Dietary RNAs: New Stories Regarding Oral Delivery

Abstract
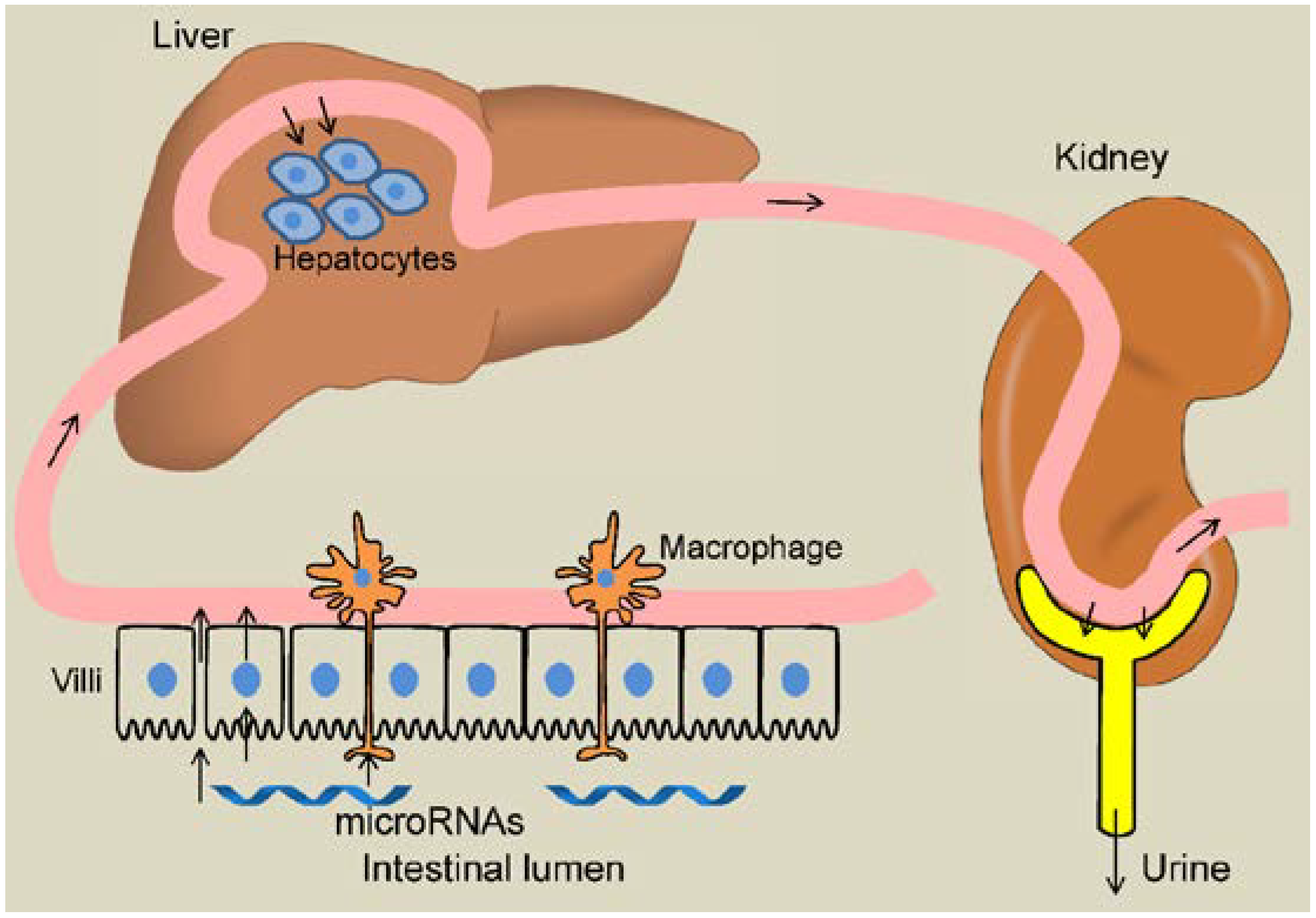
:1. Introduction
2. Overview of Main Techniques Used to Measure microRNA Levels
3. Initial Publication on Uptake and Function of Dietary microRNAs in Mammals
4. Contradicting Evidence for Dietary Uptake of microRNA
4.1. Unsuccessful Replication of the Seminal Study Using Rice Diet in Mice
4.2. Human Athletes, Honeybees Did Not Show Obvious Absorption of miRNAs from Plant Diets
4.3. Neither Normal Mice nor miR-21 Null Mice Showed Any Absorption of Dietary miRNAs
4.4. Nonhuman Primates Failed to Absorb Plant miRNAs
5. Evidence Supporting Dietary Uptake of microRNAs
5.1. An Independent NGS Study Suggests General Presence of Foregin sRNAs from a Wide Range of Organisms in Human Bloodstream
