
Brucella: Reservoirs and Niches in Animals and Humans


Aix Marseille Univ, CNRS, INSERM, CIML, Centre d’Immunologie de Marseille-Luminy, 13288 Marseille, France
*
Authors to whom correspondence should be addressed.
†
Co-senior authorship.
Pathogens 2021, 10(2), 186; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020186
Submission received: 14 January 2021
/
Revised: 2 February 2021
/
Accepted: 5 February 2021
/
Published: 9 February 2021
(This article belongs to the Special Issue Host Immune Responses and Pathogenesis to Brucella spp. Infection)
{kind=link}
Abstract
:Brucella is an intracellular bacterium that causes abortion, reproduction failure in livestock and leads to a debilitating flu-like illness with serious chronic complications if untreated in humans. As a successful intracellular pathogen, Brucella has developed strategies to avoid recognition by the immune system of the host and promote its survival and replication. In vivo, Brucellae reside mostly within phagocytes and other cells including trophoblasts, where they establish a preferred replicative niche inside the endoplasmic reticulum. This process is central as it gives Brucella the ability to maintain replicating-surviving cycles for long periods of time, even at low bacterial numbers, in its cellular niches. In this review, we propose that Brucella takes advantage of the environment provided by the cellular niches in which it resides to generate reservoirs and disseminate to other organs. We will discuss how the favored cellular niches for Brucella infection in the host give rise to anatomical reservoirs that may lead to chronic infections or persistence in asymptomatic subjects, and which may be considered as a threat for further contamination. A special emphasis will be put on bone marrow, lymph nodes, reproductive and for the first time adipose tissues, as well as wildlife reservoirs.
1. Introduction
Brucella is a Gram-negative facultative intracellular coccobacillus, responsible for brucellosis, a worldwide zoonotic disease that affects livestock, wildlife, and humans. Brucellosis remains endemic in many parts of the world, including the Middle East, Africa, Latin America, Central Asia, and several regions of the Mediterranean [1,2,3,4]. Brucella melitensis, Brucella abortus, and Brucella suis are the most important members of the genus because they are responsible for the human disease [5,6] and for significant economic losses in livestock [7,8], while Brucella canis, Brucella neotomae, and Brucella ovis display less or non-zoonotic potential [9]. Recently, many other species have been described in aquatic mammals (like Brucella pinnipedialis and Brucella ceti) and other wildlife such as Brucella papionis, Brucella microti, Brucella inopinata, and Brucella vulpis [10,11,12,13,14,15,16].
1.1. Brucellosis, a Zoonosis
Humans are accidental hosts for Brucella and are mainly infected through direct contact with infected animals, inhalation of airborne agents, or by ingestion of contaminated dairy products [17]. Human-to-human transmission may occur by organ transplantation, blood transfusion, or vertical transmission via breastfeeding [18,19,20,21,22]. Human brucellosis is at the origin of many symptoms namely undulating fever, malaise, fatigue, and anorexia. If untreated, it may progress into a chronic phase, characterized by the appearance of severe complications like endocarditis, orchitis, spondylitis, osteomyelitis, arthritis, meningoencephalitis, and recurring febrile conditions [23,24,25,26].
In domestic animals, such as cattle, sheep, goats, and swine, major consequences include abortion and metritis in females, and orchiepididymitis and infertility in males [27], resulting in reduced fertility and a significant decline in milk production [28]. Animal brucellosis is highly contagious for both animals and humans, and cross-species transmission of certain Brucella spp. exists [29]. Natural infection occurs by direct contact with infected animals or their secretions [30], like aborted fetuses and fetal membranes that contain large amounts of the bacteria [31].
1.2. Infection and Dissemination
Brucella organisms enter into their host through the mucosal membranes of the respiratory and digestive tracts [32]. Once inside, local professional phagocytes such as macrophages, dendritic cells, and neutrophils internalize the bacteria and move to the closest draining lymph nodes following the normal sampling of the immune system. This leads to subsequent dissemination to the different organs of the reticuloendothelial system, including lungs, spleen, liver, and bone marrow [33]. In pregnant animals, Brucella displays a strong tropism for placental trophoblasts [34,35,36] and also for mammary glands [37], in which it replicates extensively causing placentitis and abortion in the last trimester of pregnancy in ruminants [33]. In humans, brucellosis is a systemic infection and any organ can become infected, albeit with some predilection for joints and liver, and at lower levels for the brain and heart [38].
1.3. Acute and Chronic Infections
Human brucellosis presents a broad range of clinical manifestations from asymptomatic to severe disease. When symptoms are present, the incubation period is 1–4 weeks, but sometimes it takes several months with or without signs and symptoms. According to the duration of the symptoms, human brucellosis is classified into three different phases: acute (initial, 2 months), sub-acute (2–12 months), and chronic (more than 12 months) [39,40]. When focusing on the chronic phase, patients fall into three more categories: relapse (with fever or high IgG antibody titers after antibiotics therapy), chronic localized infection (recurrence of infectious foci and intermittent fever for long periods of time), and delayed convalescence (persistence of some symptoms without fever or other objective signs) [41].
In animals, brucellosis comprises three phases: the onset of the infection or incubation period, when Brucella invades the host without any clinical symptoms; the acute phase, when the bacteria replicate actively and infection remains unapparent in most cases or first pathological signs arise; and the chronic phase, during which bacterial loads reach a plateau before decreasing and sporadic clinical symptoms become visible, the infection localizing in sexually mature animals in the reproductive system to produce epididymitis or orchitis in males, and placentitis and abortion in females. Susceptibility to infection in females increases during the late pregnancy stages (third trimester of gestation) [31,42]. There is no pyrexia like in humans and death is rare; infection is self-limiting most of the time and does not involve other systemic lesions, but for Brucella suis infection in swine. Clinical symptoms for the latter include spondylitis of the lumbar and sacral regions, arthritis, paralysis of hind limbs, lameness, and abscesses in tissues [42]. In any case, chronicity relies on the continuous shedding of Brucella from the mammary gland or reproductive organ secretions for a protracted period. Therefore, infertility, repetitive abortions, and premature stillborn ensure Brucella’s permanence in the environment and liability for dissemination.
The long incubation period and the absence of obvious clinical signs in infected animals and patients correlate with low activation of innate immunity [43]. Brucella has indeed modified its pathogen-associated molecular patterns (PAMPs) into less detectable molecules, opening a permissive immunological window to spread stealthily throughout the reticuloendothelial system and find its target host cells [44]. Then, invasion of target cells occurs thanks to specific molecular determinants that drive not only ingress [45] but also resistance to intracellular killing allowing Brucella to reach its intracellular replicative niche within professional and non-professional phagocytes [46]. Even if innate immunity first restrains Brucella proliferation, a Th1 response with IFNγ and IL-12 production is absolutely required to eradicate it, leading either to complete clearance of the bacteria or to a chronic infection, which arises from the ability of Brucella to persist undetected for prolonged periods of time within its host reservoirs [44,47].
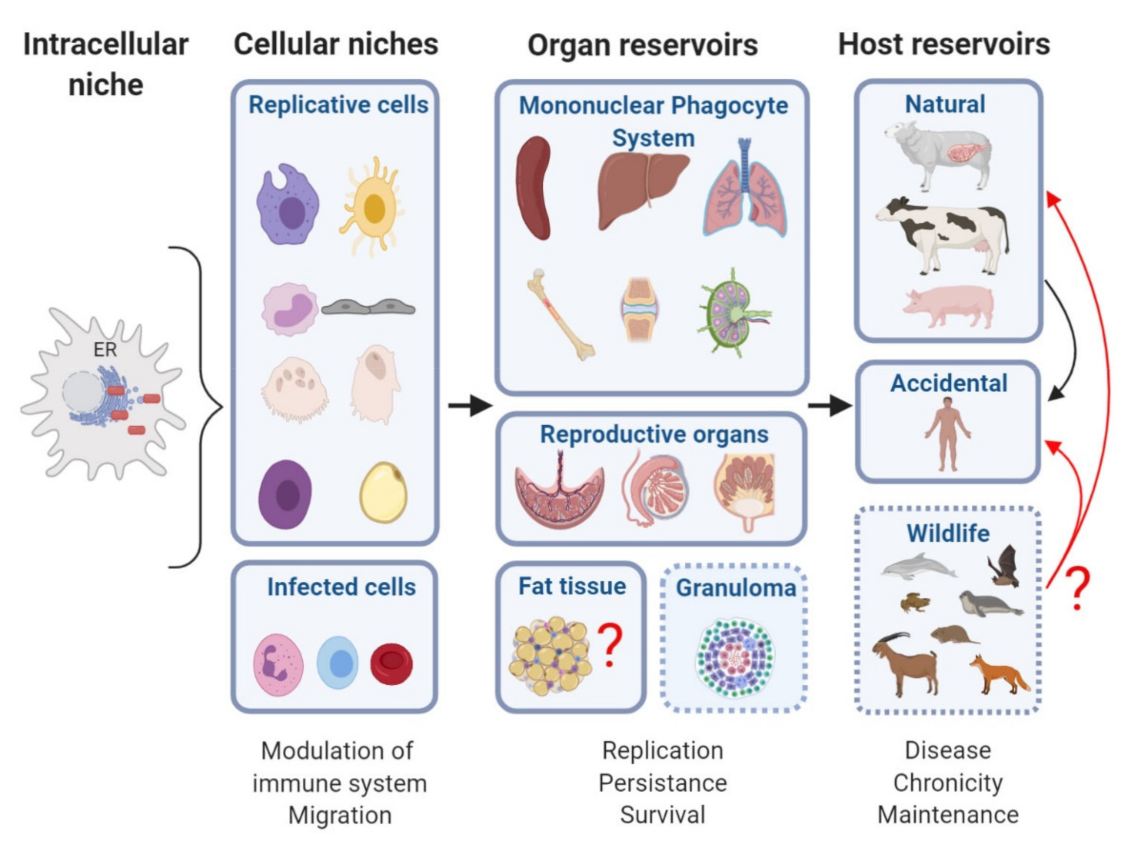
We propose that the intracellular replication niche conditions the basis for the establishment of the reservoirs in which Brucella persists inside its host. What is currently known of the suitable niches for Brucella infection in its various hosts and of the reservoirs of Brucella that support chronic infection?
2. Niche and Reservoir
2.1. Definitions
The concepts of niche and reservoir may be extrapolated to the infection context. In general, Niche is referred to as the “interrelationship of a species and its relational position in a particular ecosystem including the relationship of the species with the components of the ecosystem itself” [48]. As such, the niche may be influenced by all the factors included in its ecosystem and the niche of a species in a particular ecosystem helps setting up the features of its environment, as the latter ones are crucial for its survival [48]. A specific niche is thus defined as: (i) the specific area where an organism inhabits, (ii) the role or function of an organism or species in an ecosystem, and (iii) the interrelationship of a species with all the factors affecting it. The niche of an organism depicts how it lives and survives as a part of its environment. Brucella is a pathogenic genus exceedingly well-adapted to its hosts, which does not survive for extended periods of time in open conditions. This is why it has been called a facultative extracellular intracellular parasite [49]. It also means that Brucella has a niche in the intracellular environment of host cells that is specific in a specific cell of the host. This environment sustains extensive replication, in order to facilitate bacterial expansion and subsequent transmission to new host cells, frequently achieved for example, through the heavily infected tissues like the aborted fetus [49].
In the case of Reservoir, the term is referred to an ecologic species that maintain live circulating organisms through the ecosystem over time. For instance, within the niche from a host reservoir, Brucella can remain at a low replication rate for a long time, and under favorable conditions, egress to infect other cells and start new replicative cycles. In addition, Brucella infections in humans perdure in the ecosystem due to the lack of control of the infection in natural hosts [6,50]. Lymph nodes, spleen, lungs, and the reproductive organs, including placenta, testicular and mammary glands, are well-known target organs for Brucella infection [27]. Some predilection for joint articulations has also been reported in human brucellosis [23], meaning that all these organs allow Brucella replication and that some may contribute to persistence.
2.2. Intracellular Niche
Brucella replicates extensively in the endoplasmic reticulum (ER) compartment within host cells. The mechanisms of entry of the bacterium are still elusive but involve lipid raft-, adhesin- and opsonin-dependent processes [51,52,53,54]. After internalization, Brucella transits inside the cell engulfed in a phagosome, and multiple virulence factors help the bacteria evade the phagocytic pathway by restricting fusion of the Brucella containing vacuole (BCV) with a lysosome. These factors include the cyclic beta-1,2-glucan that operates most probably via cholesterol release [55], the two-component system BvrR/BvrS [56], SepA, which proceeds by excluding the LAMP1 lysosomal protein and preventing the maturation of an active lysosome [57], RicA that regulates vesicle trafficking [58,59] and other possible proteins secreted by the type IV secretion system (T4SS), encoded by the virB operon [60,61]. In vitro experiments using macrophage cell lines have indeed shown that the T4SS is required for maturation of the BCV into an ER-like compartment [62]. Brucella strains lacking a functional T4SS are unable to escape the fusion with lysosomes, and therefore, highly attenuated in mice and in their natural hosts [63]. Once Brucella has impaired phagosome-lysosomal fusion, it replicates in the ER compartment, its replicative niche. Following replication, BCVs interact with host autophagic proteins Beclin1, ULK1, Atg14, and the IRE1α-UPR signaling axis for bacterial egress and the start of replication cycles within newly infected cells [64,65,66,67].
