Evaluation of Antifungal Phenolics from Helianthus tuberosus L. Leaves against Phytophthora capsici Leonian by Chemometric Analysis

Abstract
:1. Introduction
2. Results
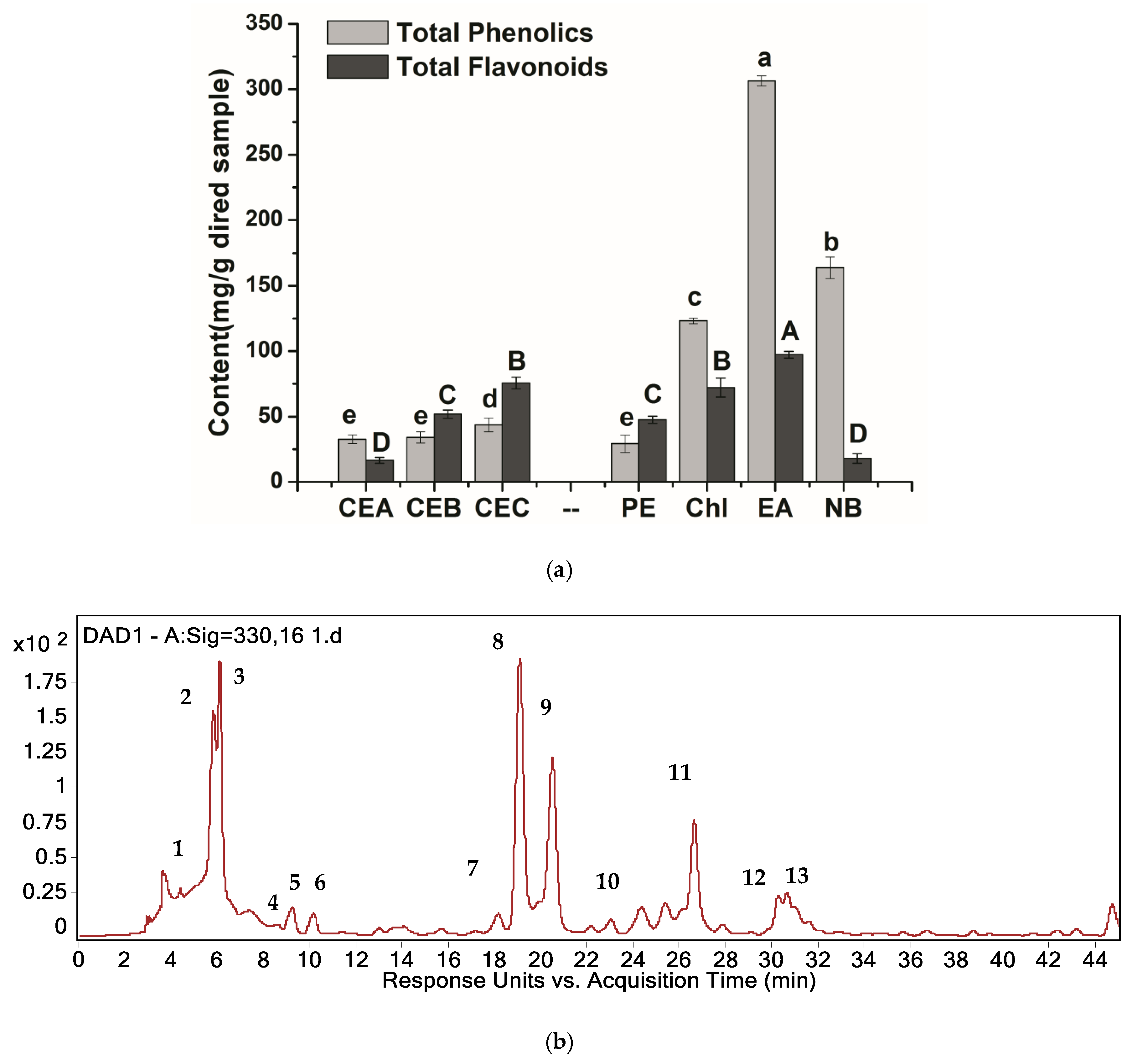
2.1. Analysis of Major Components in H. tuberosus Leaves
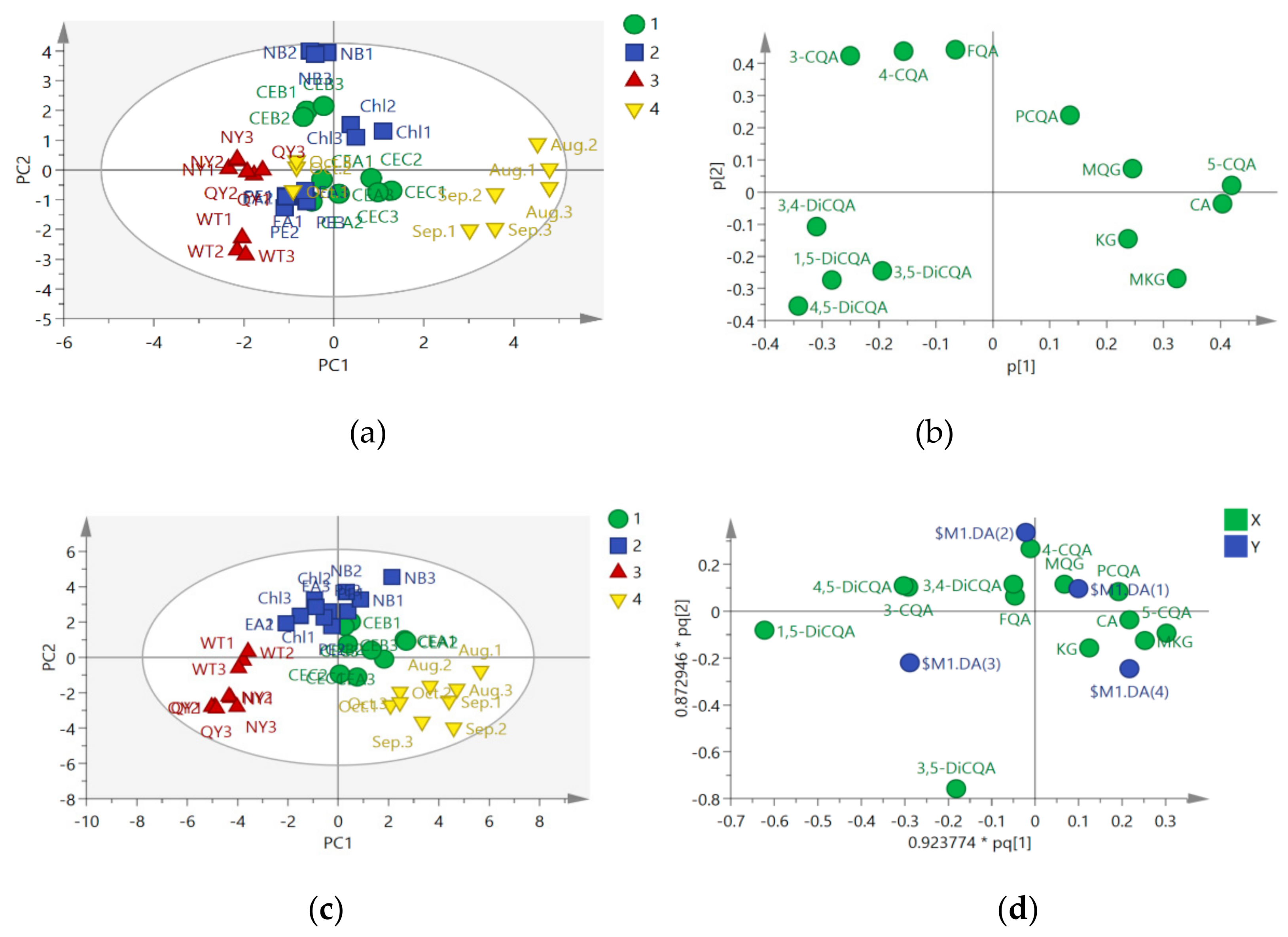
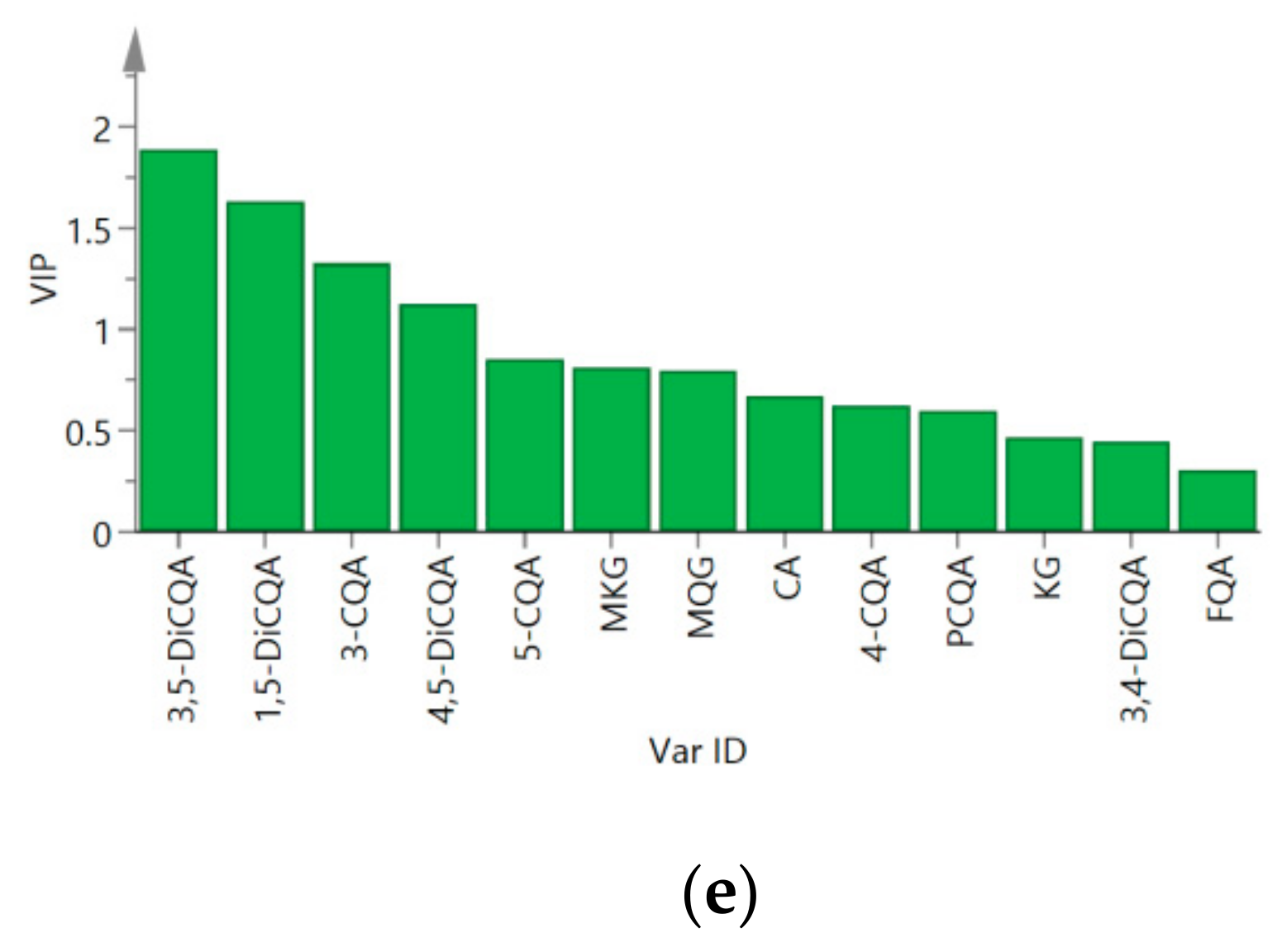
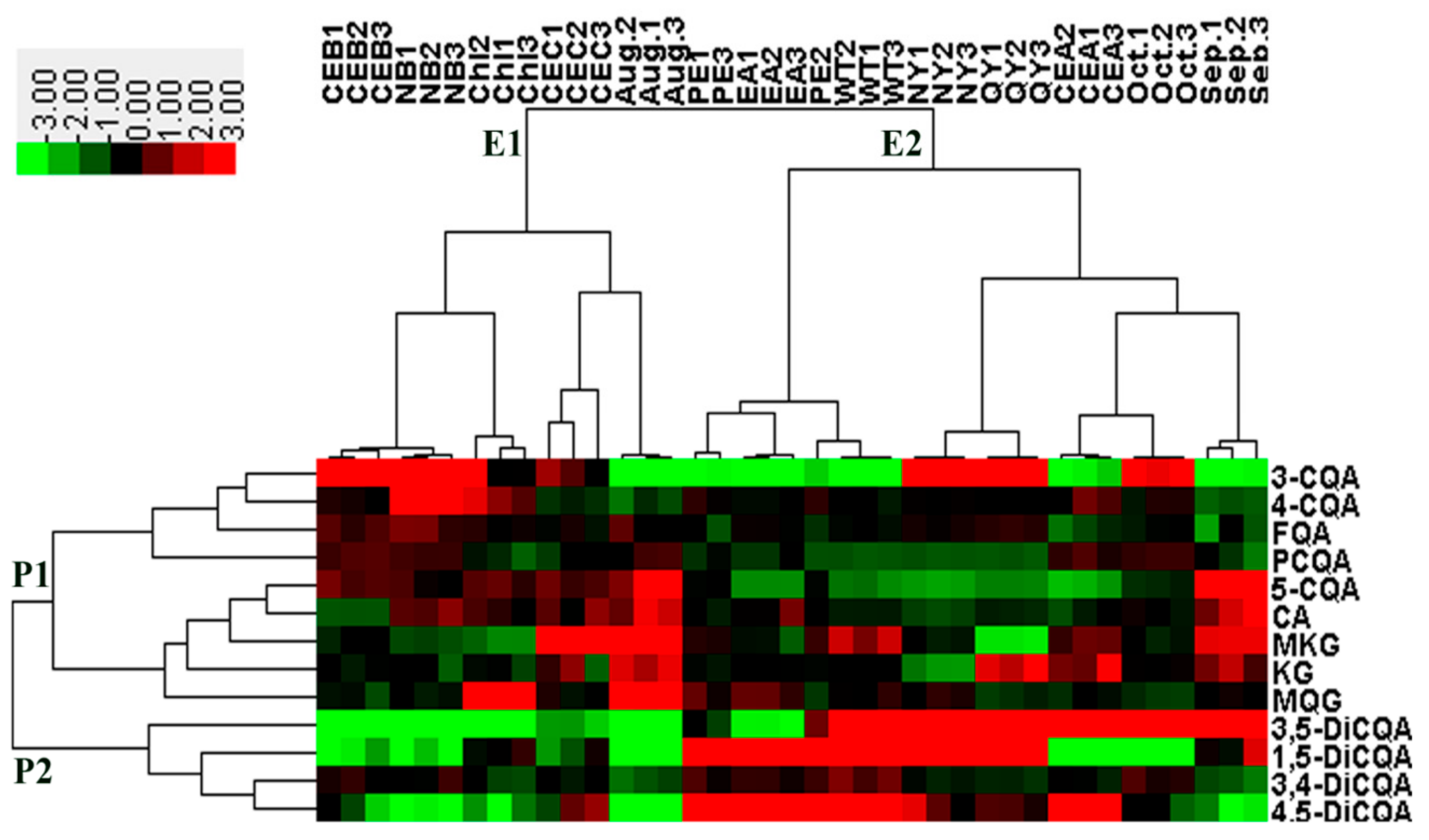
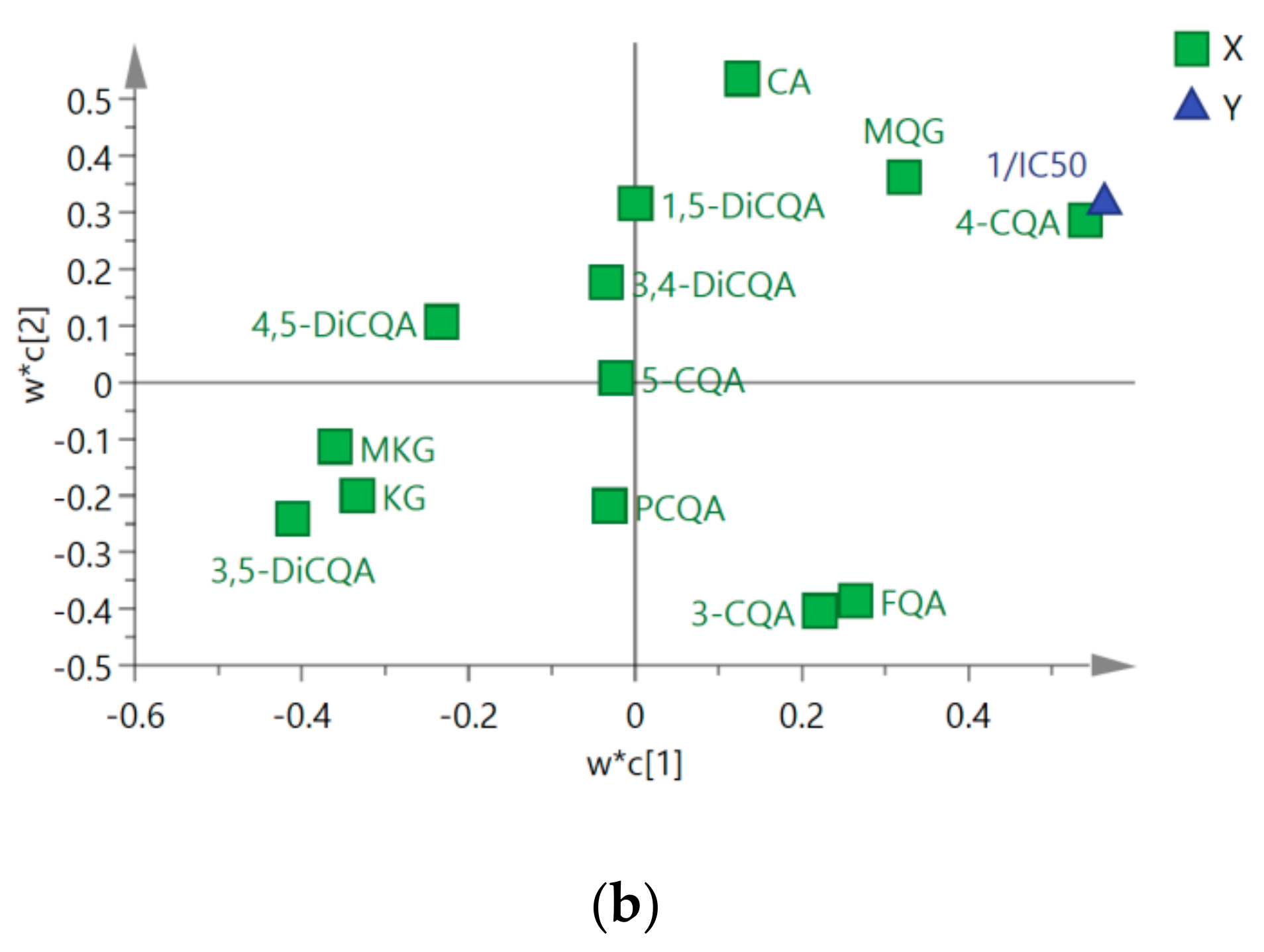
2.2. Chemometric Determination of Phenolic Acids
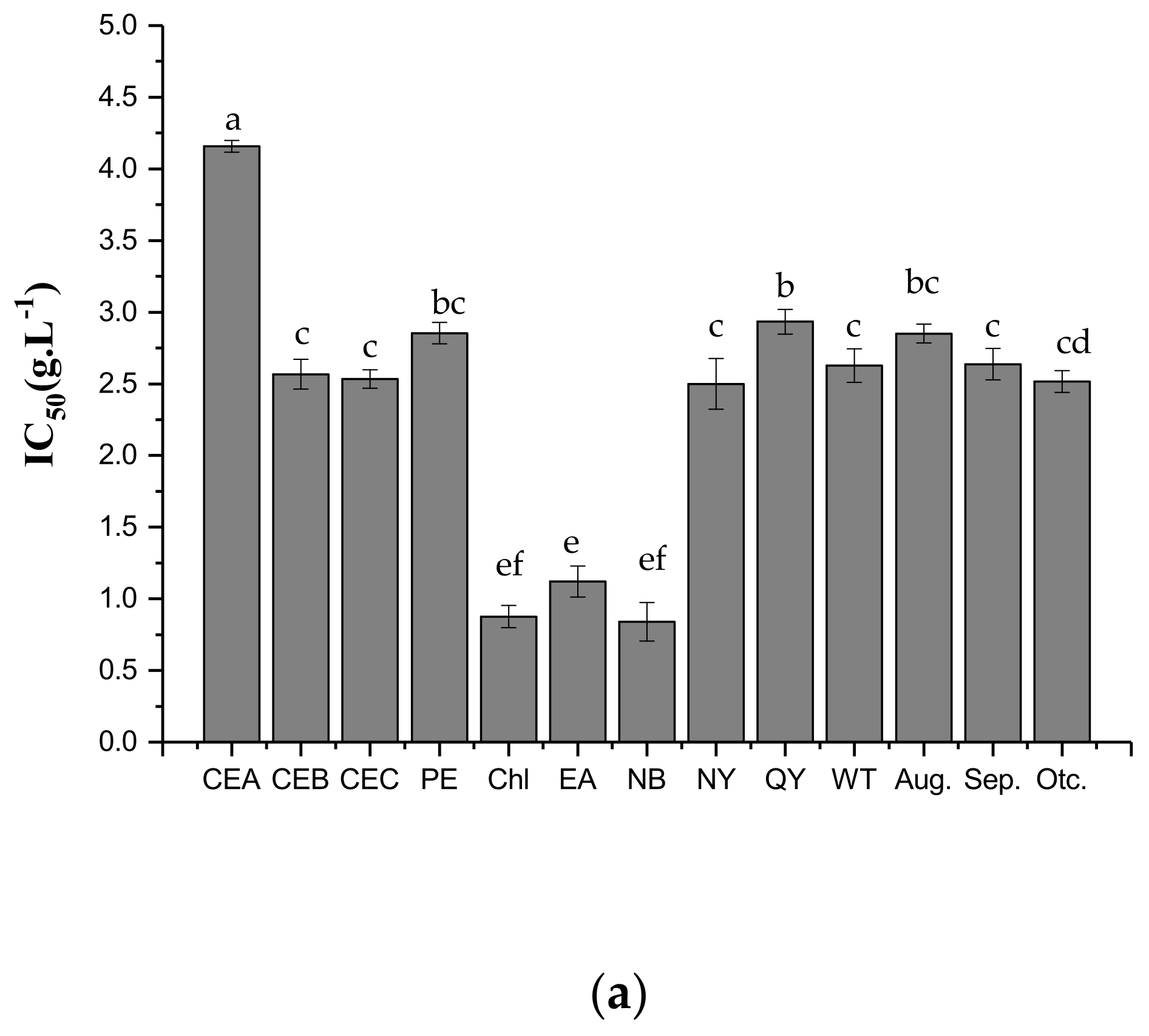
2.3. Antifungal Activities of Different Extracts and PLS-DA
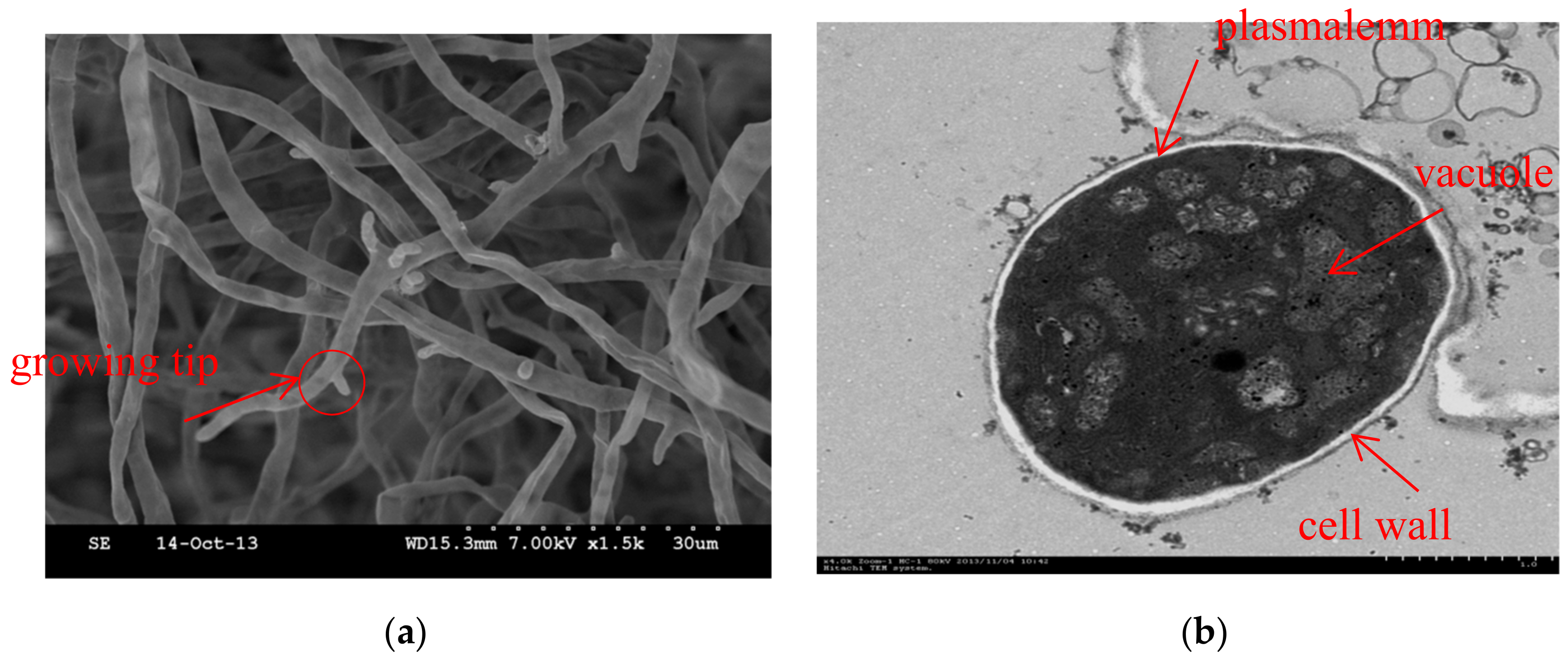
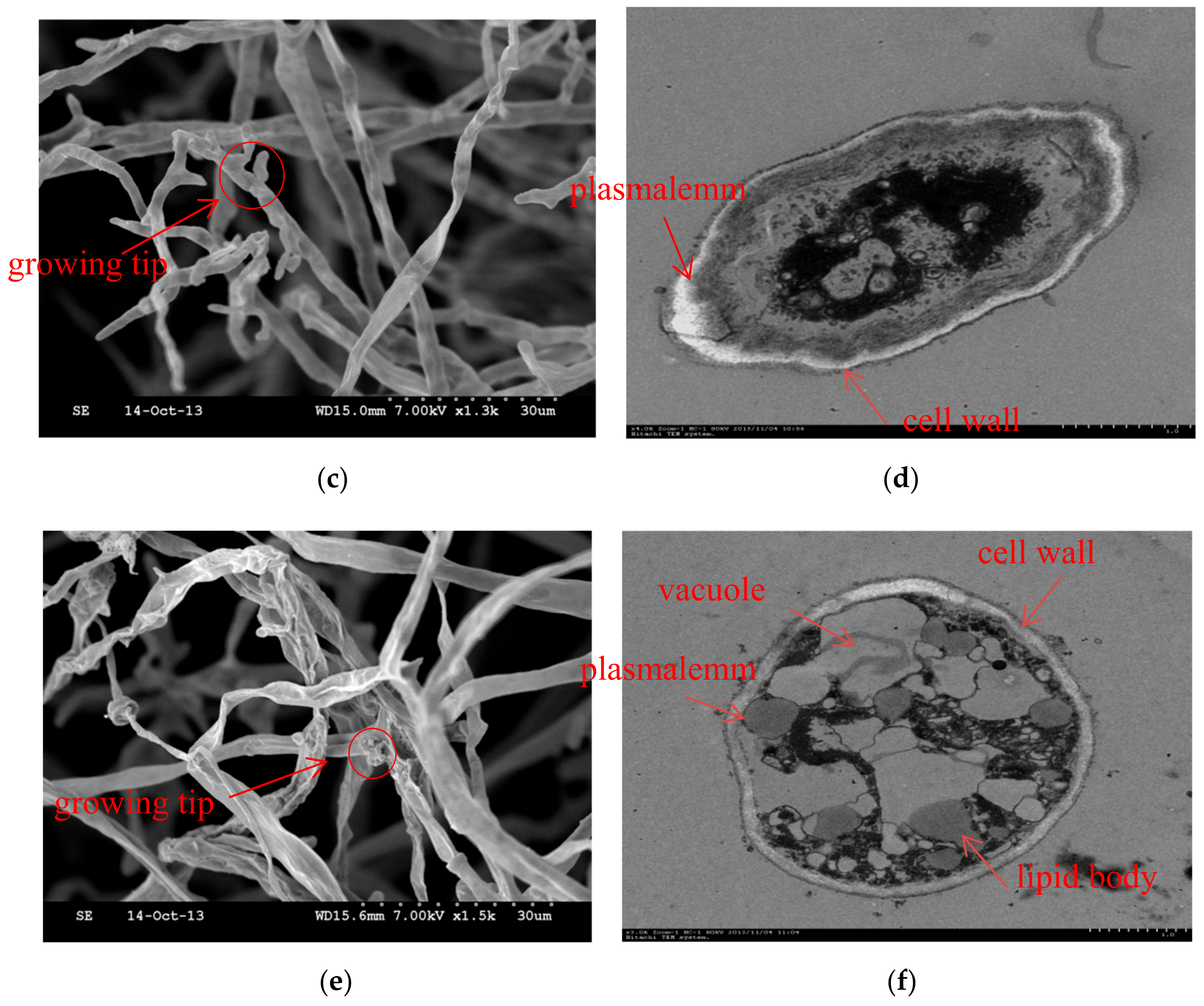
2.4. Scanning Electron Microscopy
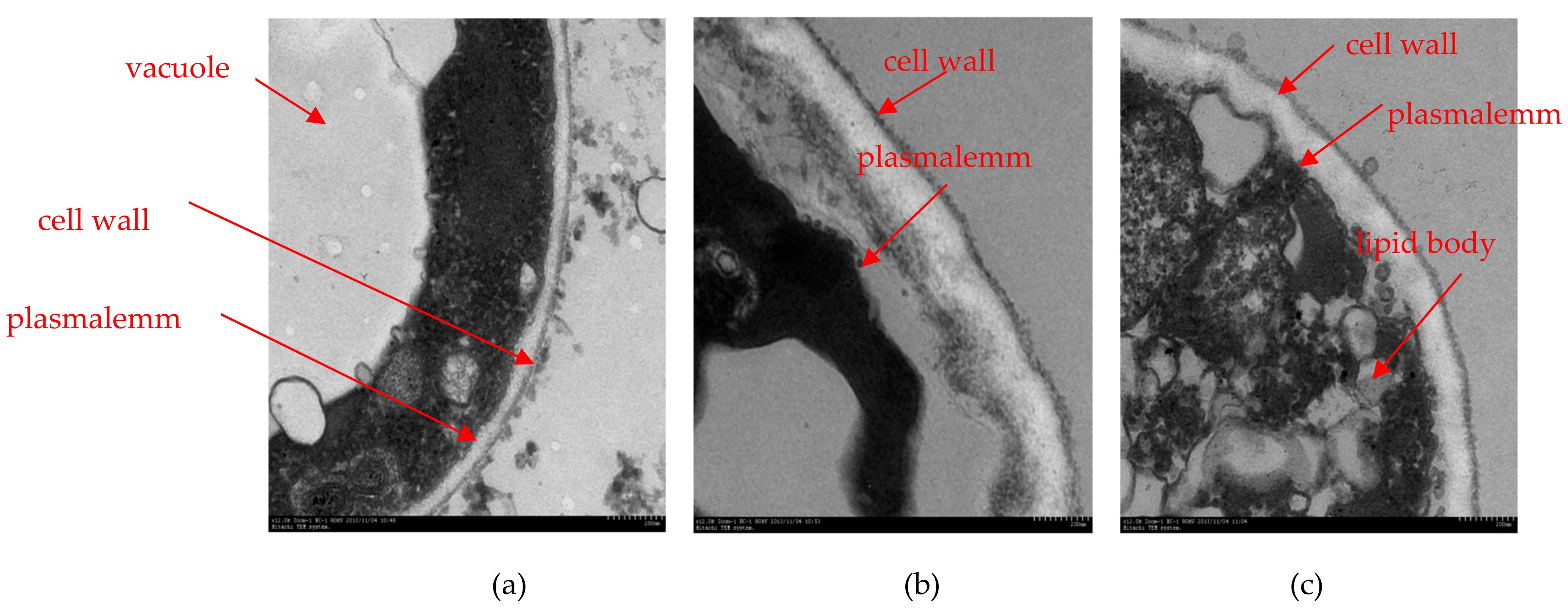
2.5. Transmission Electron Microscopy
3. Discussion
4. Materials and Methods
4.1. Chemicals and Plants
4.2. Preparation of Extracts
4.3. HPLC-MS/MS Analysis
4.4. Measurement of TPC and TFC
4.5. Microorganism
4.6. Antifungal Assay
4.7. SEM
4.8. TEM
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Lee, B.K.; Kim, B.S.; Chang, S.W.; Hwang, B.K. Aggressiveness to pumpkin cultivars of isolates of Phytophthora capsici from pumpkin and pepper. Plant Dis. 2001, 85, 497–500. [Google Scholar] [CrossRef]
- Lamour, K.H.; Hausbeck, M.K. Mefenoxam insensitivity and the sexual stage of Phytophthora capsici in Michigan cucurbit fields. Phytopathology. 2000, 90, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Lee, J.Y.; Hwang, B.K. In vivo control and in vitro antifungal activity of rhamnolipid B, a glycolipid antibiotic, against Phytophthora capsici and Colletotrichum orbiculare. Pest Manag. Sci. 2000, 56, 1029–1035. [Google Scholar] [CrossRef]
- Chen, F.; Long, X.; Yu, M.; Liu, Z.; Liu, L.; Shao, H. Phenolics and antifungal activities analysis in industrial crop jerusalem artichoke (Helianthus Tuberosus L.) leaves. Ind. Crop. Prod. 2013, 47, 339–345. [Google Scholar] [CrossRef]