5.2. Plant miRNAs Are Found in Mammalian Milk Exosomes
5.3. Cabbage MIR172 is Absorbed in Mice
5.4. Does Milk Do a miRNA Good?
5.5. Dietary and Pharmacological Influences on Dietary RNA Uptake
5.6. Anticancer Potential for Plant-Based miRNAs
6. Implication of Dietary microRNAs Regulating Consumer Genes

{kind=link}
| Species | Highly Expressed microRNAs |
|---|---|
| Rice (Oryza Sativa) | osa-miR156a, osa-miR168a, osa-miR1846e, osa-miR167d, osa-miR168b [30] |
| Corn (Zea May) | zma-miR156a, zma-miR168a, zma-miR 169c, zma-miR399e, zma-miR167a, zma-miR156 [31] |
| Soybean (Glycine max) | gma-MIR3522b, gma-MIR-1507a, gma-MIR 1509a, gma-MIR482*, gma-MIR-1510a-5p, gma-MIR167d, gma-MIR 166a, gma-MIR-166b; gma-MIR396e [32] |
| Tomato (Solanum lycopersicum) | miR159 a, miR162b, miR168a, miR164c, miR164d [33] |
| Barley (Hordeum vulgare) | hvu-miR168, hvu-miR156, hvu-miR167, hvu-miR165/166, hvu-miR172, hvu-miR2005 [34] |
| Wild wheat (Triticum dicoccoides) | miR159, miR1450, miR319, miR896, miR168, miR474, miR167, miR164, miR528, miR1436, miR396 and miR894 [35] |
| Peanut (Arachis hypogaea) | ahy-miR157a, ahy-miR156a, ahy-miR168a, ahy-miR166a, ahy-miR166h, ahy-miR167f [36] |
| Sweet Orange (Citrus sinensis) | csi-miR168a, csi-miR172a, csi-miR166j, csi-miR167a, csi-miR157a, csi-miR479, csi-miR156a [37] |
| Oilseed (Brassica napus) | miR156, miR157, miR168a, miR166, miR167, miR399 [38] |
7. Future Directions

8. Conclusions
| Dietary microRNA Uptake and Function in Mammalian Consumers? | |
|---|---|
| Evidences Against |
|
| Evidences For |
|
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: Artefacts no more. Trends Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.T.; Schmittgen, T.D.; et al. Detection of microRNA expression in human peripheral blood microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, H.; Miller, N.; Kerin, M. Circulating microRNAs: Promising breast cancer Biomarkers. Breast Cancer Res. 2011, 13, 402. [Google Scholar] [CrossRef] [PubMed]
- Winston, W.M.; Molodowitch, C.; Hunter, C.P. Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science 2002, 295, 2456–2459. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, A.; Fraser, A. Uncover Genetic Interactions in Caenorhabditis elegans by RNA Interference. Biosci. Rep. 2005, 25, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W. XenomiRs and miRNA homeostasis in health and disease: Evidence that diet and dietary miRNAs directly and indirectly influence circulating miRNA profiles. RNA Biol. 2012, 9, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.; Zhang, Y.; Petrick, J.S.; Heck, G.; Ivashuta, S.; Marshall, W.S. Lack of detectable oral bioavailability of plant microRNAs after feeding in mice. Nat. Biotechnol. 2013, 31, 965–967. [Google Scholar] [CrossRef] [PubMed]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs Are Absorbed in Biologically Meaningful Amounts from Nutritionally Relevant Doses of Cow Milk and Affect Gene Expression in Peripheral Blood Mononuclear Cells, HEK-293 Kidney Cell Cultures, and Mouse Livers. J. Nutr. 2014, 144, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res. 2014, 25, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Mlotshwa, S.; Pruss, G.J.; MacArthur, J.L.; Endres, M.W.; Davis, C.; Hofseth, L.J.; Peña, M.M.; Vance, V. A novel chemopreventive strategy based on therapeutic microRNAs produced in plants. Cell Res. 2015, 25, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Farmer, L.M.; Agyekum, A.A.A.; Hirschi, K.D. Detection of dietary plant-based small RNAs in animals. Cell Res. 2015, 25, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Yang, Z.; Li, J.; Minakhina, S.; Yang, M.; Padgett, R.W.; Steward, R.; Chen, X. Methylation as a crucial step in plant microRNA biogenesis. Science 2005, 307, 932–935. [Google Scholar] [CrossRef] [PubMed]