Why does the ER make a good intracellular niche for Brucella? The ER is a critical intracellular organelle that not only synthesizes cellular molecules (proteins, lipids, carbohydrates, etc.) but also regulates the transport of the newly synthesized proteins in the exocytic, endocytic and phagosomal pathways. As such, the association of Brucella with the host cell ER, like a few successful intracellular pathogens, is expected to be highly beneficial from a nutrient acquisition perspective. Taking advantage of the biosynthetic routes of the host cell, substantial levels of metabolites and nutrients on a local supply base fulfill the complex nutritional requirements of Brucella and provide optimal bacterial growth at minimum cost [27,68]. Furthermore, when considering the immune response, localization of Brucella in the ER provides an excellent strategy to hide from detection by the immune system and to limit exposure to the cytosolic immune surveillance pathways by avoiding lysosomal fusion for instance. Moreover, the fact that the MHC I peptide loading complex resides in the ER of the two immune cells where Brucella replicates, the dendritic cells (DC) and the macrophages, is likely not meaningless. It may suggest a yet unidentified regulatory role of Brucella at the level of the setting up of the cross-presentation within the ER and explain its predilection for such cell types.
Indeed, the preference of Brucella to replicate within the ER is mostly restricted to phagocytic cells, professional ones as macrophages [69] and DC [70], and non-professional ones, such as placental trophoblasts of pregnant ruminants [35], and fibroblasts [71] or cell lines (Hela) [72]. Even though other cells, including neutrophils [73], lymphocytes [74], and erythrocytes [75], are infectable by Brucella, there is no efficient replication inside, and their function is more associated with bacterial dispersion, conferring a regulatory role of these cells in persistence. Of note, if in most cell types Brucella replicates within an ER-derived compartment, in extravillous HLA-G+ trophoblasts, B. abortus, and B. suis fail to reach the “normal” ER-derived niche, in contrast to B. melitensis, and replicate within single-membrane acidic lysosomal membrane-associated protein 1 (LAMP1)-positive inclusions [76].
3. Gold Organs in Brucellosis
The “gold organs” for nesting Brucella, in which Brucella replicates in cells of the reticular endothelial system, include the spleen, lymph nodes, liver, bone marrow, epididymis, and placenta.
3.1. The Reticuloendothelial System
The reticuloendothelial system was originally described in 1924 by K. Aschoff as a group of cells able to incorporate vital dyes from the circulation, “reticulo” referring to their propensity to form a network or reticulum by their cytoplasmic extensions and “endothelial” referring to their vicinity to the endothelium. In 1969, a group of pathologists proposed another term, the monocyte phagocyte system (MPS) [77]. Nowadays the reticuloendothelial system or MPS embraces a family of cells that include committed precursors in the bone marrow, circulating blood monocytes, tissue macrophages, and DC in almost every organ in the body [78].
Brucella has a predilection for organs rich in reticuloendothelial cells (including spleen, liver, bone marrow, and lymph nodes) and is able to replicate successfully in any of them. Intracellular replication is directly linked to Brucella pathogenicity and it is not a coincidence that in humans, the most frequent clinical features of brucellosis are an enlarged liver in 65% of the cases, splenomegaly in 52% of the cases (from 40 cases), and lymphadenopathies in children [32,79]. Even in the chicken embryo model, replication of B. abortus detected within the rough ER of mesenchymal, mesothelial, and yolk endodermal cells, spreads to all tissues, with the liver and spleen being the most severely infected [80].
In tissues, the typical histopathological response to Brucella infection is a granulomatous inflammation, which contains representative members of the MPS, including macrophages with an epithelioid shape, i.e., with an increased amount of cytoplasm. Examination of biopsies from humans and livestock animals reveals granulomas in the liver, spleen, bone marrow, and other tissues [79,80,81,82]. As such, the initial replication niche of Brucella serves as a platform to establish a chronic infection. Brucella infected animals develop granulomatous inflammatory lesions in lymphoid tissues, including the supramammary lymph nodes, reproductive organs, notably the udder, and sometimes joints and synovial membranes. Those granulomas and their intratissular location are responsible for the chronicity of the disease, which can last for months or years [81,83] and in that respect, resemble the granulomas extensively studied in tuberculosis. In fact, in the absence of antibiotic treatment in the acute phase, Brucella is able to persist for months without causing significant morbidity or mortality. In the acute phase of infection in a resistant mouse model, the C57BL/6 mice, the formation of granuloma (comprising NOSII+ monocyte-derived inflammatory DC, T cells, and granulocytes) is mediated by MyD88, IL-12, and IFNγ and essential for the control of the bacteria [81,83]. However, these granulomas were not detected in a susceptible murine model of infection, the BALB/c mice, at that stage [81,83]. In B. melitensis acutely infected livers, discrete pyogranulomatous inflammatory areas, characterized by a similar influx of neutrophils, macrophages, and monocyte-derived DC, were detected amongst normal hepatocytes in both mouse models [81,83]. At the chronic phase, infected livers displayed well established demarcated infiltration areas of macrophages, lymphocytes, and neutrophils [81]. In chronic granulomas, the presence of lymphocytes is thought to reflect the former activation of the immune system, whereas recruitment of neutrophils suggests that live Brucella is still present. The fact that the granuloma areas were typically found surrounding or associated with liver portal tracts and that neutrophils may function as vehicles for dispersion, according to the Trojan horse model [84], supports a dynamic role of granulomas in the development of Brucella chronicity. Remarkably, granulomas provide a rich nutrient source, as shown for the dormant non-replicative Mycobacterium bacilli that internalize inside the granuloma, lipids from foamy macrophage lipid droplets [85].
3.2. Genital-Reproductive Organs: Placenta and Epididymis
Brucella has a pronounced tropism for genital organs in its natural hosts, placenta in females, and epididymis in males. The placenta is one of the paradisiac organs in terms of replication, containing up to 1014 Brucellae in the cow [86,87]. This particular environment allows high replication rates, leading consequently to abortion, the most common clinical feature of brucellosis in livestock. As the main route of infection in these farm animals is aborted fetuses, this seems to be a very efficient strategy to spread Brucella progeny to new hosts.
Some common properties in these reproductive organs have shed light on Brucella’s tropism. Firstly, high concentrations of erythritol are present in uterine, epididymal, and fetal tissues from ruminants [87,88,89,90]. Why is this important? Erythritol has been shown to be the preferred carbon/energy source for Brucella spp., promoting their massive growth [91]. In addition, the ruminant placenta produces progesterone, which further enhances in vitro B. abortus growth [92]. However, B. abortus vaccine strain S19 is not stimulated by erythritol [93,94], although it is capable of causing genital infection and abortion [95]. This suggests the existence of other trophic factors. Indeed, the dominance of fructose over glucose takes place in the placenta of cows, sows, ewes, and to a lesser extent in that of other animals [91,96,97]. The same preference applies to the epididymis, seminal fluids, and oviducts of several mammals [91]. As such, both organs play a trophic role and provide effective sources of carbon, nitrogen, and energy for Brucella spp. [49,91].
Secondly, the immune-privileged status of the testis and semen, and local immunosuppression at the feto-maternal interface in the placenta might also account for Brucella tropism [91].
Thirdly, Brucella preferentially replicates within trophoblasts, highly metabolically active cells that adjust their production of proteins and steroids throughout gestation. Intracellular Brucella likely induces the synthesis of steroids and modifies the metabolism of prostaglandin precursors, such as arachidonic acid, which together with the COX-2 enzyme are essential for Brucella lymph node persistence and subversion of the immune response [98].
Finally, the high hydrophobicity of the outer-membrane of Brucella together with its propensity to replicate within the ER [35,99], may represent an evolutionary adaptation for using hydrophobic substances available within this sub-cellular compartment in trophoblasts [49].
In humans, the genital tropism holds true as Brucella induces epididymorchitis [100] and may infect the placenta, even if abortion is very uncommon [76,101].
Therefore, both the localization and abundant multiplication in the reproductive tract of animals is crucial in the biology of this pathogen.
4. Reservoirs
4.1. Bone Marrow
The presence of Brucella organisms in humans has been highlighted in the bone marrow in both the acute and chronic phases. In the mouse, the infection remains sequestered within bone marrow cells for a prolonged period of time (until more than 3 months), without significant changes in the bacterial load [102]. For this reason, the bone marrow has recently been proposed as a reservoir [102].
4.1.1. Bone Marrow in the Mouse Model
Murine models have validated bone marrow as an intermittent colonized organ by Brucella. Indeed 3 weeks p.i., Brucella melitensis has been detected in multiple sites of the murine axial skeleton by in vivo imaging, and immunohistochemistry confirmed its presence in bones, particularly in the lower spine vertebrae, where it preferentially located in a small subset of IBA-1+ monocytes [103]. Similarly, B. canis bacterial loads increased in mouse bone marrow from 9 weeks p.i. onward till 12 weeks, a time which coincides with the persistence and chronicity of the infection [104]. Recently, Gutiérrez et al. observed that Brucella abortus burden remained constant in bone marrow for up to 168 days p.i., including the acute, chronic, and chronic declining phase in the murine model [102]. However, histopathological alterations varied accordingly to the stage of infection. A granulomatous inflammation, accompanied by augmented numbers of myeloid granulocyte-monocytes progenitors (GMP), granulocytes, and CD4+ lymphocytes, was more severe and diffuse during the acute phase than that of the multifocal chronic phase [102].
The vast number of granulomas in the bone marrow and most importantly their permanence indicates the difficulty for immune cells to eliminate B. abortus in such an environment. Interestingly, the cells harboring Brucella in higher proportion were granulocytes, monocytes, and GMP progenitors. The fact that monocytes are constantly infected and controlled by Brucella strongly suggests that bone marrow is a proper reservoir for persistence in brucellosis.
Moreover, the possibility that Brucella infects progenitor stem cells, which lack developed phagocytic machinery, is striking. Recently, it has been shown that hematopoietic stem cells (HSC) sense pathogens, eliciting enhanced myeloid commitment to promoting pathogen clearance of E. coli and Salmonella Typhimurium [105,106]. In the case of Brucella, an increased transient myeloid commitment is triggered by the interaction of Brucella Omp25 and host SLAMF1 in the bone marrow, favoring bacterium survival [107]. This may be one of the multiple strategies developed by Brucella to promote chronic infection and drive bacterial dissemination.
4.1.2. Bone Marrow in Humans
Bone marrow infection by Brucella also occurs in humans in both acute and chronic phases. Usually, a definitive diagnosis of brucellosis is made by culturing Brucella from body fluids or tissues [108,109]. The bone marrow has been a recommended tissue to investigate in suspicious cases of brucellosis when the blood culture test is negative. Gotuzzo et al. [110] determined that out of fifty patients, 92% of Brucella yielded in bone marrow versus 70% in blood. As mentioned above, granulomas are formed as a reaction to particulate or indigestible agents that persist in tissues for long periods. Additionally, 25% of Brucella-infected bone marrows show granulomas together with other pathological changes such as hypercellularity (73% of all cases) or hemophagocytosis (31%) [108,111]. These conglomerates of macrophages, which try to destroy the microbial agent and interact at the same time with lymphocytes might explain rare cases of pancytopenia that are observed in some patients with brucellosis [112].
Human to human brucellosis transmission is extremely rare. In fact, the transmission is due to external factors that promote the transfer of Brucella. Bone marrow transplantation facilitates such a transfer, as a concentration of bone marrow cells that are sheltering Brucella are transferred to a new host. This means that even in the case of an asymptomatic patient where Brucella is present, hidden within bone marrow cells, it will then replicate in the acute phase, or remain there establishing a chronic infection in the recipient host [113,114].
Bones appear to be support structures metabolically inert and resistant to infection by pathogens. However Brucella has a tropism for such location and osteoarticular brucellosis is the most common complication in Brucella-infected humans (40% from total complications) involving sacroiliitis, spondylitis, and peripheral arthritis [38]. Importantly, osteoarticular lesions are also reported in natural hosts, as infected cattle may exhibit bursitis, arthritis, and hygromas [115,116]. Previous studies have demonstrated the ability of B. abortus to invade and replicate within osteoblasts (bone-forming cells), osteocytes (bone-resorbing cells), osteoclasts (multinucleated giant formed by monocyte fusion) [117,118], and in the ER of primary human synoviocytes [119]. Remarkably, osteoclasts originate from the same myeloid precursor cells that give rise to macrophages and myeloid DC and many of the soluble factors (cytokines and growth factors) of immune cells that are regulated by Brucella, may regulate the activities of osteoblasts and osteoclasts. A common feature of patients with osteoarticular brucellosis is the presence of leukocyte infiltrates (including monocytes and neutrophils) in the synovial fluid of the joints [120]. The fact that Brucella inhibits apoptosis in some of those cells or uses them as a vector to spread to other organs, suggests that its osteoarticular location acts as a reservoir of bacteria to progress towards chronicity.