- Chen, F.; Long, X.; Liu, Z.; Shao, H.; Liu, L. Analysis of phenolic acids of jerusalem artichoke (Helianthus Tuberosus L.) responding to salt-stress by liquid chromatography/tandem mass spectrometry. Scientific World J. 2014, 2014, 568043. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Sinden, M.R.; Kennedy, A.H.; Chai, H.; Watson, L.E.; Graham, T.L.; Kinghorn, A.D. Bioactive constituents of Helianthus tuberosus (Jerusalem artichoke). Phytochem. Lett. 2009, 2, 15–18. [Google Scholar] [CrossRef]
- Vokou, D.; Chalkos, D.; Karamanoli, K. Microorganisms and allelopathy: A one-sided approach. In Allelopathy; Springer: Dordrecht, The Netherlands, 2006; pp. 341–371. [Google Scholar]
- Jabeen, R.; Shahid, M.; Jamil, A.; Ashraf, M. Microscopic evaluation of the antimicrobial activity of seed extracts of Moringa oleifera. Pak J. Bot. 2008, 40, 1349–1358. [Google Scholar]
- Liu, X.; Zhan, H.; Qiao, Z.; Zheng, M.; Liu, W.; Feng, F.; Yan, F. Chemometric analysis based on HPLC multi-wavelength fingerprints for prediction of antioxidant components in Turpiniae Folium. Chemometr. Intell. Lab. 2016, 152, 54–61. [Google Scholar] [CrossRef]
- Goh, L.M.; Barlow, P.J. Flavonoid recovery and stability from Ginkgo biloba subjected to a simulated digestion process. Food Chem. 2004, 86, 195–202. [Google Scholar] [CrossRef]
- Fraser, K.; Lane, G.A.; Otter, D.E.; Harrison, S.J.; Quek, S.Y.; Hemar, Y.; Rasmussen, S. Non-targeted analysis by LC-MS of major metabolite changes during the oolong tea manufacturing in New Zealand. Food Chem. 2014, 151, 394–403. [Google Scholar] [CrossRef]
- Fu, J.H.; Tang, J.; Liao, X.; Gao, J. Study on composition-activity relationship of lavender essential oil based on antibacterial activity. J. Chinese Mass Spectrom. Soc. 2015, 36, 403–410. [Google Scholar]
