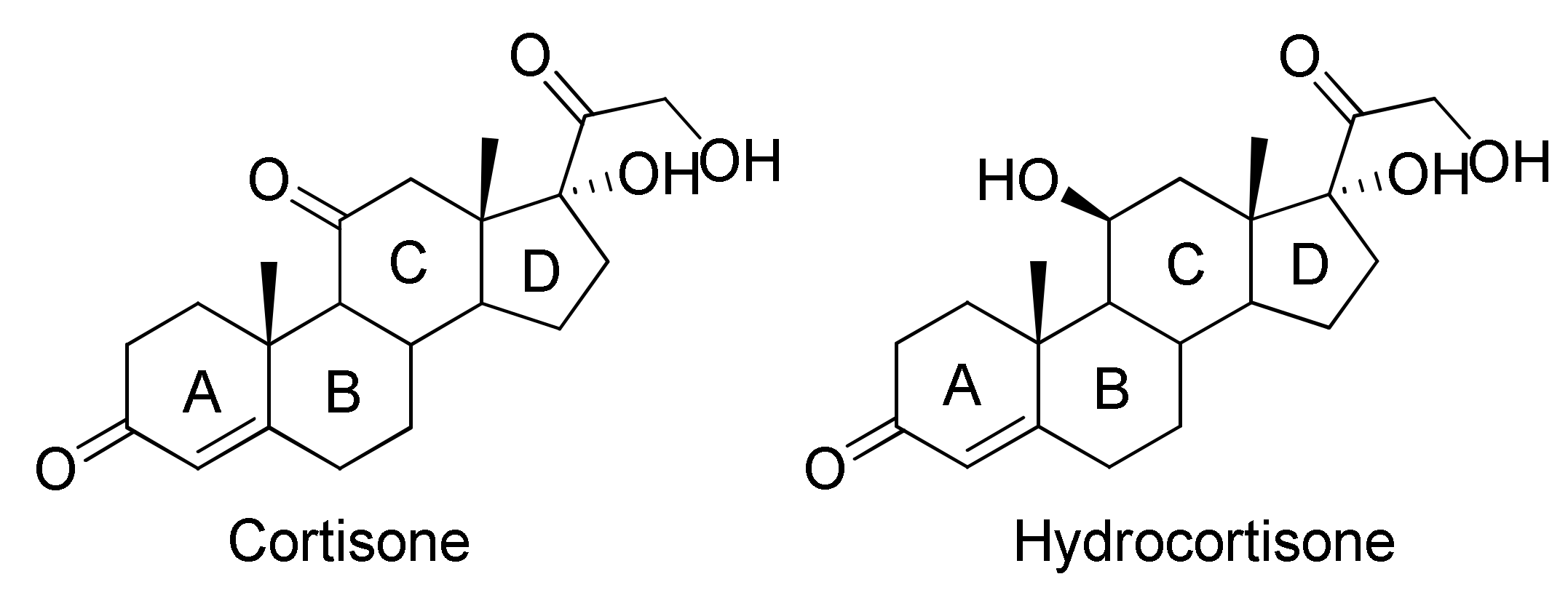
Δ1-Dehydrogenation and C20 Reduction of Cortisone and Hydrocortisone Catalyzed by Rhodococcus Strains

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Rhodococcus Screening and Selection
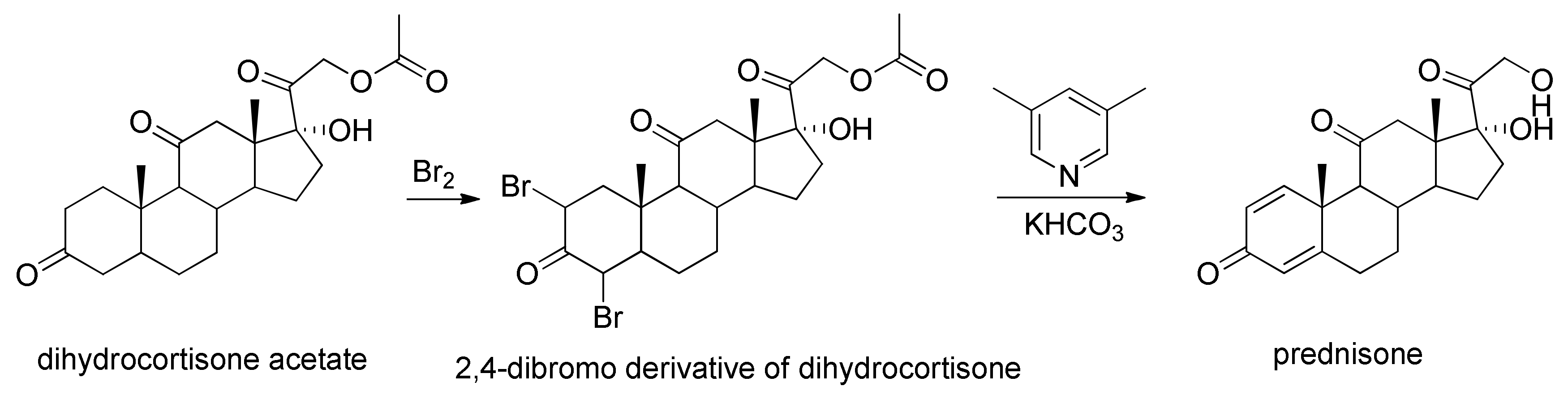
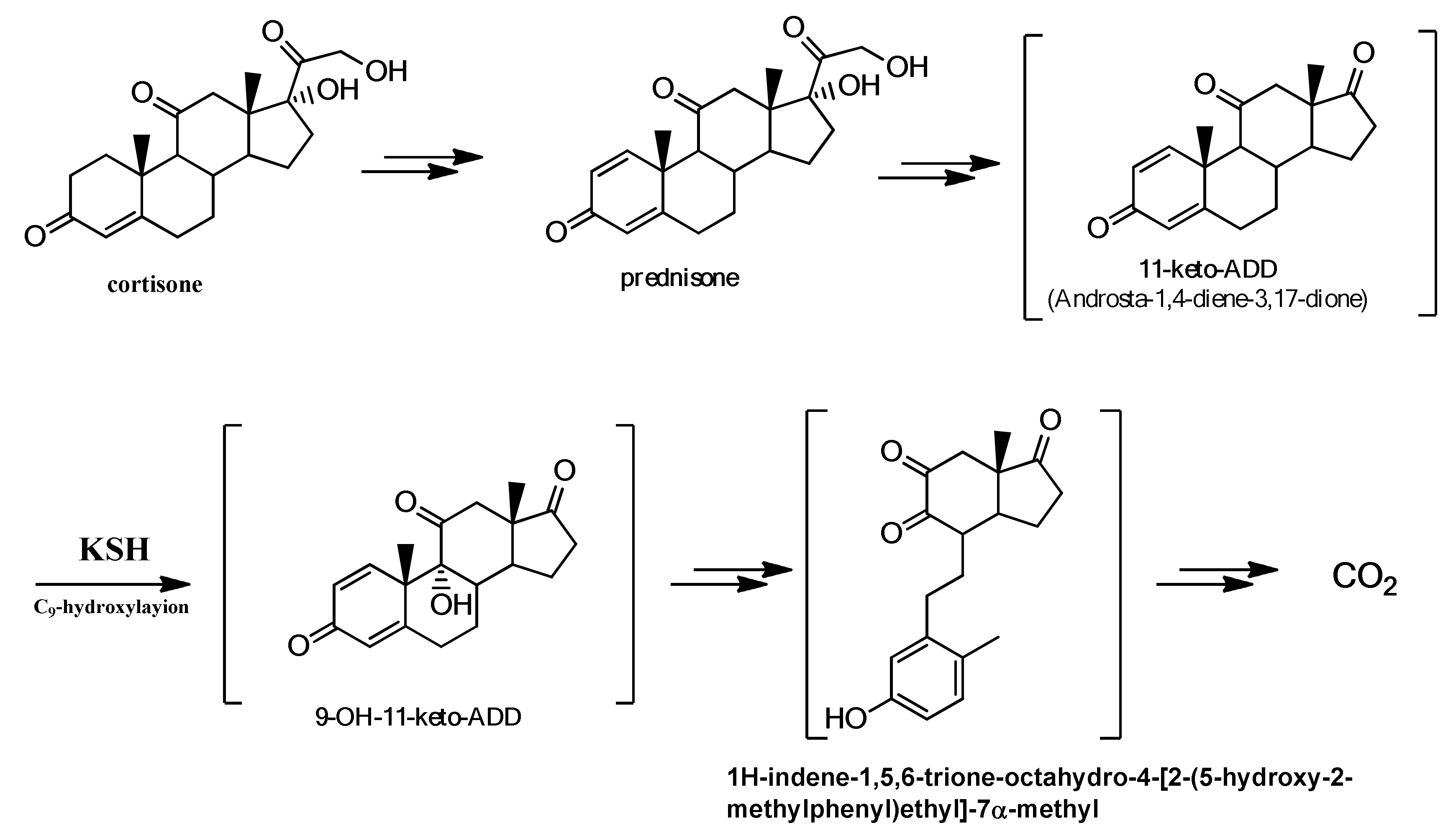
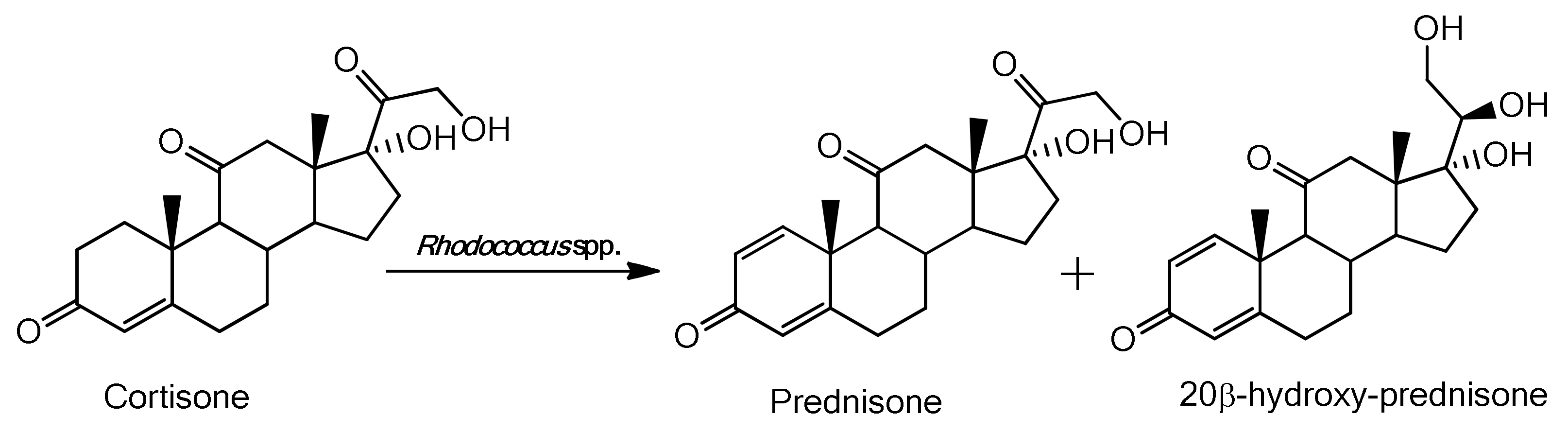
2.2. Characterization of the Biotransformation Products of Cortisone
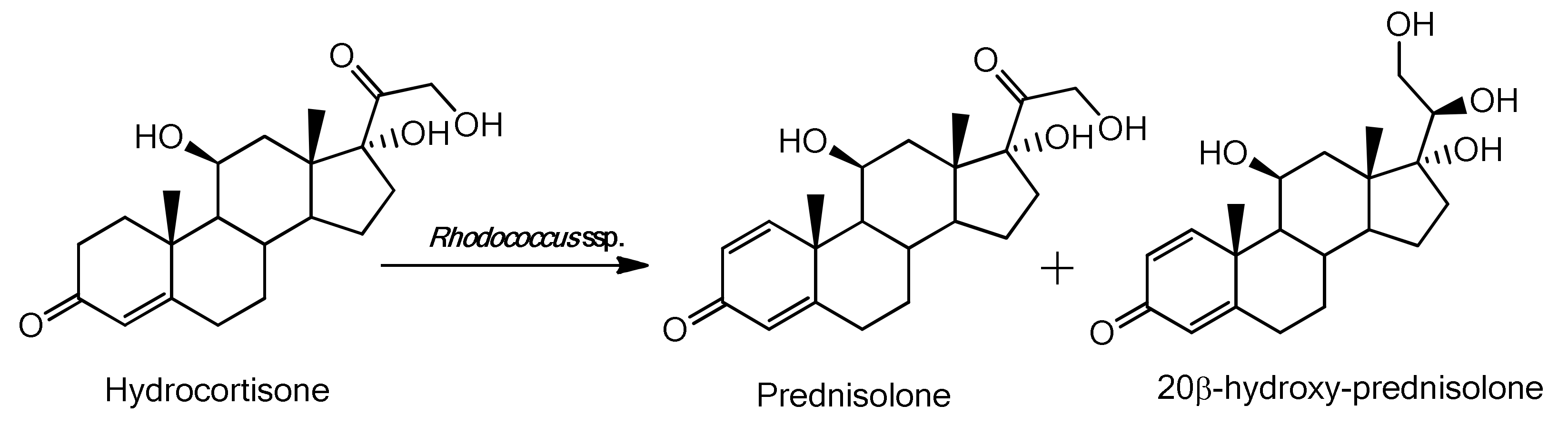
2.3. Characterization of the Biotransformation Products of Hydrocortisone
3. Materials and Methods
3.1. Chemicals and Rhodococcus Strains
3.2. Rhodococcus Biotransformation Screening Conditions
3.3. Semi-Preparative Biotransformation Protocol
3.4. Analitical Methods
3.5. Compound Data
