Efflux MexAB-Mediated Resistance in P. aeruginosa Isolated from Patients with Healthcare Associated Infections

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
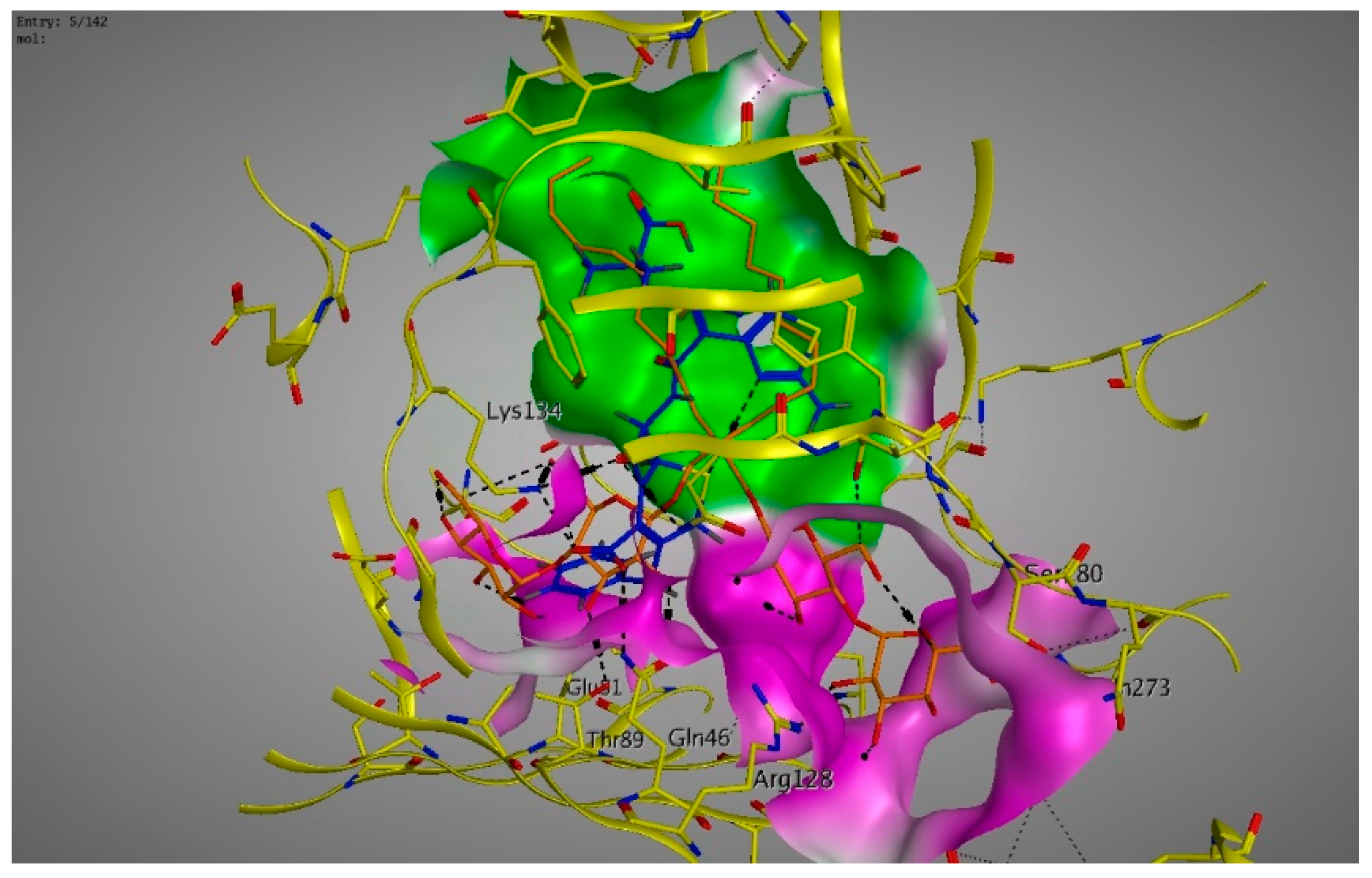
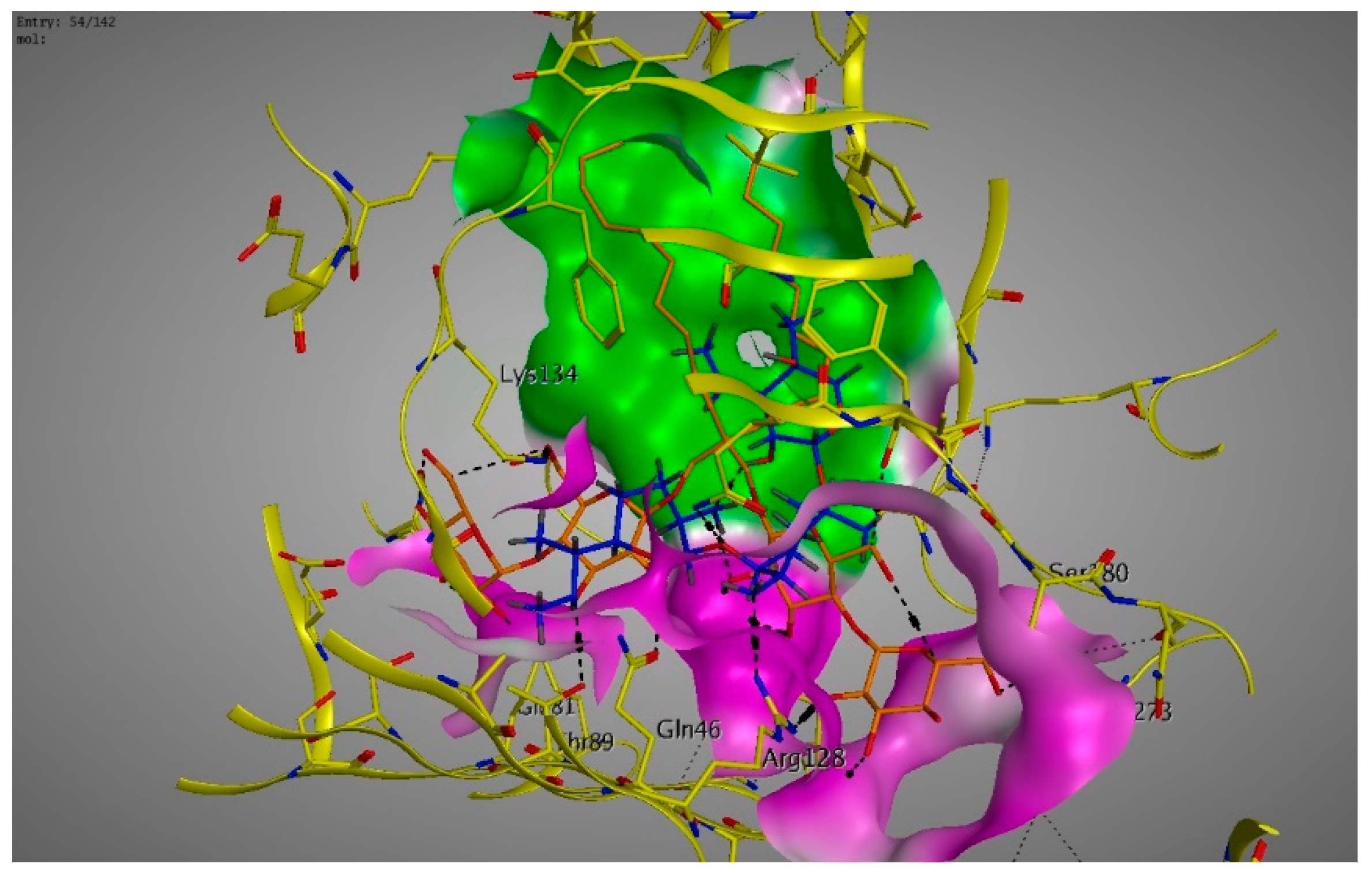
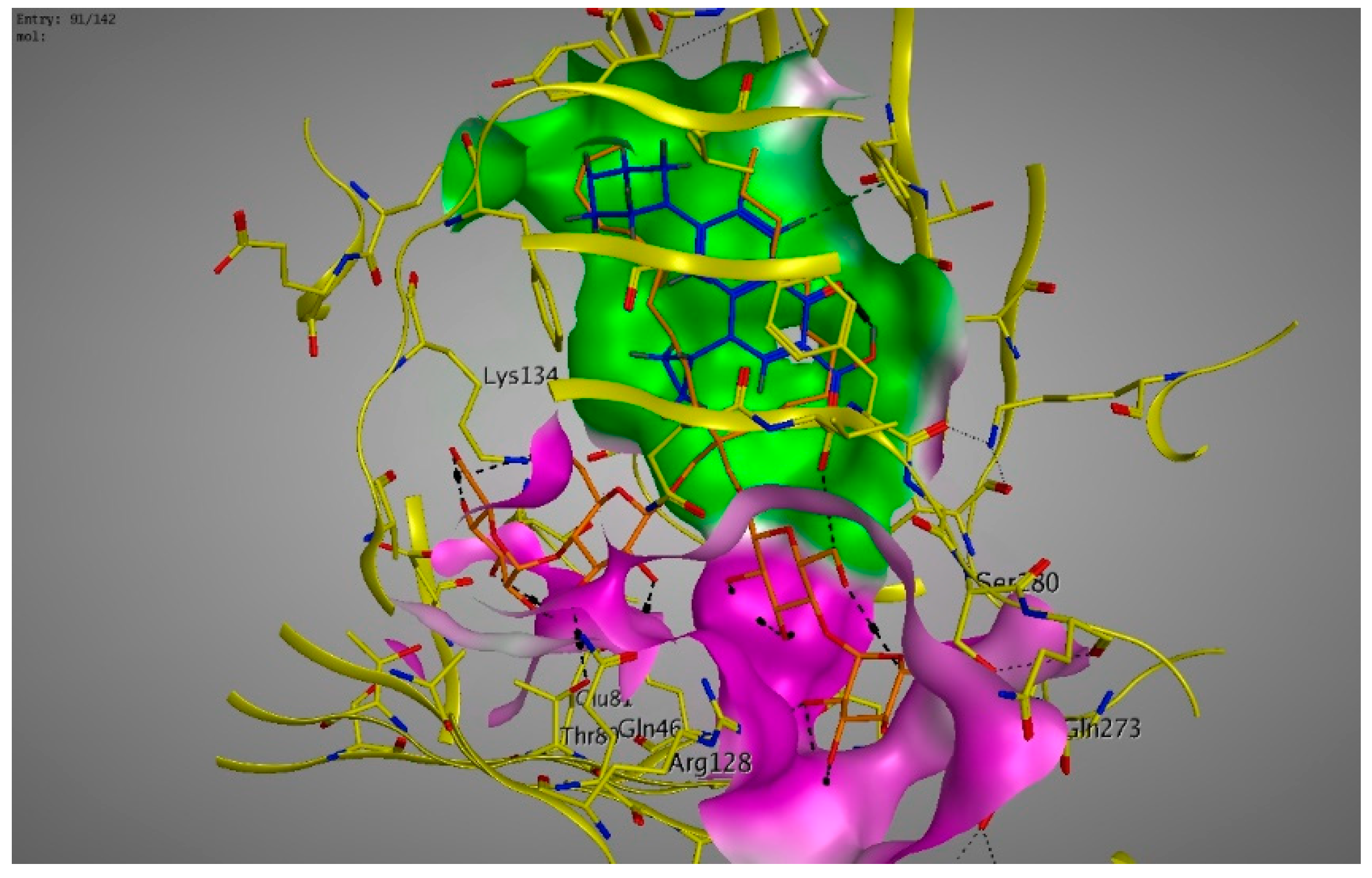
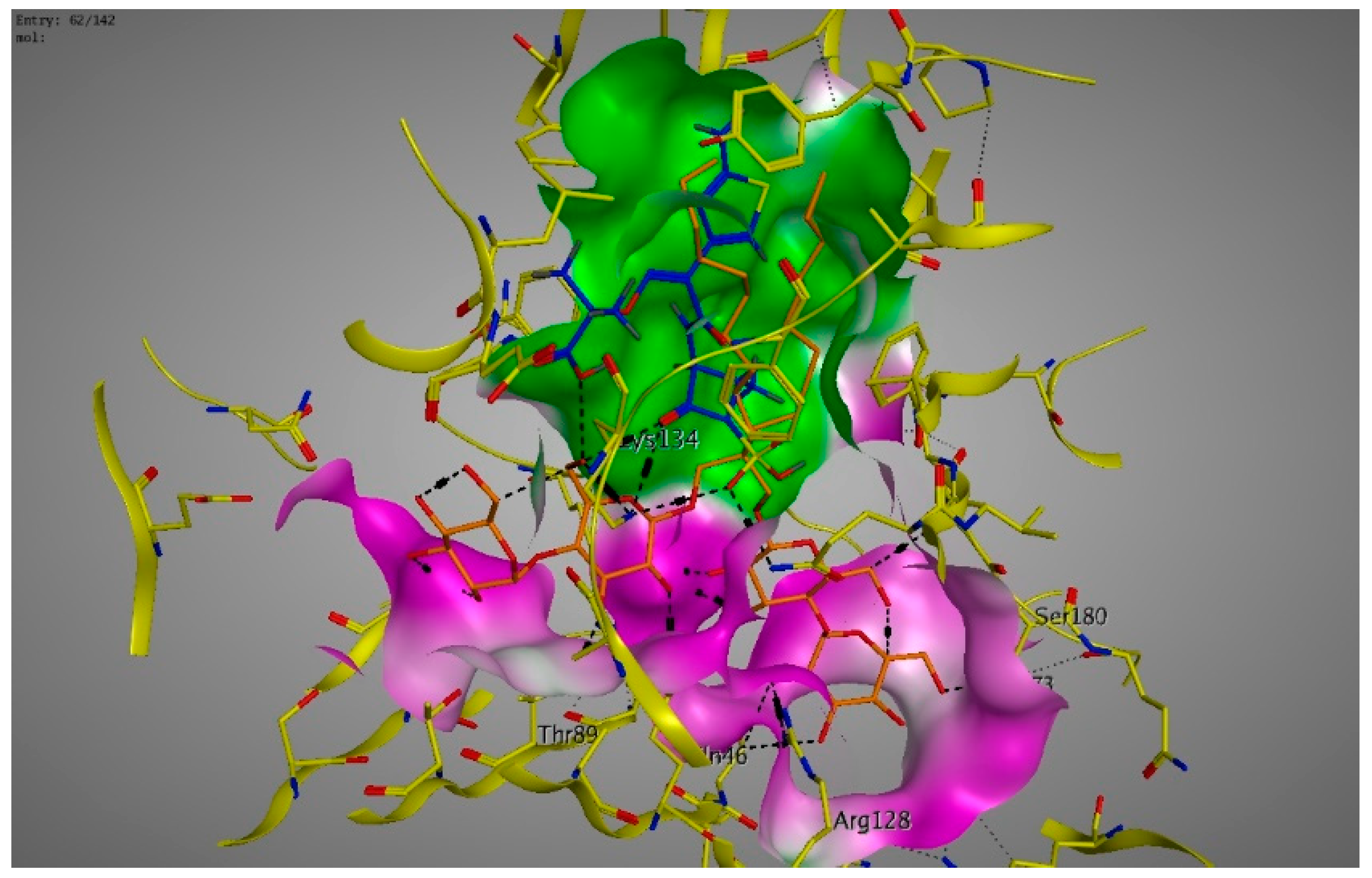
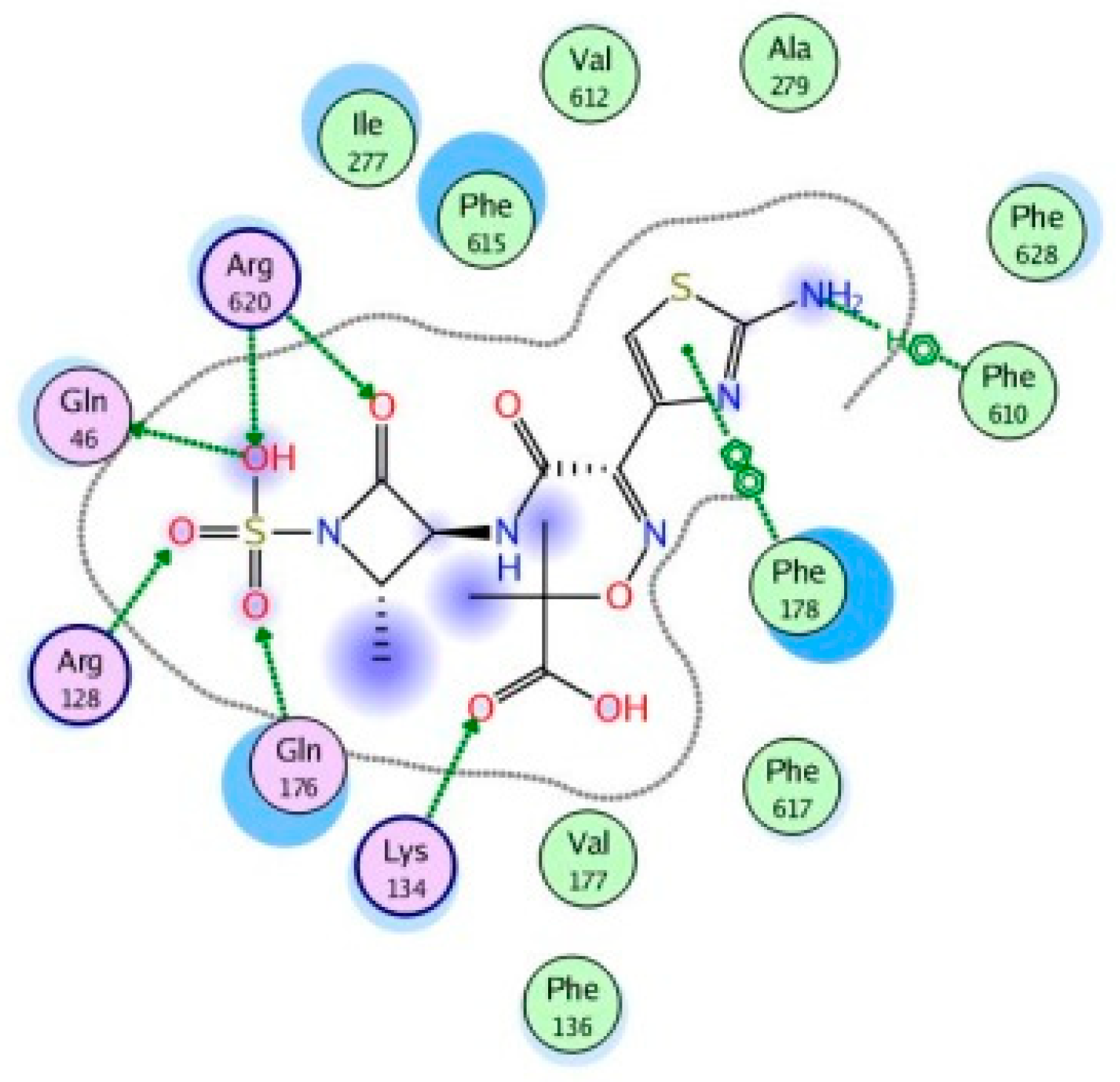
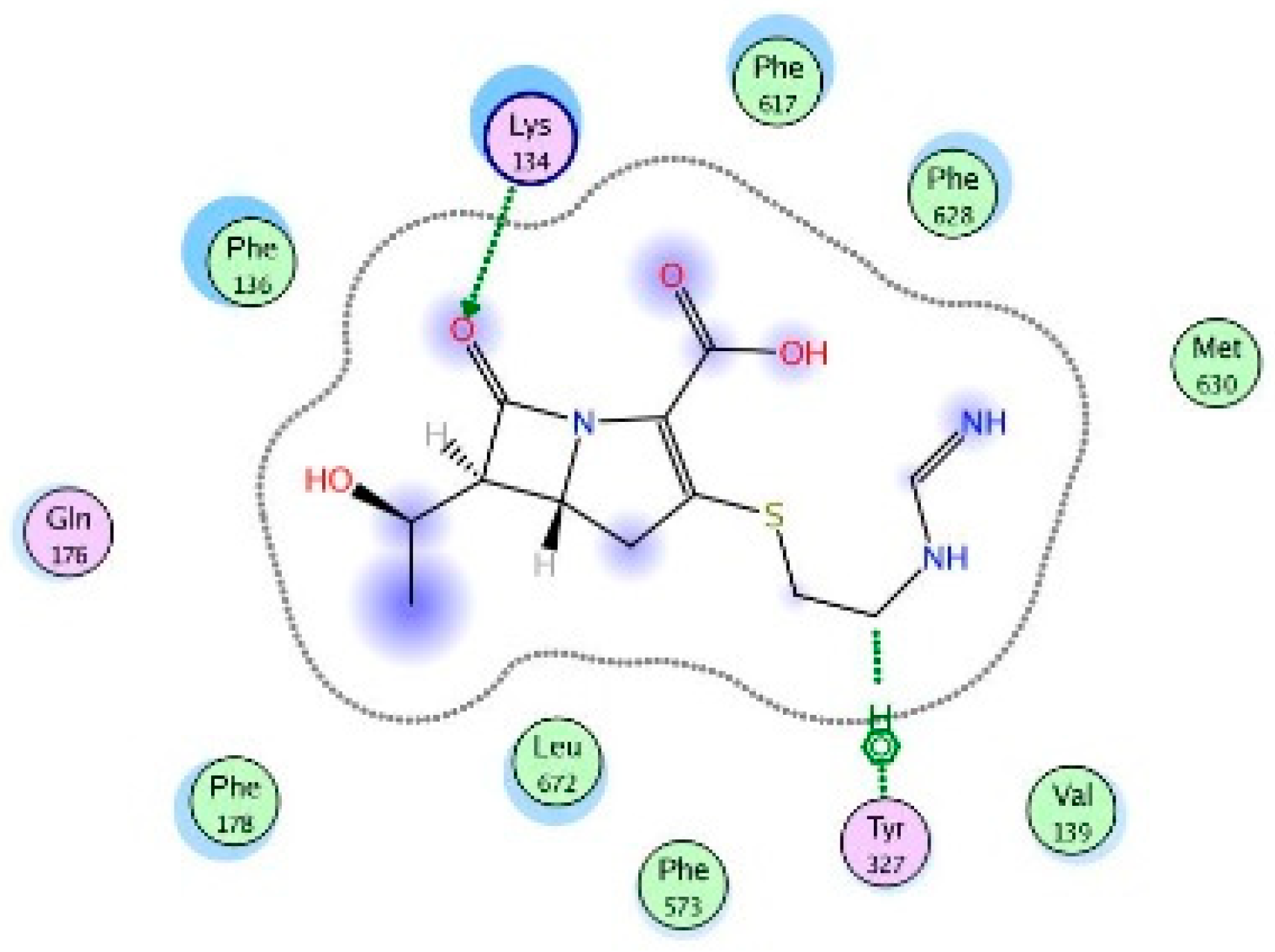
2. Results
3. Discussions
4. Materials and Methods
4.1. Bacterial Isolation and Identification
4.2. Antimicrobial Susceptibility Testing
4.3. Role of Efflux Pump in Antibiotic Resistance
4.4. Molecular Detection of mexA and mexB Genes by PCR
4.5. Statistical Analysis
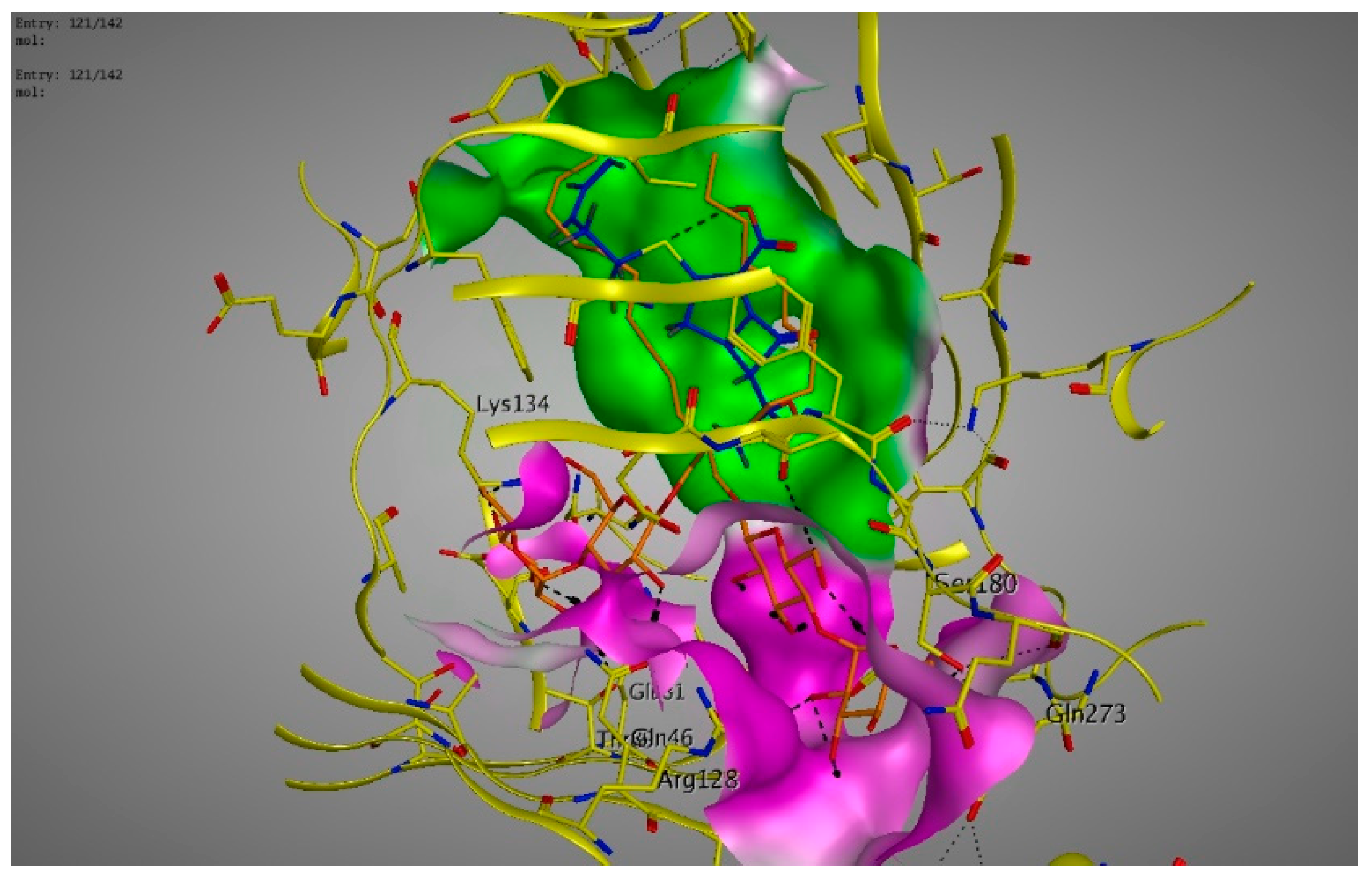
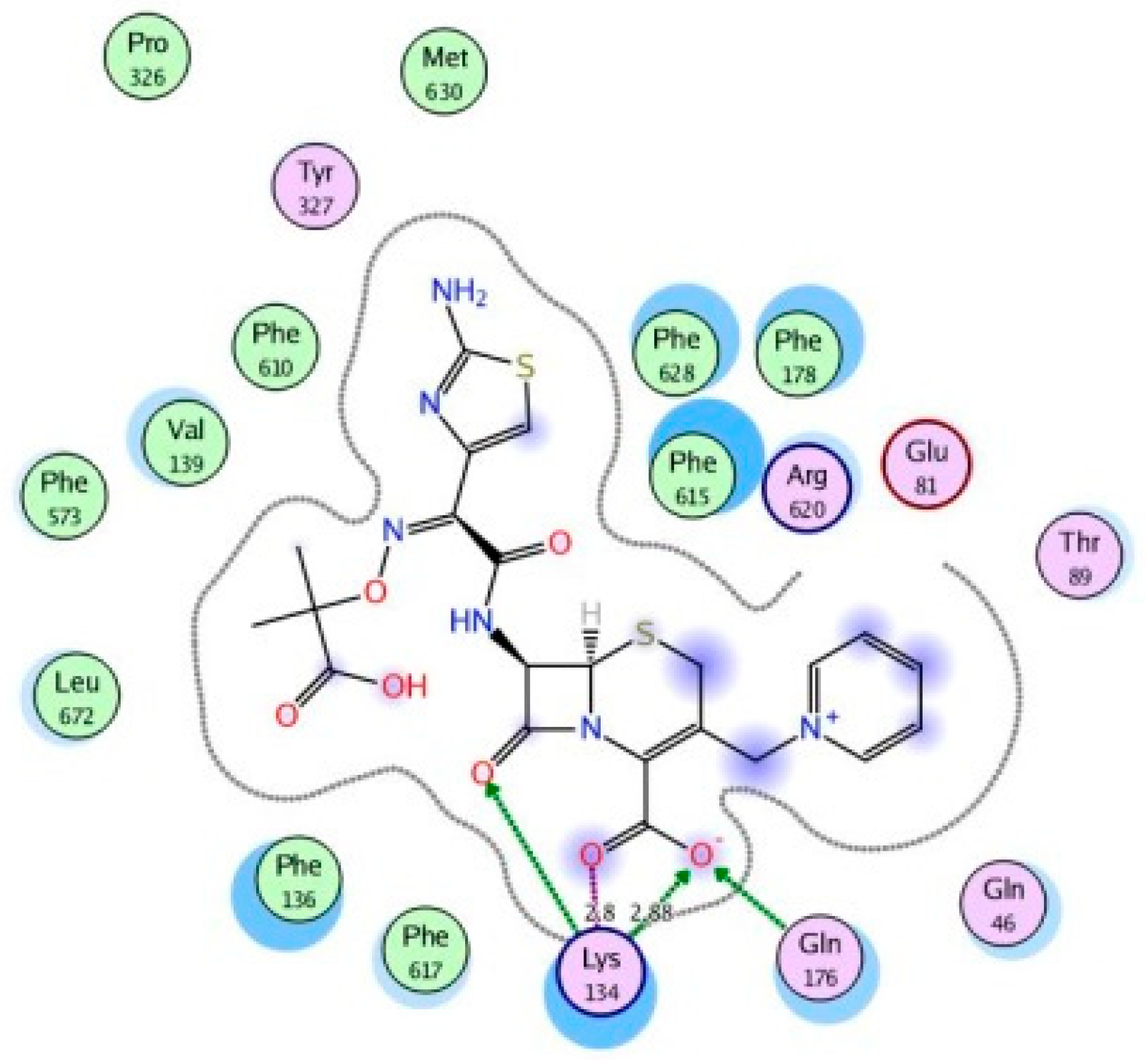
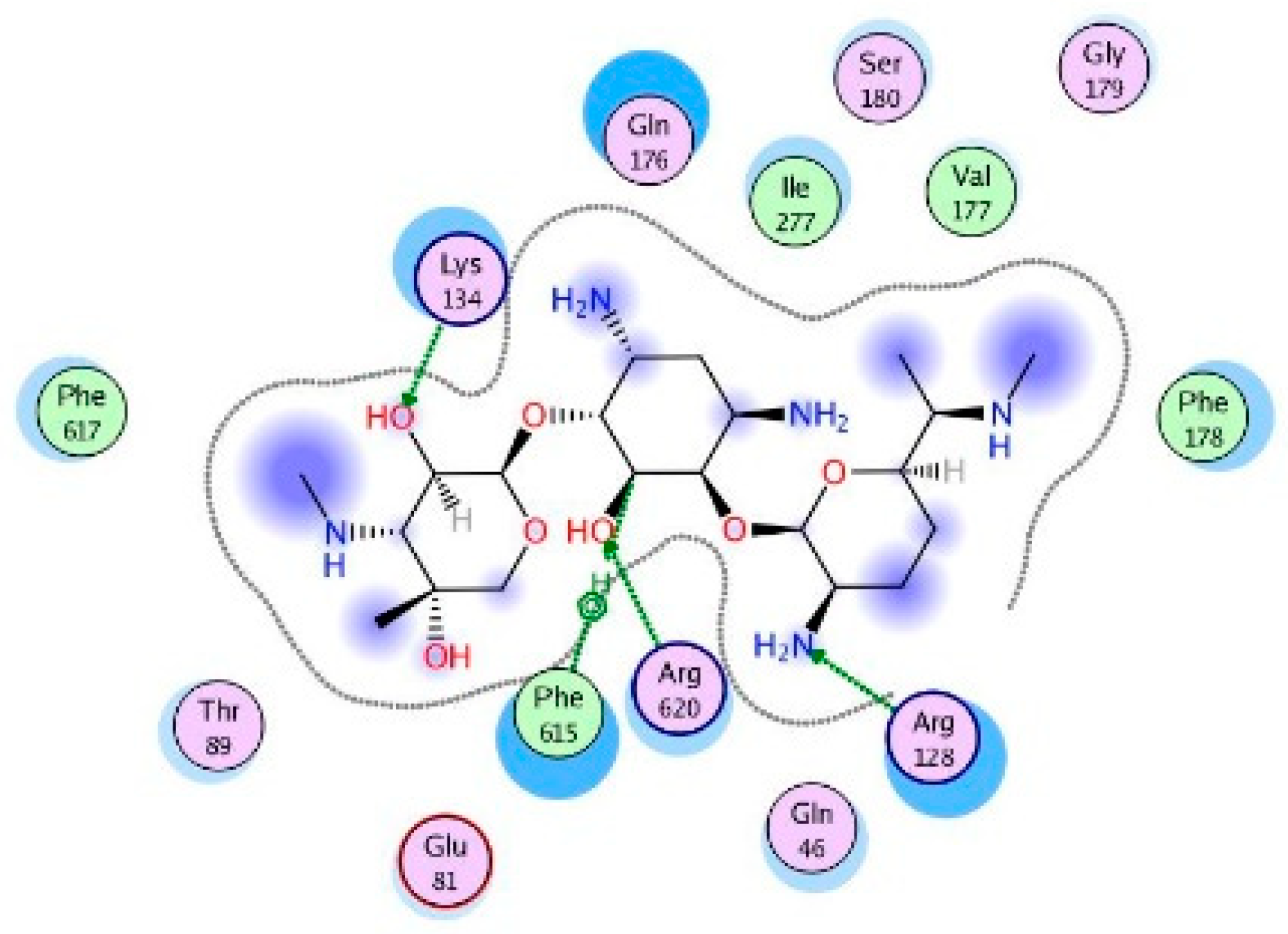
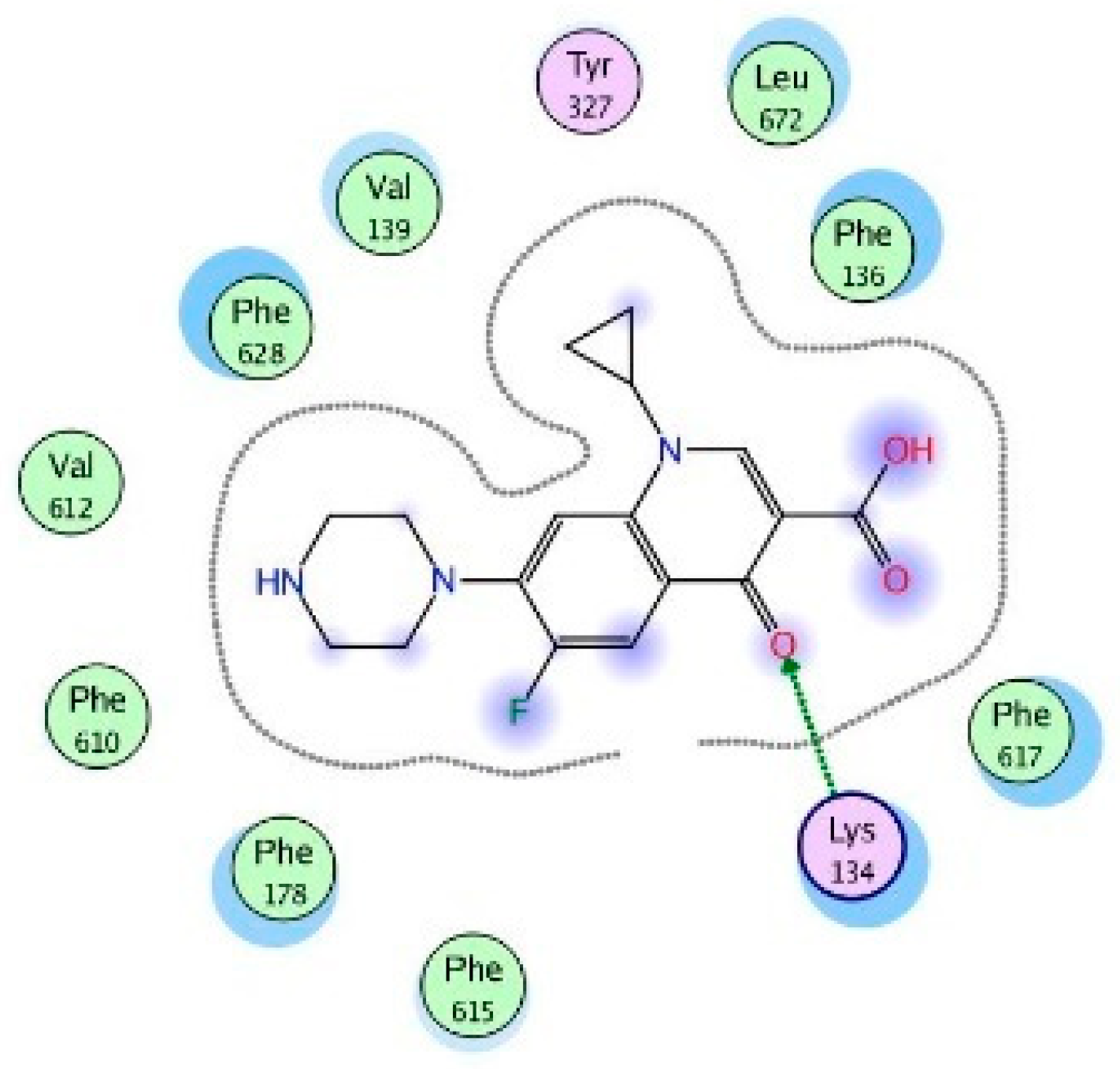
4.6. Molecular Modeling and Docking
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Defez, C.; Fabbro-Peray, P.; Bouziges, N.; Gouby, A.; Mahamat, A.; Daures, J.P. Risk factors for multidrug-resistant P. aeruginosa nosocomial infection. J. Hosp. Infect. 