
Jasmonate Signalling and Defence Responses in the Model Legume Medicago truncatula—A Focus on Responses to Fusarium Wilt Disease

Abstract
:1. Introduction
1.1. Pathogen Background
1.2. Host Jasmonate Signaling and F. oxysporum Disease Outcome
1.3. Fusarium Wilt of Legumes
2. Genomic and Transcriptomic M. truncatula Resources
Utilizing Gene Expression Resources to Study Medicago Responses to F. oxysporum Infection

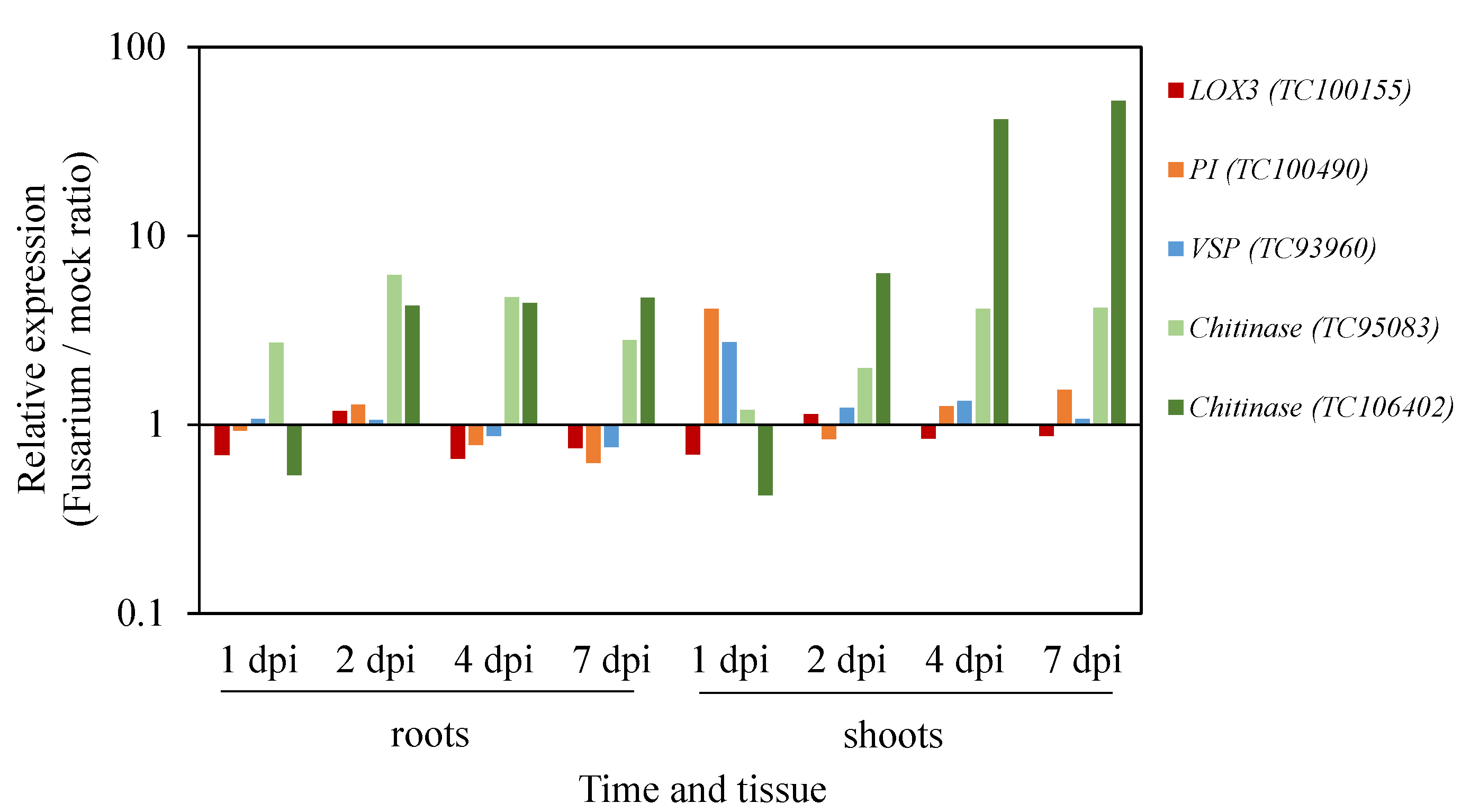{kind=link}
{kind=link}
| Term | Description | FDR |
|---|---|---|
| GO:0044255 | cellular lipid metabolic process | 1.40E-17 |
| GO:0019748 | secondary metabolic process | 1.40E-15 |
| GO:0008610 | lipid biosynthetic process | 1.40E-15 |
| GO:0032787 | monocarboxylic acid metabolic process | 2.40E-14 |
| GO:0050896 | response to stimulus | 4.80E-14 |
| GO:0009607 | response to biotic stimulus | 2.60E-13 |
| GO:0010033 | response to organic substance | 2.30E-11 |
| GO:0016053 | organic acid biosynthetic process | 3.10E-11 |
| GO:0046394 | carboxylic acid biosynthetic process | 3.10E-11 |
| GO:0006629 | lipid metabolic process | 3.80E-11 |
| GO:0043436 | oxoacid metabolic process | 4.80E-11 |
| GO:0006082 | organic acid metabolic process | 4.80E-11 |
| GO:0019752 | carboxylic acid metabolic process | 4.80E-11 |
| GO:0042180 | cellular ketone metabolic process | 4.80E-11 |
| GO:0006720 | isoprenoid metabolic process | 5.20E-11 |
| GO:0006631 | fatty acid metabolic process | 7.20E-11 |
| GO:0006633 | fatty acid biosynthetic process | 1.90E-10 |
