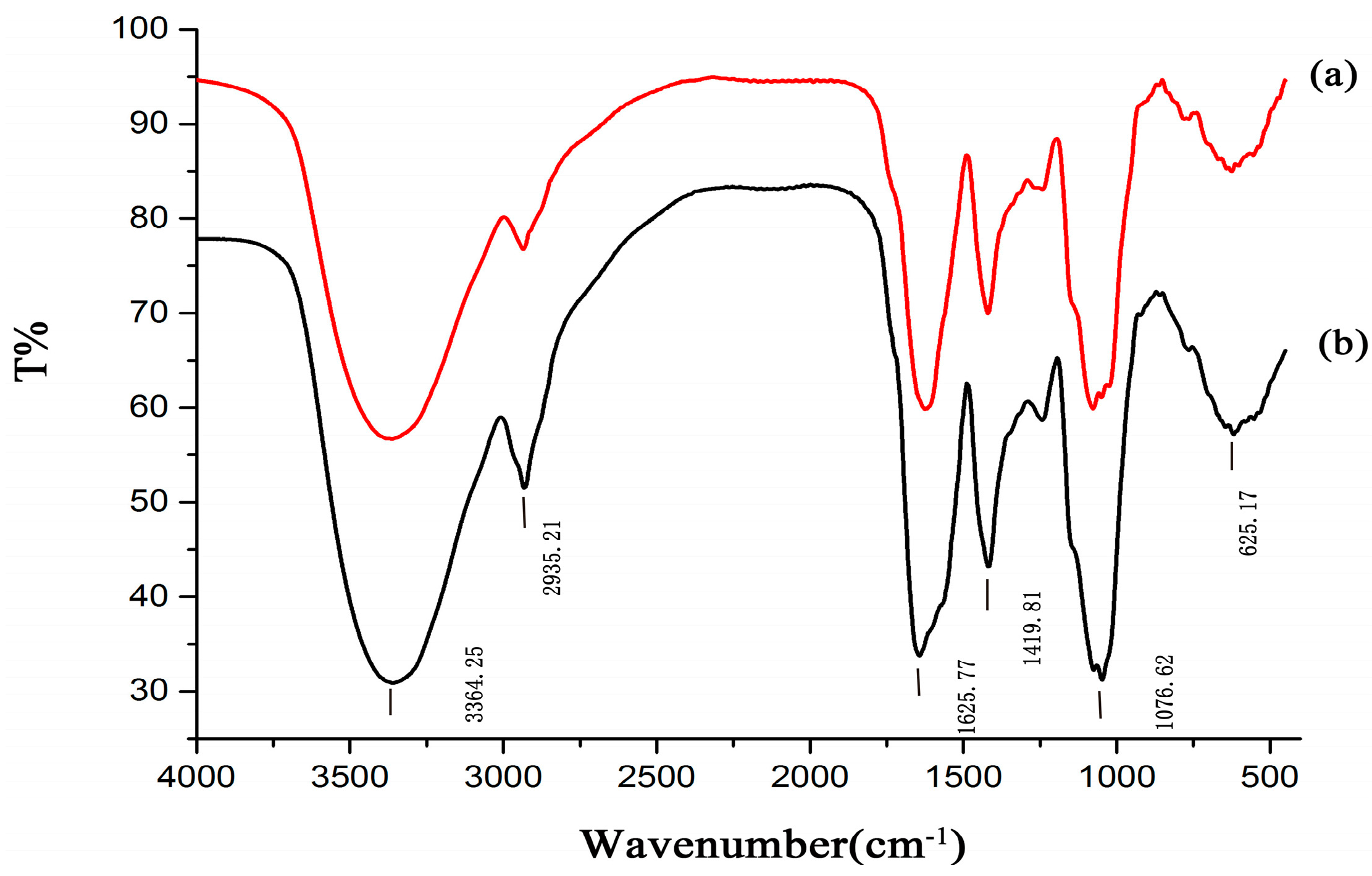
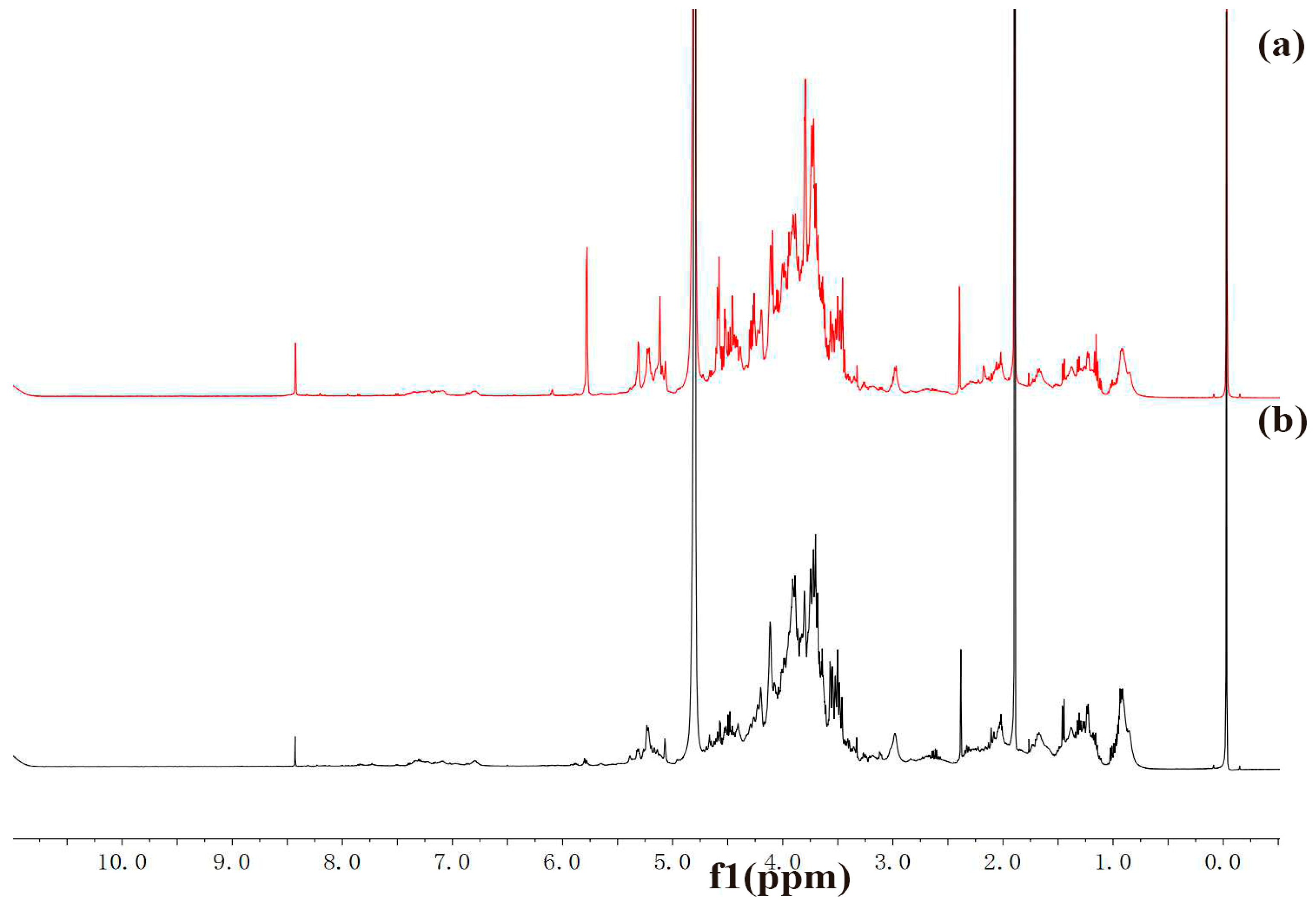
2.3. FT-IR and NMR Analysis
2.4. Anti-Inflammatory Activity of HAPS In Vitro
2.4.1. Effects of the HAPS on Cell Viability and Inhibition of NO Expression in LPS-Stimulated RAW264.7 Cells
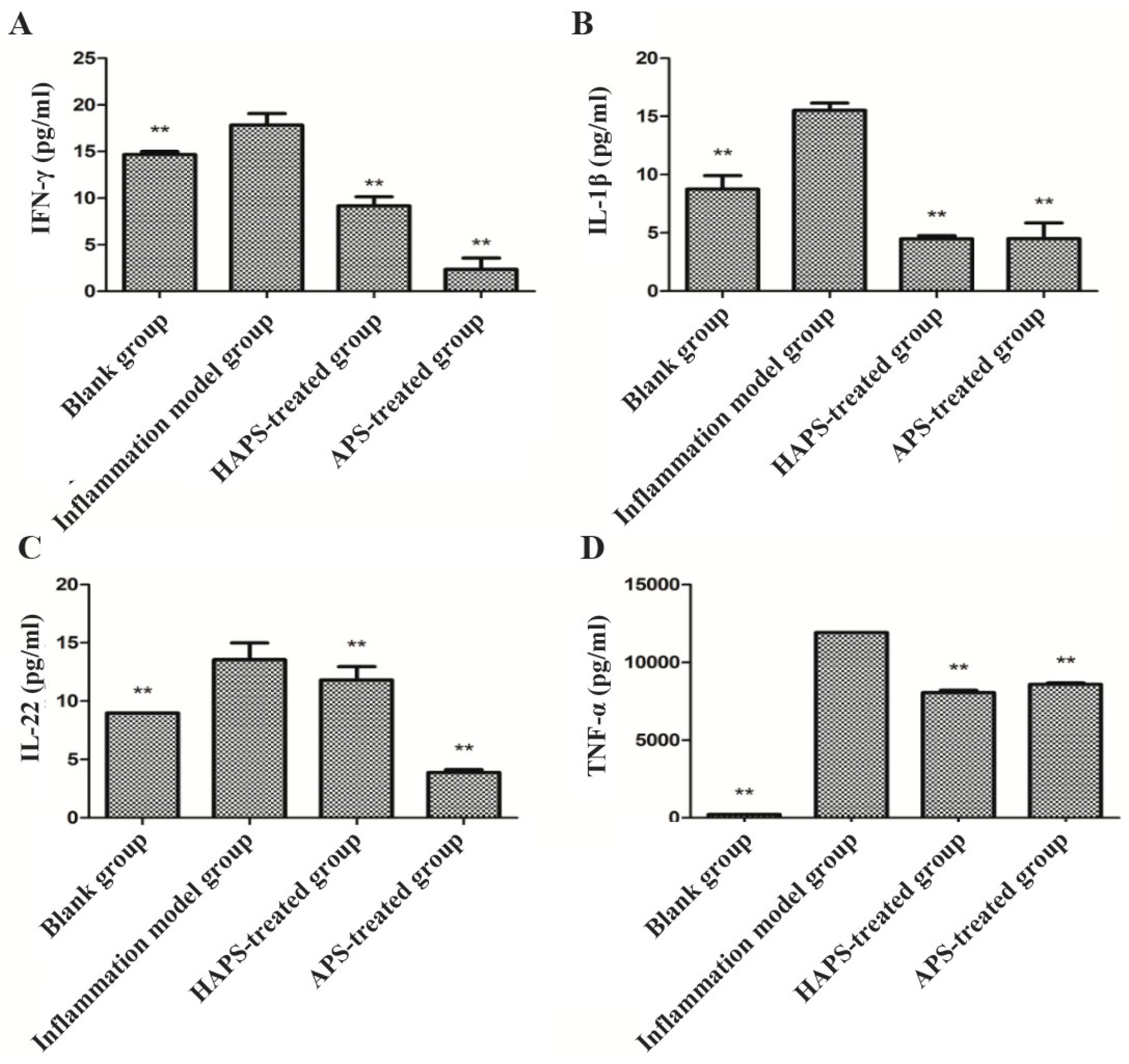
2.4.2. Effects of HAPS on IFN-γ, IL-1β, IL-22 and TNF-α Expression in LPS-Stimulated RAW264.7 Cells
3. Experimental
3.1. Materials and Reagents
3.2. Sample Preparation
3.3. Determination of the Molecular Weight of the Polysaccharides
3.4. Hydrolysis and PMP Derivatization
3.5. UPLC/Q-TOF-MS Analysis of the Composition of Monosaccharide
3.6. Method Validation for Monosaccharide Composition Testing
3.7. FT-IR and NMR Analysis
3.8. Cell Viability Assay & Measurement of NO Release
3.9. ELISA for the Detection of IFN-γ, IL-1β, IL-22 and TNF-α Cytokines Expression
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pharmacopoeia Commission of PRC. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015; Volume 1, p. 302. [Google Scholar]
- Ma, X.Q.; Shi, Q.; Duan, J.A.; Dong, T.T.; Tsim, K.W. Chemical Analysis of Radix Astragalus (Huangqi) in China: A Comparison with Its Adulterants and Seasonal Variations. Food Chem. 2002, 50, 4861–4866. [Google Scholar] [CrossRef]
- Jin, M.; Zhao, K.; Huang, Q.; Shang, P. Structural features and biological activities of the polysaccharides from Astragalus membranaceus. Int. J. Biol. Macromol. 2014, 64, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Xiao, B.; Sun, T. Antitumor and immunomodulatory activity of Astragalus membranaceus polysaccharides in H22 tumor-bearing mice. Int. J. Biol. Macromol. 