Bovine Babesiosis in Turkey: Impact, Current Gaps, and Opportunities for Intervention



 ,
,  and
and
Abstract
:1. Introduction
2. An Overview of the Cattle Industry in Turkey
3. Economic Impact of Bovine Babesiosis on the Cattle Industry in Turkey
4. Competent Tick Vectors for Babesia Parasites Identified in Turkey
5. Current Control Methods
5.1. Anti-Babesia Vaccines
5.2. Anti-Tick Control Strategies
5.3. Babesia Drug-Control Strategies
6. Diagnosis of Bovine Babesiosis
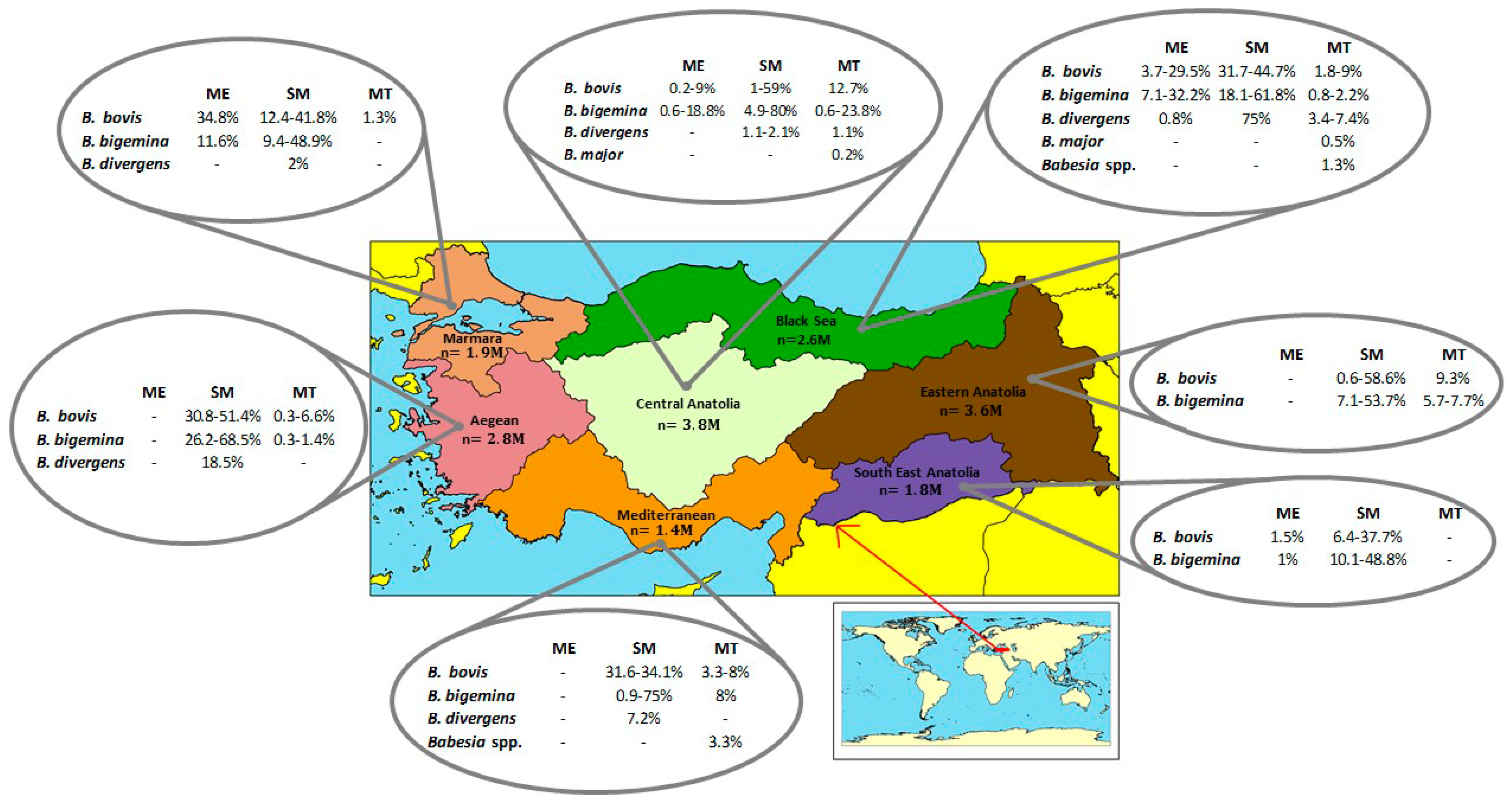
7. Current Status of Bovine Babesiosis in Turkey
8. Current Gaps on Bovine Babesiosis Research in Turkey and Suggestions for Intervention
9. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Babes, V. Sur l’hémoglobinurie bactérienne du boeuf. CR Acad. Sci. Paris 1888, 107, 692–694. [Google Scholar]
- Mihalca, A.D.C.; Suteu, E.V.; Marinculic, A.; Boireanu, P. The quest for piroplasms: From babes to smith to molecules. Sci. Parasitol. 2010, 11, 14–19. [Google Scholar]
- Smith, T.; Kilbourne, F.L. Investigations into the Nature, Causation, and Prevention of Texas or Southern Cattle Fever, 8th and 9th Reports; U.S. Department of Agriculture, Bureau of Animal Industry: Washington, DC, USA, 1893. [Google Scholar]
- Suarez, C.E.; Noh, S. Emerging perspectives in the research of bovine babesiosis and anaplasmosis. Vet. Parasitol. 2011, 180, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; Estrada-Peña, A.; Zintl, A. Vectors of Babesiosis. Annu. Rev. Èntomol. 2019, 64, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. Babesia—A historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef]
- Bock, R.; Jackson, L.; De Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.C.; Estes, D.M.; Chantler, S.E.; Kegerreis, K.A.; Suarez, C.E. DNA and a CpG Oligonucleotide Derived from Babesia bovis Are Mitogenic for Bovine B Cells. Infect. Immun. 1998, 66, 5423–5432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, W.C.; Norimine, J.; Goff, W.L.; Suarez, C.E.; McElwain, T.F. Prospects for recombinant vaccines against Babesia bovis and related parasites. Parasite Immunol. 2006, 28, 315–327. [Google Scholar] [CrossRef]
- Gallego-Lopez, G.M.; Cooke, B.M.; Suarez, C.E. Interplay between Attenuation- and Virulence-Factors of Babesia bovis and Their Contribution to the Establishment of Persistent Infections in Cattle. Pathogens 2019, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Shoda, L.K.M.; Palmer, G.H.; Florin-Christensen, J.; Florin-Christensen, M.; Godson, D.L.; Brown, W.C. Babesia bovis-Stimulated Macrophages Express Interleukin-1β, Interleukin-12, Tumor Necrosis Factor Alpha, and Nitric Oxide and Inhibit Parasite Replication In Vitro. Infect. Immun. 2000, 68, 5139–5145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goff, W.; Storset, A.K.; Johnson, W.C.; Brown, W.C. Bovine splenic NK cells synthesize IFN-gamma in response to IL-12-containing supernatants from Babesia bovis-exposed monocyte cultures. Parasite Immunol. 2006, 28, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.C.; Logan, K.S. Babesia bovis: Bovine helper T cell lines reactive with soluble and membrane antigens of merozoites. Exp. Parasitol. 1992, 74, 188–199. [Google Scholar] [CrossRef]
- Wright, I.G.; Goodger, B.V.; Leatch, G.; Aylward, J.H.; Rode-Bramanis, K.; Waltisbuhl, D.J. Protection of Babesia bigemina-immune animals against subsequent challenge with virulent Babesia bovis. Infect. Immun. 1987, 55, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R.; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Florin-Christensen, M.; Suarez, C.E.; Rodriguez, A.E.; Flores, D.A.; Schnittger, L. Vaccines against bovine babesiosis: Where we are now and possible roads ahead. Parasitology 2014, 141, 1563–1592. [Google Scholar] [CrossRef]
- Ganzinelli, S.R.; Schnittger, L.A.E.; Florin-Christensen, M. Babesia of Domestic Ruminants. In Parasitic Protozoa of Farm Animals and Pets; Florin-Christensen, M., Ed.; Springer Nature: Berlin, Germany, 2018; pp. 215–239. [Google Scholar]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a Bovine Blood Parasite of Veterinary and Zoonotic Importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.E.; Mason, T.E. Isolation and transmission of an unidentified Babesia sp. infective for cattle. Onderstepoort J. Vet. Res. 1981, 48, 155–158. [Google Scholar]
- De Waal, D.T.; Potgieter, F.T.; Combrink, M.P.; Mason, T.E. The isolation and transmission of an unidentified Babesia sp. to cattle by Hyalomma truncatum Koch 1844. Onderstepoort J. Vet. Res. 1990, 57, 229–232. [Google Scholar]
- Ohta, M.; Tsuji, M.; Tsuji, N.; Fujisaki, K. Morphological, Serological and Antigenic Characteristics, and Protein Profile of Newly Isolated Japanese Bovine Babesia Parasite with Particular Reference to Those of B. ovata. J. Vet. Med. Sci. 1995, 57, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Yin, H.; Liu, Z.; Yang, D.; Guan, G.; Liu, A.; Ma, M.; Dang, S.; Lu, B.; Sun, C.; et al. Molecular phylogenetic studies on an unnamed bovine Babesia sp. based on small subunit ribosomal RNA gene sequences. Vet. Parasitol. 2005, 133, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M. A survey of ixodid tick species and molecular identification of tick-borne pathogens. Vet. Parasitol. 2014, 200, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Inci, A.; Yildirim, A.; Düzlü, O.; Doganay, M.; Aksoy, S. Tick-Borne Diseases in Turkey: A Review Based on One Health Perspective. PLoS Neglected Trop. Dis. 2016, 10, e0005021. [Google Scholar] [CrossRef] [PubMed]