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fernandes, P.; Cabral, J.M.S. Phytosterols: Applications and recovery methods. Bioresour. Technol. 2007, 98, 2335–2350. [Google Scholar] [CrossRef] [PubMed]
- Hannich, J.T.; Umebayashi, K.; Riezman, H. Additional Perspectives on The Biology of Lipids. Cold Spring Harbour Perspect. Biol. 2011, 3, 4762–4763. [Google Scholar] [CrossRef] [PubMed]
- Muthukrishnan, S.; Merzendorfer, H.; Arakane, Y.; Kramer, K.J. Chitin Metabolism in Insects. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 193–235. [Google Scholar]
- Tong, W.Y.; Dong, X. Microbial Biotransformation: Recent Developments on Steroid Drugs. Recent Pat. Biotechnol. 2009, 3, 141–153. [Google Scholar] [CrossRef]
- Fragkaki, A.G.; Angelis, Y.S.; Koupparis, M.; Tsantili-Kakoulidou, A.; Kokotos, G.; Georgakopoulos, C. Structural characteristics of anabolic androgenic steroids contributing to binding to the androgen receptor and to their anabolic and androgenic activities. Applied modifications in the steroidal structure. Steroids 2009, 74, 172–197. [Google Scholar] [CrossRef] [PubMed]
- Agoston, E.S.; Hatcher, M.A.; Kensler, T.W.; Posner, G.H. Vitamin D Analogs as Anti-Carcinogenic Agents. Anti. Cancer Agents Med. Chem. 2006, 3, 53–71. [Google Scholar] [CrossRef]
- Tuba, Z.; Bardin, C.W.; Dancsi, A.; Francsics–Czinege, E.; Molnár, C.; Csörgei, J.; Falkay, G.; Samuel, S. Synthesis and biological activity of a new progestogen, 16-methylene- 17α-hydroxy-18-methyl-19-norpregn-4-ene-3,20-dione acetate. Steroids 2000, 65, 266–274. [Google Scholar] [CrossRef]
- Díaz-Chico, N.; Germán Rodríguez, F.; González, A.; Ramírez, R.; Bilbao, C.; Cabrera de León, A.; Aguirre Jaime, A.; Chirino, R.; Navarro, D.; Díaz-Chico, J.C. Androgens and androgen receptors in breast cancer. J.Steroid Biochem. 2007, 105, 1–15. [Google Scholar] [CrossRef]
- Chrousos, G.P. Basic & Clinical Pharmacology. In Adrenocorticosteroids & Adrenocortical Antagonists, 13th ed.; McGraw-Hill Medical: New York, NY, USA, 2015. [Google Scholar]
- Craigie, E.; Mullins, J.J.; Bailey, M.A. Glucocorticoids and mineralocorticoids. In Cardiovascular Hormone Systems: From Molecular Mechanisms to Novel Therapeutics; Bader, M., Ed.; Wiley-VCH Verlag GmbH & Co: Weinheim, Germany, 2009; pp. 1–64. [Google Scholar]
- Chung, S.K.; Ryooa, C.H.; Yang, H.W.; Shim, J.-Y.; Kang, M.G.; Lee, K.W.; Kang, H.I. Synthesis and bioactivities of steroid derivatives as antifungal agents. Tetrahedron 1998, 54, 1589–1591. [Google Scholar] [CrossRef]
- Suzuki, K.; Nakata, T.; Shimizu, T. Anti-Obesity Agents Patent 5,846,962, 8 December 1998.
