Antioxidant Effect of Extracts from Native Chilean Plants on the Lipoperoxidation and Protein Oxidation of Bovine Muscle

Abstract
:1. Introduction
2. Results and Discussion
2.1. Content of Phenolic Compounds, Flavonoid Compounds, ORAC-PGR, and FRAP Values of Plant Extracts
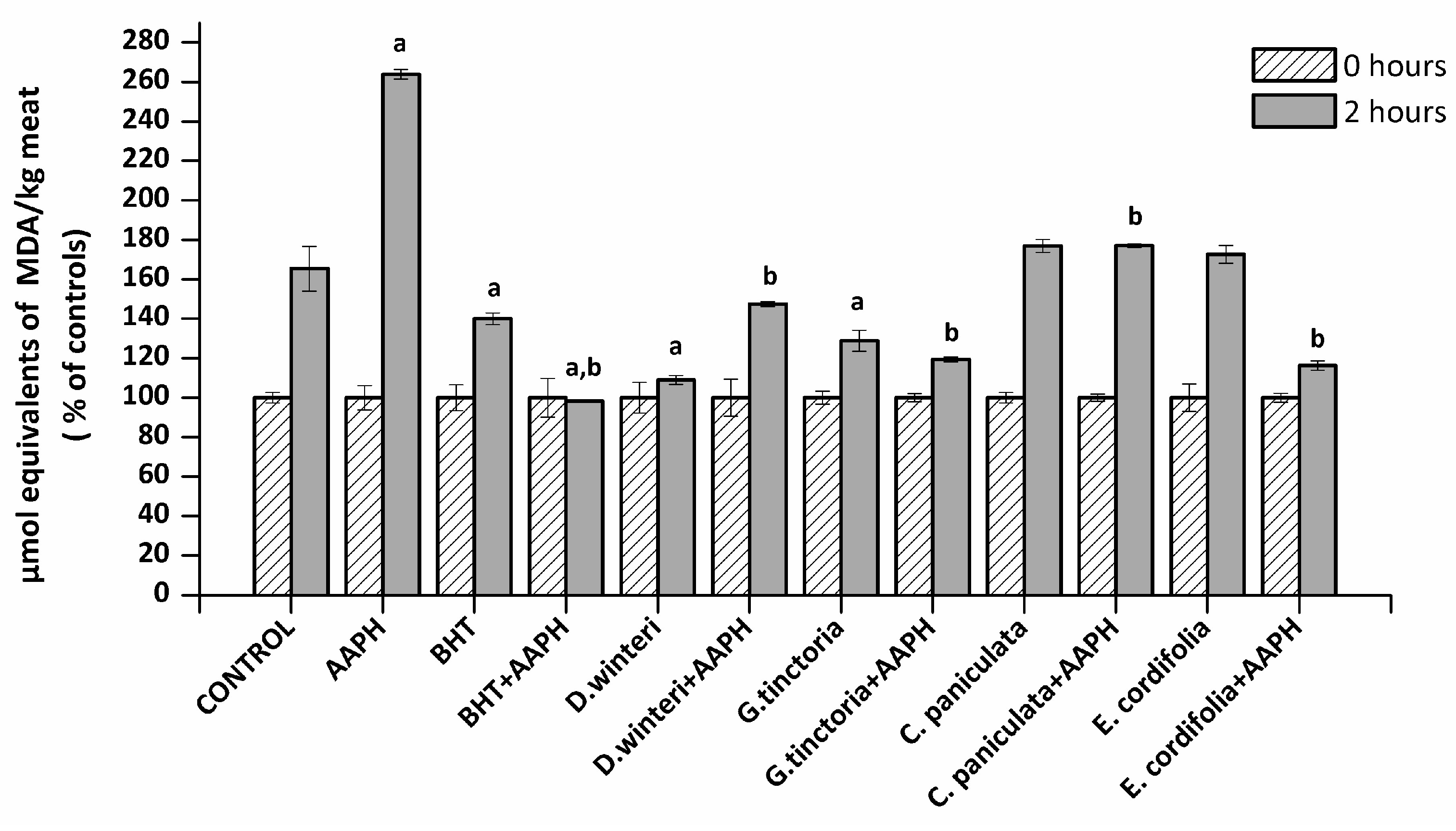
2.2. Lipid Oxidation in Meat Extract–TBARS
2.3. Protein Oxidation
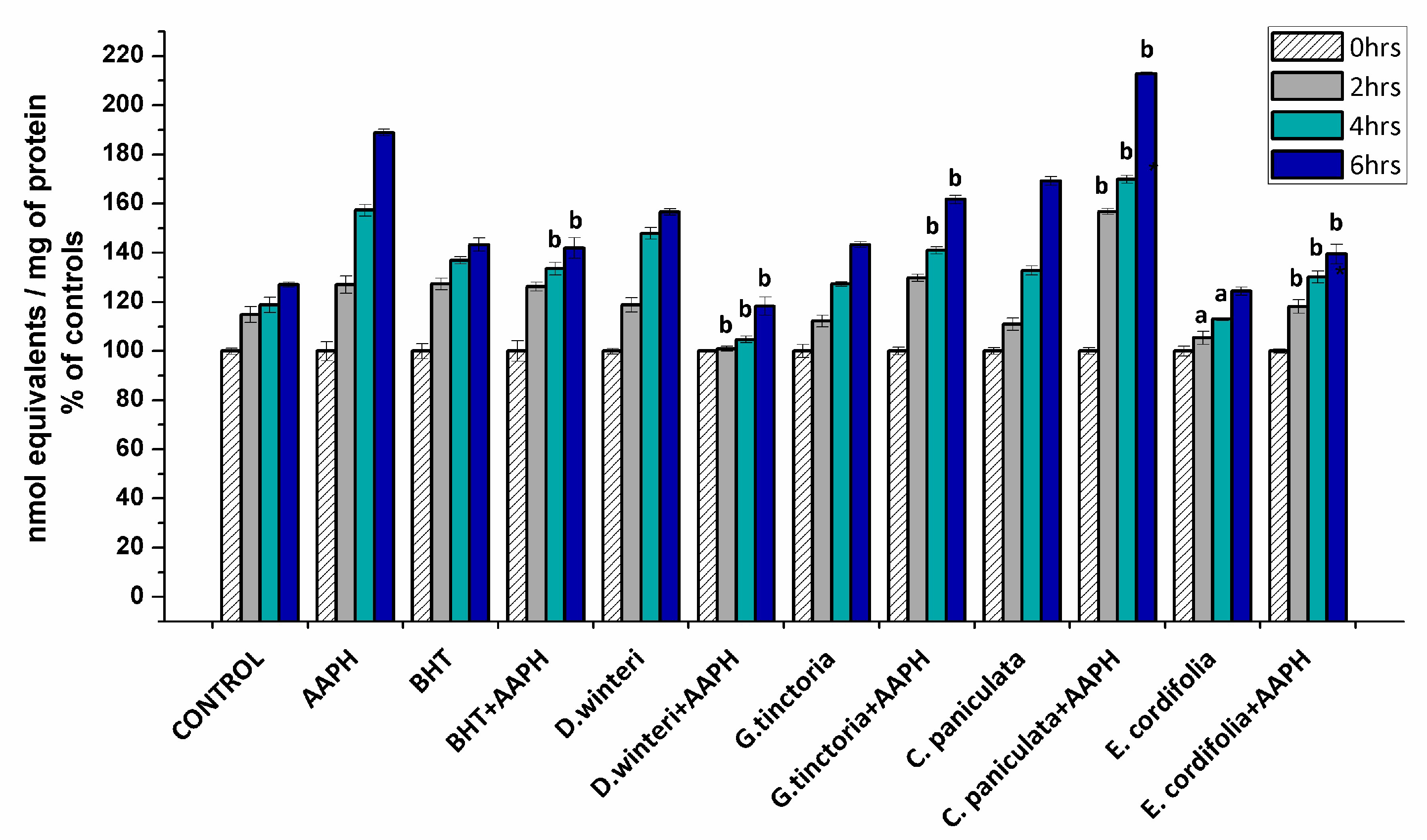
2.3.1. Determination of Carbonyl Content in Myofibrillar Proteins
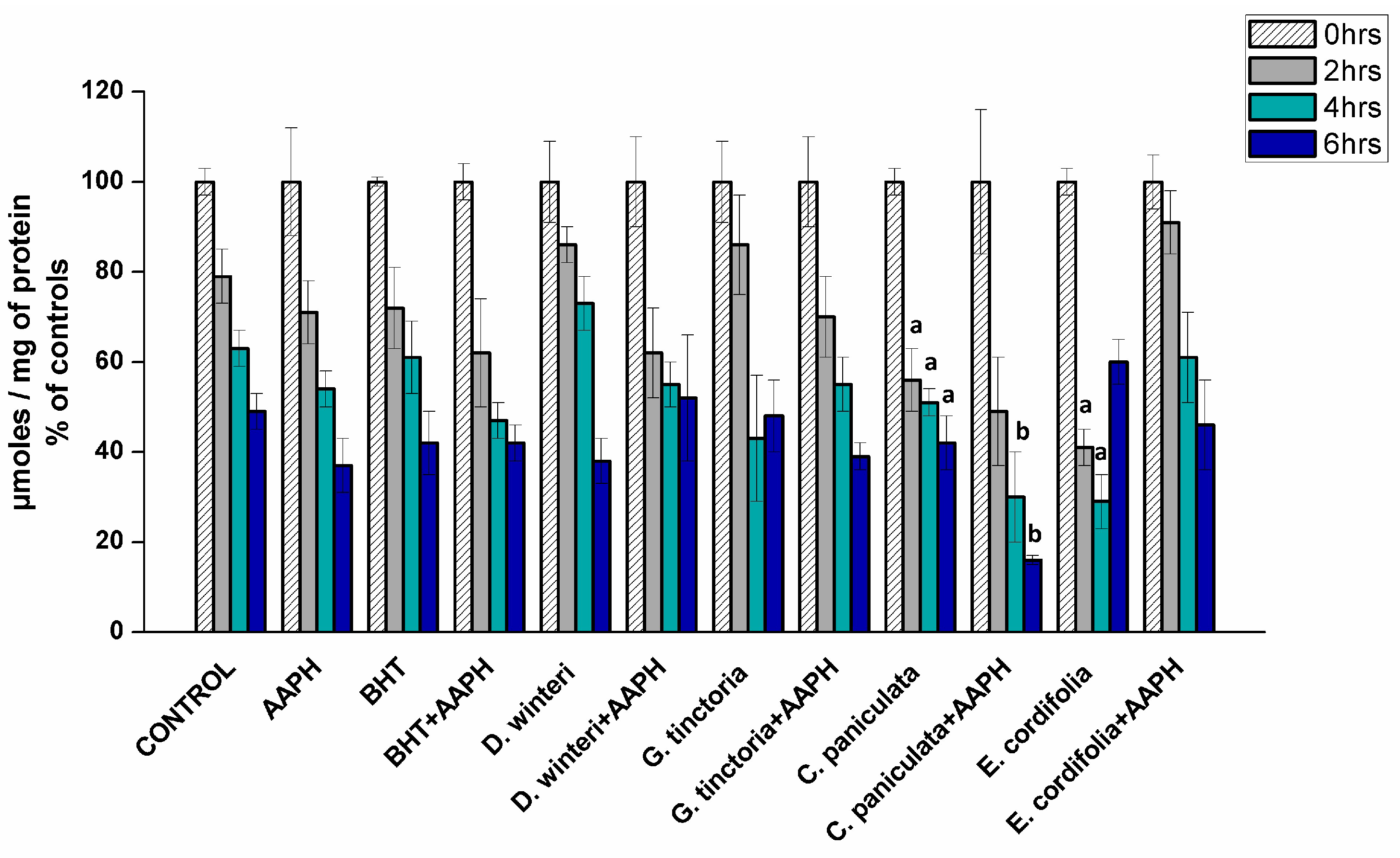
2.3.2. Determination of Total Sulfhydryl Groups in Myofibrillar Proteins
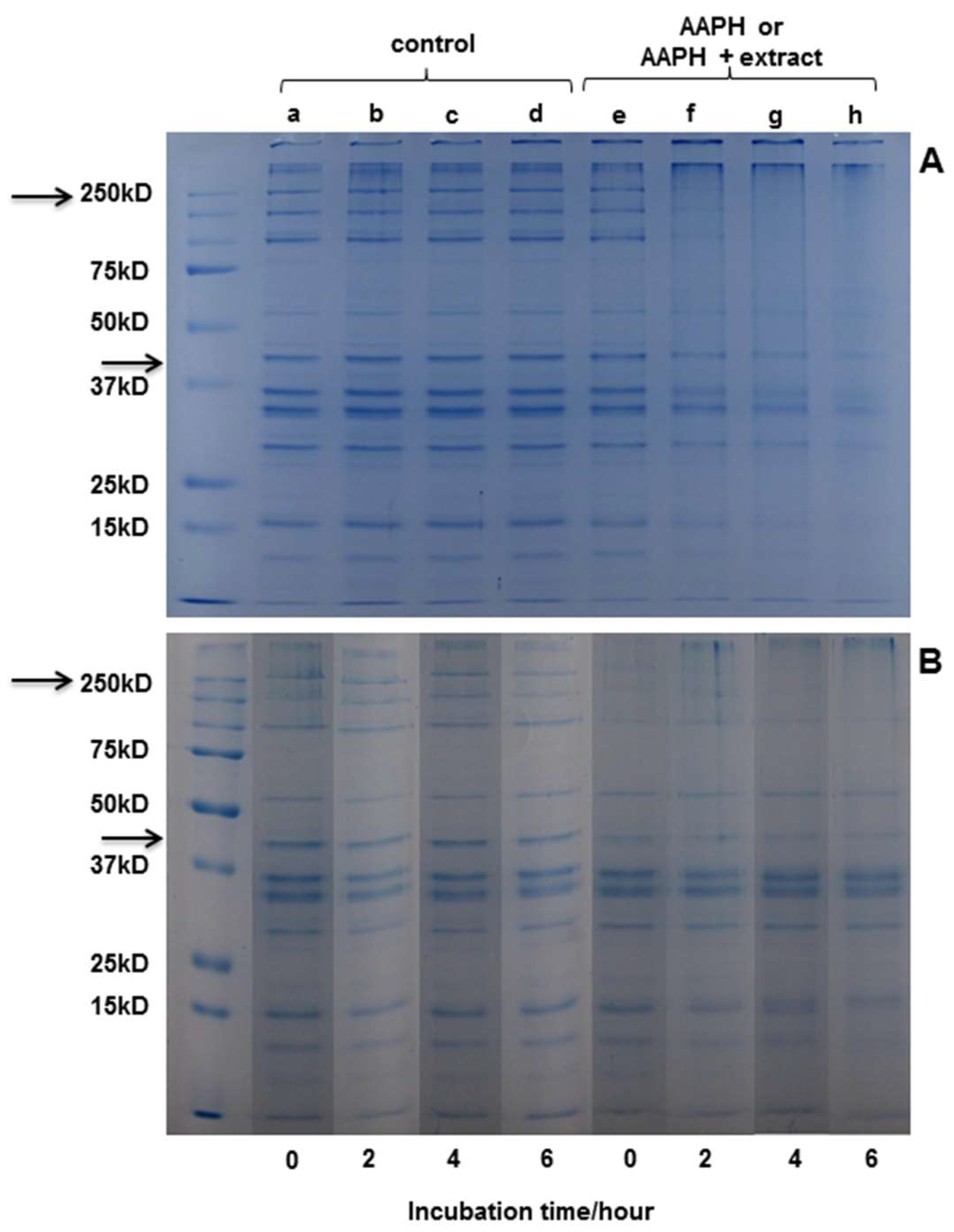
2.3.3. SDS-PAGE Electrophoresis
3. Materials and Methods
3.1. Plant Material
3.2. Plant Extracts
3.3. Total Phenolic Content (TPC)
3.4. Flavonoids Determination
3.5. Identification and Quantification of Flavonoids and Phenolic Acids
3.6. Determination of Antioxidant Capacity
3.7. Lipid Oxidation
3.7.1. Preparation and Incubation of Meat Extract
3.7.2. TBARS Determination
3.8. Protein Oxidation
3.8.1. Preparation and Incubation of Myofibrillar Proteins
3.8.2. Determination of Carbonyl Content
3.8.3. Determination of Total Sulfhydryl Groups
3.8.4. SDS-PAGE Electrophoresis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, J.R.; Liu, X.M.; Zhang, Y.S.; Zhang, Y.H.; Chen, Z.Y.; Tang, D.B.; Wang, J.Y. Protective effects of Momordica grosvenori extract against lipid and protein oxidation-induced damage in dried minced pork slices. Meat Sci. 2017, 133, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.K.; Laosinwattana, C.; Teerarak, M.; Pilasombut, K. Potential antioxidant and lipid peroxidation inhibition of Phyllanthus acidus leaf extract in minced pork. Asian Australas. J. Anim. Sci. 2017, 30, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M.; Kylli, P.; Puolanne, E.; Kivikari, R.; Heinonen, M. Fluorescence spectroscopy as a novel approach for the assessment of myofibrillar protein oxidation in oil-in-water emulsions. Meat Sci. 2008, 80, 1290–1296. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zhao, M.; Zhao, H.; Sun, W.; Cui, C. Effects of oxidative modification on gel properties of isolated porcine myofibrillar protein by peroxyl radicals. Meat Sci. 2014, 96, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Guyon, C.; Meynier, A.; de Lamballerie, M. Protein and lipid oxidation in meat: A review with emphasis on high-pressure treatments. Trends Food Sci. Technol. 2016, 50, 131–143. [Google Scholar] [CrossRef]
- Bañón, S.; Díaz, P.; Rodríguez, M.; Garrido, M.D.; Price, A. Ascorbate, green tea and grape seed extracts increase the shelf life of low sulphite beef patties. Meat Sci. 2007, 77, 626–633. [Google Scholar] [CrossRef]
- Lara, M.S.; Gutierrez, J.I.; Timón, M.; Andrés, A.I. Evaluation of two natural extracts (Rosmarinus officinalis L. and Melissa officinalis L.) as antioxidants in cooked pork patties packed in MAP. Meat Sci. 2011, 88, 481–488. [Google Scholar] [CrossRef]
- Karre, L.; Lopez, K.; Getty, K.J.K. Natural antioxidants in meat and poultry products. Meat Sci. 2013, 94, 220–227. [Google Scholar] [CrossRef]