- Sevinc, F.; Xuan, X. Major tick-borne parasitic diseases of animals: A frame of references in Turkey. Eurasian J. Vet. Sci. 2015, 31, 132. [Google Scholar] [CrossRef]
- Turkstat. Turkısh Statistical Institute, I.A. Available online: http://www.turkstat.gov.tr/ (accessed on 5 May 2020).
- Pupin, R.C.; Guizelini, C.D.C.; De Lemos, R.A.A.; Martins, T.B.; Borges, F.D.A.; Borges, D.G.L.; Gomes, D.C. Retrospective study of epidemiological, clinical and pathological findings of bovine babesiosis in Mato Grosso do Sul, Brazil (1995–2017). Ticks Tick-Borne Dis. 2019, 10, 36–42. [Google Scholar] [CrossRef]
- Graham, O.H.; Hourrigan, J.L. Review Article1: Eradication Programs for the Arthropod Parasites of Livestock2. J. Med. Èntomol. 1977, 13, 629–658. [Google Scholar] [CrossRef]
- Esteve-Gasent, M.D.; Rodríguez-Vivas, R.I.; Medina, R.F.; Ellis, D.; Schwartz, A.; Garcia, B.C.; Hunt, C.; Tietjen, M.; Bonilla, D.; Thomas, D.; et al. Research on Integrated Management for Cattle Fever Ticks and Bovine Babesiosis in the United States and Mexico: Current Status and Opportunities for Binational Coordination. Pathogens 2020, 9, 871. [Google Scholar] [CrossRef]
- De León, A.A.P.; Teel, P.D.; Auclair, A.N.; Messenger, M.T.; Guerrero, F.D.; Schuster, G.; Miller, R.J. Integrated Strategy for Sustainable Cattle Fever Tick Eradication in USA Is Required to Mitigate the Impact of Global Change. Front. Physiol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- León, A.A.P.D.; Strickman, D.; Knowles, D.P.; Fish, D.; Thacker, E.L.; De La Fuente, J.; Krause, P.J.; Wikel, S.K.; Miller, R.S.; Wagner, G.G.; et al. One Health approach to identify research needs in bovine and human babesioses: Workshop report. Parasites Vectors 2010, 3. [Google Scholar] [CrossRef] [Green Version]
- Ekici, O.D.; Sevinc, F. Seroepidemiology of Babesia bigemina in Cattle in the Konya Province, Turkey: Endemic Status. B. Vet. I. Pulawy. 2009, 53, 645–649. [Google Scholar]
- Inci, A.; Ica, A.; Yildirim, A.; Vatansever, Z.; Cakmak, A.; Albasan, H.; Cam, Y.; Atasever, A.; Sariozkan, S.; Duzlu, O. Economical impact of tropical theileriosis in the Cappadocia region of Turkey. Parasitol. Res. 2007, 101, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.E.S.; Tomazic, M.L.L.; Florin-Christensen, M. Current and Prospective Tools for the Control of Cattle-Infecting Babesia Parasites. In Protozoa: Biology, Classification and Role in Disease; Castillo, V., Harris, R., Eds.; Nova Publishers: Hauppauge, NY, USA, 2013; pp. 1–44. [Google Scholar]
- Hadani, A.P.E.; Tsafrir, N.; Rauchbach, K.; Mayer, E. The transmission of Babesia bigemina and Babesiella berbera and Anaplasma centrale by Boophilus annulatus (Say). Refuah Vet. 1974, 31, 149–154. [Google Scholar]
- Mahoney, D.; Mirre, G. A note on the transmission of Babesia bovis (syn B. argentina) by the one-host tick, Boophilus microplus. Res. Vet. Sci. 1979, 26, 253–254. [Google Scholar] [CrossRef]
- Smith, R.; Osorno, B.; Brener, J.; De La Rosa, R.; Ristic, M. Bovine babesiosis: Severity and reproducibility of Babesia bovis infections induced by Boophilus microplus under laboratory conditions. Res. Vet. Sci. 1978, 24, 287–292. [Google Scholar] [CrossRef]
- Callow, L.L. The infection of Boophilus microplus with Babesia bigemina. Parasitology 1968, 58, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Morzaria, S.P.; Young, A.S.; Hudson, E.B. Babesia bigemina in Kenya: Experimental transmission by Boophilus decoloratus and the production of tick-dervied stabilates. Parasitology 1977, 74, 291–298. [Google Scholar] [CrossRef]
- Büscher, G. The infection of various tick species with Babesia bigemina, its transmission and identification. Parasitol. Res. 1988, 74, 324–330. [Google Scholar] [CrossRef]
- Donnelly, J.; Peirce, M. Experiments on the transmission of Babesia divergens to cattle by the tick Ixodes ricinus. Int. J. Parasitol. 1975, 5, 363–367. [Google Scholar] [CrossRef]
- Gray, J.S.; De Vos, A.J. Studies on a bovine Babesia transmitted by Hyalomma marginatum rufipes Koch, 1844. Onderstepoort J. Vet. Res. 1981, 48, 215–223. [Google Scholar]
- Ma, L.H.L.; Zhao, J.L. An investigation of water buffalo babesiosis in Hubei province V. adult Rhipicephalus haemaphysaloides transmits the parasites transovarially. Chin. J. Anim. Vet. Sci. 1989, 1, 67–70. [Google Scholar]
- Takahashi, K.W.; Kawai, S.A.; Yokota, H.; Kurosawa, T.; Sonoda, M. Investigation of isolation, transmission and virulence of bovine Babesia sp. in Hokkaido. J. Coll. Dairying 1983, 10, 171–181. [Google Scholar]