- Jin, Y.; Ma, Y.; Xie, W.; Hou, L.; Xu, H.; Zhang, K.; Zhang, L.; Du, Y. UHPLC-Q-TOF-MS/MS-oriented characteristic components dataset and multivariate statistical techniques for the holistic quality control of Usnea. RSC Adv. 2018, 8, 15487–15500. [Google Scholar] [CrossRef]
- Tan, T.; Zhang, J.; Xu, X.; Huang, W.P.; Luo, Y. Geographical discrimination of Glechomae Herba based on fifteen phenolic constituents determined by LC-MS/MS method combined with chemometric methods. Biomed. Chromatogr. 2018, 32, e4239. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Kim, J.K.; Lee, J.K.; Kim, S.H. Discrimination of geographical origin of rice (Oryza sativa L.) by multielement analysis using inductively coupled plasma atomic emission spectroscopy and multivariate analysis. J. Cereal Sci. 2015, 65, 252–259. [Google Scholar] [CrossRef]
- Xiao, R.; Ma, Y.; Zhang, D.; Qian, L. Discrimination of conventional and organic rice using untargeted lc-ms-based metabolomics. J. Cereal Sci. 2018, 82, 73–81. [Google Scholar] [CrossRef]
- Ngadze, R.T.; Verkerk, R.; Nyanga, L.K.; Fogliano, V.; Ferracane, R.; Troise, A.D.; Linnemann, A.R. Effect of heat and pectinase maceration on phenolic compounds and physicochemical quality of Strychnos cocculoides juice. PLoS ONE 2018, 13, e0202415. [Google Scholar] [CrossRef]
- Kong, D.; Li, Y.; Bai, M.; Deng, Y.; Liang, G.; Wu, H. A comparative study of the dynamic accumulation of polyphenol components and the changes in their antioxidant activities in diploid and tetraploid Lonicera japonica. Plant Physiol. Bioch. 2017, 112, 87–96. [Google Scholar] [CrossRef]
- Duke, S.O.; Baerson, S.R.; Dayan, F.E.; Rimando, A.M.; Scheffler, B.E.; Tellez, M.R.; Wedge, D.E.; Schrader, K.K.; Akey, D.H.; Arthur, F.H.; et al. United States Department of Agriculture–Agricultural Research Service research on natural products for pest management. Pest Manag. Sci. 2003, 59, 708–717. [Google Scholar] [CrossRef]