- Dombrowski, A.W.; Hazuda, D.J.; Polishook, J.D.; Felock, P.J.; Singh, S.B.; Zink, D.L. HIV-Integrase-Inhibitors. World Patent 0036132, 22 June 2000. [Google Scholar]
- Arthan, D.; Svasti, J.; Kittakoop, P.; Pittayakhachonwut, D.; Tanticharoen, M.; Thebtaranonth, Y. Antiviral isoflavonoid sulfate and steroidal glycosides from the fruits of Solanum torvum. Phytochemistry 2002, 59, 459–463. [Google Scholar] [CrossRef]
- Funder, J.W. Minireview: Aldosterone and mineralocorticoid receptors: Past, present, and future. Endocrinology 2010, 151, 5098–5102. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tian, Y.; Wang, J.; Li, Y.; Wang, H.; Mao, S.; Liu, X.; Wang, C.; Bie, S.; Lu, F. Construction of engineered Arthrobacter simplex with improved performance for cortisone acetate biotransformation. Appl. Microbiol. Biotechnol. 2013, 97, 9503–9514. [Google Scholar] [CrossRef] [PubMed]
- Samuel, S.; Nguyen, T.; Choi, H.A. Pharmacologic Characteristics of Corticosteroids. J. Neurocritical Care 2017, 10, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Olivetq, E.P.; Gould, D.H. US Patent 2.897.216, 28 July 1959.
- Fernández-Cabezón, L.; Galán, B.; García, J. New Insights on Steroid Biotechnology. Front Microbiol. 2018, 15, 958–973. [Google Scholar] [CrossRef]
- Mao, S.; Yu, L.; Ji, S.; Liu, X.; Lu, F. Evaluation of deep eutectic solvents as co-solvent for steroids 1-en-dehydrogenation biotransformation by Arthrobacter simplex. J. Chem. Technol. Biotechnol. 2016, 91, 1099–1104. [Google Scholar] [CrossRef]
- Gao, Q.; Shen, Y.B.; Huang, W.; Wang, M. Effect of Natural Cyclodextrins on Cell Growth, Activity and Permeability of Arthrobacter simplex. Appl. Mech. Mater. 2015, 730, 249–253. [Google Scholar] [CrossRef]
- Luo, J.; Ning, J.; Wang, Y.; Cheng, Y.; Zheng, Y.; Shen, Y.; Wang, M. The effect of ethanol on cell properties and steroid 1-en-dehydrogenation biotransformation of Arthrobacter simplex. Biotechnol. Appl. Biochem. 2014, 61, 555–564. [Google Scholar] [CrossRef]
- Spassov, G.; Krützfeldt, R.; Sheldrick, W.S.; Wania, W.; Vlahov, R.; Snatzke, G. Crystallographic monitoring of microbiological steroid transformations. Eur. J. Appl. Microbiol. Biotechnol. 1983, 17, 80–84. [Google Scholar] [CrossRef]
- Vlahov, R.; Pramatarova, V.; Spassov, G.; Suchodolskaya, G.V.; Koshcheenko, K.A. Transformation of microcrystalline hydrocortisone by free and immobilized cells of Arthrobacter simplex. Appl. Microbiol. Biotechnol. 1990, 33, 172–175. [Google Scholar] [CrossRef]
- Wang, X.; Feng, J.; Zhang, D.; Wu, Q.; Zhu, D.; Ma, Y. Characterization of new recombinant 3-ketosteroid-Δ1-dehydrogenases for the biotransformation of steroids. Appl. Microbiol. Biotechnol. 2017, 101, 6049–6060. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Rasoul-Amini, S.; Morowvat, M.H.; Raee, M.J.; Ghoshoon, M.B.; Nouri, F.; Negintaji, N.; Parvizi, R.; Mosavi-Azam, S.B. Characterization of Hydrocortisone Biometabolites and 18S rRNA Gene in Chlamydomonas reinhardtii Cultures. Molecules 2008, 13, 2416–2425. [Google Scholar] [CrossRef] [Green Version]
- Bie, S.; Lu, F.; Liu, X.; Mao, S.; Li, J. Method for Manufacturing Prednisolone with Biotransformation. Patent CN 2012 102505024 A 20120620, 20 June 2012. [Google Scholar]