- Aydin, L.; Bakirci, S. Geographical distribution of ticks in Turkey. Parasitol. Res. 2007, 101, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Ozubek, S.; Dumanlı, N. A survey of ixodid ticks feeding on cattle and prevalence of tick-borne pathogens in the Black Sea region of Turkey. Vet. Parasitol. 2012, 187, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Ica, A.; Vatansever, Z.; Yildirim, A.; Duzlu, O.; Inci, A. Detection of Theileria and Babesia species in ticks collected from cattle. Vet. Parasitol. 2007, 148, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Meyilli, T.D.; Yildirim, A.O.; Onder, Z.; Ciloglu, A.; Inci, A. Investigation of Babesia bovis and Babesia bigemina by real time pcr in tick species collected from cattle in Kayseri province. Erciyes Üniv. Vet. Fak Derg 2016, 13, 201–208. [Google Scholar]
- Aktas, M.; Vatansever, Z.; Ozubek, S. Molecular evidence for trans-stadial and transovarial transmission of Babesia occultans in Hyalomma marginatum and Rhipicephalus turanicus in Turkey. Vet. Parasitol. 2014, 204, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Orkun, Ö.; Çakmak, A.; Nalbantoğlu, S.; Karaer, Z. Turkey tick news: A molecular investigation into the presence of tick-borne pathogens in host-seeking ticks in Anatolia; Initial evidence of putative vectors and pathogens, and footsteps of a secretly rising vector tick, Haemaphysalis parva. Ticks Tick-Borne Dis. 2020, 11. [Google Scholar] [CrossRef]
- Callow, L.; Mellors, L.; McGregor, W. Reduction in virulence of Babesia bovis due to rapid passage in splenectomized cattle. Int. J. Parasitol. 1979, 9, 333–338. [Google Scholar] [CrossRef]
- Dalgliesh, R.J.; Callow, L.L.; Mellors, L.T.; McGregor, W. Development of a highly infective babesia bigemina vaccine of reduced virulence. Aust. Vet. J. 1981, 57, 8–11. [Google Scholar] [CrossRef]
- Pedroni, M.J.; Sondgeroth, K.S.; Gallego-Lopez, G.M.; Echaide, I.; Lau, A.O.T. Comparative transcriptome analysis of geographically distinct virulent and attenuated Babesia bovis strains reveals similar gene expression changes through attenuation. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Lopez, G.M.; Lau, A.O.T.; Brown, W.C.; Johnson, W.C.; Ueti, M.W.; Suarez, C.E. Spherical Body Protein 2 truncated copy 11 as a specific Babesia bovis attenuation marker. Parasites Vectors 2018, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Späth, E.; Mangold, A.J.; Guglielmone, A. Estimation of the potential demand for a bovine babesiosis and anaplasmosis vaccine in argentina. Vet. Parasitol. 1990, 36, 131–140. [Google Scholar] [CrossRef]
- Ojeda, J.J.; Orozco, L.; Flores, R.; Rojas, C.; Figueroa, J.V.; Martínez, J.A.A. Validation of an Attenuated Live Vaccine against Babesiosis in Native Cattle in an Endemic Area. Transbound. Emerg. Dis. 2010, 57, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Alzan, H.F.; Cooke, B.M.; Suarez, C.E. Transgenic Babesia bovis lacking 6-Cys sexual-stage genes as the foundation for non-transmissible live vaccines against bovine babesiosis. Ticks Tick-Borne Dis. 2019, 10, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Rathinasamy, V.; Poole, W.A.; Bastos, R.G.; Suarez, C.E.; Cooke, B.M. Babesiosis Vaccines: Lessons Learned, Challenges Ahead, and Future Glimpses. Trends Parasitol. 2019, 35, 622–635. [Google Scholar] [CrossRef] [PubMed]
- Sibinovic, K.H.; Sibinovic, S.; Ristic, M.; Cox, H.W. Immunogenic Properties of Babesial Serum Antigens. J. Parasitol. 1967, 53, 1121. [Google Scholar] [CrossRef]
- Montenegro-James, S.; Meuwissen, J. Immunoprophylactic control of bovine babesiosis: Role of exoantigens of Babesia. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 85–94. [Google Scholar] [CrossRef]
- Moreau, Y.; Vidor, E.; Bissuel, G.; Dubreuil, N. Vaccination against canine babesiosis: An overview of field observations. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 95–96. [Google Scholar] [CrossRef]
- Valentin, A.; Precigout, E.; L’Hostis, M.; Carcy, B.; Gorenflot, A.; Schrevel, J. Cellular and humoral immune responses induced in cattle by vaccination with Babesia divergens culture-derived exoantigens correlate with protection. Infect. Immun. 1993, 61, 734–741. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Liu, Z.; Yao, B.; Ma, L. Culture-derived Babesia orientalis exoantigens used as a vaccine against buffalo babesiosis. Parasitol. Res. 2002, 88, S38–S40. [Google Scholar] [CrossRef]
- Bastos, R.G.; Suarez, C.E.; Laughery, J.M.; Johnson, W.C.; Ueti, M.W.; Knowles, D.P. Differential Expression of Three Members of the Multidomain Adhesion CCp Family in Babesia bigemina, Babesia bovis and Theileria equi. PLoS ONE 2013, 8, e67765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, H.E.; Bastos, R.G.; Schneider, D.A.; Johnson, W.C.; Adham, F.K.; Davis, W.C.; Laughery, J.M.; Herndon, D.R.; Alzan, H.F.; Ueti, M.W.; et al. The Babesia bovis hap2 gene is not required for blood stage replication, but expressed upon in vitro sexual stage induction. PLoS Neglected Trop. Dis. 2017, 11, e0005965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Ellis, R.D.; Shaffer, D.; Fontes, E.; Malkin, E.M.; Mahanty, S.; Fay, M.P.; Narum, D.L.; Rausch, K.; Miles, A.P.; et al. Phase 1 Trial of Malaria Transmission Blocking Vaccine Candidates Pfs25 and Pvs25 Formulated with Montanide ISA 51. PLoS ONE 2008, 3, e2636. [Google Scholar] [CrossRef] [PubMed]
- Alzan, H.F.; Lau, A.O.T.; Knowles, D.P.; Herndon, D.R.; Ueti, M.W.; Scoles, G.A.; Kappmeyer, L.S.; Suarez, C.E. Expression of 6-Cys Gene Superfamily Defines Babesia bovis Sexual Stage Development within Rhipicephalus microplus. PLoS ONE 2016, 11, e0163791. [Google Scholar] [CrossRef]