2013, 62, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zheng, K.; Zhao, H.; Lu, C.; Liu, B.; Yu, C.; Zhang, G.; Bian, Z.; Lu, A.; He, X. Regulatory Effect of Astragalus Polysaccharides on Intestinal Intraepithelial γδT Cells of Tumor Bearing Mice. Molecules 2014, 19, 15224–15236. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, Z.F.; Ding, Y.; Wang, J.B.; Li, Y. Astragalus Polysaccharide Improves Palmitate-Induced Insulin Resistance by Inhibiting PTP1B and NF-κB in C2C12 Myotubes. Molecules 2012, 17, 7083–7092. [Google Scholar] [CrossRef] [PubMed]
- Dun, C.; Liu, J.; Qiu, F.; Wu, X.; Wang, Y.; Zhao, Y.; Gu, P. Effects of Astragalus polysaccharides on memory impairment in a diabetic rat model. Neuropsychiatr. Dis. Treat. 2016, 12, 1617–1621. [Google Scholar] [PubMed]
- Jiang, J.; Wu, C.; Gao, H.; Song, J.; Li, H. Effect of Astragalus polysaccharides on immunologic function of erythrocyte in chickens infected with infectious bursa disease virus. Vaccine 2010, 28, 5614–5616. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Xiao, H.T.; Bao, W.R.; Ma, D.L.; Leung, C.H.; Han, X.Q.; Ko, C.H.; Lau, C.B.S.; Chun-Kwok, W.O.N.G.; Fung, K.P.; et al. TLR-4 may mediate signaling pathways of Astragalus polysaccharide RAP induced cytokine expression of RAW 264.7 cells. J. Ethnopharmacol. 2016, 179, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, Z.; Wang, Z.; Yu, S.; Long, T.; Zhou, X.; Bao, Y. Astragalus polysaccharides exerts immunomodulatory effects via TLR4-mediated MyD88-dependent signaling pathway in vitro and in vivo. Sci. Rep. 2017, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Zhu, F.; Qian, J.M.; Dai, J.Y. Therapeutic and Immunoregulatory Effect of GATA-Binding Protein-3/T-Box Expressed in T-Cells Ratio of Astragalus Polysaccharides on 2,4,6-Trinitrobenzene Sulfonic Acid-Induced Colitis in Rats. Chin. J. Integr. Med. 2016, 22, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, Z.; Zhou, L.; Long, T.; Zhou, X.; Bao, Y. Immunomodulatory effect of APS and PSP is mediated by Ca2+-cAMP and TLR4/NF-kB signaling pathway in macrophage. Int. J. Biol. Macromol. 