2004, 57, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, M.A.; Walsh, C.T. Antibiotics for emerging pathogens. Science 2009, 325, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Prevention of drug access to bacterial targets: Permeability barriers and active efflux. Science 1994, 264, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E. Resistance mechanisms in P. aeruginosa and other nonfermentative gram-negative bacteria. Clin. Infect. Dis. 1998, 27, S93–S99. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.; Martinez, J. Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Ali, H.M.; Alayafi, A.A.; Witczak, J.; Ahmad, M. Genetic variation and alleviation of salinity stress in barley. Molecules 2018, 23, 2488. [Google Scholar] [CrossRef] [Green Version]
- Falagas, M.E.; Koletsi, P.K.; Bliziotis, I.A. The diversity of definitions of multidrug-resistant (MDR) and pandrug-resistant (PDR) Acinetobacter baumannii and Pseudomonas aeruginosa. J. Med. Microbiol. 2006, 55, 1619–1629. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Efflux-mediated multiresistance in Gram-negative bacteria. Clin. Microbiol. Infect. 2004, 10, 12–26. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llanes, C.; Hocquet, D.; Vogne, C.; Benali-Baitich, D.; Neuwirth, C.; Plesiat, P. Clinical strains of P. aeruginosa overproducing MexAB-OprM and MexXY efflux pumps simultaneously. Antimicrob. Agents Chemother. 