- Widmer, T.L.; Laurent, N. Plant extracts containing caffeic acid and rosmarinic acid inhibit zoospore germination of Phytophthora spp. pathogenic to Theobroma cacao. Eur. J. Plant Pathol. 2006, 115, 377–388. [Google Scholar] [CrossRef]
- Martínez, J.A. Natural fungicides obtained from plants. In Fungicides for Plant and Animal Diseases; Dhanasekaran, D., Thajuddin, N., Panneerselvam, A., Eds.; IntechOpen: Rijeka, Croatia, 2012; pp. 3–28. [Google Scholar] [CrossRef]
- Koley, T.K.; Singh, S.; Khemariya, P.; Sarkar, A.; Kaur, C.; Chaurasia, S.N.S.; Naik, P.S. Evaluation of bioactive properties of Indian carrot (Daucus carota L.): A chemometric approach. Food Res. Int. 2014, 60, 76–85. [Google Scholar] [CrossRef]
- Chan, K.M.; Yue, G.G.; Li, P.; Wong, E.C.; Lee, J.K.; Kennelly, E.J.; Lau, C.B. Screening and analysis of potential anti-tumor components from the stipe of Ganoderma sinense using high-performance liquid chromatography/time-of-flight mass spectrometry with multivariate statistical tool. J. Chromatogr. A 2017, 1487, 162–167. [Google Scholar] [CrossRef]
- Farag, M.A.; Sakna, S.T.; Elfiky, N.M.; Shabana, M.M.; Wessjohann, L.A. Phytochemical, antioxidant and antidiabetic evaluation of eight Bauhinia L. species from Egypt using UHPLC-PDA-qTOF-MS and chemometrics. Phytochemistry 2015, 119, 41–50. [Google Scholar] [CrossRef]
- Semeniuc, C.; Socaciu, M.I.; Socaci, S.; Mureșan, V.; Fogarasi, M.; Rotar, A. Chemometric comparison and classification of some essential oils extracted from plants belonging to Apiaceae and Lamiaceae families based on their chemical composition and biological activities. Molecules 2018, 23, 2261. [Google Scholar] [CrossRef] [PubMed]
- Chua, L.S.; Lau, C.H.; Chew, C.Y.; Ismail, N.; Soontorngun, N. Phytochemical profile of Orthosiphon aristatus extracts after storage: rosmarinic acid and other caffeic acid derivatives. Phytomedicine 2018, 39, 49. [Google Scholar] [CrossRef] [PubMed]