4.1.3. Bone Marrow Environment
Blood cell formation or hematopoiesis is constantly occurring in the bone marrow, starting from self-replicating-pluripotent HSC, which give rise to multipotent progenitors and further on to lineage-committed cells, such as common lymphoid progenitors (CLP) or common myeloid progenitors (CMP). This sequence of events is tightly regulated by key cytokines and growth factors. The need for such a regulated process suggests the existence of a proper nursing environment called “HSC niche” provided by non-hematopoietic cells and by the undefined architecture of the bone marrow if compared to the spleen or lymph node. Cells forming the niche of HSC have to stay close to the progenitors, produce growth and maintenance of soluble factors, sense signals, and respond to the surrounding environment. This group is made up of endothelial, mesenchymal stem cells, macrophages, DC, granulocytes, megakaryocytes, and lymphocytes. In the HSC niche, amongst others, neutrophils modulate endothelium to produce vascular and HSC regeneration, while DCs control vascularity permeabilization processes [121]. Macrophages physically retain the HSC in the bone marrow and regulate HSC fate by fine-tuning the expression of pro-hematopoietic factors in other cells [122]. Moreover, macrophages via activation of cholesterol sensing LXR receptors eat apoptotic neutrophils that return to the bone marrow to be cleared and allow hematopoietic progenitor release into the blood circulation [123]. Memory CD8+ T lymphocytes and CD4+ Tregs are also localized in the bone marrow and protect HSC and downstream progenitors from various types of stress including inflammation [124].
The bone marrow environment is not only beneficial for HSC, pathogens have also taken advantage of those favorable conditions. Mycobaterium tuberculosis is found in mesenchymal cells and HSC of the bone marrow [125,126], which can also host Listeria monocytogenes. However, Listeria persists rarely in the bone marrow of humans and mouse bones, from where it ultimately seeds the central nervous system [127]. As mentioned before, Brucella is able to infect GMP, macrophages, and granulocytes. Brucella bone marrow infection is a successful strategy for the following reasons.
- (i)
- Colonization of an immune-privileged organ that lacks intracellular antibacterial mechanisms in early-stage stem cells is easier.
- (ii)
- Expression of drug efflux pumps by bone marrow cells facilitates resistance to antibiotic therapy.
- (iii)
- Cells from bone marrow are highly motile and reside in close proximity to arterial vessels thus offering a wide distribution system for dissemination of the pathogen to target organs.
- (iv)
- Brucella’s control of HSC release might explain the splenomegaly observed in human cases as a result of the signal triggered in the bone marrow niche by the apoptotic infected neutrophils to the macrophages, which promotes ensuing cycles of HSC release and extramedullary hematopoiesis.
- (v)
- All Brucella preferential target cells differentiate from progenitor stem cells in the bone marrow.
Moreover, the fact that Brucella is transmittable by bone marrow transplantation in humans and adoptive transfer experiments in the mouse [107,128], indicates that Brucella is able to infect and persist for prolonged periods of time in bone marrow progenitors. It also means that the differentiated infected cells might derive from a GMP, which was already infected and recolonized target tissues in relapse or chronic cases.
4.2. Lymph Nodes
Lymph nodes (LNs) are essential compartments of the immune system, composed of highly organized dynamic leukocyte aggregates, where pathogen defense and adaptive immunity take place [129]. A tightly controlled balance of responses upon challenge results in the induction of either tolerance or immunity [130]. LNs filter fluids from the lymphatic vessels that form an extensive network connecting one LN to another. As such, this network provides conduits for the trafficking of DCs, neutrophils, macrophages, and T cells along the process of immune activation after capturing, transportation, and presentation of antigens in regional lymph nodes [131].
Some bacterial pathogens have exploited the lymphatic system for host colonization in LNs and systemic dissemination [132]. Lymphadenopathy is one of the most common signs of brucellosis in humans, present in approximately more than one-third of cases [4,32,133]. As an example, out of 307 children diagnosed with brucellosis, 112 (36%) exhibited lymphadenopathy [32]. LNs most commonly affected are cervical and axillary LNs due to their proximity to the oral route of infection, the natural route of infection for Brucella in humans.
In ruminants, B. abortus and B. melitensis have a marked affinity for mammary glands and reproductive organs together with the supramammary and genital LNs [134,135,136]. In goats, about two-thirds of infections acquired naturally during pregnancy lead to infection of the udder and excretion of the bacteria in the milk during successive lactations [137]. In contrast, in sheep, excretion of bacteria in the milk does not last more than two months generally, continuing for up to 140 or 180 days exceptionally [137].
In camels, B. abortus and B. melitensis have been isolated from LNs, and other organs or compounds like milk, aborted fetus, and placenta [138,139].
The constant or intermittent shedding of Brucella in the milk and genital secretions is held by the colonization of LNs and the subsequent spreading through lymphatics. Hence, Brucellae have been detected in macrophages [32] and neutrophils in lymphatic draining sites of inoculation or in mammary gland LNs [136]. Since LN leukocytes eventually enter the blood, transportation of infected phagocytes in the lymphatic network might disseminate bacterial organisms throughout the host. In fact, the spleen and the iliac, mammary, and prefemoral LNs are the most reliable samples for isolation purposes in necropsied animals thus proving efficient dissemination [140]. The persistent infection of LNs leads to a constant or intermittent shedding of Brucella in the system providing a source of persistent infection in the host and for other animals or humans. Indeed, close contacts between farmers and pastoralists with domesticated animals and the frequent consumption of fresh unpasteurized dairy products increase the maintenance of Brucella and the risk of brucellosis in rural and pastoral areas.
4.3. Adipose Tissue
Microorganisms show incredible diversity with respect to which environment they preferentially colonize or invade. Recently, the emergence of new investigation techniques has allowed the analysis of the tropism of several microbes in yet unexamined organs. This is the case for adipose tissue. Adipose tissue is no longer studied as inert lipid storage but as a central regulator of energy homeostasis and immunity. In the past years, fat-associated lymphoid clusters (FALCs) have received much attention [141]. These structures are quite peculiar because contrary to secondary lymphoid organs, FALCs lack both a surrounding capsule and structured compartmentalization of cells. In contrast, they are in direct contact with adipocytes at mucosal surfaces, including omental, mesenteric, mediastinal, gonadal, and pericardial fat [141]. Adipose tissue comprises a vast range of cellular and non-cellular components that support a large network of cells. These include fibroblasts, preadipocytes, cells with mesenchymal and hematopoietic stem cell capacity together with myeloid cells (macrophages, neutrophils, etc.), lymphocytes (T cells, B cells, innate lymphoid cells (ILCs)), eosinophils, mast cells, and NK cells [142]. These immune cells mostly aggregate in clusters or are scattered around them.
The presence of myeloid cell precursors and mature macrophages in the FALC milieu suggests that the lymphoid clusters form permissive microenvironments, where progenitor cells may proliferate locally to generate free macrophages within the cavity where they are located. Another feature of FALCs is that they are rich in vascularization; they are always closely associated with both blood and lymphatic vessels. In terms of response to pathogens, a rapid formation of acute or chronic inflammation occurs in FALCs, suggesting a prominent role of their clusters in the formation of local immune responses.
Many microorganisms, from viruses like HIV and SIV, to bacteria, like Mycobacterium tuberculosis, Ricketssia prowazekii, Coxiella burnetti, and parasites, such as Plasmodium berghei, Trypanozoma cruzi, and Trypanozoma brucei, have been shown to infect adipocytes from humans or mice [143,144,145,146,147,148]. Their pattern of infection varies according to each pathogen. Some of them establish an intracellular infection specifically within the adipocyte, whereas others remain close to or surround adipocytes and vasculature, or alternatively infect non-fat cells like macrophages.
Not so long ago, nothing was known about Brucella and adipocytes or fat tissues. One report in 2018 described the presence of B. canis in fat cells of the gastro-splenic ligament, next to lipid droplets and precisely where ER is located, in naturally infected fetuses and neonates [149]. More recently, B. abortus has been shown to replicate in a murine fibroblastic derived adipocyte-like cell line, the 3T3-L1 cells, and in its differentiated adipocyte derivative, albeit with less efficiency [150]. However, in this system, bacterial loads start to decline steadily from 3 days p.i. onwards, suggesting that this cell type may not serve as a long-term cellular niche for Brucella per se.
Based on these new findings, we would like to propose the adipose tissue in its whole as another singular reservoir for Brucella and speculate on the benefits this furtive bacterium might gain from such a location.
- (i)
- Fat tissues are enriched in immune cells as aforementioned. With respect to macrophage populations, M2 polarized ones together with other immune cells recruited locally, like neutrophils, monocytes, and DC, might help to maintain Brucella survival and take control of the immune response during infection as suggested by the anti-inflammatory polarization of adipose tissue macrophages potentiated by chronic T. cruzi infection [145].
- (ii)
- The majority of drugs are lipophilic compounds, meaning that efficient distribution within the adipose tissue is hampered, as illustrated by some HIV treatment failures. In human brucellosis, 4% of cases undergo to chronic phase [151]. This incidence may rise because of delayed administration of antibiotics or its inefficient delivery to specific organs, such as fat tissue, in the acute phase of infection.
- (iii)
- The localization of fat depots in the perigonadal region of rodents, humans, and ruminants [152] and the preference of Brucella to infect epididymis might not just be a simple coincidence and reflect an essential role of adipose tissue in the persistence of this bacteria in reproductive organs. It also suggests that fat tissue provides Brucella with a proper environment, i.e., a rich source of nutrients.
- (iv)
- FALCs respond dynamically to stimuli. They expand in size and numbers in response to acute or chronic peritoneal insult [153]. More specifically, an increase of B1 cells, macrophages, and neutrophils recruited via the high endothelial venules has been described in omental FALCs upon injection of E. coli LPS or infection [154]. FALCs also support a unique population of CD4+ regulatory cells producers of IL-10 called visceral adipocyte tissue-associated (VAT) Tregs, mostly studied in large fat depots like epididymal fat [155] but also present in the omentum, where it likely regulates local immune responses [156]. Given the essential role of IL-10 in promoting B. abortus persistence and pathology [157] and the balance between activation and regulation of immune state that exists in adipocyte tissue, it is conceivable that pathogens, and Brucella, in particular, find a golden reservoir here, in which regulatory cells would control the local immune response and create a tolerogenic environment for progression to chronicity.
5. Reservoirs in Wildlife
In addition to the anatomic reservoirs described above, it is imperative to also consider wildlife host reservoirs. The control of brucellosis in humans depends on the control of disease in livestock. However, the creation of new bridges between livestock and wildlife due to human activity is one of the most important factors in disease transmission. As an example, brucellosis has been eradicated in domestic ruminants from most European countries and wild ruminants were not reckoned important hitherto reservoirs. This view changed recently after the notification of two humans cases and the re-emergence of B. melitensis in a dairy cattle farm [158], suggesting a possible implication of wildlife as B. melitensis infection was identified in a French population of Alpine ibex (Capra ibex) [159]. Interestingly, among the 88 seropositive Alpine ibex tested, 58% showed at least one isolation of B. melitensis from a urogenital organ (testes, genital tract, urine, or bladder) or a lymph node from the pelvic area (supramammary, internal iliac, and inguinal LNs), meaning that active infection in the pelvic area is at risk of shedding Brucella [159]. Moreover, microbiome analysis of bats’ guano in India found B. melitensis affiliated sequences [160], and Brucella sequences were detected in the spleen of two different species of bats from Georgia that were coinfected with Bartonella and Leptospira [161]. These data indicate that bats may serve as a wildlife reservoir of Brucella for grazing goats and sheep.
Regarding other classical Brucella species, infections that are recognized as sustainable in wildlife are B. abortus in buffalo (Syncerus caffer) [162] and bison (Bison bison) [163], B. suis biovar 2 in wild boar (Sus Scrofa), and European hare (Lepus europeaus) [164], B. suis biovar 4 in reindeer (Rangifer tarandus) [165], Brucella ceti in cetaceans (Cetacea) [166], Brucella microti in voles (Microtus arvalis) [167] and red fox (Vulpes vulpes) [168].
Other species genetically related to an atypical group within the genus have been described in different host species: Brucella microti-like in marsh frogs (Pelophylax ridibundus) [11]; Brucella vulpis in the red fox (V. vulpes) [13]; Brucella inopinata in White’s and Denny’s tree frogs, (Ranoidea caerulea and Zhangixalus dennysi) [169] and humans; Brucella papionis in baboons (Papio spp.) [15], without mentioning Brucella strains isolates from lungworms in porpoise (Phocoena phocoena) [170], blue-spotted ribbontail ray (Taeniura lymma) [171] and reptile panther chameleon (Furcifer pardalis) [172].
Interestingly, B. microti positivity was evidenced in mandibular lymph nodes from apparently healthy foxes (V. vulpes) assumed to have been contaminated by rodent predation [168].