- Snow, J.W.; Hale, A.E.; Isaacs, S.K.; Baggish, A.L.; Chan, S.Y. Ineffective delivery of diet-derived microRNAs to recipient animal organisms. RNA Biol. 2013, 10, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; McAlexander, M.A.; Queen, S.E.; Adams, R.J. Real-time quantitative PCR and droplet digital PCR for plant miRNAs in mammalian blood provide little evidence for general uptake of dietary miRNAs: Limited evidence for general uptake of dietary plant xenomiRs. RNA Biol. 2013, 10, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Stokowy, T.; Eszlinger, M.; Świerniak, M.; Fujarewicz, K.; Jarząb, B.; Paschke, R.; Krohn, K. Analysis options for high-throughput sequencing in miRNA expression profiling. BMC Res. Notes 2014, 7, 144. [Google Scholar] [CrossRef] [PubMed]
- Bizouarn, F. Introduction to digital PCR. Methods Mol. Biol. 2014, 1160, 27–41. [Google Scholar] [PubMed]
- Naccache, S.N.; Greninger, A.L.; Lee, D.; Coffey, L.L.; Phan, T.; Rein-Weston, A.; Aronsohn, A.; Hackett, John, Jr.; Delwart, E.L.; Chiu, C.Y. The perils of pathogen discovery: Origin of a novel parvovirus-like hybrid genome traced to nucleic acid extraction spin columns. J. Virol. 2013, 87, 11966–11977. [Google Scholar] [CrossRef] [PubMed]
- Charlson, E.S.; Bittinger, K.; Haas, A.R.; Fitzgerald, A.S.; Frank, I.; Yadav, A.; Bushman, F.D.; Collman, R.G. Topographical continuity of bacterial populations in the healthy human respiratory tract. Am. J. Respir. Crit. Care Med. 2011, 184, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Hirschi, K.D. Transfer and functional consequences of dietary microRNAs in vertebrates: Concepts in search of corroboration: Negative results challenge the hypothesis that dietary xenomiRs cross the gut and regulate genes in ingesting vertebrates, but important questions persist. Bioessays 2014, 36, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wiggins, B.E.; Lawrence, C.; Petrick, J.; Ivashuta, S.; Heck, G. Analysis of plant-derived miRNAs in animal small RNA datasets. BMC Genomics 2012, 13, 381. [Google Scholar] [CrossRef] [PubMed]
- Petrick, J.S.; Brower-Toland, B.; Jackson, A.L.; Kier, L.D. Safety assessment of food and feed from biotechnology-derived crops employing RNA-mediated gene regulation to achieve desired traits: A scientific review. Regul. Toxicol. Pharmacol. 2013, 66, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, H.; Yuan, Y.; Etheridge, A.; Zhou, Y.; Huang, D.; Wilmes, P.; Galas, D. The complex exogenous RNA spectra in human plasma: An interface with human gut biota? PLoS ONE 2012, 7, e51009. [Google Scholar] [CrossRef] [PubMed]
- Lukasik, A.; Zielenkiewicz, P. In Silico Identification of Plant miRNAs in Mammalian Breast Milk Exosomes—A Small Step Forward? PLoS ONE 2014, 9, e99963. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Zhu, Y.; Sun, B.; Shao, Y.; Jing, A.; Wang, J.; Xiao, Z. Assessing the survival of exogenous plant microRNA in mice. Food Sci. Nutr. 2014, 2, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Munch, E.M.; Harris, R.A.; Mohammad, M.; Benham, A.L.; Pejerrey, S.M.; Showalter, L.; Hu, M.; Shope, C.D.; Maningat, P.D.; Gunaratne, P.H.; et al. Transcriptome profiling of microRNA by Next-Gen deep sequencing reveals known and novel miRNA species in the lipid fraction of human breast milk. PLoS ONE 2013, 8, e50564. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W. Diet-responsive mammalian miRNAs are likely endogenous. J. Nutr. 2014, 144, 1880–1881. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.J.; Zhang, J.J.; Xue, H.W. Characterization and expression profiles of miRNAs in rice seeds. Nucleic Acids Res. 2009, 37, 916–930. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Elling, A.A.; Li, X.; Li, N.; Peng, Z.; He, G.; Sun, H.; Qi, Y.; Liu, X.S.; Deng, X.W. Genome-wide and organ-specific landscapes of epigenetic modifications and their relationships to mRNA and small RNA transcriptomes in maize. Plant Cell 2009, 21, 1053–1069. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.X.; Liu, Y.F.; Hu, X.Y.; Zhang, W.K.; Ma, B.; Chen, S.Y.; Zhang, J.S. Identification of miRNAs and their target genes in developing soybean seeds by deep sequencing. BMC Plant Biol. 2011, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zeng, R.; Chen, J.; Liu, X.; Liao, Q. Identification of conserved microRNAs and their targets from Solanum lycopersicum Mill. Gene 2008, 423, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, A.W.; Shi, B.J.; Huang, C.Y.; Langridge, P.; Baumann, U. Discovery of barley miRNAs through deep sequencing of short reads. BMC Genomics 2011, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Kantar, M.; Lucas, S.J.; Budak, H. miRNA expression patterns of Triticum dicoccoides in response to shock drought stress. Planta 2011, 233, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Z.; Xia, H.; Frazier, T.P.; Yao, Y.Y.; Bi, Y.P.; Li, A.Q.; Li, M.J.; Li, C.S.; Zhang, B.H.; Wang, X.J. Deep sequencing identifies novel and conserved microRNAs in peanuts (Arachis hypogaea L.). BMC Plant Biol. 2010, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Liu, Y.; Zhu, A.; Wu, X.; Ye, J.; Yu, K.; Guo, W.; Deng, X. Discovery and comparative profiling of microRNAs in a sweet orange red-flesh mutant and its wild type. BMC Genomics 2010, 11, 246. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.T.; Wang, M.; Fu, S.X.; Yang, W.C.; Qi, C.K.; Wang, X.J. Small RNA profiling in two Brassica napus cultivars identifies microRNAs with oil production- and development-correlated expression and new small RNA classes. Plant Physiol. 2012, 158, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Luo, H. MicroRNA-mediated gene regulation: Potential applications for plant genetic engineering. Plant Mol. Biol. 2013, 83, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Castellano, L.; Stebbing, J. Deep sequencing of small RNAs identifies canonical and non-canonical miRNA and endogenous siRNAs in mammalian somatic tissues. Nucleic Acids Res. 2013, 41, 3339–3351. [Google Scholar] [CrossRef] [PubMed]
- Helwak, A.; Kudla, G.; Dudnakova, T.; Tollervey, D. Mapping the human miRNA interactome by CLASH reveals frequent noncanonical binding. Cell 2013, 153, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Grobe, A.; Rissanen, M.E. Nanotechnologies in agriculture and food—An overview of different fields of application, risk assessment and public perception. Recent Pat. Food Nutr. Agric. 2012, 4, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Barratt, B.I.; Todd, J.H.; Burgess, E.P.; Malone, L.A. Developing biosafety risk hypotheses for invertebrates exposed to GM plants using conceptual food webs: A case study with elevated triacylglyceride levels in ryegrass. Environ. Biosafety Res. 2010, 9, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, H.A.; Kok, E.J.; Davies, H.V. New EU legislation for risk assessment of GM food: No scientific justification for mandatory animal feeding trials. Plant Biotechnol. J. 2013, 11, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Poortinga, W.; Pidgeon, N.F. Trust in risk regulation: Cause or consequence of the acceptability of GM food? Risk Anal. 2005, 25, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sisk, J.M.; Gama, L.; Clements, J.E.; Witwer, K.W. Tristetraprolin expression and microRNA-mediated regulation during simian immunodeficiency virus infection of the central nervous system. Mol. Brain 2013, 6, 40. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Hirschi, K.D.; Farmer, L.M. Dietary RNAs: New Stories Regarding Oral Delivery. Nutrients 2015, 7, 3184-3199. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7053184
Yang J, Hirschi KD, Farmer LM. Dietary RNAs: New Stories Regarding Oral Delivery. Nutrients. 2015; 7(5):3184-3199. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7053184
Chicago/Turabian StyleYang, Jian, Kendal D. Hirschi, and Lisa M. Farmer. 2015. "Dietary RNAs: New Stories Regarding Oral Delivery" Nutrients 7, no. 5: 3184-3199. https://0-doi-org.brum.beds.ac.uk/10.3390/nu7053184