- Bohaliga, G.A.R.; Johnson, W.C.; Taus, N.S.; Hussein, H.E.; Bastos, R.G.; Suarez, C.E.; Scoles, G.A.; Ueti, M.W. Identification of proteins expressed by Babesia bigemina kinetes. Parasites Vectors 2019, 12, 271. [Google Scholar] [CrossRef]
- Camacho-Nuez, M.; Hernández-Silva, D.J.; Castañeda-Ortiz, E.J.; Paredes-Martínez, M.E.; Rocha-Martínez, M.K.; Álvarez-Sánchez, M.E.; Mercado-Curiel, R.F.; Aguilar-Tipacamú, G.; Gualito, J.M. Hap2, a novel gene in Babesia bigemina is expressed in tick stages, and specific antibodies block zygote formation. Parasites Vectors 2017, 10, 568. [Google Scholar] [CrossRef] [Green Version]
- Fish, L.; Leibovich, B.; Krigel, Y.; McElwain, T.; Shkap, V. Vaccination of cattle against B. bovis infection with live attenuated parasites and non-viable immunogens. Vaccine 2008, 26, G29–G33. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Lopez, U.; Rojas, C.; Borgonio, V.M.; Sanchez, V.; Castañeda, R.; Vargas, P.; Figueroa-Millán, J.V. Immunization of Bos taurus Steers with Babesia bovis Recombinant Antigens MSA-1, MSA-2c and 12D3. Transbound. Emerg. Dis. 2010, 57, 87–90. [Google Scholar] [CrossRef]
- Gaffar, F.R.; Yatsuda, A.P.; Franssen, F.F.; De Vries, E. A Babesia bovis merozoite protein with a domain architecture highly similar to the thrombospondin-related anonymous protein (TRAP) present in Plasmodium sporozoites. Mol. Biochem. Parasitol. 2004, 136, 25–34. [Google Scholar] [CrossRef]
- Gaffar, F.R.; Yatsuda, A.P.; Franssen, F.F.J.; De Vries, E. Erythrocyte Invasion by Babesia bovis Merozoites Is Inhibited by Polyclonal Antisera Directed against Peptides Derived from a Homologue of Plasmodium falciparum Apical Membrane Antigen 1. Infect. Immun. 2004, 72, 2947–2955. [Google Scholar] [CrossRef] [Green Version]
- Torina, A.; Agnone, A.; Sireci, G.; Mosqueda, J.J.; Blanda, V.; Albanese, I.; La Farina, M.; Cerrone, A.; Cusumano, F.; Caracappa, S. Characterization of the Apical Membrane Antigen-1 in Italian Strains of Babesia bigemina. Transbound. Emerg. Dis. 2010, 57, 52–56. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, J.; Kopáček, P.; Lew-Tabor, A.; Maritz-Olivier, C. Strategies for new and improved vaccines against ticks and tick-borne diseases. Parasite Immunol. 2016, 38, 754–769. [Google Scholar] [CrossRef] [PubMed]
- Karunaratne, S.H.P.P. Insecticide resistance in insects: A review. Cey. J. Sci. (Bio. Sci.) 1998, 25, 72–99. [Google Scholar]
- De La Fuente, J. Controlling ticks and tick-borne diseases looking forward. Ticks Tick-Borne Dis. 2018, 9, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, F.D.; Miller, R.J.; De León, A.A.P. Cattle tick vaccines: Many candidate antigens, but will a commercially viable product emerge? Int. J. Parasitol. 2012, 42, 421–427. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, J.; Rodríguez, M.; Montero, C.V.; Redondo, M.L.; García-García, J.C.; Méndez, L.; Serrano, E.; Valdes, M.; Enríquez, A.; Canales, M.; et al. Vaccination against ticks (Boophilus spp.): The experience with the Bm86-based vaccine Gavac™. Genet. Anal. Biomol. Eng. 1999, 15, 143–148. [Google Scholar] [CrossRef]
- Suarez, M.; Rubi, J.; Perez, D.; Cordova, V.; Salazar, Y.; Vielma, A.; Barrios, F.; Gil, C.A.; Segura, N.; Carrillo, Y.; et al. High impact and effectiveness of Gavac™ vaccine in the national program for control of bovine ticks Rhipicephalus microplus in Venezuela. Livest. Sci. 2016, 187, 48–52. [Google Scholar] [CrossRef]
- De La Fuente, J.; Almazán, C.; Canales, M.; De La Lastra, J.M.P.; Kocan, K.M.; Willadsen, P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim. Heal. Res. Rev. 2007, 8, 23–28. [Google Scholar] [CrossRef]
- De La Fuente, J. Vaccines for vector control: Exciting possibilities for the future. Vet. J. 2012, 194, 139–140. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, J.; Merino, O. Vaccinomics, the new road to tick vaccines. Vaccine 2013, 31, 5923–5929. [Google Scholar] [CrossRef]
- Merino, O.; Antunes, S.; Gualito, J.M.; Moreno-Cid, J.A.; De La Lastra, J.M.P.; Rosario-Cruz, R.; Rodríguez, S.; Domingos, A.; De La Fuente, J.; Rosario-Cruz, R. Vaccination with proteins involved in tick–pathogen interactions reduces vector infestations and pathogen infection. Vaccine 2013, 31, 5889–5896. [Google Scholar] [CrossRef] [PubMed]
- Merino, O.; Almazán, C.; Canales, M.; Villar, M.; Moreno-Cid, J.A.; Galindo, R.C.; De La Fuente, J. Targeting the tick protective antigen subolesin reduces vector infestations and pathogen infection by Anaplasma marginale and Babesia bigemina. Vaccine 2011, 29, 8575–8579. [Google Scholar] [CrossRef] [PubMed]
- Gualito, J.M.; Olvera-Ramirez, A.; Aguilar-Tipacamu, G.; Canto, G.J. Current Advances in Detection and Treatment of Babesiosis. Curr. Med. Chem. 2012, 19, 1504–1518. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.R.; Bruno, P.M.; Gilbert, L.A.; Capron, K.L.; Lauffenburger, D.A.; Hemann, M.T. Defining principles of combination drug mechanisms of action. Proc. Natl. Acad. Sci. USA 2013, 110, E170–E179. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.-C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; Terkawi, M.A.; Youssef, M.A.; Said, E.S.E.S.E.; Elsayed, G.; El-Khodery, S.; El-Ashker, M.; Elsify, A.; Omar, M.; et al. Optimization of a Fluorescence-Based Assay for Large-Scale Drug Screening against Babesia and Theileria Parasites. PLoS ONE 2015, 10, e0125276. [Google Scholar] [CrossRef]