In 2017, a B. microti-like strain was identified in internal organs (heart, lung, spleen, kidney, liver, and reproductive organs) and sometimes in hind limb muscles of marsh frogs in a French farm producing frogs for human consumption [11]. The human pathogenicity of amphibian strains has not been formally demonstrated but cannot be ruled out because the pathogenicity of B. microti in wild rodents has been confirmed experimentally in a mouse infection model, with high replication rates in murine macrophages [173].
In cetaceans, B. ceti has also been recovered from mandibular, pulmonary, mesenteric, and gastric lymph nodes, spleen, liver, joints, urinary system, and other organs. Noteworthily, Brucellae have been isolated from the female reproductive system, mammary glands, milk and placenta, and in multiple fetal organs, resembling the pathology of terrestrial animals [3,174]. However, it is not the case for pinnipeds (Pinnipedia), where most of the isolates came from the spleen, liver, lungs from healthy animals with non-associated pathology. The risk of contamination due to direct contact between coast animals or due to occupational exposure or direct contact with infected aquatic mammals or fomites for humans increases [174].
In both groups, marine mammal parasites, such as lungworms (Parafilaroides spp., Otostrongylus circumlitus, and Pseudalius inflexus), can serve as vectors of marine Brucellae [170,175,176]. In pinnipeds, lungworms are shed in the feces of an infected marine mammal host into the water, then eaten by coprophagic fish; the worms then migrate from the host gastrointestinal tract to their lungs [177], supporting the maintenance and distribution of Brucella in aquatic reservoirs.
Recently, B. pinnipedialis was recovered for at least 28 days from experimentally infected Atlantic codfish (Gadus morhua), suggesting fish as new potential bacterial reservoirs for Brucella spp. [178]. Most intriguingly, B. melitensis (biovar 3) has been isolated from experimentally and naturally infected Nile catfish (Clarias gariepinus) in the delta region of the Nile in Egypt [179]. These findings raise a concern about the role fish and possibly invertebrates may have in transmission or as reservoirs of infection in aquatic environments and for humans, who ingest or handle raw seafood.
B. inopinata was originally described in human infections [12,180]. With the current isolation of B. inopinata-like (B13-0095) strain from Pac-Man frogs (Ceratophyrus ornate) [181] and the first human case of brucellosis caused by an isolate whose genome is identical, the role of wildlife reservoirs should not be underestimated [182].
Considering that multiple Brucella species circulate in totally different hosts and environments in wildlife, control and eradication strategies that had succeeded for livestock contexts require adaptation to wildlife reservoir conditions. For instance, Alpine ibexes and goats after experimental conjunctival vaccination with B. melitensis Rev.1 vaccine strain display differences in tissue localization and shedding of the bacteria, as well as humoral immune responses [183]. Likewise, retrospective analysis of the effect of vaccination of elks (Cervus canadensis), attending winter feed grounds and adjacent areas of western Wyoming, USA, with the S19 vaccine revealed a failure at reducing post-vaccination seroprevalence of B. abortus [184]. Vaccination in wildlife reservoirs involves additional challenges to face, as vaccines need to be validated for safety and efficacy in wild animals. Delivery route and cost being also important issues, its application is by far more complicated than that for livestock [185].
Epidemiological surveillance of brucellosis in terrestrial and marine wildlife reservoirs takes place via several methodologies, amongst which serology is favored [186]. In terrestrial wild animals, if serology is the most commonly applied diagnostic approach, PCR appears to be the most sensitive (36.62% of positive results). Isolation from blood samples and visceral organs constitutes the great majority of specimens used for the detection of Brucella spp., noting again lymph nodes as a highly prevalent reservoir (94.6%) [187]. Panorama for surveillance in wildlife is challenging given the diversity of laboratory tests, animal species, environments, cross-reactivity, and non-validation of tests for wildlife [188]. This results in uncertain estimates by serological means of the true prevalence for brucellosis in wildlife, requiring cautious interpretations and other technics when available.
Wildlife reservoirs raise major issues in brucellosis. It is clear that carrying or shedding Brucella by wild animals bring a potential risk associated with transmission, persistence, and control in this population as well as domestic ones. However, it is still unknown if wildlife hosts are preferential hosts and if wildlife infections represent a critical reservoir of Brucella strains for livestock and therefore humans. New results are needed to elucidate the infection cell cycle of Brucella in cells of wildlife and better understand the pathological traits of infection related to disease or persistence.
6. Conclusions
Thanks to its stealthy nature, upon infection, Brucella after entering inside a cell, reaches its intracellular niche, the ER, to replicate sheltered from detection by the immune system. This process is central to Brucella as it gives the bacterium the ability to maintain replicating-surviving cycles for long periods of time, even at low bacterial numbers, in its cellular niches. Eventually, Brucella will take advantage of the environment provided by its anatomic reservoirs, where the cellular niches reside, to disseminate to other organs, where high replication rates can occur. Of course, an organ reservoir would not exist without a pre-existing intracellular niche. It is also generally well interconnected with the other organs of the host to facilitate dissemination. Secretions and products of natural hosts of Brucella, livestock, and wildlife, contribute to contamination spreading. Figure 1 illustrates the journey of Brucella inside its hosts and recapitulates the intracellular replicative niche, cellular niches, organ reservoirs, and various hosts described in this report. In this challenging time where the world has seen the rapid emergence of a new viral zoonosis transmitted most probably from a pangolin, it is essential to better understand how another zoonosis, such as brucellosis, develops in its numerous wildlife hosts including bats, and livestock ones. Unraveling the molecular and cellular bases of Brucella host preference and reservoirs should be continued to preclude opportunities for Brucella to jump hosts.
Moreover, the persistence of viable furtive bacteria for extended periods of time highlights the ability of Brucella to maintain a chronic state, a feature that complicates brucellosis treatment, control, and eradication programs. A deeper understanding of the different organ reservoirs of Brucella should help to design new therapies, which would overcome the inability of current treatments to reach this surreptitious bacterium in certain cells and organs, as is the case for the bone marrow of infected patients.
Brucella niche is distributed in different anatomic reservoirs in the host and especially in some organs, such as the adipose tissue, the role of which in brucellosis is still speculative. This opens up new avenues of research that will undoubtedly contribute to a deeper knowledge of brucellosis and more generally of the mechanisms leading to the chronicity of intracellular pathogens.
Author Contributions
Conceptualization, G.G.-E., V.A.-G., S.M. and J.-P.G.; supervision, S.M. and J.-P.G.; writing-original draft preparation, G.G.-E. and S.M.; project administration, S.M. and J.-P.G. All authors have read and agreed to the published version of the manuscript.
Funding
Work is supported by institutional funding from the Centre National de la Recherche Scientifique (CNRS) and the Institut National de la Santé et de la Recherche Médicale (INSERM), and by the Fondation pour la Recherche Médicale (FRM) grant DEQ20170336745, “Laboratories of excellence” programme (Labex INFORM, ANR-11-LABX-0054), the Excellence Initiative of Aix-Marseille Université (AMU) (A*MIDEX), a French “Investissements d’Avenir” programme. GGE is a recipient of o fellowship from the Fondation de Coopération Scientifique “Méditerranée Infection” and from Campus France.
Acknowledgments
Authors thank all members of the JPG lab for their support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fouskis, I.; Sandalakis, V.; Christidou, A.; Tsatsaris, A.; Tzanakis, N.; Tselentis, Y.; Psaroulaki, A. The epidemiology of Brucellosis in Greece, 2007–2012: A ‘One Health’ approach. Trans. R. Soc. Trop. Med. Hyg. 2018, 112, 124–135. [Google Scholar] [CrossRef]
- Guzmán-Hernández, R.L.; Contreras-Rodríguez, A.; Ávila-Calderón, E.D.; Morales-García, M.R. Brucelosis: Zoonosis de importancia en México. Rev. Chil. Infectol. 2016, 33, 656–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Mora, G.; Bonilla-Montoya, R.; Barrantes-Granados, O.; Esquivel-Suárez, A.; Montero-Caballero, D.; González-Barrientos, R.; Fallas-Monge, Z.; Palacios-Alfaro, J.D.; Baldi, M.; Campos, E.; et al. Brucellosis in mammals of Costa Rica: An epidemiological survey. PLoS ONE 2017, 12, e0182644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, R.; Xie, S.; Lu, X.; Sun, L.; Zhou, Y.; Zhang, Y.; Wang, K. A Systematic Review and Meta-Analysis of Epidemiology and Clinical Manifestations of Human Brucellosis in China. Biomed. Res. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, M.P.; Mulder, M.; Gilman, R.H.; Smits, H.L. Human brucellosis. Lancet Infect. Dis. 2007, 7, 775–786. [Google Scholar] [CrossRef]
- O’Callaghan, D. Human brucellosis: Recent advances and future challenges. Infect. Dis. Poverty 2020, 9, 101. [Google Scholar] [CrossRef]
- Bardhan, D.; Kumar, S.; Verma, M.R.; Bangar, Y.C. Economic losses due to brucellosis in India. Indian J. Comp. Microbiol. Immunol. Infect. Dis. 2020, 41, 19. [Google Scholar] [CrossRef]
- Zeng, J.Y.; Robertson, I.D.; Ji, Q.M.; Dawa, Y.L.; Bruce, M. Evaluation of the economic impact of brucellosis in domestic yaks of Tibet. Transbound. Emerg. Dis. 2019, 66, 476–487. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Esquivel, M.; Chaves-Olarte, E.; Moreno, E.; Guzmán-Verri, C. Brucella Genomics: Macro and Micro Evolution. Int. J. Mol. Sci. 2020, 21, 7749. [Google Scholar] [CrossRef]
- Foster, G.; Osterman, B.S.; Godfroid, J.; Jacques, I.; Cloeckaert, A. Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2688–2693. [Google Scholar] [CrossRef] [Green Version]
- Jaÿ, M.; Freddi, L.; Mick, V.; Durand, B.; Girault, G.; Perrot, L.; Taunay, B.; Vuilmet, T.; Azam, D.; Ponsart, C.; et al. Brucella microti-like prevalence in French farms producing frogs. Transbound. Emerg. Dis. 2020, 67, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Scholz, H.C.; Nöckler, K.; Göllner, C.; Bahn, P.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Kämpfer, P.; Cloeckaert, A.; Maquart, M.; et al. Brucella inopinata sp. nov., isolated from a breast implant infection. Int. J. Syst. Evol. Microbiol. 2010, 60, 801–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, H.C.; Revilla-Fernández, S.; Dahouk, S.A.; Hammerl, J.A.; Zygmunt, M.S.; Cloeckaert, A.; Koylass, M.; Whatmore, A.M.; Blom, J.; Vergnaud, G.; et al. Brucella vulpis sp. nov., isolated from mandibular lymph nodes of red foxes (Vulpes vulpes). Int. J. Syst. Evol. Microbiol. 2016, 66, 2090–2098. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Esquivel, M.; Baker, K.S.; Ruiz-Villalobos, N.; Hernández-Mora, G.; Barquero-Calvo, E.; González-Barrientos, R.; Castillo-Zeledón, A.; Jiménez-Rojas, C.; Chacón-Díaz, C.; Cloeckaert, A.; et al. Brucella Genetic Variability in Wildlife Marine Mammals Populations Relates to Host Preference and Ocean Distribution. Genome Biol. Evol. 2017, 9, 1901–1912. [Google Scholar] [CrossRef]