2017, 93, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Chen, H.; Shi, Z.; Feng, Y.; Rui, W. Rapid and reliable method for analysis of raw and honey-processed Astragalus by UPLC/ESI-Q-TOF-MS using HSS T3 columns. Anal. Methods 2014, 6, 8045–8054. [Google Scholar] [CrossRef]
- Honda, S. Posteolumn derivatization for chromatographic analysis of carbohydrates. J. Chromatogr. A 1996, 720, 183–199. [Google Scholar] [CrossRef]
- Cai, J.F.; Dai, Y.T.; Xiao, Y.Q.; Zhao, R.; Zhang, L.W. Systemic Evaluation of Effect of Honey-processing on Therapeutical Basis of Astragalus Radix. Chin. J. Exp. Tradit. Med. Formul. 2016, 22, 47–52. [Google Scholar]
- Ni, D.J.; Chen, Y.Q.; Xie, B.J.; Zhang, Y.; Zhou, J.R. Spectrum, Morphological and Thermal Characteristics of OTPS 2-1 in Polysaccharides from Oolong Tea. Chem. Res. Chin. Univ. 2004, 25, 2263–2268. [Google Scholar]
- Lv, J.; Zhang, Y.; Tian, Z.; Liu, F.; Shi, Y.; Liu, Y.; Xia, P. Astragalus polysaccharides protect agansit dextran sulfate sodium-induced colitis by inhibiting NF-κB, activation. Int. J. Biol. Macromol. 2017, 98, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Villarino, A.V.; Kanno, Y.; Ferdinand, J.R.; O’Shea, J.J. Mechanisms of Jak/STAT signaling in immunity and disease. J. Immunol. 2015, 194, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.Y.; Chan, B.C.L.; Yu, H.; Lau, I.Y.K.; Han, X.Q.; Cheng, S.W.; Wong, C.K.; Lau, C.B.S.; Xie, M.Y.; Fung, K.P.; et al. Separation, structure characterization, conformation and immunomodulating effect of a hyperbranched heteroglycan from Radix Astragali. Carbohydr. Polym. 2012, 87, 667–675. [Google Scholar] [CrossRef]