- Pogorzelska-Nowicka, E.; Atanasov, A.G.; Horbańczuk, J.; Wierzbicka, A. Bioactive Compounds in Functional Meat Products. Molecules 2018, 23, 307. [Google Scholar] [CrossRef]
- Shah, M.A.; Bosco, S.J.D.; Mir, S.A. Plant extracts as natural antioxidants in meat and meat products. Meat Sci. 2014, 98, 21–33. [Google Scholar] [CrossRef]
- Bridi, R.; Montenegro, G. The Value of Chilean Honey: Floral Origin Related to their Antioxidant and Antibacterial activities. In Honey Analysis; Arnaut, V., Ed.; Intechopen: Rijeka, Croatia, 2017; pp. 63–78. [Google Scholar]
- Cechinel Filho, V.; Schlemper, V.; Santos, A.R.S.; Pinheiro, T.R.; Yunes, R.A.; Mendes, G.L.; Calixto, J.B.; Delle Monache, F. Isolation and identification of active compounds from Drimys winteri barks. J. Ethnopharmacol. 1998, 62, 223–227. [Google Scholar] [CrossRef]
- Mendes, G.L.; Santos, A.R.S.; Campos, M.M.; Tratsk, K.S.; Yunes, R.A.; Filho, V.C.; Calixto, J.B. Anti-hyperalgesic properties of the extract and of the main sesquiterpene polygodial isolated from the barks of Drymis winteri (Winteraceae). Life Sci. 1998, 63, 369–381. [Google Scholar] [CrossRef]
- Montenegro, G. Chile, Nuestra Flora Útil: Guía de Plantas de Uso Apícola, en Medicina Folklórica, Artesanal y Ornamental; Universidad Católica de Chile: Santiago, Chile, 2000. [Google Scholar]
- Barrero, A.; Herrador, M.; Arteaga, P.; Lara, A.; Cortés, M. Chemical Composition of the Essential Oil from Drimys winteri Forst. Wood. J. Essent. Oil Res. 2000, 12, 685–688. [Google Scholar] [CrossRef]