- D’Urso, G.; Pizza, C.; Piacente, S.; Montoro, P. Combination of LC-MS based metabolomics and antioxidant activity for evaluation of bioactive compounds in Fragaria vesca leaves from Italy. J. Pharmaceut. Biomed. 2018, 150, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Čižmárik, J.; Matel, I. Examination of the chemical composition of propolis 2. Isolation and identification of 4-hydroxy-3-methoxycinnamic acid (ferulic acid) from propolis. J. Apicult. Res. 1973, 12, 52–54. [Google Scholar] [CrossRef]
- Wen, A.; Delaquis, P.; Stanich, K.; Toivonen, P. Antilisterial activity of selected phenolic acids. Food Microb. 2003, 20, 305–311. [Google Scholar] [CrossRef]
- Furuhata, K.; Dogasaki, C.; Hara, M.; Fukuyama, M. Inactivation of Legionella pneumophila by phenol compounds contained in coffee. Bokin Bobai. 2002, 30, 291–298. [Google Scholar]
- Wu, W.J. From Natural Products to the New Pesticide-Principle and Method; Chem. Ind. Press: Beijing, China, 2006; p. 69. ISBN 7-5025-8243-6. [Google Scholar]
- Yan, X.; Qin, W.; Sun, L.; Qi, S.; Yang, D.; Qin, Z.; Yuan, H. Study of inhibitory effects and action mechanism of the novel fungicide pyrimorph against Phytophthora capsici. J. Agric. Food chem. 2009, 58, 2720–2725. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J. Fungal Cell Wall; Structure, Synthesis, and Assembly, 2nd ed.; Ringgold Inc: Portland, OR, USA, 2012. [Google Scholar]
- Yamaguchi, M.U.; Garcia, F.P.; Cortez, D.A.G.; Ueda-Nakamura, T.; Filho, B.P.D.; Nakamura, C.V. Antifungal effects of Ellagitannin isolated from leaves of Ocotea odorifera(Lauraceae). Antonie van Leeuwenhoek 2011, 99, 507. [Google Scholar] [CrossRef]
- Ozgonen, H.; Yardimci, N.; Kilic, H.C. Induction of phenolic compounds and pathogenesis-related proteins by mycorrhizal fungal inoculations against Phytophthora capsici Leonian in pepper. Pak. J. Biol. Sci. 2009, 12, 1181. [Google Scholar] [CrossRef] [PubMed]