| GO:0009719 | response to endogenous stimulus | 2.30E-10 |
| GO:0042221 | response to chemical stimulus | 7.20E-10 |
| GO:0008299 | isoprenoid biosynthetic process | 1.10E-09 |

3. Genetic/Mutant Resources Available in M. truncatula
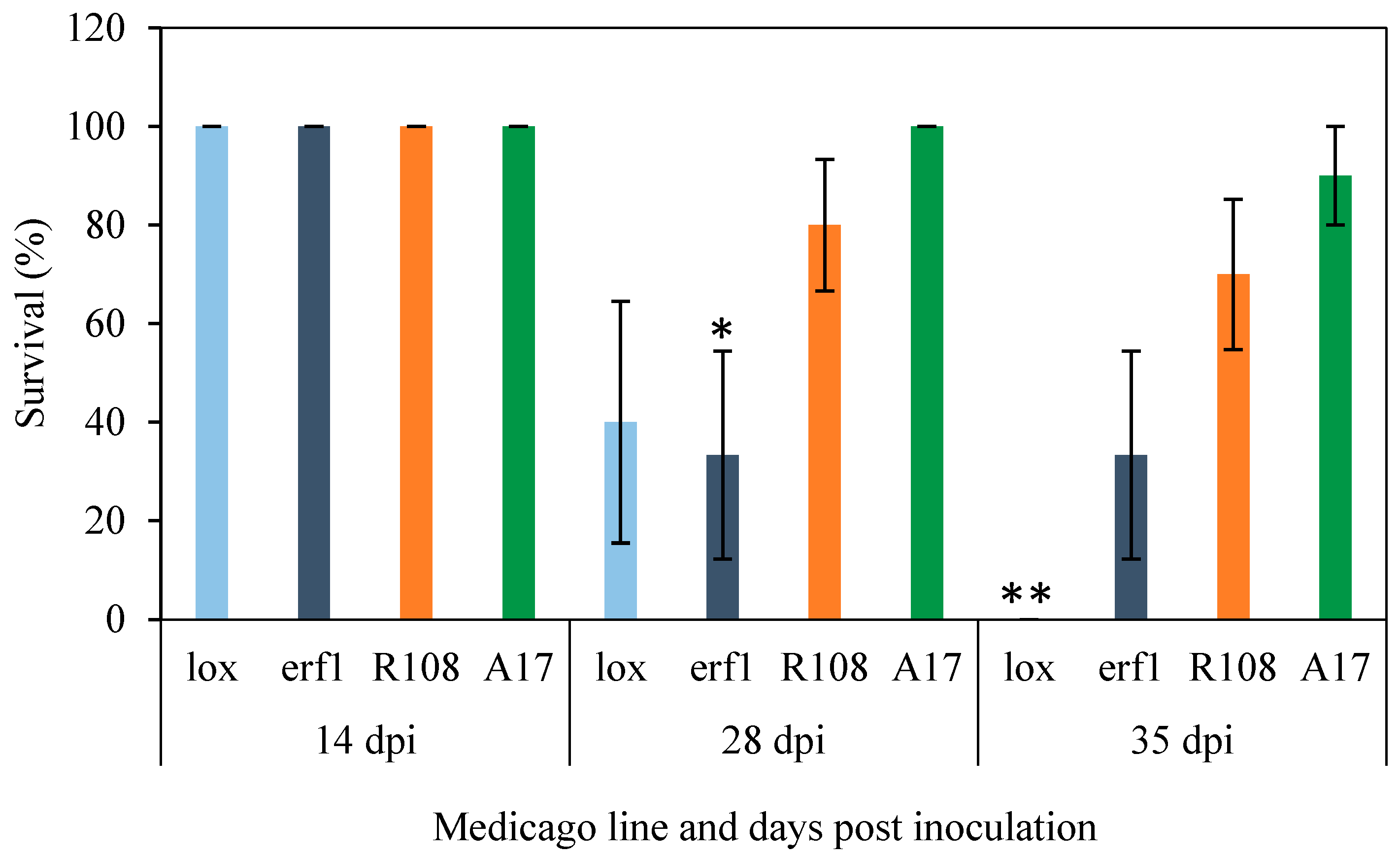
Utilizing M. truncatula Mutant Resources to Dissect Host JA-Responses under F. oxysporum Infection
| Putative Mutant | Medtr ID | TC | Insertion Line | Gene Function | Arabidopsis Homologue |
|---|---|---|---|---|---|
| Mtlox1 | Medtr8g018430 | TC132688 | NF0217 insertion Ase8 | 9S-lipoxygenase | AT1G55020 LOX1 AT3G22400 LOX5 |
| Mterf1 | Medtr4g100380 | TC114237 | NF1858 insertion 26 | MtERF1-A transcription factor | AT4G17500 ATERF-1 |

4. Experimental Section
4.1. Plant Growth Conditions
4.2. Pathogen Assays
4.3. qRT-PCR
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Beckman, C.H. (Ed.) The Nature of Wilt Diseases of Plants; Aps Press: St. Paul, MN, USA, 1987.
- Gordon, T.R.; Martyn, R.D. The Evolutionary Biology of Fusarium oxysporum. Annu. Rev. Phytopathol. 1997, 35, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Recorbet, G.; Steinberg, C.; Olivain, C.; Edel, V.; Trouvelot, S.; Dumas-Gaudot, E.; Gianinazzi, S.; Alabouvette, C. Wanted: Pathogenesis-Related Marker Molecules for Fusarium oxysporum. New Phytol. 2003, 159, 73–92. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen Profile Update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Kidd, B.N.; Kadoo, N.Y.; Dombrecht, B.; Tekeoglu, M.; Gardiner, D.M.; Thatcher, L.F.; Aitken, E.A.; Schenk, P.M.; Manners, J.M.; Kazan, K. Auxin Signaling and Transport Promote Susceptibility to the Root-Infecting Fungal Pathogen Fusarium oxysporum in Arabidopsis. Mol. Plant Microbe Interact. 2011, 24, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Perez-Nadales, E.; Almeida Nogueira, M.F.; Baldin, C.; Castanheira, S.; El Ghalid, M.; Grund, E.; Lengeler, K.; Marchegiani, E.; Mehrotra, P.V.; Moretti, M.; et al. Fungal Model Systems and the Elucidation of Pathogenicity Determinants. Fungal Genet. Biol. 2014, 70, 42–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrios, G.N. Plant Diseases Caused By Fungi. In Plant Pathology, 5th ed.; Agrios, G.N., Ed.; Academic Press: San Diego, CA, USA, 2005; Chapter Eleven; pp. 385–614. [Google Scholar]
- Gupta, S.; Bhar, A.; Das, S. Understanding the Molecular Defence Responses of Host During Chickpea–Fusarium Interplay: Where do We Stand? Funct. Plant Biol. 2013, 40, 1285–1297. [Google Scholar] [CrossRef]
- Pietro, A.D.; Madrid, M.P.; Caracuel, Z.; Delgado-Jarana, J.; Roncero, M.I.G. Fusarium oxysporum: Exploring the Molecular Arsenal of a Vascular Wilt Fungus. Mol. Plant Pathol. 2003, 4, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Berrocal-Lobo, M.; Molina, A. Arabidopsis Defense Response against Fusarium oxysporum. Trends Plant Sci. 2008, 13, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations, Statistics Division. Available online: http://faostat3.fao.org/home/e (accessed on 15 May 2015).