2004, 48, 1797–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneyama, H.; Maseda, H.; Kamiguchi, H.; Nakae, T. Function of the membrane fusion protein, MexA, of the MexA, B-OprM efflux pump in P. aeruginosa without an anchoring membrane. J. Biol. Chem. 2000, 275, 4628–4634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweizer, H.P. Efflux as a mechanism of resistance to antimicrobials in P. aeruginosa and related bacteria: Unanswered questions. Genet. Mol. Res. GMR 2003, 2, 48–62. [Google Scholar]
- Nikaido, H.; Pages, J.M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Efflux-mediated antimicrobial resistance. In Antibiotic Discovery and Development; Springer: Berlin/Heidelberg, Germany, 2012; pp. 349–395. [Google Scholar]
- Armstrong, E.S.; Miller, G.H. Combating evolution with intelligent design: The neoglycoside ACHN-490. Curr. Opin. Microbiol. 2010, 13, 565–573. [Google Scholar] [CrossRef]
- Tsutsui, A.; Suzuki, S.; Yamane, K.; Matsui, M.; Konda, T.; Marui, E. Genotypes and infection sites in an outbreak of multidrug-resistant Pseudomonas aeruginosa. J. Hosp. Infect. 2011, 78, 317–322. [Google Scholar] [CrossRef]