- Whatmore, A.M.; Davison, N.; Cloeckaert, A.; Al Dahouk, S.; Zygmunt, M.S.; Brew, S.D.; Perrett, L.L.; Koylass, M.S.; Vergnaud, G.; Quance, C.; et al. Brucella papionis sp. nov., isolated from baboons (Papio spp.). Int. J. Syst. Evol. Microbiol. 2014, 64, 4120–4128. [Google Scholar] [CrossRef] [Green Version]
- Whatmore, A.M.; Dale, E.J.; Stubberfield, E.; Muchowski, J.; Koylass, M.; Dawson, C.; Gopaul, K.K.; Perrett, L.L.; Jones, M.; Lawrie, A. Isolation of Brucella from a White’s tree frog (Litoria caerulea). JMM Case Rep. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Godfroid, J.; Al Dahouk, S.; Pappas, G.; Roth, F.; Matope, G.; Muma, J.; Marcotty, T.; Pfeiffer, D.; Skjerve, E. A “One Health” surveillance and control of brucellosis in developing countries: Moving away from improvisation. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Ay, N.; Kaya, S.; Anil, M.; Alp, V.; Beyazit, U.; Yuksel, E.; Danis, R. Pulmonary Involvement in Brucellosis, a Rare Complication of Renal Transplant: Case Report and Brief Review. Exp. Clin. Transplant. Off. J. Middle East. Soc. Organ. Transplant. 2018, 16, 757–760. [Google Scholar] [CrossRef]
- Bosilkovski, M.; Arapović, J.; Keramat, F. Human brucellosis in pregnancy—An overview. Bosn. J. Basic Med. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ting, I.-W.; Ho, M.-W.; Sung, Y.-J.; Tien, N.; Chi, C.-Y.; Ho, H.-C.; Huang, C.-C. Brucellosis in a renal transplant recipient. Transpl. Infect. Dis. 2013. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Gondolfo, R.B.; Cerchiari, N. Human-to-human transmission of Brucella—A systematic review. Trop. Med. Int. Health 2017, 22, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Wallach, J.C.; Samartino, L.E.; Efron, A.; Baldi, P.C. Human infection by Brucella melitensis: An outbreak attributed to contact with infected goats. FEMS Immunol. Med. Microbiol. 1997, 19, 315–321. [Google Scholar] [CrossRef]
- Aygen, B.; Doğanay, M.; Sümerkan, B.; Yildiz, O.; Kayabaş, Ü. Clinical manifestations, complications and treatment of brucellosis: A retrospective evaluation of 480 patients. Médecine Mal. Infect. 2002, 32, 485–493. [Google Scholar] [CrossRef]
- Buzgan, T.; Karahocagil, M.K.; Irmak, H.; Baran, A.I.; Karsen, H.; Evirgen, O.; Akdeniz, H. Clinical manifestations and complications in 1028 cases of brucellosis: A retrospective evaluation and review of the literature. Int. J. Infect. Dis. 2010, 14, e469–e478. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.; Splitter, G.A. Molecular Host-Pathogen Interaction in Brucellosis: Current Understanding and Future Approaches to Vaccine Development for Mice and Humans. Clin. Microbiol. Rev. 2003, 16, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Köse, Ş.; Serin Senger, S.; Akkoçlu, G.; Kuzucu, L.; Ulu, Y.; Ersan, G.; Oğuz, F. Clinical manifestations, complications, and treatment of brucellosis: Evaluation of 72 cases. Turk. J. Med. Sci. 2014, 44, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Moriyón, I. The Genus Brucella. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 315–456. [Google Scholar]
- Mcdermott, J.J.; Grace, D.; Zinsstag, J. Economics of brucellosis impact and control in low-income countries. Rev. Sci. Tech. De L’oie 2013, 32, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, S.C.; Palmer, M.V. Advancement of Knowledge of Brucella over the Past 50 Years. Vet. Pathol. 2014, 51, 1076–1089. [Google Scholar] [CrossRef]
- Moreno, E. Retrospective and prospective perspectives on zoonotic brucellosis. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Poester, F.P.; Samartino, L.E.; Santos, R.L. Pathogenesis and pathobiology of brucellosis in livestock. Rev. Sci. Tech. De L’oie 2013, 32, 105–115. [Google Scholar] [CrossRef] [Green Version]
- von Bargen, K.; Gagnaire, A.; Arce-Gorvel, V.; de Bovis, B.; Baudimont, F.; Chasson, L.; Bosilkovski, M.; Papadopoulos, A.; Martirosyan, A.; Henri, S.; et al. Cervical Lymph Nodes as a Selective Niche for Brucella during Oral Infections. PLoS ONE 2015, 10, e0121790. [Google Scholar] [CrossRef] [Green Version]
- Moreno, E.; Barquero-Calvo, E. The Role of Neutrophils in Brucellosis. Microbiol. Mol. Biol. Rev. 2020, 84, e00048-20. [Google Scholar] [CrossRef]
- Anderson, T.D.; Meador, V.P.; Cheville, N.F. Pathogenesis of Placentitis in the Goat Inoculated with Brucella abortus. I. Gross and Histologic Lesions. Vet. Pathol. 1986, 23, 219–226. [Google Scholar] [CrossRef]
- Meador, V.P.; Deyoe, B.L. Intracellular Localization of Brucella abortus in Bovine Placenta. Vet. Pathol. 1989, 26, 513–515. [Google Scholar] [CrossRef]
- Tobias, L.; Cordes, D.O.; Schurig, G.G. Placental Pathology of the Pregnant Mouse Inoculated with Brucella abortus Strain 2308. Vet. Pathol. 1993, 30, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Harmon, B.G.; Adams, L.G.; Frey, M. Survival of rough and smooth strains of Brucella abortus in bovine mammary gland macrophages. Am. J. Vet. Res. 1988, 49, 1092–1097. [Google Scholar] [PubMed]
- Hull, N.C.; Schumaker, B.A. Comparisons of brucellosis between human and veterinary medicine. Infect. Ecol. Epidemiol. 2018, 8, 1500846. [Google Scholar] [CrossRef] [PubMed]
- Amjadi, O.; Rafiei, A.; Mardani, M.; Zafari, P.; Zarifian, A. A review of the immunopathogenesis of Brucellosis. Infect. Dis. 2019, 51, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Pabuccuoglu, O.; Ecemis, T.; El, S.; Coskun, A.; Akcali, S.; Sanlidag, T. Evaluation of serological tests for diagnosis of brucellosis. Jpn. J. Infect. Dis. 2011, 64, 272–276. [Google Scholar] [PubMed]
- Brucellosis in Humans and Animals; Weltgesundheitsorganisation; FAO (Eds.) WHO: Geneva, Switzerland, 2006; p. 89. [Google Scholar]
- Megid, J.; Antonio Mathias, L.; Robles, C. Clinical Manifestations of Brucellosis in Domestic Animals and Humans. Open Vet. Sci. J. 2010, 4, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Barquero-Calvo, E.; Chaves-Olarte, E.; Weiss, D.S.; Guzmán-Verri, C.; Chacón-Díaz, C.; Rucavado, A.; Moriyón, I.; Moreno, E. Brucella abortus Uses a Stealthy Strategy to Avoid Activation of the Innate Immune System during the Onset of Infection. PLoS ONE 2007, 2, e631. [Google Scholar] [CrossRef] [PubMed]
- Martirosyan, A.; Moreno, E.; Gorvel, J.-P. An evolutionary strategy for a stealthy intracellular Brucella pathogen: Brucella pathogenenesis. Immunol. Rev. 2011, 240, 211–234. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Verri, C.; Manterola, L.; Sola-Landa, A.; Parra, A.; Cloeckaert, A.; Garin, J.; Gorvel, J.-P.; Moriyon, I.; Moreno, E.; Lopez-Goni, I. The two-component system BvrR/BvrS essential for Brucella abortus virulence regulates the expression of outer membrane proteins with counterparts in members of the Rhizobiaceae. Proc. Natl. Acad. Sci. USA 2002, 99, 12375–12380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizarro-Cerdá, J.; Desjardins, M.; Moreno, E.; Akira, S.; Gorvel, J.P. Modulation of endocytosis in nuclear factor IL-6(−/−) macrophages is responsible for a high susceptibility to intracellular bacterial infection. J. Immunol. 1999, 162, 3519–3526. [Google Scholar] [PubMed]
- Gomes, M.T.R.; Campos, P.C.; de Almeida, L.A.; Oliveira, F.S.; Costa, M.M.S.; Marim, F.M.; Pereira, G.S.M.; Oliveira, S.C. The role of innate immune signals in immunity to Brucella abortus. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Polechová, J.; Storch, D. Ecological Niche. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 72–80. [Google Scholar]
- Gorvel, J.P.; Moreno, E. Brucella intracellular life: From invasion to intracellular replication. Vet. Microbiol. 2002, 90, 281–297. [Google Scholar] [CrossRef]
- Godfroid, J.; Garin-Bastuji, B.; Saegerman, C.; Blasco, J.M. Brucellosis in terrestrial wildlife. Rev. Sci. Tech. De L’oie 2013, 32, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Watarai, M.; Suzuki, H.; Makino, S.-i.; Kodama, T.; Shirahata, T. Lipid raft microdomains mediate class A scavenger receptor-dependent infection of Brucella abortus. Microb. Pathog. 2004, 37, 11–19. [Google Scholar] [CrossRef]
- Naroeni, A.; Porte, F. Role of Cholesterol and the Ganglioside GM1 in Entry and Short-Term Survival of Brucella suis in Murine Macrophages. Infect. Immun. 2002, 70, 1640–1644. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ranwez, V.; Posadas, D.M.; Van der Henst, C.; Estein, S.M.; Arocena, G.M.; Abdian, P.L.; Martín, F.A.; Sieira, R.; De Bolle, X.; Zorreguieta, A. BtaE, an adhesin that belongs to the trimeric autotransporter family, is required for full virulence and defines a specific adhesive pole of Brucella suis. Infect. Immun. 2013, 81, 996–1007. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ranwez, V.; Posadas, D.M.; Estein, S.M.; Abdian, P.L.; Martin, F.A.; Zorreguieta, A. The BtaF Trimeric Autotransporter of Brucella suis Is Involved in Attachment to Various Surfaces, Resistance to Serum and Virulence. PLoS ONE 2013, 8, e79770. [Google Scholar] [CrossRef] [PubMed]
- Arellano-Reynoso, B.; Lapaque, N.; Salcedo, S.; Briones, G.; Ciocchini, A.E.; Ugalde, R.; Moreno, E.; Moriyón, I.; Gorvel, J.-P. Cyclic β-1,2-glucan is a Brucella virulence factor required for intracellular survival. Nat. Immunol. 2005, 6, 618–625. [Google Scholar] [CrossRef]
- Sola-Landa, A.; Pizarro-Cerdá, J.; Grilló, M.J.; Moreno, E.; Moriyón, I.; Blasco, J.M.; Gorvel, J.P.; López-Goñi, I. A two-component regulatory system playing a critical role in plant pathogens and endosymbionts is present in Brucella abortus and controls cell invasion and virulence. Mol. Microbiol. 1998, 29, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Döhmer, P.H.; Valguarnera, E.; Czibener, C.; Ugalde, J.E. Identification of a type IV secretion substrate of B rucella abortus that participates in the early stages of intracellular survival: A novel Brucella virB secretion substrate. Cell. Microbiol. 2014, 16, 396–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Barsy, M.; Jamet, A.; Filopon, D.; Nicolas, C.; Laloux, G.; Rual, J.-F.; Muller, A.; Twizere, J.-C.; Nkengfac, B.; Vandenhaute, J.; et al. Identification of a Brucella spp. secreted effector specifically interacting with human small GTPase Rab2: A Brucella effector interacting with Rab2. Cell. Microbiol. 2011, 13, 1044–1058. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.P.; Cotto-Rosario, A.; Borghesan, E.; Held, K.; Miller, C.N.; Celli, J. Epistatic Interplay between Type IV Secretion Effectors Engages the Small GTPase Rab2 in the Brucella Intracellular Cycle. mBio 2020, 11, e03350-19. [Google Scholar] [CrossRef] [Green Version]
- Celli, J.; de Chastellier, C.; Franchini, D.-M.; Pizarro-Cerda, J.; Moreno, E.; Gorvel, J.-P. Brucella Evades Macrophage Killing via VirB-dependent Sustained Interactions with the Endoplasmic Reticulum. J. Exp. Med. 2003, 198, 545–556. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, D.; Cazevieille, C.; Allardet-Servent, A.; Boschiroli, M.L.; Bourg, G.; Foulongne, V.; Frutos, P.; Kulakov, Y.; Ramuz, M. A homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl type IV secretion systems is essential for intracellular survival of Brucella suis: Brucella type IV secretion. Mol. Microbiol. 2002, 33, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Comerci, D.J.; Martinez-Lorenzo, M.J.; Sieira, R.; Gorvel, J.-P.; Ugalde, R.A. Essential role of the VirB machinery in the maturation of the Brucella abortus-containing vacuole. Cell. Microbiol. 2001, 3, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.C.; Tsolis, R.M.; Ficht, T.A. Identification of Genes Required for Chronic Persistence of Brucella abortus in Mice. Infect. Immun. 2000, 68, 4102–4107. [Google Scholar] [CrossRef] [Green Version]