- Koc, E.; Üstün, A.S. Differential Induction of Phenylalanine Ammonia Lyase and Phenolics in Peppers (Capsicum annuum) in Response to Inoculation with Phytophthora capsici. Int. J. Agr. Biol. 2011, 13, 881–887. [Google Scholar]
- Wang, T.; Wang, G.; Jia, Z.H.; Pan, D.L.; Zhang, J.Y.; Guo, Z.R. Transcriptome analysis of kiwifruit in response to Pseudomonas syringae pv. actinidiae infection. Int. J. Mol. Sci. 2018, 19, 373. [Google Scholar] [CrossRef] [PubMed]
- Tawaha, K.; Alali, F.Q.; Gharaibeh, M.; Mohammad, M.; El-Elimat, T. Antioxidant activity and total phenolic content of selected Jordanian plant species. Food Chem. 2007, 104, 1372–1378. [Google Scholar] [CrossRef]
- Zhou, K.; Yu, L. Total phenolic contents and antioxidant properties of commonly consumed vegetables grown in Colorado. LWT-Food Sci. Technol. 2006, 39, 1155–1162. [Google Scholar] [CrossRef]
- Rauter, A.P.; Martins, A.; Borges, C.; Ferreira, J.; Justino, J.; Bronze, M.; Coelho, A.V.; Choi, Y.H.; Verpoorte, R. Liquid chromatography–diode array detection–electrospray ionisation mass spectrometry/nuclear magnetic resonance analyses of the anti-hyperglycemic flavonoid extract of Genista tenera structure elucidation of a flavonoid-C-glycoside. J. Chromatogr. A 2005, 1089, 59–64. [Google Scholar] [CrossRef]
- Mihai, C.M.; Mărghitaş, L.A.; Dezmirean, D.S.; Chirilă, F.; Moritz, R.F.; Schlüns, H. Interactions among flavonoids of propolis affect antibacterial activity against the honeybee pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2012, 110, 68–72. [Google Scholar] [CrossRef]
- Chander, P.A.; Sri, H.Y.; Sravanthi, N.; Susmitha, U.V. In vitro anthelmintic activity of Barleria buxifolia on Indian adult earthworms and estimation of total flavonoid content. Asian Pac. J. Trop. Dis. 2014, 4, S233–S235. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | 5-CQA | 3-CQA | 4-CQA | CA | PCQA | FQA | 3,4-DiCQA | 3,5-DiCQA | 1,5-DiCQA | MQG | 4,5-DiCQA | MKG | KG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CEA | 0.49 ± 0.06 g | 14.14 ± 0.12 g | 2.51 ± 0.43 bc | 1.26 ± 0.19 def | 2.52 ± 0.41 a | 1.11 ± 0.16 de | 2.96 ± 0.53 bc | 23.76 ± 0.19 c | 3.43 ± 0.19 j | 1.54 ± 0.23 de | 17.08 ± 0.65 b | 5.27 ± 0.16 c | 4.19 ± 0.67 a |