- Pagès, J.M.; Amaral, L. Mechanisms of drug efflux and strategies to combat them: Challenging the efflux pump of Gram-negative bacteria. Biochim. Biophys. Acta 2009, 1794, 826–833. [Google Scholar] [CrossRef]
- Poole, K.; Lomovskaya, O. Can efflux inhibitors really counter resistance? Infect Dis. 2006, 3, 145–152. [Google Scholar] [CrossRef]
- Mahamoud, A.; Chevalier, J.; Alibert-Franco, S.; Kern, W.V.; Pagès, J.M. Antibiotic efflux pumps in gram-negative bacteria: The inhibitor response strategy. J. Antimicrob. Chemother. 2007, 59, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Fanélus, I.; Desrosiers, R.R. Mitochondrial uncoupler carbonyl cyanide M-chlorophenylhydrazone induces the multimer assembly and activity of repair enzyme protein L-isoaspartyl methyltransferase. J. Mol. Neurosci. 2013, 50, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Kim, J.S.; Song, S.; Shigematsu, H.; Yokoyama, T.; Hyun, J.; Ha, N.C. Pseudoatomic Structure of the Tripartite Multidrug Efflux Pump AcrAB-TolC Reveals the Intermeshing Cogwheel-like Interaction between AcrA and TolC. Structure 2016, 24, 272–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakata, Y.; Srikumar, R.; Poole, K.; Gotoh, N.; Suematsu, T.; Kohno, S. Multidrug efflux systems