- Wollenweber, E.; Dörr, M.; Rozefelds, A.; Minchin, P.; Forster, P. Variation in flavonoid exudates in Eucryphia species from Australia and South America. Biochem. Syst. Ecol. 2000, 28, 111–118. [Google Scholar] [CrossRef]
- Pacheco, P.; Crawford, D.J.; Stuessy, T.F.; Silva, O.M. Flavonoid chemistry and evolution of Gunnera (Gunneraceae) in the Juan Fernandez Islands, Chile. Gayana Bot. 1993, 50, 17–28. [Google Scholar]
- Segovia, F.J.; Hidalgo, G.I.; Villasante, J.; Ramis, X.; Almajano, M.P. Avocado Seed: A Comparative Study of Antioxidant Content and Capacity in Protecting Oil Models from Oxidation. Molecules 2018, 23, 2421. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Kozakiewicz, Z. Ochratoxin A in grapes and grape-derived products. Trends Food Sci. Technol. 2006, 17, 72–81. [Google Scholar] [CrossRef]
- Wu, J.G.; Wang, P.J.; Chen, S.C. Antioxidant and antimicrobial effectiveness of catechin-impregnated pva–starch film on red meat. J. Food Qual. 2010, 33, 780–801. [Google Scholar] [CrossRef]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Lund, M.; Heinonen, M.; Baron, C.; Estévez, M. Protein oxidation in muscle foods: A review. Mol. Nutr. Food Res. 2011, 55, 83–95. [Google Scholar] [CrossRef]
- Estévez, M.; Heinonen, M. Effect of Phenolic Compounds on the Formation of α-Aminoadipic and γ-Glutamic Semialdehydes from Myofibrillar Proteins Oxidized by Copper, Iron, and Myoglobin. J. Agric. Food Chem. 2010, 58, 4448–4455. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed]