| CEB | 3.20 ± 0.24 c | 25.64 ± 0.72 c | 1.89 ± 0.09 de | 0.48 ± 0.11 f | 2.53 ± 0.23 a | 2.49 ± 0.27 ab | 3.18 ± 0.17 abc | 6.42 ± 0.37 h | 8.37 ± 0.75 g | 1.13 ± 0.25 de | 11.14 ± 1.14 e | 4.03 ± 0.35 e | 2.22 ± 0.24 bc |
| CEC | 3.06 ± 0.26 cd | 17.46 ± 0.77 f | 1.08 ± 0.10 gh | 2.17 ± 0.78 bc | 1.47 ± 0.42 bc | 1.68 ± 0.30 cde | 2.73 ± 0.20 cd | 9.70 ± 0.34 g | 10.04 ± 1.03 f | 1.54 ± 0.33 de | 12.73 ± 1.23 d | 7.10 ± 0.24 b | 2.62 ± 1.27 b |
| PE | 2.30 ± 0.25 e | 13.94 ± 0.47 g | 2.12 ± 0.09 cd | 1.44 ± 0.10 cde | 1.29 ± 0.50 cd | 1.36 ± 0.31 cde | 3.54 ± 0.14 a | 11.96 ± 0.87 f | 14.63 ± 0.90 d | 1.88 ± 0.79 cd | 17.62 ± 0.49 b | 4.71 ± 0.17 d | 2.29 ± 0.09 bc |
| Chl | 2.65 ± 0.28 de | 17.28 ± 1.39 f | 2.76 ± 0.47 b | 1.46 ± 0.12 cde | 0.70 ± 0.21 de | 1.61 ± 0.21 cde | 1.94 ± 0.21 ef | 7.42 ± 0.18 h | 10.62 ± 0.69 ef | 6.57 ± 0.13 a | 9.63 ± 1.15 f | 2.33 ± 0.12 g | 1.62 ± 0.28 c |
| EA | 0.67 ± 0.02 g | 11.32 ± 0.51 i | 1.53 ± 0.13 efg | 1.93 ± 0.61 bcd | 1.33 ± 0.35 cd | 1.86 ± 0.18 bcd | 3.34 ± 0.18 ab | 8.73 ± 0.30 g | 19.20 ± 0.50 c | 2.41 ± 0.30 c | 15.57 ± 0.18 c | 3.65 ± 0.50 ef | 2.37 ± 0.05 bc |
| NB | 2.50 ± 0.29 e | 30.64 ± 1.03 a | 4.71 ± 0.31 a | 2.38 ± 0.51 b | 2.26 ± 0.18 a | 2.65 ± 0.22 a | 2.96 ± 0.59 bc | 4.06 ± 0.14 j | 7.24 ± 1.39 hi | 1.26 ± 0.25 de | 8.59 ± 0.53 f | 3.22 ± 0.14 f | 1.91 ± 0.42 bc |
| NY | 0.25 ± 0.04 g | 27.79 ± 0.52 b | 1.54 ± 0.07 efg | 0.48 ± 0.06 f | 0.59 ± 0.04 e | 1.82 ± 0.07 bcd | 2.40 ± 0.06 de | 26.44 ± 1.18 b | 14.18 ± 0.37 d | 1.69 ± 0.35 cde | 13.12 ± 1.12 d | 3.75 ± 0.13 e | 0.56 ± 0.16 d |