- Pu, X.; Ma, X.; Liu, L.; Ren, J.; Li, H.; Li, X.; Yu, S.; Zhang, W.; Fan, W. Structural characterization and antioxidant activity in vitro of polysaccharides from angelica and astragalus. Carbohydr. Polym. 2016, 137, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Kärkkäinen, J.; Lappalainen, K.; Joensuu, P.; Lajunen, M. HPLC-ELSD analysis of six starch species heat-dispersed in [BMIM]Cl ionic liquid. Carbohydr. Polym. 2011, 84, 509–516. [Google Scholar] [CrossRef]
- Fu, D.T.; Oneill, R.A. Monosaccharide composition analysis of oligosaccharides and glycoproteins by high-performance liquid chromatography. Anal. Biochem. 1995, 227, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, Z.; Huang, L.J.; Bai, Q.; Jia, J.F. An improved PMP derivatization method for analyzing monosaccharide composition. Chem. J. Chin. Univ. Chin. Ed. 2006, 27, 1456. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Miles, A.M.; Wink, D.A.; Cook, J.C.; Grisham, M.B. Determination of nitric oxide using fluorescence spectroscopy. Methods Enzymol. 1996, 268, 105–120. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monosaccharides | Relative Retention Time (RSD, %) | Relative Peak Area (RSD, %) |
|---|---|---|
| Mannose | 0.00 | 0.09 |
| Glucose | 0.00 | 0.11 |
| Xylose | 0.00 | 0.12 |
| Arabinose | 0.00 | 0.14 |
| Glucuronic acid | 0.00 | 0.12 |
| Rhamnose | 0.01 | 0.10 |
| Monosaccharides | Relative Retention Time (RSD, %) | Relative Peak Area (RSD,%) |
|---|---|---|
| Mannose | 0.00 | 0.09 |
| Glucose | 0.00 | 0.12 |
| Xylose | 0.01 | 0.06 |
| Arabinose | 0.01 | 0.12 |
| Glucuronic acid | 0.01 | 0.11 |
| Rhamnose | 0.00 | 0.13 |
| Monosaccharide | Standard Curve (n = 6) | R2 (n = 6) | Linear Range (nmol) |