- Silva, M.G.; Domingos, A.; Esteves, M.A.; Cruz, M.; Suarez, C.E. Evaluation of the growth-inhibitory effect of trifluralin analogues on in vitro cultured Babesia bovis parasites. Int. J. Parasitol. Drugs Drug Resist. 2013, 3, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.G.; Villarino, N.F.; Knowles, D.P.; Suarez, C.E. Assessment of Draxxin® (tulathromycin) as an inhibitor of in vitro growth of Babesia bovis, Babesia bigemina and Theileria equi. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 265–270. [Google Scholar] [CrossRef]
- Tayebwa, D.S.; Tuvshintulga, B.; Guswanto, A.; Nugraha, A.B.; Batiha, G.E.-S.; Gantuya, S.; Rizk, M.A.; Vudriko, P.; Sivakumar, T.; Yokoyama, N.; et al. The effects of nitidine chloride and camptothecin on the growth of Babesia and Theileria parasites. Ticks Tick-Borne Dis. 2018, 9, 1192–1201. [Google Scholar] [CrossRef]
- Tuvshintulga, B.; Aboulaila, M.; Davaasuren, B.; Ishiyama, A.; Sivakumar, T.; Yokoyama, N.; Iwatsuki, M.; Otoguro, K.; Ōmura, S.; Igarashi, I. Clofazimine Inhibits the Growth of Babesia and Theileria ParasitesIn VitroandIn Vivo. Antimicrob. Agents Chemother. 2016, 60, 2739–2746. [Google Scholar] [CrossRef] [Green Version]
- Tuvshintulga, B.; Aboulaila, M.; Sivakumar, T.; Tayebwa, D.S.; Gantuya, S.; Naranbaatar, K.; Ishiyama, A.; Iwatsuki, M.; Otoguro, K.; Ōmura, S.; et al. Chemotherapeutic efficacies of a clofazimine and diminazene aceturate combination against piroplasm parasites and their AT-rich DNA-binding activity on Babesia bovis. Sci. Rep. 2017, 7, 13888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guswanto, A.; Sivakumar, T.; Rizk, M.A.; Elsayed, S.A.E.; Youssef, M.A.; Elsaid, E.E.S.; Yokoyama, N.; Igarashi, I. Evaluation of a Fluorescence-Based Method for Antibabesial Drug Screening. Antimicrob. Agents Chemother. 2014, 58, 4713–4717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalovecká, M.; Hartmann, D.; Miyamoto, Y.; Eckmann, L.; Hajdusek, O.; O’Donoghue, A.J.; Sojka, D. Validation of Babesia proteasome as a drug target. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 394–402. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, S.A.E.-S.; Rizk, M.A.; Yokoyama, N.; Igarashi, I. Evaluation of the in vitro and in vivo inhibitory effect of thymoquinone on piroplasm parasites. Parasites Vectors 2019, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; Nassif, M.; Gualito, J.M.; Xuan, X.; Igarashi, I. Assay methods for in vitro and in vivo anti-Babesia drug efficacy testing: Current progress, outlook, and challenges. Vet. Parasitol. 2020, 279. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Ozubek, S. Molecular and Parasitological Survey of Bovine Piroplasms in the Black Sea Region, Including the First Report of Babesiosis Associated with Babesia divergensin Turkey. J. Med. Èntomol. 2015, 52, 1344–1350. [Google Scholar] [CrossRef]
- Wang, G.; Wormser, G.P.; Zhuge, J.; Villafuerte, P.; Ip, D.; Zeren, C.; Fallon, J.T. Utilization of a real-time PCR assay for diagnosis of Babesia microti infection in clinical practice. Ticks Tick-Borne Dis. 2015, 6, 376–382. [Google Scholar] [CrossRef]
- Ozubek, S.; Aktas, M. Molecular evidence of a new Babesia sp. in goats. Vet. Parasitol. 2017, 233, 1–8. [Google Scholar] [CrossRef]
- Roy, B.C.; Krücken, J.; Ahmed, J.S.; Majumder, S.; Baumann, M.P.; Clausen, P.-H.; Nijhof, A.M. Molecular identification of tick-borne pathogens infecting cattle in Mymensingh district of Bangladesh reveals emerging species of Anaplasma and Babesia. Transbound. Emerg. Dis. 2017, 65, e231–e242. [Google Scholar] [CrossRef]
- Kim, C.-M.; Yokoyama, N.; Blanco, L.B.C.; Iseki, H.; Igarashi, I.; Xuan, X.; Alhassan, A. Development of a rapid immunochromatographic test for simultaneous serodiagnosis of bovine babesioses caused by Babesia bovis and Babesia bigemina. Am. J. Trop. Med. Hyg. 2008, 78, 117–121. [Google Scholar] [CrossRef]
- Guswanto, A.; Allamanda, P.; Mariamah, E.S.; Munkjargal, T.; Tuvshintulga, B.; Takemae, H.; Sivakumar, T.; Aboulaila, M.; Terkawi, M.A.; Ichikawa-Seki, M.; et al. Evaluation of immunochromatographic test (ICT) strips for the serological detection of Babesia bovis and Babesia bigemina infection in cattle from Western Java, Indonesia. Vet. Parasitol. 2017, 239, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Tayebwa, D.S.; Beshbishy, A.M.; Batiha, G.E.-S.; Komugisha, M.; Byaruhanga, J.; Vudriko, P.; Yahia, R.; Alkazmi, L.; Hetta, H.F.; Yokoyama, N.; et al. Assessing the Immunochromatographic Test Strip for Serological Detection of Bovine Babesiosis in Uganda. Microorganisms 2020, 8, 1110. [Google Scholar] [CrossRef]
- Curasson, D.A. Famille des Piroplasmides. In Traite de Parasitologie Veterinare et Comparee; Freres, V., Ed.; Tome III: Paris, France, 1943; pp. 160–190. [Google Scholar]
- Cakmak, A. Untersuchungen zur Inzidenz von Haemoparasiten in der Provinz Ankara; Tieraerztliche Hochschule Hannover: Hanover, Germany, 1987. [Google Scholar]