- Byndloss, M.X.; Tsai, A.Y.; Walker, G.T.; Miller, C.N.; Young, B.M.; English, B.C.; Seyffert, N.; Kerrinnes, T.; de Jong, M.F.; Atluri, V.L.; et al. Brucella abortus Infection of Placental Trophoblasts Triggers Endoplasmic Reticulum Stress-Mediated Cell Death and Fetal Loss via Type IV Secretion System-Dependent Activation of CHOP. mBio 2019, 10, e01538-19. [Google Scholar] [CrossRef] [Green Version]
- Celli, J. The changing nature of the B rucella-containing vacuole: The Brucella -containing vacuole. Cell. Microbiol. 2015, 17, 951–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Lin, F.; Cabello, A.L.; da Costa, L.F.; Feng, X.; Feng, H.-Q.; Zhang, M.-Z.; Iwawaki, T.; Rice-Ficht, A.; Ficht, T.A.; et al. Activation of Host IRE1α-Dependent Signaling Axis Contributes the Intracellular Parasitism of Brucella melitensis. Front. Cell. Infect. Microbiol. 2018, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.; Child, R.; Wehrly Tara, D.; Hansen, B.; Hwang, S.; López-Otin, C.; Virgin Herbert, W.; Celli, J. Selective Subversion of Autophagy Complexes Facilitates Completion of the Brucella Intracellular Cycle. Cell Host Microbe 2012, 11, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Roy, C.R.; Salcedo, S.P.; Gorvel, J.-P.E. Pathogen–endoplasmic-reticulum interactions: In through the out door. Nat. Rev. Immunol. 2006, 6, 136–147. [Google Scholar] [CrossRef]
- Celli, J. Surviving inside a macrophage: The many ways of Brucella. Res. Microbiol. 2006, 157, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, S.P.; Marchesini, M.I.; Lelouard, H.; Fugier, E.; Jolly, G.; Balor, S.; Muller, A.; Lapaque, N.; Demaria, O.; Alexopoulou, L.; et al. Brucella Control of Dendritic Cell Maturation Is Dependent on the TIR-Containing Protein Btp1. PLoS Pathog. 2008, 4, e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demars, A.; Lison, A.; Machelart, A.; Van Vyve, M.; Potemberg, G.; Vanderwinden, J.-M.; De Bolle, X.; Letesson, J.-J.; Muraille, E. Route of Infection Strongly Impacts the Host-Pathogen Relationship. Front. Immunol. 2019, 10, 1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizarro-Cerdá, J.; Méresse, S.; Parton, R.G.; van der Goot, G.; Sola-Landa, A.; Lopez-Goñi, I.; Moreno, E.; Gorvel, J.-P. Brucella abortus Transits through the Autophagic Pathway and Replicates in the Endoplasmic Reticulum of Nonprofessional Phagocytes. Infect. Immun. 1998, 66, 5711–5724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barquero-Calvo, E.; Mora-Cartín, R.; Arce-Gorvel, V.; de Diego, J.L.; Chacón-Díaz, C.; Chaves-Olarte, E.; Guzmán-Verri, C.; Buret, A.G.; Gorvel, J.-P.; Moreno, E. Brucella abortus Induces the Premature Death of Human Neutrophils through the Action of Its Lipopolysaccharide. PLoS Pathog. 2015, 11, e1004853. [Google Scholar] [CrossRef] [Green Version]
- Goenka, R.; Guirnalda, P.D.; Black, S.J.; Baldwin, C.L. B Lymphocytes Provide an Infection Niche for Intracellular Bacterium Brucella abortus. J. Infect. Dis. 2012, 206, 91–98. [Google Scholar] [CrossRef]
- Vitry, M.-A.; Hanot Mambres, D.; Deghelt, M.; Hack, K.; Machelart, A.; Lhomme, F.; Vanderwinden, J.-M.; Vermeersch, M.; De Trez, C.; Pérez-Morga, D.; et al. Brucella melitensis invades murine erythrocytes during infection. Infect. Immun. 2014, 82, 3927–3938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo, S.P.; Chevrier, N.; Lacerda, T.L.S.; Ben Amara, A.; Gerart, S.; Gorvel, V.A.; de Chastellier, C.; Blasco, J.M.; Mege, J.-L.; Gorvel, J.-P. Pathogenic Brucellae Replicate in Human Trophoblasts. J. Infect. Dis. 2013, 207, 1075–1083. [Google Scholar] [CrossRef]
- Yona, S.; Gordon, S. From the Reticuloendothelial to Mononuclear Phagocyte System—The Unaccounted Years. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Summers, K.M.; Bush, S.J.; Hume, D.A. Network analysis of transcriptomic diversity amongst resident tissue macrophages and dendritic cells in the mouse mononuclear phagocyte system. PLoS Biol. 2020, 18, e3000859. [Google Scholar] [CrossRef]
- Cervantes, F.; Carbonell, J.; Bruguera, M.; Force, L.; Webb, S. Liver disease in brucellosis. A clinical and pathological study of 40 cases. Postgrad. Med. J. 1982, 58, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Detilleux, P.G.; Cheville, N.F.; Deyoe, B.L. Pathogenesis of Brucella abortus in Chicken Embryos. Vet. Pathol. 1988, 25, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daggett, J.; Rogers, A.; Harms, J.; Splitter, G.A.; Durward-Diioia, M. Hepatic and splenic immune response during acute vs. chronic Brucella melitensis infection using in situ microscopy. Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101490. [Google Scholar] [CrossRef]
- Karim, M.F.; Maruf, A.A.; Yeasmin, F.; Shafy, N.M.; Khan, A.H.N.A.; Rahman, A.K.M.A.; Bhuiyan, M.J.S.; Hasan, M.M.; Karim, M.R.; Hasan, M.T.; et al. Histopathological changes of brucellosis in experimentally infected guinea pig. Bangladesh J. Vet. Med. 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- Copin, R.; Vitry, M.-A.; Hanot Mambres, D.; Machelart, A.; De Trez, C.; Vanderwinden, J.-M.; Magez, S.; Akira, S.; Ryffel, B.; Carlier, Y.; et al. In Situ Microscopy Analysis Reveals Local Innate Immune Response Developed around Brucella Infected Cells in Resistant and Susceptible Mice. PLoS Pathog. 2012, 8, e1002575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Jiménez, C.; Mora-Cartín, R.; Altamirano-Silva, P.; Chacón-Díaz, C.; Chaves-Olarte, E.; Moreno, E.; Barquero-Calvo, E. Neutrophils as Trojan Horse Vehicles for Brucella abortus Macrophage Infection. Front. Immunol. 2019, 10, 1012. [Google Scholar] [CrossRef] [Green Version]
- Peyron, P.; Vaubourgeix, J.; Poquet, Y.; Levillain, F.; Botanch, C.; Bardou, F.; Daffé, M.; Emile, J.-F.; Marchou, B.; Cardona, P.-J.; et al. Foamy Macrophages from Tuberculous Patients’ Granulomas Constitute a Nutrient-Rich Reservoir for M. tuberculosis Persistence. Plos Pathog. 2008, 4, e1000204. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.; Schnurrenberger, P.R.; Brown, R.R. Numbers of Brucella abortus in the placenta, umbilicus and fetal fluid of two naturally infected cows. Vet. Rec. 1981, 108, 500. [Google Scholar] [CrossRef]
- Smith, H.; Williams, A.E.; Pearce, J.H.; Keppie, J.; Harris-Smith, P.W.; Fitz-George, R.B.; Witt, K. Fœtal Erythritol: A Cause of the Localization of Brucella abortus in Bovine Contagious Abortion. Nature 1962, 193, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.B.K.; Graham, E.F.; Lewis, B.A.; Smith, F. D-mannitol, erythritol and glycerol in bovine semen. Reproduction 1967, 13, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Essenberg, R.C.; Seshadri, R.; Nelson, K.; Paulsen, I. Sugar metabolism by Brucellae. Vet. Microbiol. 2002, 90, 249–261. [Google Scholar] [CrossRef]
- Keppie, J.; Williams, A.E.; Witt, K.; Smith, H. The role of erythritol in the tissue localization of the brucellae. Br. J. Exp. Pathol. 1965, 46, 104–108. [Google Scholar]
- Letesson, J.-J.; Barbier, T.; Zúñiga-Ripa, A.; Godfroid, J.; De Bolle, X.; Moriyón, I. Brucella Genital Tropism: What’s on the Menu. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Samartino, L.E.; Enright, F.M. Pathogenesis of abortion of bovine brucellosis. Comp. Immunol. Microbiol. Infect. Dis. 1993, 16, 95–101. [Google Scholar] [CrossRef]
- Keppie, J.; Witt, K.; Smith, H. The Effect of Erythritol on the Growth of S19 and other Attenuated Strains of Brucella abortus. Res. Vet. Sci. 1967, 8, 294–296. [Google Scholar] [CrossRef]
- Rodríguez, M.C.; Viadas, C.; Seoane, A.; Sangari, F.J.; López-Goñi, I.; García-Lobo, J.M. Evaluation of the Effects of Erythritol on Gene Expression in Brucella abortus. PLoS ONE 2012, 7, e50876. [Google Scholar] [CrossRef] [Green Version]
- Corner, L.A.; Alton, G.G. Persistence of Brucella abortus strain 19 infection in adult cattle vaccinated with reduced doses. Res. Vet. Sci. 1981, 31, 342–344. [Google Scholar] [CrossRef]
- Alexander, D.P.; Huggett, A.S.G.; Nixon, D.A.; Widdas, W.F. The placental transfer of sugars in the sheep: The influence of concentration gradient upon the rates of hexose formation as shown in umbilical perfusion of the placenta. J. Physiol. 1955, 129, 367–383. [Google Scholar] [CrossRef] [Green Version]
- Hastein, T.; Velle, W. Placental aldose reductase activity and foetal blood fructose during bovine pregnancy. Reproduction 1968, 15, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Gagnaire, A.; Gorvel, L.; Papadopoulos, A.; Von Bargen, K.; Mège, J.-L.; Gorvel, J.-P. COX-2 Inhibition Reduces Brucella Bacterial Burden in Draining Lymph Nodes. Front. Microbiol. 2016, 7, 1987. [Google Scholar] [CrossRef] [Green Version]
- Anderson, T.D.; Cheville, N.F.; Meador, V.P. Pathogenesis of Placentitis in the Goat Inoculated with Brucella abortus. II. Ultrastructural Studies. Vet. Pathol. 1986, 23, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Queipo-Ortuño, M.I.; Colmenero, J.D.; Muñoz, N.; Baeza, G.; Clavijo, E.; Morata, P. Rapid Diagnosis of Brucella Epididymo-Orchitis by Real-Time Polymerase Chain Reaction Assay in Urine Samples. J. Urol. 2006, 176, 2290–2293. [Google Scholar] [CrossRef] [PubMed]
- Al-Tawfiq, J.; Memish, Z. Pregnancy Associated Brucellosis. Recent Pat. Anti-Infect. Drug Discov. 2013, 8, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Jiménez, C.; Hysenaj, L.; Alfaro-Alarcón, A.; Mora-Cartín, R.; Arce-Gorvel, V.; Moreno, E.; Gorvel, J.P.; Barquero-Calvo, E. Persistence of Brucella abortus in the Bone Marrow of Infected Mice. J. Immunol. Res. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Magnani, D.M.; Lyons, E.T.; Forde, T.S.; Shekhani, M.T.; Adarichev, V.A.; Splitter, G.A. Osteoarticular tissue infection and development of skeletal pathology in murine brucellosis. Dis. Models Mech. 2013, 6, 811–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacón-Díaz, C.; Altamirano-Silva, P.; González-Espinoza, G.; Medina, M.-C.; Alfaro-Alarcón, A.; Bouza-Mora, L.; Jiménez-Rojas, C.; Wong, M.; Barquero-Calvo, E.; Rojas, N.; et al. Brucella canis Is an Intracellular Pathogen That Induces a Lower Proinflammatory Response than Smooth Zoonotic Counterparts. Infect. Immun. 2015, 83, 4861–4870. [Google Scholar] [CrossRef] [Green Version]
- Burberry, A.; Zeng, M.Y.; Ding, L.; Wicks, I.; Inohara, N.; Morrison, S.J.; Núñez, G. Infection Mobilizes Hematopoietic Stem Cells through Cooperative NOD-like Receptor and Toll-like Receptor Signaling. Cell Host Microbe 2014, 15, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Takizawa, H.; Fritsch, K.; Kovtonyuk, L.V.; Saito, Y.; Yakkala, C.; Jacobs, K.; Ahuja, A.K.; Lopes, M.; Hausmann, A.; Hardt, W.-D.; et al. Pathogen-Induced TLR4-TRIF Innate Immune Signaling in Hematopoietic Stem Cells Promotes Proliferation but Reduces Competitive Fitness. Cell Stem Cell 2017, 21, 225–240.e225. [Google Scholar] [CrossRef]
- Hysenaj, L.; De Laval, B.; Arce-Gorvel, V.; Bosilkovski, M.; Gonzalez-Espinoza, G.; Debroas, G.; Sieweke, M.; Sarrazin, S.; Gorvel, J.-P. Brucella subverts myeloid commitment of hematopoietic stem cells by a SLAMF1 (CD150)-dependent mechanism. Cell Stem Cell 2021, 26, 657–674. [Google Scholar]
- Demir, C.; Karahocagil, M.K.; Esen, R.; Atmaca, M.; Gönüllü, H.; Akdeniz, H. Bone marrow biopsy findings in brucellosis patients with hematologic abnormalities. Chin. Med. J. 2012, 125, 1871–1876. [Google Scholar]