- Choudhary, M.I.; Siddiqui, Z.A.; Musharraf, S.G.; Nawaz, S.A. Microbial transformation of prednisone. Nat. Prod. Res. 2005, 19, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cui, L.; Wu, M. Transformation of prednisolone to a 20β-hydroxy-prednisolone compound by Streptomyces roseochromogenes TS79. Appl. Microbiol. Biotechnol. 2011, 92, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.S.; El-Hadi, A.A. Biotransformation of prednisolone to hydroxyl derivatives by Penicillium aurantium. Biocatal. Biotransformation 2017, 35, 215–222. [Google Scholar] [CrossRef]
- Donova, M.V. Transformation of steroids by actinobacteria: A review. Appl. Biochem. Micro. 2007, 43, 1–14. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Kuyukina, M.; Ivshina, I. Advanced Rhodococcus Biocatalysts for Environmental Biotechnologies. Catalysts 2019, 9, 236–255. [Google Scholar] [CrossRef] [Green Version]
- Lichtinger, T.; Reiss, G.; Benz, R. Biochemical identification and biophysical characterization of a channel-forming protein from Rhodococcus erythropolis. J. Bacteriol. 2000, 182, 764–770. [Google Scholar] [CrossRef] [Green Version]
- Patrauchan, M.A.; Florizone, C.; Dosanjh, M.; Mohn, W.W.; Davies, J.; Eltis, L.D. Catabolism of benzoate and phthalate in Rhodococcus sp. strain RHA1: Redundancies and convergence. J. Bacteriol. 2005, 187, 4050–4063. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.; Giovannini, P.P.; Fantin, G.; Medici, A.; Pedrini, P. New 9,10-secosteroids from biotransformations of bile acids with Rhodococcus ruber. Helv. Chim. Acta 2013, 96, 2124–2133. [Google Scholar] [CrossRef]
- Costa, S.; Giovannini, P.P.; Fantin, G.; Medici, A.; Pedrini, P. New 9,10-secosteroids from biotransformations of hyodeoxycholic acid with Rhodococcus spp. Helv. Chim. Acta 2013, 96, 1062–1071. [Google Scholar] [CrossRef]
- Horinouchi, M.; Hayashi, T.; Kudo, T. Steroid degradation in Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2012, 129, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Van Der Geize, R.; Besra, G.S.; Gurcha, S.S.; Liu, A.; Rohde, M.; Singh, M.; Coates, A. 3-Ketosteroid 9α-hydroxylase is an essential factor in the pathogenesis of Mycobacterium tuberculosis. Mol. Microbiol. 2010, 75, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Capyk, J.K.; D’Angelo, I.; Strynadka, N.C.; Eltis, L.D. Characterization of 3-ketosteroid 9α-hydroxylase, a Rieske oxygenase in the cholesterol degradation pathway of Mycobacterium tuberculosis. J. Biol. Chem. 2009, 284, 9937–9946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrusma, M.; Dijkhuizen, L.; Van Der Geize, R. Rhodococcus rhodochrous DSM 43269 3-ketosteroid 9α-hydroxylase, a two-component iron-sulfur-containing monooxygenase with subtle steroid substrate specificity. Appl. Environ. Microbiol. 2009, 75, 5300–5307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, R.J. Phenolic 9,10-secosteroids as products of the catabolism of bile acids by a Pseudomonas sp. Steroids 1984, 44, 175–193. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Products from Cortisone | Products from Hydrocortisone |
|---|---|---|
| R. zopfii | Degradation | Rf 0.45 |
| R. opacus | no product | no product |
| R. fascians | no product | no product |