- Navas-Cortés, J.A.; Hau, B.; Jiménez-Díaz, R.M. Yield Loss in Chickpeas in Relation to Development of Fusarium Wilt Epidemics. Phytopathology 2000, 90, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Trapero-Casas, A.; Jiménez-Díaz, R.M. Fungal Wilt and Root Rot Diseases of Chickpea in Southern Spain. Phytopathology 1985, 75, 1146–1151. [Google Scholar] [CrossRef]
- Sharma, K.D.; Chen, W.; Muehlbauer, F.J. Genetics of Chickpea Resistance to Five Races of Fusarium Wilt and a Concise Set of Race Differentials for Fusarium oxysporum F. Sp. Ciceris. Plant Dis. 2005, 89, 385–390. [Google Scholar] [CrossRef]
- Diener, A.C.; Ausubel, F.M. Resistance to Fusarium oxysporum 1, a Dominant Arabidopsis Disease-Resistance Gene, Is Not Race Specific. Genetics 2005, 171, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; Maclean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic Interaction Between Abscisic Acid and Jasmonate-Ethylene Signaling Pathways Modulates Defense Gene Expression and Disease Resistance in Arabidopsis. Plant Cell 2004, 16, 3460–3479. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Manners, J.M.; Kazan, K. Fusarium oxysporum Hijacks Coi1-Mediated Jasmonate Signaling to Promote Disease Development in Arabidopsis. Plant J. 2009, 58, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Edgar, C.I.; Mcgrath, K.C.; Dombrecht, B.; Manners, J.M.; Maclean, D.C.; Schenk, P.M.; Kazan, K. Salicylic Acid Mediates Resistance to the Vascular Wilt Pathogen Fusarium oxysporum in the Model Host Arabidopsis thaliana. Australas. Plant Pathol. 2006, 35, 581–591. [Google Scholar] [CrossRef]
- Kidd, B.N.; Edgar, C.I.; Kumar, K.K.; Aitken, E.A.; Schenk, P.M.; Manners, J.M.; Kazan, K. The Mediator Complex Subunit Pft1 Is a Key Regulator of Jasmonate-Dependent Defense in Arabidopsis. Plant Cell 2009, 21, 2237–2252. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Kamphuis, L.G.; Hane, J.K.; Oñate-Sánchez, L.; Singh, K.B. The Arabidopsis Kh-Domain Rna-Binding Protein Esr1 Functions in Components of Jasmonate Signalling, Unlinking Growth Restraint and Resistance to Stress. PLoS ONE 2015, 10, E0126978. [Google Scholar] [CrossRef] [PubMed]
- Takken, F.; Rep, M. The Arms Race between Tomato and Fusarium oxysporum. Mol. Plant Pathol. 2010, 11, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Laluk, K.; Mengiste, T. Necrotroph Attacks on Plants: Wanton Destruction or Covert Extortion? Arabidopsis Book 2010, 8, E0136. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.; Eggermont, K.; Penninckx, I.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.; Broekaert, W.F. Separate Jasmonate-Dependent and Salicylate-Dependent Defense-Response Pathways in Arabidopsis Are Essential for Resistance to Distinct Microbial Pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Gardiner, D.M.; Kazan, K.; Manners, J.M. A Highly Conserved Effector in Fusarium oxysporum Is Required for Full Virulence on Arabidopsis. Mol. Plant Microbe Interact. 2012, 25, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Lyons, R.; Stiller, J.; Powell, J.; Rusu, A.; Manners, J.M.; Kazan, K. Fusarium oxysporum Triggers Tissue-Specific Transcriptional Reprogramming in Arabidopsis thaliana. PLoS ONE 2015, 10, E0121902. [Google Scholar] [CrossRef] [PubMed]