- Ganado, W.; Bannister, W. Bacteraemia in Human Brucellosis. BMJ 1960, 1, 601–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotuzzo, E.; Carrillo, C.; Guerra, J.; Llosa, L. An Evaluation of Diagnostic Methods for Brucellosis—The Value of Bone Marrow Culture. J. Infect. Dis. 1986, 153, 122–125. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Yrivarren, J.L.; Argumans, C.; Crosby, E.; Carrillo, C.; Gotuzzo, E. Evaluation of the bone marrow in patients with brucellosis. Clinico-pathological correlation. Enferm. Infecc. Y Microbiol. Clin. 1990, 8, 19–24. [Google Scholar]
- Crosby, E.; Llosa, L.; Quesada, M.M.; Carrillo, P.C.; Gotuzzo, E. Hematologic Changes in Brucellosis. J. Infect. Dis. 1984, 150, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Ertem, M.; Kürekçi, A.; Aysev, D.; Ünal, E.; Ikincioğulları, A. Brucellosis transmitted by bone marrow transplantation. Bone Marrow Transplant. 2000, 26, 225–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naparstek, E. Transmission of brucellosis by bone marrow transplantation. Lancet 1982, 319, 574–575. [Google Scholar] [CrossRef]
- Bracewell, C.; Corbel, M. An association between arthritis and persistent serological reactions to Brucella abortus in cattle from apparently brucellosis-free herds. Vet. Rec. 1980, 106, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Corbel, M.J.; Stuart, F.A.; Brewer, R.A.; Jeffrey, M.; Bradley, R. Arthropathy associated with Brucella abortus strain 19 vaccination in cattle. I. Examination of field cases. Br. Vet. J. 1989, 145, 337–346. [Google Scholar] [CrossRef]
- Delpino, M.V.; Fossati, C.A.; Baldi, P.C. Proinflammatory Response of Human Osteoblastic Cell Lines and Osteoblast-Monocyte Interaction upon Infection with Brucella spp. Infect. Immun. 2009, 77, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Khalaf, O.H.; Chaki, S.P.; Garcia-Gonzalez, D.G.; Suva, L.J.; Gaddy, D.; Arenas-Gamboa, A.M. Interaction of Brucella abortus with Osteoclasts: A Step toward Understanding Osteoarticular Brucellosis and Vaccine Safety. Infect. Immun. 2020, 88, e00822-19. [Google Scholar] [CrossRef]
- Scian, R.; Barrionuevo, P.; Giambartolomei, G.H.; De Simone, E.A.; Vanzulli, S.I.; Fossati, C.A.; Baldi, P.C.; Delpino, M.V. Potential Role of Fibroblast-Like Synoviocytes in Joint Damage Induced by Brucella abortus Infection through Production and Induction of Matrix Metalloproteinases. Infect. Immun. 2011, 79, 3619–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giambartolomei, G.H.; Arriola Benitez, P.C.; Delpino, M.V. Brucella and Osteoarticular Cell Activation: Partners in Crime. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Supakorndej, T.; Krambs, J.R.; Rao, M.; Abou-Ezzi, G.; Ye, R.Y.; Li, S.; Trinkaus, K.; Link, D.C. Bone marrow dendritic cells regulate hematopoietic stem/progenitor cell trafficking. J. Clin. Investig. 2019, 129, 2920–2931. [Google Scholar] [CrossRef]
- Winkler, I.G.; Sims, N.A.; Pettit, A.R.; Barbier, V.; Nowlan, B.; Helwani, F.; Poulton, I.J.; van Rooijen, N.; Alexander, K.A.; Raggatt, L.J.; et al. Bone marrow macrophages maintain hematopoietic stem cell (HSC) niches and their depletion mobilizes HSCs. Blood 2010, 116, 4815–4828. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Acebes, M.; Pitaval, C.; Weiss, L.A.; Nombela-Arrieta, C.; Chèvre, R.; A-González, N.; Kunisaki, Y.; Zhang, D.; van Rooijen, N.; Silberstein, L.E.; et al. Rhythmic Modulation of the Hematopoietic Niche through Neutrophil Clearance. Cell 2013, 153, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Saraiva, M.; Gomes, M.S. The bone marrow hematopoietic niche and its adaptation to infection. Semin. Cell Dev. Biol. 2020, S1084952119302617. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Kashino, S.S.; Pulu, I.; Kalita, D.; Swami, V.; Yeger, H.; Felsher, D.W.; Campos-Neto, A. CD271+ Bone Marrow Mesenchymal Stem Cells May Provide a Niche for Dormant Mycobacterium tuberculosis. Sci. Transl. Med. 2013, 5, 170ra113. [Google Scholar] [CrossRef] [Green Version]
- Tornack, J.; Reece, S.T.; Bauer, W.M.; Vogelzang, A.; Bandermann, S.; Zedler, U.; Stingl, G.; Kaufmann, S.H.E.; Melchers, F. Human and Mouse Hematopoietic Stem Cells Are a Depot for Dormant Mycobacterium tuberculosis. PLoS ONE 2017, 12, e0169119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, J.; Chu, P.; Contag, C.H. Foci of Listeria monocytogenes persist in the bone marrow. Dis. Models Mech. 2009, 2, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, E.E. Brucellosis as a Hazard of Blood Transfusion. BMJ 1955, 1, 27–28. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, G.; Ataide, M.; Kastenmüller, W. Lymph node—An organ for T-cell activation and pathogen defense. Immunol. Rev. 2016, 271, 200–220. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurty, A.T.; Turley, S.J. Lymph node stromal cells: Cartographers of the immune system. Nat. Immunol. 2020, 21, 369–380. [Google Scholar] [CrossRef]
- Jackson, D.G. Leucocyte Trafficking via the Lymphatic Vasculature— Mechanisms and Consequences. Front. Immunol. 2019, 10, 471. [Google Scholar] [CrossRef]
- Lynskey, N.N.; Banerji, S.; Johnson, L.A.; Holder, K.A.; Reglinski, M.; Wing, P.A.C.; Rigby, D.; Jackson, D.G.; Sriskandan, S. Rapid Lymphatic Dissemination of Encapsulated Group A Streptococci via Lymphatic Vessel Endothelial Receptor-1 Interaction. PLoS Pathog. 2015, 11, e1005137. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, W.P. The Nature of Brucellosis. Am. J. Trop. Med. Hyg. 1957, 6, 591–592. [Google Scholar] [CrossRef]
- Fensterbank, R. Some aspects of experimental bovine brucellosis. Ann. Rech. Vet. Vet. Ann. Vet. Res. 1987, 18, 421–428. [Google Scholar]
- Meador, V.P.; Deyoe, B.L.; Cheville, N.F. Effect of Nursing on Brucella abortus Infection of Mammary Glands of Goats. Vet. Pathol. 1989, 26, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Meador, V.P.; Deyoe, B.L.; Cheville, N.F. Pathogenesis of Brucella abortus Infection of the Mammary Gland and Supramammary Lymph Node of the Goat. Vet. Pathol. 1989, 26, 357–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scientific Committe on Animal Health and Animal Welfare. Brucellosis in Sheep and Goats (Brucella melitensis). 2001. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/sci-com_scah_out59_en.pdf (accessed on 15 December 2020).
- Damir, H.A.; Tageldin, M.H.; Kenyon, S.J.; Idris, O.F. Isolation of Brucella abortus from experimentally infected dromedary camels in Sudan: A preliminary report. Vet. Res. Commun. 1989, 13, 403–406. [Google Scholar] [CrossRef]
- Foster, J.T.; Walker, F.M.; Rannals, B.D.; Hussain, M.H.; Drees, K.P.; Tiller, R.V.; Hoffmaster, A.R.; Al-Rawahi, A.; Keim, P.; Saqib, M. African Lineage Brucella melitensis Isolates from Omani Livestock. Front. Microbiol. 2018, 8, 2702. [Google Scholar] [CrossRef]
- Marin, C.M.; Jimenez de Bagues, M.P.; Barberan, M.; Blasco, J.M. Comparison of two selective media for the isolation of Brucella melitensis from naturally infected sheep and goats. Vet. Rec. 1996, 138, 409–411. [Google Scholar] [CrossRef]
- Cruz-Migoni, S.; Caamaño, J. Fat-Associated Lymphoid Clusters in Inflammation and Immunity. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef]
- Ferrante, A.W. The immune cells in adipose tissue. Diabetes Obes. Metab. 2013, 15, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Bechah, Y.; Paddock, C.D.; Capo, C.; Mege, J.-L.; Raoult, D. Adipose Tissue Serves as a Reservoir for Recrudescent Rickettsia prowazekii Infection in a Mouse Model. PLoS ONE 2010, 5, e8547. [Google Scholar] [CrossRef]
- Beigier-Bompadre, M.; Montagna, G.N.; Kühl, A.A.; Lozza, L.; Weiner, J.; Kupz, A.; Vogelzang, A.; Mollenkopf, H.-J.; Löwe, D.; Bandermann, S.; et al. Mycobacterium tuberculosis infection modulates adipose tissue biology. PLoS Pathog. 2017, 13, e1006676. [Google Scholar] [CrossRef] [Green Version]
- Cabalén, M.E.; Cabral, M.F.; Sanmarco, L.M.; Andrada, M.C.; Onofrio, L.I.; Ponce, N.E.; Aoki, M.P.; Gea, S.; Cano, R.C. Chronic Trypanosoma cruzi infection potentiates adipose tissue macrophage polarization toward an anti-inflammatory M2 phenotype and contributes to diabetes progression in a diet-induced obesity model. Oncotarget 2016, 7, 13400–13415. [Google Scholar] [CrossRef] [Green Version]
- Damouche, A.; Lazure, T.; Avettand-Fènoël, V.; Huot, N.; Dejucq-Rainsford, N.; Satie, A.-P.; Mélard, A.; David, L.; Gommet, C.; Ghosn, J.; et al. Adipose Tissue Is a Neglected Viral Reservoir and an Inflammatory Site during Chronic HIV and SIV Infection. PLoS Pathog. 2015, 11, e1005153. [Google Scholar] [CrossRef] [Green Version]
- Neyrolles, O.; Hernández-Pando, R.; Pietri-Rouxel, F.; Fornès, P.; Tailleux, L.; Payán, J.A.B.; Pivert, E.; Bordat, Y.; Aguilar, D.; Prévost, M.-C.; et al. Is Adipose Tissue a Place for Mycobacterium tuberculosis Persistence? PLoS ONE 2006, 1, e43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, L. The Parasite in the Fat: A Serendipitous Finding. Trends Parasitol. 2017, 33, 255. [Google Scholar] [CrossRef]
- De Souza, T.D.; de Carvalho, T.F.; Mol, J.P.d.S.; Lopes, J.V.M.; Silva, M.F.; da Paixão, T.A.; Santos, R.L. Tissue distribution and cell tropism of Brucella canis in naturally infected canine foetuses and neonates. Sci. Rep. 2018, 8, 7203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesce Viglietti, A.I.; Giambartolomei, G.H.; Quarleri, J.; Delpino, M.V. Brucella abortus Infection Modulates 3T3-L1 Adipocyte Inflammatory Response and Inhibits Adipogenesis. Front. Endocrinol. 2020, 11, 585923. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Gao, H.; Pappas, G.; Chen, Q.; Li, M.; Xu, J.; Lai, S.; Liao, Q.; Yang, W.; Yi, Z.; et al. Clinical features of 2041 human brucellosis cases in China. PLoS ONE 2018, 13, e0205500. [Google Scholar] [CrossRef] [Green Version]
- Hansel, W. The Essentiality of the Epididymal Fat Pad for Spermatogenesis. Endocrinology 2010, 151, 5565–5567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elewa, Y.H.A.; Ichii, O.; Otsuka, S.; Hashimoto, Y.; Kon, Y. Characterization of mouse mediastinal fat-associated lymphoid clusters. Cell Tissue Res. 2014, 357, 731–741. [Google Scholar] [CrossRef]
- Buscher, K.; Wang, H.; Zhang, X.; Striewski, P.; Wirth, B.; Saggu, G.; Lütke-Enking, S.; Mayadas, T.N.; Ley, K.; Sorokin, L.; et al. Protection from septic peritonitis by rapid neutrophil recruitment through omental high endothelial venules. Nat. Commun. 2016, 7, 10828. [Google Scholar] [CrossRef]
- Cipolletta, D.; Cohen, P.; Spiegelman, B.M.; Benoist, C.; Mathis, D. Appearance and disappearance of the mRNA signature characteristic of T reg cells in visceral adipose tissue: Age, diet, and PPARγ effects. Proc. Natl. Acad. Sci. USA 2015, 112, 482–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meza-Perez, S.; Randall, T.D. Immunological Functions of the Omentum. Trends Immunol. 2017, 38, 526–536. [Google Scholar] [CrossRef]
- Xavier, M.N.; Winter, M.G.; Spees, A.M.; Nguyen, K.; Atluri, V.L.; Silva, T.M.A.; Bäumler, A.J.; Müller, W.; Santos, R.L.; Tsolis, R.M. CD4+ T Cell-derived IL-10 Promotes Brucella abortus Persistence via Modulation of Macrophage Function. PLoS Pathog. 2013, 9, e1003454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garin-Bastuji, B.; Hars, J.; Drapeau, A.; Cherfa, M.-A.; Game, Y.; Le Horgne, J.-M.; Rautureau, S.; Maucci, E.; Pasquier, J.-J.; Jay, M.; et al. Reemergence of Brucella melitensis Infection in Wildlife, France. Emerg. Infect. Dis. 2014, 20, 1570–1571. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.; Gilot-Fromont, E.; Freycon, P.; Thébault, A.; Game, Y.; Toïgo, C.; Petit, E.; Barthe, M.-N.; Reynaud, G.; Jaÿ, M.; et al. High Shedding Potential and Significant Individual Heterogeneity in Naturally-Infected Alpine ibex (Capra ibex) With Brucella melitensis. Front. Microbiol. 2018, 9, 1065. [Google Scholar] [CrossRef] [PubMed]