play an important role in the invasiveness of Pseudomonas aeruginosa. J. Exp. Med. 2002, 196, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, R.; Sakurai, K.; Yamasaki, S.; Nishino, K.; Yamaguchi, A. Structures of the multidrug exporter AcrB reveal a proximal multisite drug-binding pocket. Nature 2011, 480, 565–569. [Google Scholar] [CrossRef]
- Sakurai, K.; Yamasaki, S.; Nakao, K.; Nishino, K.; Yamaguchi, A.; Nakashima, R. Crystal structures of multidrug efflux pump MexB bound with high-molecular-mass compounds. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, J.A.; Brody, S.L.; Kollef, M.H. The epidemiology, pathogenesis and treatment of P. aeruginosa infections. Drugs 2007, 67, 351–368. [Google Scholar] [CrossRef]
- Nehme, D.; Poole, K. Assembly of the MexAB-OprM multidrug pump of Pseudomonas aeruginosa: Component interactions defined by the study of pump mutant suppressors. J. Bacteriol. 2007, 189, 6118–6127. [Google Scholar] [CrossRef] [Green Version]
- Khosravi, A.D.; Mohammadian, A. Efflux MexAB-mediated resistance in multidrug and pan-drug resistant strains of P. aeruginosa isolated from patients with burn and wound infections. Jundishapur J. Nat. Pharm. Prod. 2016, 11, e25352. [Google Scholar] [CrossRef]