- Schenk, P.M.; Kazan, K.; Rusu, A.G.; Manners, J.M.; Maclean, D.J. The Sen1 Gene of Arabidopsis Is Regulated By Signals that Link Plant Defence Responses and Senescence. Plant Physiol. Biochem. 2005, 43, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Mcgrath, K.C.; Dombrecht, B.; Manners, J.M.; Schenk, P.M.; Edgar, C.I.; Maclean, D.J.; Scheible, W.R.; Udvardi, M.K.; Kazan, K. Repressor- and Activator-Type Ethylene Response Factors Functioning in Jasmonate Signaling and Disease Resistance Identified via a Genome-Wide Screen of Arabidopsis Transcription Factor Gene Expression. Plant Physiol. 2005, 139, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Powell, J.J.; Aitken, E.A.; Kazan, K.; Manners, J.M. The Lateral Organ Boundaries Domain Transcription Factor Lbd20 Functions in Fusarium Wilt Susceptibility and Jasmonate Signaling in Arabidopsis. Plant Physiol. 2012, 160, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, Perception, Signal Transduction and Action in Plant Stress Response, Growth and Development. An Update to the 2007 Review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.J.; Yoon, A.J.; Faull, K.F.; Diener, A.C. Host Perception of Jasmonates Promotes Infection By Fusarium oxysporum Formae Speciales that Produce Isoleucine- and Leucine-Conjugated Jasmonates. Mol. Plant Pathol. 2014, 15, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Feys, B.; Benedetti, C.E.; Penfold, C.N.; Turner, J.G. Arabidopsis Mutants Selected for Resistance to the Phytotoxin Coronatine Are Male Sterile, Insensitive to Methyl Jasmonate, and Resistant to a Bacterial Pathogen. Plant Cell 1994, 6, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Thaler, J.S.; Owen, B.; Higgins, V.J. The Role Of The Jasmonate Response in Plant Susceptibility to Diverse Pathogens With a Range of Lifestyles. Plant Physiol. 2004, 135, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, Y.; Mccaig, B.C.; Wingerd, B.A.; Wang, J.; Whalon, M.E.; Pichersky, E.; Howe, G.A. The Tomato Homolog of Coronatine-Insensitive1 Is Required for the Maternal Control of Seed Maturation, Jasmonate-Signaled Defense Responses, and Glandular Trichome Development. Plant Cell 2004, 16, 126–143. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, R.; Schaarschmidt, S.; Hause, B. Repeated Leaf Wounding Alters The Colonization of Medicago Truncatula Roots by Beneficial and Pathogenic Microorganisms. Plant Cell Environ. 2012, 35, 1344–1357. [Google Scholar] [CrossRef] [PubMed]
- Hilou, A.; Zhang, H.; Franken, P.; Hause, B. Do Jasmonates Play A Role in Arbuscular Mycorrhiza-Induced Local Bioprotection of Medicago truncatula against Root Rot Disease Caused by Aphanomyces Euteiches? Mycorrhiza 2014, 24, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Uppalapati, S.R.; Marek, S.M.; Lee, H.K.; Nakashima, J.; Tang, Y.; Sledge, M.K.; Dixon, R.A.; Mysore, K.S. Global Gene Expression Profiling During Medicago Truncatula-Phymatotrichopsis Omnivora Interaction Reveals a Role for Jasmonic Acid, Ethylene, and the Flavonoid Pathway in Disease Development. Mol. Plant Microbe Interact. 2009, 22, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Kamphuis, L.G.; Williams, A.H.; Kuster, H.; Trengove, R.D.; Singh, K.B.; Oliver, R.P.; Ellwood, S.R. Phoma medicaginis Stimulates the Induction of the Octadecanoid and Phenylpropanoid Pathways in Medicago truncatula. Mol. Plant Pathol. 2012, 13, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.; Schaarschmidt, S. The Role of Jasmonates in Mutualistic Symbioses between Plants and Soil-Born Microorganisms. Phytochemistry 2009, 70, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Van Der Ent, S.; Van Wees, S.C.M.; Pieterse, C.M.J. Jasmonate Signaling in Plant Interactions With Resistance-Inducing Beneficial Microbes. Phytochemistry 2009, 70, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Ballare, C.L. Jasmonate-Induced Defenses: A Tale of Intelligence, Collaborators and Rascals. Trends Plant Sci. 2011, 16, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone Crosstalk in Plant Disease and Defense: More Than Just Jasmonate-Salicylate Antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Browse, J. Jasmonate Passes Muster: A Receptor and Targets for the Defense Hormone. Annu. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Kombrink, E. Jasmonates: Structural Requirements for Lipid-Derived Signals Active in Plant Stress Responses and Development. Acs Chem. Biol. 2010, 5, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, S.; Chini, A.; Hamberg, M.; Adie, B.; Porzel, A.; Kramell, R.; Miersch, O.; Wasternack, C.; Solano, R. (+)-7-Iso-Jasmonoyl-l-Isoleucine Is the Endogenous Bioactive Jasmonate. Nat. Chem. Biol. 2009, 5, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Cevik, V.; Kidd, B.N.; Zhang, P.; Hill, C.; Kiddle, S.; Denby, K.J.; Holub, E.B.; Cahill, D.M.; Manners, J.M.; Schenk, P.M.; et al. Mediator25 Acts as an Integrative Hub for the Regulation of Jasmonate-Responsive Gene Expression in Arabidopsis. Plant Physiol. 2012, 160, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.; Kang, J.-H.; Howe, G. Jasmonate-Triggered Plant Immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Devoto, A.; Ellis, C.; Magusin, A.; Chang, H.-S.; Chilcott, C.; Zhu, T.; Turner, J. Expression Profiling Reveals Coi1 to Be a Key Regulator of Genes Involved in Wound- and Methyl Jasmonate-Induced Secondary Metabolism, Defence, and Hormone Interactions. Plant. Mol. Biol. 2005, 58, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Devoto, A.; Turner, J.G. Regulation of Jasmonate-Mediated Plant Responses in Arabidopsis. Ann. Bot. 2003, 92, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Rispail, N.; Rubiales, D. Identification of Sources of Quantitative Resistance to Fusarium oxysporum F. Sp. Medicaginis in Medicago truncatula. Plant Dis. 2014, 98, 667–673. [Google Scholar] [CrossRef]
- Rispail, N.; Bani, M.; Rubiales, D. Resistance reaction of Medicago truncatula genotypes to Fusarium oxysporum: Effect of plant age, substrate and inoculation method. Crop Pasture Sci. 2015, 66, 506–515. [Google Scholar] [CrossRef]
- Ramírez-Suero, M.; Khanshour, A.; Martinez, Y.; Rickauer, M. A Study on the Susceptibility of the Model Legume Plant Medicago truncatula to the Soil-Borne Pathogen Fusarium oxysporum. Eur. J. Plant Pathol. 2010, 126, 517–530. [Google Scholar] [CrossRef]
- Chatterjee, M.; Gupta, S.; Bhar, A.; Chakraborti, D.; Basu, D.; Das, S. Analysis of Root Proteome Unravels Differential Molecular Responses during Compatible and Incompatible Interaction between Chickpea (Cicer arietinum L.) and Fusarium oxysporum F. Sp. Ciceri Race1 (Foc1). BMC Genom. 2014, 15, 949. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Wu, J.; Zhu, Z.; Wang, L.; Wang, X.; Wang, S.; Blair, M.W. Differentially Expressed Genes in Resistant and Susceptible Common Bean (Phaseolus vulgaris L.) Genotypes in Response to Fusarium oxysporum F. Sp. Phaseoli. PLoS ONE 2015, 10, E0127698. [Google Scholar] [CrossRef] [PubMed]
- Journet, E.-P.; Van Tuinen, D.; Gouzy, J.; Crespeau, H.; Carreau, V.; Farmer, M.-J.; Niebel, A.; Schiex, T.; Jaillon, O.; Chatagnier, O.; et al. Exploring Root Symbiotic Programs in the Model Legume Medicago Truncatula Using Est Analysis. Nucleic Acids Res. 2002, 30, 5579–5592. [Google Scholar] [CrossRef] [PubMed]