- Caro, A.A.; Davis, A.; Fobare, S.; Horan, N.; Ryan, C.; Schwab, C. Antioxidant and pro-oxidant mechanisms of (+) catechin in microsomal CYP2E1-dependent oxidative stress. Toxicol. In Vitro 2019, 54, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Platt, A.A.; Gieseg, S.P. Inhibition of protein hydroperoxide formation by protein thiols. Redox Rep. 2003, 8, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Jongberg, S.; Tørngren, M.; Skibsted, L. Dose-Dependent Effects of Green Tea or Maté Extracts on Lipid and Protein Oxidation in Brine-Injected Retail-Packed Pork Chops. Medicines 2018, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Bridi, R.; Montenegro, G.; Nuñez-Quijada, G.; Giordano, A.; Morán-Romero, M.F.; Jara-Pezoa, I.; Speisky, H.; Atala, E.; López-Alarcón, C. International regulations of propolis quality: Required assays do not necessarily reflect their polyphenolic-related in vitro activities. J. Food Sci. 2015, 80, C1188–C1195. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, P.; Rodriguez, K.; Retamal, M.; Giordano, A.; Valenzuela, L.M.; Montenegro, G. Relation between composition, antioxidant and antibacterial activities and botanical origin of multi-floral bee pollen. J. Appl. Bot. Food Qual. 2017, 90, 306–314. [Google Scholar]
- Esterbauer, H.; Cheeseman, K.H. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990; Volume 186, pp. 407–421. [Google Scholar]