- Lari, A.R.; Alaghehbandan, R.; Akhlaghi, L. Burn wound infections and antimicrobial resistance in Tehran, Iran: An increasing problem. Ann. Burns Fire Disasters 2005, 18, 68. [Google Scholar]
- Pellegrino, F.L.; Teixeira, L.M.; Carvalho Md Mda, G.; Aranha Nouer, S.; Pinto De Oliveira, M.; Mello Sampaio, J.L. Occurrence of a multidrug-resistant P. aeruginosa clone in different hospitals in Rio de Janeiro, Brazil. J. Clin. Microbiol. 2002, 40, 2420–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deplano, A.; Denis, O.; Poirel, L.; Hocquet, D.; Nonhoff, C.; Byl, B. Molecular characterization of an epidemic clone of panantibiotic-resistant Pseudomonas aeruginosa. J. Clin. Microbiol. 2005, 43, 1198–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosravi, A.D.; Mihani, F. Detection of metallo-beta-lactamase-producing P. aeruginosa strains isolated from burn patients in Ahwaz, Iran. Diagn. Microbiol. Infect. Dis. 2008, 60, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Forbes, B.A.; Sahm, D.F.; Weissfeld, A. Bailey and Scott’s Diagnostic Microbiology, 12th ed.; Mosby Elsevier: St Louis, MO, USA, 2007. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Pumbwe, L.; Glass, D.; Wexler, H.M. Efflux pump overexpression in multiple-antibiotic-resistant mutants of Bacteroides fragilis. Antimicrob. Agents Chemother. 2006, 50, 3150–3153. [Google Scholar] [CrossRef] [Green Version]