- Frugoli, J.; Harris, J. Medicago truncatula on the Move! Plant Cell 2001, 13, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Kim, D.J.; Baek, J.M.; Choi, H.K.; Ellis, L.C.; Kuester, H.; Mccombie, W.R.; Peng, H.M.; Cook, D.R. Syntenic Relationships Between Medicago truncatula and Arabidopsis Reveal Extensive Divergence of Genome Organization. Plant Physiol. 2003, 131, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Salzer, P.; Bonanomi, A.; Beyer, K.; Vogeli-Lange, R.; Aeschbacher, R.A.; Lange, J.; Wiemken, A.; Kim, D.; Cook, D.R.; Boller, T. Differential Expression of Eight Chitinase Genes in Medicago truncatula Roots during Mycorrhiza Formation, Nodulation, and Pathogen Infection. Mol. Plant Microbe Interact. 2000, 13, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Ben, C.; Toueni, M.; Montanari, S.; Tardin, M.C.; Fervel, M.; Negahi, A.; Saint-Pierre, L.; Mathieu, G.; Gras, M.C.; Noel, D.; et al. Natural Diversity in the Model Legume Medicago Truncatula Allows Identifying Distinct Genetic Mechanisms Conferring Partial Resistance to Verticillium Wilt. J. Exp. Bot. 2013, 64, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Medicago truncatula Genome Project. Available online: http://medicago.jcvi.org/medicago/index.php (accessed on 20 November 2015).
- Medicago truncatula Hapmap Project. Available online: http://medicagohapmap.org/ (accessed on 20 November 2015).
- Bell, C.J.; Dixon, R.A.; Farmer, A.D.; Flores, R.; Inman, J.; Gonzales, R.A.; Harrison, M.J.; Paiva, N.L.; Scott, A.D.; Weller, J.W.; et al. The Medicago Genome Initiative: A Model Legume Database. Nucleic Acids Res. 2001, 29, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Medicago truncatula Gene Expression Atlas. Available online: http://mtgea.noble.org/v3/ (accessed on 20 November 2015).
- He, J.; Benedito, V.A.; Wang, M.; Murray, J.D.; Zhao, P.X.; Tang, Y.; Udvardi, M.K. The Medicago truncatula Gene Expression Atlas Web Server. BMC Bioinform. 2009, 10, 441. [Google Scholar] [CrossRef] [PubMed]
- Benedito, V.A.; Torres-Jerez, I.; Murray, J.D.; Andriankaja, A.; Allen, S.; Kakar, K.; Wandrey, M.; Verdier, J.; Zuber, H.; Ott, T.; et al. A Gene Expression Atlas of the Model Legume Medicago truncatula. Plant J. 2008, 55, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Genevestigator. Available online: https://genevestigator.com/gv/ (accessed on 5 August 2015).
- Naoumkina, M.; Farag, M.A.; Sumner, L.W.; Tang, Y.; Liu, C.J.; Dixon, R.A. Different Mechanisms for Phytoalexin Induction by Pathogen and Wound Signals in Medicago Truncatula. Proc. Natl. Acad. Sci. USA 2007, 104, 17909–17915. [Google Scholar] [CrossRef] [PubMed]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated Plant Defense Responses in Arabidopsis Revealed by Microarray Analysis. Proc. Natil. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. Agrigo: A Go Analysis Toolkit for the Agricultural Community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.-L.; Anderson, J.P.; Klingler, J.P.; Nair, R.M.; Edwards, O.R.; Singh, K.B. Involvement of the Octadecanoid Pathway in Bluegreen Aphid Resistance in Medicago Truncatula. Mol. Plant Microbe Interact. 2007, 20, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, M.; Silverstein, K.A.; Nallu, S.; Wang, L.; Botanga, C.J.; Gomez, S.K.; Costa, L.M.; Harrison, M.J.; Samac, D.A.; Glazebrook, J.; et al. Spatio-Temporal Expression Patterns of Arabidopsis thaliana And Medicago truncatula Defensin-Like Genes. PLoS ONE 2013, 8, E58992. [Google Scholar] [CrossRef] [PubMed]
- Tadege, M.; Wang, T.L.; Wen, J.; Ratet, P.; Mysore, K.S. Mutagenesis and Beyond! Tools for Understanding Legume Biology. Plant Physiol. 2009, 151, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Ane, J.M.; Zhu, H.; Frugoli, J. Recent Advances in Medicago truncatula Genomics. Int. J. Plant Genomics 2008, 2008, 256597. [Google Scholar] [CrossRef] [PubMed]