- Martinaud, A.; Mercier, Y.; Marinova, P.; Tassy, C.; Gatellier, P.; Renerre, M. Comparison of Oxidative Processes on Myofibrillar Proteins from Beef during Maturation and by Different Model Oxidation Systems. J. Agric. Food Chem. 1997, 45, 2481–2487. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990; Volume 186, pp. 464–478. [Google Scholar]
- Lund, M.N.; Hviid, M.S.; Skibsted, L.H. The combined effect of antioxidants and modified atmosphere packaging on protein and lipid oxidation in beef patties during chill storage. Meat Sci. 2007, 76, 226–233. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Sun, W.; Cui, C.; Zhao, M.; Zhao, Q.; Yang, B. Effects of composition and oxidation of proteins on their solubility, aggregation and proteolytic susceptibility during processing of Cantonese sausage. Food Chem. 2011, 124, 336–341. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Total Phenolic Content (mg GAE/100g DW) | Flavonoid Content (mg QE/100g DW) | ORAC-PGR (µmol TE/100g DW) | FRAP (mg FeSO4·7H2O /100g DW) |
|---|---|---|---|---|
| D. winteri (canelo) | 438 ± 16 a | 90 ± 0.8 a | 14 ± 0.4 a | 95 ± 1.1 a |
| G. tinctoria (nalca) | 211 ± 15 b | 64 ± 0.1 b | 5 ± 1.2 b | 55 ± 0.9 b |
| C. paniculata (tiaca) | 231 ± 3 b | 47 ± 0.3 c | 3 ± 0.9 b | 61 ± 1.6 c |
| E. cordifolia (ulmo) | 91 ± 1 c | 89 ± 0.7 a | 5 ± 1.1 b | 27 ± 1.1 d |
| Concentration (mg / kg DW) 1 | ||||
|---|---|---|---|---|
| D. winteri (canelo) | G. tinctoria (nalca) | C. paniculata (tiaca) | E. cordifolia (ulmo) | |
| Gallic acid | 3.88 ± 0.39 | 2.96 ± 0.24 | 0.62 ± 0.06 | 9.03 ± 0.33 |
| Chlorogenic acid | 0.81 ± 0.05 | ND | 1.13 ± 0.07 | ND |