- Banskar, S.; Bhute, S.S.; Suryavanshi, M.V.; Punekar, S.; Shouche, Y.S. Microbiome analysis reveals the abundance of bacterial pathogens in Rousettus leschenaultii guano. Sci. Rep. 2016, 6, 36948. [Google Scholar] [CrossRef]
- Bai, Y.; Urushadze, L.; Osikowicz, L.; McKee, C.; Kuzmin, I.; Kandaurov, A.; Babuadze, G.; Natradze, I.; Imnadze, P.; Kosoy, M. Molecular Survey of Bacterial Zoonotic Agents in Bats from the Country of Georgia (Caucasus). PLoS ONE 2017, 12, e0171175. [Google Scholar] [CrossRef]
- Shirima, G.M.; Kunda, J.S. Prevalence of brucellosis in the human, livestock and wildlife interface areas of Serengeti National Park, Tanzania. Onderstepoort J. Vet. Res. 2016, 83, a1032. [Google Scholar] [CrossRef]
- Tessaro, S.V.; Forbes, L.B.; Turcotte, C. A survey of brucellosis and tuberculosis in bison in and around Wood Buffalo National Park, Canada. Can. Vet. J. La Rev. Vet. Can. 1990, 31, 174–180. [Google Scholar]
- Kreizinger, Z.; Foster, J.T.; Rónai, Z.; Sulyok, K.M.; Wehmann, E.; Jánosi, S.; Gyuranecz, M. Genetic relatedness of Brucella suis biovar 2 isolates from hares, wild boars and domestic pigs. Vet. Microbiol. 2014, 172, 492–498. [Google Scholar] [CrossRef]
- Forbes, L.B.; Tessaro, S.V. Transmission of brucellosis from reindeer to cattle. J. Am. Vet. Med. Assoc. 1993, 203, 289–294. [Google Scholar]
- Foster, G.; Jahans, K.L.; Reid, R.J.; Ross, H.M. Isolation of Brucella species from cetaceans, seals and an otter. Vet. Rec. 1996, 138, 583–586. [Google Scholar] [CrossRef]
- Scholz, H.C.; Hubalek, Z.; Sedlacek, I.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Melzer, F.; Kampfer, P.; Neubauer, H.; Cloeckaert, A.; et al. Brucella microti sp. nov., isolated from the common vole Microtus arvalis. Int. J. Syst. Evol. Microbiol. 2008, 58, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, H.C.; Hofer, E.; Vergnaud, G.; Fleche, P.L.; Whatmore, A.M.; Dahouk, S.A.; Pfeffer, M.; Krüger, M.; Cloeckaert, A.; Tomaso, H. Isolation of Brucella microti from Mandibular Lymph Nodes of Red Foxes, Vulpes vulpes, in Lower Austria. Vector Borne Zoonotic Dis. 2009, 9, 153–156. [Google Scholar] [CrossRef]
- Kimura, M.; Une, Y.; Suzuki, M.; Park, E.-S.; Imaoka, K.; Morikawa, S. Isolation of Brucella inopinata -Like Bacteria from White’s and Denny’s Tree Frogs. Vector Borne Zoonotic Dis. 2017, 17, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Dawson, C.E.; Perrett, L.L.; Stubberfield, E.J.; Stack, J.A.; Farrelly, S.S.; Cooley, W.A.; Davison, N.J.; Quinney, S. Isolation and characterization of Brucella from the lungworms of a harbor porpoise (Phocoena phocoena). J. Wildl. Dis. 2008, 44, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenberg, T.; Riße, K.; Schauerte, N.; Geiger, C.; Blom, J.; Scholz, H.C. Isolation of a novel ‘atypical’ Brucella strain from a bluespotted ribbontail ray (Taeniura lymma). Antonie Van Leeuwenhoek 2017, 110, 221–234. [Google Scholar] [CrossRef]
- Eisenberg, T.; Schlez, K.; Fawzy, A.; Völker, I.; Hechinger, S.; Curić, M.; Schauerte, N.; Geiger, C.; Blom, J.; Scholz, H.C. Expanding the host range: Infection of a reptilian host (Furcifer pardalis) by an atypical Brucella strain. Antonie Van Leeuwenhoek 2020, 113, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Jiménez de Bagüés, M.P.; Ouahrani-Bettache, S.; Quintana Juan, F.; Mitjana, O.; Hanna, N.; Bessoles, S.; Sanchez, F.; Scholz, H.C.; Lafont, V.; Köhler, S.; et al. The New Species Brucella microti Replicates in Macrophages and Causes Death in Murine Models of Infection. J. Infect. Dis. 2010, 202, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Mora, G.; Palacios-Alfaro, J.D.; González-Barrientos, R. Wildlife reservoirs of brucellosis: Brucella in aquatic environments. Rev. Sci. Tech. 2013, 32, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.M.; Lambourn, D.M.; Jeffries, S.J.; Hall, P.B.; Rhyan, J.C.; Ewalt, D.R.; Polzin, L.M.; Cheville, N.F. Evidence of Brucella infection in Parafilaroides lungworms in a Pacific harbor seal (Phoca vitulina richardsi). J. Vet. Diagn. Investig. 1997, 9, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Perrett, L.L.; Dawson, C.E.; Davison, N.; Quinney, S. Brucella infection of lungworms from a harbour porpoise. Vet. Rec. 2004, 154, 800. [Google Scholar] [PubMed]
- Lambourn, D.M.; Garner, M.; Ewalt, D.; Raverty, S.; Sidor, I.; Jeffries, S.J.; Rhyan, J.; Gaydos, J.K. Brucella pinnipedialis infections in Pacific harbor seals (Phoca vitulina richardsi) from Washington State, USA. J. Wildl. Dis. 2013, 49, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Nymo, I.H.; Seppola, M.; Al Dahouk, S.; Bakkemo, K.R.; Jiménez de Bagüés, M.P.; Godfroid, J.; Larsen, A.K. Experimental Challenge of Atlantic Cod (Gadus morhua) with a Brucella pinnipedialis Strain from Hooded Seal (Cystophora cristata). PLoS ONE 2016, 11, e0159272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Tras, W.F.; Tayel, A.A.; Eltholth, M.M.; Guitian, J. Brucella infection in fresh water fish: Evidence for natural infection of Nile catfish, Clarias gariepinus, with Brucella melitensis. Vet. Microbiol. 2010, 141, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Tiller, R.V.; Gee, J.E.; Lonsway, D.R.; Gribble, S.; Bell, S.C.; Jennison, A.V.; Bates, J.; Coulter, C.; Hoffmaster, A.R.; De, B.K. Identification of an unusual Brucella strain (BO2) from a lung biopsy in a 52 year-old patient with chronic destructive pneumonia. BMC Microbiol. 2010, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Soler-Lloréns, P.F.; Quance, C.R.; Lawhon, S.D.; Stuber, T.P.; Edwards, J.F.; Ficht, T.A.; Robbe-Austerman, S.; O’Callaghan, D.; Keriel, A. A Brucella spp. Isolate from a Pac-Man Frog (Ceratophrys ornata) Reveals Characteristics Departing from Classical Brucellae. Front. Cell. Infect. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Rouzic, N.; Desmier, L.; Cariou, M.-E.; Gay, E.; Foster, J.T.; Williamson, C.H.D.; Schmitt, F.; Le Henaff, M.; Le Coz, A.; Lorléac’h, A.; et al. First Case of Brucellosis Caused by an Amphibian-type Brucella. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ponsart, C.; Riou, M.; Locatelli, Y.; Jacques, I.; Fadeau, A.; Jay, M.; Simon, R.; Perrot, L.; Freddi, L.; Breton, S.; et al. Brucella melitensis Rev.1 vaccination generates a higher shedding risk of the vaccine strain in Alpine ibex (Capra ibex) compared to the domestic goat (Capra hircus). Vet. Res. 2019, 50, 100. [Google Scholar] [CrossRef] [Green Version]
- Eric, J.M.; Brandon, M.S.; Paul, C.C.; Jared, D.R.; William, H.E.; Benjamin, W.; Scott, G.S.; Terry, J.K. Assessment of a Strain 19 Brucellosis Vaccination Program in Elk. Wildl. Soc. Bull. 2017, 41, 70–79. [Google Scholar] [CrossRef]
- Davis, D.S.; Elzer, P.H. Brucella vaccines in wildlife. Vet. Microbiol. 2002, 90, 533–544. [Google Scholar] [CrossRef]
- Godfroid, J.; Nielsen, K.; Saegerman, C. Diagnosis of brucellosis in livestock and wildlife. Croat. Med. J. 2010, 51, 296–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadar, M.; Shahali, Y.; Fakhri, Y.; Godfroid, J. The global epidemiology of Brucella infections in terrestrial wildlife: A meta-analysis. Transbound. Emerg. Dis. 2020, tbed.13735. [Google Scholar] [CrossRef] [PubMed]
- Batista, T.G.D.S.; Fornazari, F.; Joaquim, S.F.; Latosinski, G.S.; Teixeira, C.R.; Langoni, H. Serologic Screening for Smooth Brucella sp. in Wild Animals in Brazil. J. Wildl. Dis. 2019, 55, 721–723. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Summary of Brucella’s cellular niches and reservoirs. The endoplasmic reticulum is the preferred intracellular niche for Brucella, but in some extravillous HLA-G+ trophoblasts, B. abortus and B. suis are located in lysosomal membrane-associated protein 1 (LAMP1)- and CD63-positive acidic inclusions. Brucella replicates in macrophages, dendritic cells, monocytes, trophoblasts, bone cells (osteoclasts, osteoblasts), granulocyte progenitors, adipocytes, and infects other cells such as neutrophils, lymphocytes, and erythrocytes. Some infected cells like neutrophils mediate Brucella´s immune response modulation and/or serve as a Trojan horse to disseminate and infect new organs. Several anatomical compartments are populated by Brucella infected cells. Organs with high replication rates (placenta, epididymis, mammary glands, lymph nodes, spleen, liver, lungs, and bone marrow) correlate with clinical manifestations of the disease. Once an adaptive immune response is achieved or granulomas contain the infection, Brucella develops chronicity and persists at low replication rates. An organ reservoir would not exist without a pre-existing intracellular niche. The structures and physiological characteristics of organ reservoirs allow Brucella to start new infection cycles within natural or accidental hosts. Although Brucella detection in wild sheep, goats, frogs, fox, bats, and rodents seems almost inconsequential, bacterial loads might be maintained within the host. The zoonotic potential of wildlife reservoirs is still unknown but represents an important risk of transmission to livestock or humans (Created with BioRender.com (accessed on 15 December 2020)).
Figure 1.
Summary of Brucella’s cellular niches and reservoirs. The endoplasmic reticulum is the preferred intracellular niche for Brucella, but in some extravillous HLA-G+ trophoblasts, B. abortus and B. suis are located in lysosomal membrane-associated protein 1 (LAMP1)- and CD63-positive acidic inclusions. Brucella replicates in macrophages, dendritic cells, monocytes, trophoblasts, bone cells (osteoclasts, osteoblasts), granulocyte progenitors, adipocytes, and infects other cells such as neutrophils, lymphocytes, and erythrocytes. Some infected cells like neutrophils mediate Brucella´s immune response modulation and/or serve as a Trojan horse to disseminate and infect new organs. Several anatomical compartments are populated by Brucella infected cells. Organs with high replication rates (placenta, epididymis, mammary glands, lymph nodes, spleen, liver, lungs, and bone marrow) correlate with clinical manifestations of the disease. Once an adaptive immune response is achieved or granulomas contain the infection, Brucella develops chronicity and persists at low replication rates. An organ reservoir would not exist without a pre-existing intracellular niche. The structures and physiological characteristics of organ reservoirs allow Brucella to start new infection cycles within natural or accidental hosts. Although Brucella detection in wild sheep, goats, frogs, fox, bats, and rodents seems almost inconsequential, bacterial loads might be maintained within the host. The zoonotic potential of wildlife reservoirs is still unknown but represents an important risk of transmission to livestock or humans (Created with BioRender.com (accessed on 15 December 2020)).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
González-Espinoza, G.; Arce-Gorvel, V.; Mémet, S.; Gorvel, J.-P. Brucella: Reservoirs and Niches in Animals and Humans. Pathogens 2021, 10, 186. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020186
AMA Style
González-Espinoza G, Arce-Gorvel V, Mémet S, Gorvel J-P. Brucella: Reservoirs and Niches in Animals and Humans. Pathogens. 2021; 10(2):186. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020186
Chicago/Turabian StyleGonzález-Espinoza, Gabriela, Vilma Arce-Gorvel, Sylvie Mémet, and Jean-Pierre Gorvel. 2021. "Brucella: Reservoirs and Niches in Animals and Humans" Pathogens 10, no. 2: 186. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020186
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.