- Carelli, M.; Calderini, O.; Panara, F.; Porceddu, A.; Losini, I.; Piffanelli, P.; Arcioni, S.; Scotti, C. Reverse Genetics in Medicago truncatula Using a Tilling Mutant Collection. Methods Mol. Biol. 2013, 1069, 101–118. [Google Scholar] [PubMed]
- Tadege, M.; Wen, J.; He, J.; Tu, H.; Kwak, Y.; Eschstruth, A.; Cayrel, A.; Endre, G.; Zhao, P.X.; Chabaud, M.; et al. Large-Scale Insertional Mutagenesis Using the Tnt1 Retrotransposon in the Model Legume Medicago truncatula. Plant J. 2008, 54, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Penmetsa, R.V.; Cook, D.R. Production And Characterization of Diverse Developmental Mutants of Medicago truncatula. Plant Physiol. 2000, 123, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- Le Signor, C.; Savois, V.; Aubert, G.; Verdier, J.; Nicolas, M.; Pagny, G.; Moussy, F.; Sanchez, M.; Baker, D.; Clarke, J.; et al. Optimizing Tilling Populations for Reverse Genetics in Medicago Truncatula. Plant Biotech. J. 2009, 7, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, M.; Lee, H.-K.; Tadege, M.; Ratet, P.; Udvardi, M.; Mysore, K.S.; Wen, J. An Efficient Reverse Genetics Platform in the Model Legume Medicago truncatula. New Phytol. 2014, 201, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Medicago truncatula Mutant Database. Available online: http://nedicago-nutant.noble.org/mutant/ (accessed on 20 November 2015).
- Epple, P.; Apel, K.; Bohlmann, H. Overexpression of an Endogenous Thionin Enhances Resistance of Arabidopsis against Fusarium oxysporum. Plant Cell 1997, 9, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.L.; Prasad, V.; Sanjaya; Chen, K.H.; Liu, P.C.; Chan, M.T.; Cheng, C.P. Transgenic Tomato Plants Expressing an Arabidopsis Thionin (Thi2.1) Driven by Fruit-Inactive Promoter Battle against Phytopathogenic Attack. Planta 2005, 221, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Berrocal-Lobo, M.; Molina, A. Ethylene Response Factor 1 Mediates Arabidopsis Resistance to the Soilborne Fungus Fusarium oxysporum. Mol. Plant Microbe Interact. 2004, 17, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.P.; Singh, K.B. Interactions of Arabidopsis and M. truncatula with the Same Pathogens Differ in Dependence on Ethylene and Ethylene Response Factors. Plant Signal. Behav. 2011, 6, 551–552. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Wasternack, C. The Lipoxygenase Pathway. Annu. Review Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.; Chaturvedi, R. Lipid Signals in Plant–Pathogen Interactions. In Annual Plant Reviews Volume 34: Molecular Aspects Of Plant Disease Resistance; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 292–333. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thatcher, L.F.; Gao, L.-L.; Singh, K.B. Jasmonate Signalling and Defence Responses in the Model Legume Medicago truncatula—A Focus on Responses to Fusarium Wilt Disease. Plants 2016, 5, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/plants5010011
Thatcher LF, Gao L-L, Singh KB. Jasmonate Signalling and Defence Responses in the Model Legume Medicago truncatula—A Focus on Responses to Fusarium Wilt Disease. Plants. 2016; 5(1):11. https://0-doi-org.brum.beds.ac.uk/10.3390/plants5010011
Chicago/Turabian StyleThatcher, Louise F., Ling-Ling Gao, and Karam B. Singh. 2016. "Jasmonate Signalling and Defence Responses in the Model Legume Medicago truncatula—A Focus on Responses to Fusarium Wilt Disease" Plants 5, no. 1: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/plants5010011