- Pirnay, J.-P.; De Vos, D.; Duinslaeger, L.; Reper, P.; Vandenvelde, C.; Cornelis, P. Quantitation of P. aeruginosa in wound biopsy samples: From bacterial culture to rapid ‘real-time’polymerase chain reaction. Crit. Care 2000, 4, 255. [Google Scholar] [CrossRef] [Green Version]
- Xavier, D.E.; Picao, R.C.; Girardello, R.; Fehlberg, L.C.; Gales, A.C. Efflux pumps expression and its association with porin down-regulation and beta-lactamase production among P. aeruginosa causing bloodstream infections in Brazil. BMC Microbiol. 2010, 10, 217. [Google Scholar] [CrossRef] [Green Version]
- Molecular Operating Environment (MOE), 2019.01; Chemical Computing Group ULC: Montreal, QC, Canada, 2019.
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A.; Witczak, J.; Ahmad, M. Analysis of Genetic Variation and Enhancement of Salt Tolerance in French Pea (Pisum Sativum L.). Int. J. Mol. Sci. 2018, 19, 2433. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Amplified Product |
|---|---|---|
| 16s gene | ||
| 5′ATGGAAATGCTGAAATTCGGC 3′ | 530 bp |
| 5′CTTCTTCAGCTCGACGCGACG 3′ | |
| MexA gene | ||
| 5′CGACCAGGCCGTGAGCAAGCAGC3′ | 293 bp |
| 5′GGAGACCTTCGCCGCGTTGTCGC 3′ | |
| MexB gene | ||
| 5′GTGTTCGGCTCGCAGTACTC 3′ | 244 bp |
| 5′AACCGTCGGGATTGACCTTG 3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kishk, R.M.; Abdalla, M.O.; Hashish, A.A.; Nemr, N.A.; El Nahhas, N.; Alkahtani, S.; Abdel-Daim, M.M.; Kishk, S.M. Efflux MexAB-Mediated Resistance in P. aeruginosa Isolated from Patients with Healthcare Associated Infections. Pathogens 2020, 9, 471. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060471
Kishk RM, Abdalla MO, Hashish AA, Nemr NA, El Nahhas N, Alkahtani S, Abdel-Daim MM, Kishk SM. Efflux MexAB-Mediated Resistance in P. aeruginosa Isolated from Patients with Healthcare Associated Infections. Pathogens. 2020; 9(6):471. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060471
Chicago/Turabian StyleKishk, Rania M., Mohamed O. Abdalla, Abdullah A. Hashish, Nader A. Nemr, Nihal El Nahhas, Saad Alkahtani, Mohamed M. Abdel-Daim, and Safaa M. Kishk. 2020. "Efflux MexAB-Mediated Resistance in P. aeruginosa Isolated from Patients with Healthcare Associated Infections" Pathogens 9, no. 6: 471. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9060471