| Caffeic acid | 0.20 ± 0.01 | 0.12 ± 0.01 | 0.28 ± 0.02 | 4.15 ± 0.68 |
| Coumaric acid | 0.84 ± 0.03 | 0.12 ± 0.01 | 0.80 ± 0.03 | 1.37 ± 0.20 |
| Catechin | 52.1 ± 10.2 | 188.4 ± 15.1 | 519.4 ± 22.3 | 0.30 ± 0.03 |
| Pinocembrin | 0.06 ± 0.005 | ND | ND | 0.06 ± 0.01 |
| Rutin | 1.33 ± 0.13 | 5.47 ± 0.33 | 1.36 ± 0.03 | 40.4 ± 4.0 |
| Chrysin | 0.07 ± 0.01 | ND | ND | ND |
| Quercetin | 2.98 ± 0.18 | ND | 10.15 ± 0.61 | 7.37 ± 0.67 |
| Abscisic acid | ND | 0.09 ± 0.01 | 0.08 ± 0.05 | 0.10 ± 0.01 |
| Luteolin | 0.30 ± 0.03 | ND | ND | ND |
| Epicatechin | 63.66 ± 1.14 | 197.0 ± 11.8 | 491.4 ± 3.6 | 0.54 ± 0.04 |
| Apigenin | 0.26 ± 0.01 | 0.24 ± 0.02 | 0.30 ±0.01 | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bridi, R.; Giordano, A.; Peñailillo, M.F.; Montenegro, G. Antioxidant Effect of Extracts from Native Chilean Plants on the Lipoperoxidation and Protein Oxidation of Bovine Muscle. Molecules 2019, 24, 3264. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24183264
Bridi R, Giordano A, Peñailillo MF, Montenegro G. Antioxidant Effect of Extracts from Native Chilean Plants on the Lipoperoxidation and Protein Oxidation of Bovine Muscle. Molecules. 2019; 24(18):3264. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24183264
Chicago/Turabian StyleBridi, Raquel, Ady Giordano, Maria Fernanda Peñailillo, and Gloria Montenegro. 2019. "Antioxidant Effect of Extracts from Native Chilean Plants on the Lipoperoxidation and Protein Oxidation of Bovine Muscle" Molecules 24, no. 18: 3264. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24183264