| R. baikonurensis | Rf 0.23, 0.55 | Rf 0.21 |
| R. globerulus | Rf 0.55 | Rf 0.45 |
| R. aetherivorans | Rf 0.55 | no product |
| R. coprophilus | Rf 0.55 | Rf 0.45 |
| R. rhodochrous NCIMB 11216 | no product | no product |
| Rhodococcus sp. R312 | no product | no product |
| R. equi | no product | no product |
| R. ruber | Rf 0.55 | Rf 0.45 |
| R. rhodochrous DSM 43273 | Rf 0.55 | degradation |
| R. erythropholis | Rf 0.23, 0.55 | Rf 0.45, 0.21 |
| Bacteria | Time of Biotransformation (h) | Prednisone (% Yield) | 20β-Hydroxy-Prednisone (% Yield) | Cortisone Residue (%) |
|---|---|---|---|---|
| R. baikonurensis | 72 | 27 ± 3 (0.053 ± 0.005 g ) | 68 ± 4 (0.134 ± 0.007 g) | -- |
| R. globerulus | 72 | 33 ± 6 (0.065 ± 0.012 g) | -- | 65 ± 3 (0.129 ± 0.006 g) |
| R. aetherivorans | 72 | 35 ± 3 (0.069 ± 0.006 g) | -- | 63 ± 7 (0.125 ± 0.014 g) |
| R. coprophilus | 24 | 94 ± 2 (0.185 ± 0.004 g) | -- | -- |
| R. ruber | 72 | 52 ± 4 (0.102 ± 0.008 g) | -- | 47 ± 4 (0.093 ± 0.008 g) |
| R. rhodochrous DSM 43273 | 72 | 31 ± 3 (0.0065 ± 0.006) | 53 ± 3 (0.105 ± 0.006) | |
| R. erythropolis | 72 | 33 ± 5 (0.065 ± 0.01 g) | 37 ± 6 (0.073 ± 0.011 g) | 28 ± 6 (0.056 ± 0.012 g) |
| Bacteria | Time of Biotransformation (h) | Prednisolone (% Yield) | 20β-Hydroxy-Prednisolone (% Yield) | Hydrocortisone Residue (%) |
|---|---|---|---|---|
| R. baikonurensis | 72 | -- | 32 ± 3 (0.063 ± 0.006 g) | -- |
| R. globerulus | 72 | 76 ± 2 (0.151 ± 0.004 g) | -- | 23 ± 1 (0.045 ± 0.002 g) |
| R. coprophilus | 24 | 97 ± 2 (0.192 ± 0.004 g) | -- | -- |
| R. ruber | 72 | 56 ± 4 (0.111 ± 0.009 g) | -- | 40 ± 4 (0.072 ± 0.007 g) |
| R. erythropolis | 72 | 37 ± 5 (0.073 ± 0.01 g) | 40 ± 8 (0.079 ± 0.016 g) | 18 ± 6 (0.035 ± 0.012 g) |
| # | Genus | Species | Identification Code |
|---|---|---|---|
| 1 | Rhodococcus | zopfii | DSM 44108 |
| 2 | Rhodococcus | opacus | DSM 43250 |
| 3 | Rhodococcus | fascians | DSM 43985 |
| 4 | Rhodococcus | baikonurensis | DSM 44587 |
| 5 | Rhodococcus | globerulus | DSM 43954 |
| 6 | Rhodococcus | aetherivorans | DSM 44752 |
| 7 | Rhodococcus | coprophilus | DSM 43347 |
| 8 | Rhodococcus | rhodochrous | NCIMB 11216 |
| 9 | Rhodococcus | sp. R312 | CBS 717.73 |
| 10 | Rhodococcus | equi | IFO 3730 |
| 11 | Rhodococcus | ruber | DSM 6264 |
| 12 | Rhodococcus | rhodochrous | DSM 43273 |
| 13 | Rhodococcus | erythropholis | DSM 43188 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, S.; Zappaterra, F.; Summa, D.; Semeraro, B.; Fantin, G. Δ1-Dehydrogenation and C20 Reduction of Cortisone and Hydrocortisone Catalyzed by Rhodococcus Strains. Molecules 2020, 25, 2192. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25092192
Costa S, Zappaterra F, Summa D, Semeraro B, Fantin G. Δ1-Dehydrogenation and C20 Reduction of Cortisone and Hydrocortisone Catalyzed by Rhodococcus Strains. Molecules. 2020; 25(9):2192. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25092192
Chicago/Turabian StyleCosta, Stefania, Federico Zappaterra, Daniela Summa, Bruno Semeraro, and Giancarlo Fantin. 2020. "Δ1-Dehydrogenation and C20 Reduction of Cortisone and Hydrocortisone Catalyzed by Rhodococcus Strains" Molecules 25, no. 9: 2192. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25092192