- Sayın, F.D.Ş.; Karaer, Z.; Çakmak, A.; İnci, A.; Yukarı, B.A.; Eren, H.; Friehoff, K.T.; Müler, I. Studies of Sero-Prevalence Babesia Infection of Cattle in Turkey. In New Dimensions in Parasitology; Ozcel, M.A., Ed.; Turkish Society for Parasitology: İzmir, Turkey, 1996; Volume 20, pp. 505–516. [Google Scholar]
- Oncel, T.; Vural, G.; Karaer, Z.; Çakmak, A.; Yurtalan, S.; Öz, İ.; Erhan, Z.N.; Beyazıt, A.; Pışkın, Ç.; Deniz, A.; et al. Seroprevalence of Babesia bovis and B. bigemina in cattle in Turkey. Pendik Vet. Mikrobiyol. Derg 2010, 37, 33–42. [Google Scholar]
- Tanyüksel, M.; Vatansever, Z.; Karaer, Z.; Araz, E.; Haznedaroglu, T.; Yukari, B.A.; Acici, M. Sığır babesiosisinin epidemiyolojisi ve zoonotik önemi. Turkiye Parazitol. Derg. 2002, 26, 42–47. [Google Scholar]
- Yildirim, A.; Duzlu, O.; Inci, A.; Önder, Z.; Ciloglu, A. Sığırlarda Babesia bovis ve Babesia bigemina’nın Reverse Line Blotting, Nested PCR ve Real Time PCR Teknikleri İle Karşılaştırmalı Tanısı. Kafkas Univ. Vet. Fak. Derg. 2013, 19, 895–902. [Google Scholar] [CrossRef]
- Mimioğlu, M.G.K.; Sayın, F. Veteriner ve Tıbbi Protozooloji II; Ankara Üniversitesi Basımevi: Ankara, Turkey, 1969. [Google Scholar]
- Vatansever, Z.; Ica, A.; Deniz, A.; Nalbantoğlu, S.; Karaer, Z.; Çakmak, A.; Sparagano, O. Ankara yöresinde sığırlarda kene kaynaklı protozoon enfeksiyonlarının yayılışının reverse line blotting (RLB)ve indirek floresan antikor testi (IFAT) ile saptanması. In Proceedings of the13th Ulusal Parazitoloji Kongresi, Konya, Turkey, 8–12 September 2003; p. 194. [Google Scholar]
- Altay, K.; Aydin, M.F.; Dumanli, N.; Aktas, M. Molecular detection of Theileria and Babesia infections in cattle. Vet. Parasitol. 2008, 158, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Kose, O.B.H.B.; Bakirci, S.; Karagenc, T.; Aksulu, A.; Yukari, B.A.; Adanir, R.; Yildiz, S.; Eren, H. Research of Theileria and Babesia species in ruminantia of Burdur province by reverse line blot hybridisation technique. In Proceedings of the 20 Ulusal Parazitoloji Kongresi (Uluslararası Katılımlı), Eskisehir, Turkey, 25–29 September 2017. [Google Scholar]
- Altay, K.; Atas, A.D.; Ograk, Y.Z.; Ozkan, E. Survey of Theileria, Babesia and Anaplasma infections of cattle and ticks from Sivas province of Turkey. Erciyes Üniv. Vet. Fak. Derg. 2020, 17, 32–38. [Google Scholar]
- Yildirim, D. Önder; Sığırlarda Babesia bovis ve Babesia bigemina’nın Real-Time PCR ile araştırılması ve izolatların moleküler karakterizasyonu. Ankara Üniv. Vet. Fak. Derg. 2015, 62, 27–35. [Google Scholar] [CrossRef]
- Inci, Ç.A. Babesia bigemina Kayseri/Türkiye suşunun in vivo ve in vitro izolatlarında apikal membran antijen-1 proteininin moleküler karakterizasyonu ve ekspresyonu. Ankara Üniv. Vet. Fak. Derg. 2018, 65, 239–246. [Google Scholar] [CrossRef]
- Goksu, K. Ankara ve civarı sığırlarında theileriosis üzerinde sistematik araştırmalar. Ankara Üniv. Vet. Fak. Derg. 1959, 115, 60. [Google Scholar]
- Goksu, K. Yurdumuzun çeşitli bölgelerinde sığırlarda Piroplasmida enfeksiyonları (piroplasmossis, babesiosis, theileriosis ve anaplasmosis’in yayılış durumları). Türk. Vet. Hek. Dern. Derg. 1970, 40, 29–39. [Google Scholar]
- Tuzer, E. İstanbul ili ve çevresinde sığırlarda görülen Babesia, Theileria ve Anaplasma türleri ve bunlardan oluşan enfeksiyonların yayılışı üzerinde araştırma. İstanbul Üniv. Vet. Fak. Derg. 1981, 8, 97–110. [Google Scholar]
- Ozer, E.; Erdogmus, S.Z.; Köroğlu, E.; Yilmaz, F. Malatya ve Güneydoğu Anadolu illerinde sığır, koyun ve keçilerde bulunan kan parazitleri ve yayılışları. Turk. J. Vet. Am. Sci. 1993, 17, 209–215. [Google Scholar]
- Acici, M. Samsun ve yöresi sığırlarında kan parazitlerinin yayılışı. Etlik. Vet. Mik. Derg. 1995, 8, 271–277. [Google Scholar]
- Inci, A.; Cakmak, A.; Karaer, Z.; Dincer, S.; Sayin, F.; Ica, A. Seroprevalence of bovine babesiosis around Kayseri. Turk. J. Vet. Anim. Sci. 2002, 26, 1345–1350. [Google Scholar]
- Inci, A. Ankara’nın Çubuk Ilçesinde Sığırlarda Babesiosis’in Seroinsidensi Üzerine Araştırmalar. Ph.D. Thesis, Üniversitesi Sağlık Bilimleri Enstitüsü, Ankara, Turkey, 1992. [Google Scholar]
- Sayın, F.; Friedhoff, K.T.; Dincer, S.; Karaer, Z.; Cakmak, A. Beytepe köyünde sığır babesiosisi üzerinde seroepidemiyolojik araştırmalar. In Proceedings of the 6th Ulusal Parazitoloji Kongresi, Istanbul, Turkey, 26–29 September 1989. [Google Scholar]
- Eren, H. Ankara yöresinde sığır babesiosisinin sero-prevalansı. Ankara Üniv. Vet. Fak. Derg. 1993, 40, 23–27. [Google Scholar]
- Ica, A. Sığırlarda bazı Babesia türlerinin indirek floresan antikor ve reverse line blotting yöntemi ile karşılaştırmalı tanısı. Erciyes Üniv. Vet. Fak. Derg. 2004, 1, 77–85. [Google Scholar]
- Aktas, M.; Dumanli, N.; Karaer, Z.; Cakmak, A.; Sevgili, M. Elazıg, Malatya, ve Tunceli illerinde sığırlarda Babesia türlerinin seroprevalansı. Turk. J. Vet. Anim. Sci. 2001, 25, 447–451. [Google Scholar]
- Kalkan, K.; Ozcelik, S.; Malatyalı, E. Sivas’ta sıgırlarda babesiosis seroprevalansı. Turkiye Parazitol. Derg. 2010, 34, 11–16. [Google Scholar]
- Bilgin, Z. Trakya’da Sıgırlarda bulunan Theileria ve Babesia türlerinin ve bunların sıgırlarda yaygınlıgının reverse line blooting (rlb) teknigi ile arastırılması. Ph.D. Thesis, Istanbul University, Istanbul, Turkey, 2007. [Google Scholar]