|---|---|---|---|
| Mannose | y = 4.2699x − 2.8281 | R2 = 0.99132 | 0.1~10 |
| Glucose | y = 3.2251x − 7.9961 | R2 = 0.99781 | 0.1~10 |
| Xylose | y = 7.2628x − 17.148 | R2 = 0.99706 | 0.1~10 |
| Arabinose | y = 5.0057x − 13.844 | R2 = 0.99844 | 0.1~10 |
| Glucuronic acid | y = 2.2763x − 5.5898 | R2 = 0.99492 | 0.1~10 |
| Rhamnose | y = 6.4915x − 22.234 | R2 = 0.99614 | 0.1~10 |
| Treatment Groups (μg mL−1) | NO Release Rate/μM | |
|---|---|---|
| Blank group | 0.84 ± 0.50 ** | |
| Inflammation model group | 5.28 ± 0.37 | |
| HAPS | 10 | 2.80 ± 0.43 ** |
| 50 | 2.75 ± 0.66 ** | |
| 100 | 2.44 ± 1.04 ** | |
| 300 | 2.06 ± 1.13 ** | |
| APS | 10 | 3.92 ± 0.98 ** |
| 50 | 2.97 ± 0.67 ** | |
| 100 | 2.81 ± 0.47 ** | |
| 300 | 2.42 ± 0.23 ** | |
| Ibuprofen | 10 | 5.19 ± 0.001 |
| 50 | 4.87 ± 0.004 | |
| 100 | 4.27 ± 0.003 * | |
| 300 | 2.93 ± 0.002 ** | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, J.; Li, C.; Huang, J.; Liu, W.; Chen, H.; Liao, S.; Chen, H.; Rui, W. Structure Characterization of Honey-Processed Astragalus Polysaccharides and Its Anti-Inflammatory Activity In Vitro. Molecules 2018, 23, 168. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23010168
Liao J, Li C, Huang J, Liu W, Chen H, Liao S, Chen H, Rui W. Structure Characterization of Honey-Processed Astragalus Polysaccharides and Its Anti-Inflammatory Activity In Vitro. Molecules. 2018; 23(1):168. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23010168
Chicago/Turabian StyleLiao, Jingzhu, Chanyi Li, Jing Huang, Wuping Liu, Hongce Chen, Shuangye Liao, Hongyuan Chen, and Wen Rui. 2018. "Structure Characterization of Honey-Processed Astragalus Polysaccharides and Its Anti-Inflammatory Activity In Vitro" Molecules 23, no. 1: 168. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23010168