| QY | 0.65 ± 0.04 g | 24.08 ± 0.46 d | 1.53 ± 0.07 efg | 0.90 ± 0.06 ef | 0.62 ± 0.02 e | 2.10 ± 0.02 abc | 2.27 ± 0.04 de | 23.76 ± 0.36 c | 20.58 ± 0.50 b | 0.89 ± 0.20 e | 12.71 ± 0.22 d | 1.34 ± 0.04 h | 4.55 ± 0.46 a |
| WT | 0.63 ± 0.21 g | 11.54 ± 0.42 hi | 1.29 ± 0.22 fg | 0.86 ± 0.06 ef | 0.54 ± 0.02 e | 1.41 ± 0.19 cde | 3.70 ± 0.25 a | 17.48 ± 0.96 e | 23.41 ± 0.72 a | 1.55 ± 0.25 de | 19.30 ± 0.71 a | 5.72 ± 0.44 c | 2.15 ± 0.04 bc |
| Aug. | 4.5 ± 0.64 b | 7.15 ± 0.80 j | 0.57 ± 0.29 i | 3.16 ± 0.52 a | 2.07 ± 0.11 ab | 1.94 ± 0.87 abc | 1.72 ± 0.08 f | 5.39 ± 0.23 i | 2.45 ± 0.44 j | 4.88 ± 1.10 b | 5.00 ± 0.47 g | 7.62 ± 0.40 a | 4.35 ± 0.50 a |
| Sep. | 7.33 ± 0.29 a | 12.67 ± 1.03 h | 0.80 ± 0.17 hi | 3.88 ± 0.97 a | 1.25 ± 0.87 cd | 1.01 ± 0.86 e | 2.01 ± 0.48 ef | 18.52 ± 1.05 d | 11.85 ± 1.20 e | 1.86 ± 0.23 cd | 9.67 ± 1.43 f | 6.76 ± 0.16 b | 3.85 ± 0.71 a |
| Oct. | 1.70 ± 0.22 f | 19.55 ± 0.50 e | 1.70 ± 0.52 def | 1.34 ± 0.17 de | 2.23 ± 0.16 a | 1.51 ± 0.30 cde | 3.31 ± 0.17 ab | 33.56 ± 0.30 a | 6.80 ± 0.08 i | 0.84 ± 0.01 e | 11.74 ± 0.58 de | 3.81 ± 0.14 e | 2.37 ± 0.15 bc |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.-J.; Long, X.-H.; Li, E.-Z. Evaluation of Antifungal Phenolics from Helianthus tuberosus L. Leaves against Phytophthora capsici Leonian by Chemometric Analysis. Molecules 2019, 24, 4300. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24234300
Chen F-J, Long X-H, Li E-Z. Evaluation of Antifungal Phenolics from Helianthus tuberosus L. Leaves against Phytophthora capsici Leonian by Chemometric Analysis. Molecules. 2019; 24(23):4300. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24234300
Chicago/Turabian StyleChen, Fu-Jia, Xiao-Hua Long, and En-Zhong Li. 2019. "Evaluation of Antifungal Phenolics from Helianthus tuberosus L. Leaves against Phytophthora capsici Leonian by Chemometric Analysis" Molecules 24, no. 23: 4300. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24234300