- Duzlu, O.; Inci, A.; Yildirim, A. Karadeniz Bölgesi’ndeki sıgırlardan elde edilen Babesia bovis suslarının moleküler karakterizasyonu. ERÜ Sağ. Bil. Derg. 2011, 20, 18–28. [Google Scholar]
- Yavuz, A.; Inci, A.; Duzlu, O.; Bişkin, Z.; Yildirim, A. Molecular Characterization of Babesia bovis msa-2c Gene. Turk. J. Parasitol. 2011, 35, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Mimioglu, M. Samsun, Ordu, Giresun ve Bolu Vilayetlerinde “Haematurai vesicalis bovis”li sığırlarda parasitolojik araştırmalar. Ankara Üniv. Vet. Fak. Derg. 1955, 11, 183–192. [Google Scholar]
- Ozcan, C.H. Ankara ve civarında evcil hayvanlarda görülen Piroplasmose vakaları ve tedavileri üzerinde araştırmalar. A. Ü. Vet. Fak. Yay. 1961, 143. [Google Scholar]
- Hoffmann, G.H.F.; Schein, E.; Gerber, H.G. Saisonales auftreten von zecken und piroplasmen bei haustieren in des asiatischen provinzen der Turkei. Berl. Münch Tierarztl. Wschr. 1971, 84, 152–156. [Google Scholar]
- Dumanli, N.; Ozer, E. Elazığ yöresinde sığırlarda görülen kan parazitleri ve yayılışları üzerinde araştırmalar. Selçuk Üniv Vet. Fak. Derg. 1987, 3, 159–166. [Google Scholar]
- Dincer, S.; Sayin, F.; Karaer, Z.; Cakmak, A.; Friedhoff, K.T.; Muller, I.; Inci, A.; Yukarı, B.A.; Eren, H. Karadeniz bölgesi sığırlarında bulunan kan parazitlerinin sero-insidensi üzerine araştırmalar. Ankara Üniv. Vet. Fak. Derg. 1991, 38, 206–226. [Google Scholar]
- Çakmak, A.; Öz, İ. Adana yöresi sığırlarında kan protozoonlarının serodiagnozu. Ankara Üniv. Vet. Fak. Derg. 1993, 40, 70–77. [Google Scholar]
- Vatansever, Z.; Nalbantoglu, S.; Çakmak, A. Çukurova bölgesinde sığır babesiosis’nin epidemiyolojisi. In Proceedings of the 12th Ulusal Parazitoloji Kongresi, Elazig, Turkey, 24–28 September 2001. [Google Scholar]
- Sevinc, F.; Sevinc, M.; Birdane, F.M.; Altinoz, F. Prevalence of Babesia bigemina in cattle. Rev. Med. Vet. Toulouse 2001, 152, 395–398. [Google Scholar]
- Kaya, G.; Cakmak, A.; Karaer, Z. Seroprevalance of theileriosis and babesiosis of cattle. Medycyna Wet. 2006, 62, 156–158. [Google Scholar]
- Sevgili, M.; Cakmak, A.; Gokcen, A.; Altas, M.G.; Ergun, G. Prevalence of Theileria annulata and Babesia bigemina in Cattle in the Vicinity of Sanliurfa. J. Anim. Vet. Adv. 2010, 9, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Ica, A.; Inci, A.; Yildirim, A. Parasitological and molecular prevalence of bovine Theileria and Babesia species in the vicinity of Kayseri. Turk. J. Vet. Anim. Sci. 2007, 31, 33–38. [Google Scholar]
- Zhou, M.; Cao, S.; Sevinc, F.; Sevinc, M.; Ceylan, O.; Moumouni, P.F.A.; Jirapattharasate, C.; Liu, M.; Wang, G.; Iguchi, A.; et al. Molecular detection and genetic identification of Babesia bigemina, Theileria annulata, Theileria orientalis and Anaplasma marginale in Turkey. Ticks Tick-Borne Dis. 2016, 7, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Walton, T.E. The impact of diseases on the importation of animals and animal products. Ann. N. Y. Acad. Sci. 2000, 916, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Barré, N.; Happold, J.; Delathière, J.-M.; Desoutter, D.; Saléry, M.; De Vos, A.; Marchal, C.; Perrot, R.; Grailles, M.; Mortelecque, A. A campaign to eradicate bovine babesiosis from New Caledonia. Ticks Tick-Borne Dis. 2011, 2, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Gebrekidan, H.; Nelson, L.; Smith, G.; Gasser, R.B.; Jabbar, A. An outbreak of oriental theileriosis in dairy cattle imported to Vietnam from Australia. Parasitology 2016, 144, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Madder, M.; Adehan, S.; De Deken, R.; Adehan, R.; Lokossou, R. New foci of Rhipicephalus microplus in West Africa. Exp. Appl. Acarol. 2012, 56, 385–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grisi, L.; Leite, R.C.; Martins, J.R.D.S.; De Barros, A.T.M.; Andreotti, R.; Cançado, P.H.D.; De León, A.A.P.; Pereira, J.B.; Villela, H.S. Reassessment of the potential economic impact of cattle parasites in Brazil. Rev. Brasil. Parasitol. Vet. 2014, 23, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, T.; Tuvshintulga, B.; Kothalawala, H.; Silva, S.S.P.; Lan, D.T.B.; Long, P.T.; Ybañez, A.P.; Ybañez, R.H.D.; Benitez, D.F.; Tayebwa, D.S.; et al. Host range and geographical distribution of Babesia sp. Mymensingh. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Gulegen, E.; Giriskin, A.O.; Bakırcı, S.; Senlik, B.; Aydın, L. Sığırların doğal kene enfestasyonlarında bazı sentetik pyrethroid’lerin etkisi. Etlik. Vet. Mikrobiyol. Derg. 2011, 22, 27–31. [Google Scholar]
- Guven, E.; Deniz, A.; Gunduz, N.; Ozubek, S.; Kar, S.; Aktas, M.; Vatansever, Z. Kırım-Kongo Kanamalı Ateşi hastalığının endemik olduğu bir bölgede kene popülasyonunu kontrol amacıyla sığırlarda 1 Flumethrin uygulanmasının etkinliği. In Proceedings of the 18th Ulusal Parazitoloji Kongresi, Denizli, Turkey, 1–4 September 2013. [Google Scholar]
- Brayton, K.A.; Lau, A.O.T.; Herndon, D.R.; Hannick, L.; Kappmeyer, L.S.; Berens, S.J.; Bidwell, S.L.; Brown, W.C.; Crabtree, J.; Fadrosh, D.; et al. Genome Sequence of Babesia bovis and Comparative Analysis of Apicomplexan Hemoprotozoa. PLoS Pathog. 2007, 3, e148-13. [Google Scholar] [CrossRef]
- Suarez, C.E.; Bishop, R.; Alzan, H.; Poole, W.; Cooke, B.M. Advances in the application of genetic manipulation methods to apicomplexan parasites. Int. J. Parasitol. 2017, 47, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.E.; Alzan, H.F.; Cooke, B.M. Genomics and Genetic Manipulation of Protozoan Parasites Affecting Farm Animals. In Parasitic Protozoa of Farm Animals and Pets; Springer Science and Business Media LLC: Berlin, Germany, 2018; pp. 413–438. [Google Scholar]
- Vimonish, R.; Johnson, W.C.; Mousel, M.R.; Brayton, K.A.; Scoles, G.A.; Noh, S.M.; Ueti, M.W. Quantitative analysis of Anaplasma marginale acquisition and transmission by Dermacentor andersoni fed in vitro. Sci. Rep. 2020, 10, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakimi, H.; Ishizaki, T.; Kegawa, Y.; Kaneko, O.; Kawazu, S.-I.; Asada, M. Genome Editing of Babesia bovis Using the CRISPR/Cas9 System. mSphere 2019, 4, e00109-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, M.; Kasaija, P.D.; Merino, O.; De La Cruz-Hernandez, N.I.; Gortazar, C.; De La Fuente, J. Oral Vaccination with a Formulation Combining Rhipicephalus microplus Subolesin with Heat Inactivated Mycobacterium bovis Reduces Tick Infestations in Cattle. Front. Cell. Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente, J.; Estrada-Peña, A. Why New Vaccines for the Control of Ectoparasite Vectors Have Not Been Registered and Commercialized? Vaccines 2019, 7, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Species | Geographical Distribution | Vectors/ITS/TOT | References |
|---|---|---|---|
| B. bovis * | tropics, subtropics | R. annulatus Ψ/L/+ R. australis/L/+ R. microplus/L/+ | [36,37,38] |
| B. bigemina * | tropics, subtropics | R. annulatus Ψ/A/+ R. australis/N, A/+ R. decoloratus/N, A/+ R. microplus/N, A/+ R. evertsi/N/− R. bursa Ψ/N, A/± | [36,39,40,41] |
| B. divergens * | Europe, North Africa, Russia | Ixodes ricinus Ψ/L, N, A/+ | [42] |
| B. major * | Europe, North Africa, temperate Asia | Hae. punctata Ψ/L, N, A/+ | [40] |
| B. occultans * | Africa (Southern Europe, Russia) | Hy. rufipes Ψ/A/+ | [43] |
| B. orientalis | East Asia | R. haemaphysaloides/A/+ | [44] |
| B. ovata | East Asia | Hae. longicornis/L, N, A/+ | [45] |
| Species | Method | Prevelance (%) | Reference |
|---|---|---|---|
| B. bovis | ME | 0.2–34.8 | Göksu 1959 [120]; Göksu 1970 [121]; Cakmak 1987 [108]; Tuzer 1981 [122]; Ozer et al., 1993 [123]; Acıcı 1995 [124]; Inci et al., 2002 [125]; Inci 1992 [126] |
| SM | 0.6–59 | Cakmak 1987 [108]; Sayın et al., 1989 [127]; Dincer et al., 1991; Inci 1992 [126]; Eren 1993 [128]; Sayın et al., 1996 [109]; Ica 2004 [129]; Vatansever et al., 2003 [114]; Aktas et al., 2001 [130]; Inci et al. 2002 [125]; Sayın et al. 1996 [109]; Kalkan et al., 2010 [131]; Oncel et al., 2010 [110] | |
| MT | 0.4–12.7 | Tanyüksel et al., 2002 [111]; Bilgin 2007 [132]; Duzlu et al., 2011 [133]; Yavuz et al., 2011 [134]; Aktas and Ozubek 2015 [100]; Duzlu et al., 2015 [118]; Kose et al., 2017 [116] | |
| B. bigemina | ME | 0.6–32.2 | Mimioglu et al., 1955 [135]; Goksu et al., 1959 [120]; Ozcan 1961 [136]; Hoffman et al., 1971 [137]; Tüzer 1981 [122]; Dumanlı and Özer 1987 [138]; Sayin et al., 1989 [127]; Inci 1992 [126]; Ozer et al., 1993 [123]; Acıcı 1995 [124]; Inci et al., 2002 [125] |
| SM | 0.9–80 | Cakmak 1987 [108]; Dincer et al., 1991 [139]; Eren 1993 [128]; Sayın et al., 1996 [109]; Ica 2004 [129]; Vatansever et al., 2003 [114]; Cakmak and Oz 1993 [140]; Vatansever et al., 2001 [141]; Aktas et al., 2001 [130]; Sevinc et al., 2001 [142]; Inci et al., 2002 [125]; Kaya et al., 2006 [143]; Sayın et al., 1996 [109]; Ekici and Sevinc 2009 [33]; Sevgili et al., 2010 [144]; Kalkan et al., 2010 [131] | |
| MT | 0.6–18.2 | Tanyuksel et al., 2002 [111]; Ica 2004 [129]; Ica et al., 2007 [145]; Altay et al., 2008 [115]; Duzlu et al., 2011 [133]; Aktas and Ozubek 2015 [100]; Duzlu et al., 2015 [118]; Zhou et al., 2016 [146] | |
| B. divergens | ME | 0.8 | Aktas and Ozubek 2015 [100] |
| SM | 0.6–75 | Sayın et al., 1996 [109]; Aktas et al., 2001 [130]; Inci et al., 2002 [125] | |
| MT | 1.1–7.4 | Tanyüksel et al., 2002 [111]; Vatansever et al., 2003 [114]; Aktas and Ozubek 2015 [100] | |
| B. major | MT | 0.2–0.5 | Vatansever et al., 2003 [114]; Altay et al., 2008 [115] |
| B. occultans | MT | 3 | Aktas et al., 2014 [100]; Aktas and Ozubek 2015 [100] |
| Babesia spp. | MT | 1.3 | Altay et al., 2008 [115] |
| Babesia spp. | MT | 3.3 | Kose et al., 2017 [116] |
| Babesia sp. Sivas | MT | 1.9 | Altay et al., 2020 [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozubek, S.; Bastos, R.G.; Alzan, H.F.; Inci, A.; Aktas, M.; Suarez, C.E. Bovine Babesiosis in Turkey: Impact, Current Gaps, and Opportunities for Intervention. Pathogens 2020, 9, 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9121041
Ozubek S, Bastos RG, Alzan HF, Inci A, Aktas M, Suarez CE. Bovine Babesiosis in Turkey: Impact, Current Gaps, and Opportunities for Intervention. Pathogens. 2020; 9(12):1041. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9121041
Chicago/Turabian StyleOzubek, Sezayi, Reginaldo G. Bastos, Heba F. Alzan, Abdullah Inci, Munir Aktas, and Carlos E. Suarez. 2020. "Bovine Babesiosis in Turkey: Impact, Current Gaps, and Opportunities for Intervention" Pathogens 9, no. 12: 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9121041