Isolation and Characterization of Plant Growth-Promoting Endophytic Fungi from the Roots of Dendrobium moniliforme


Abstract
:1. Introduction
2. Results
2.1. Molecular Identification of Endophytic Fungi
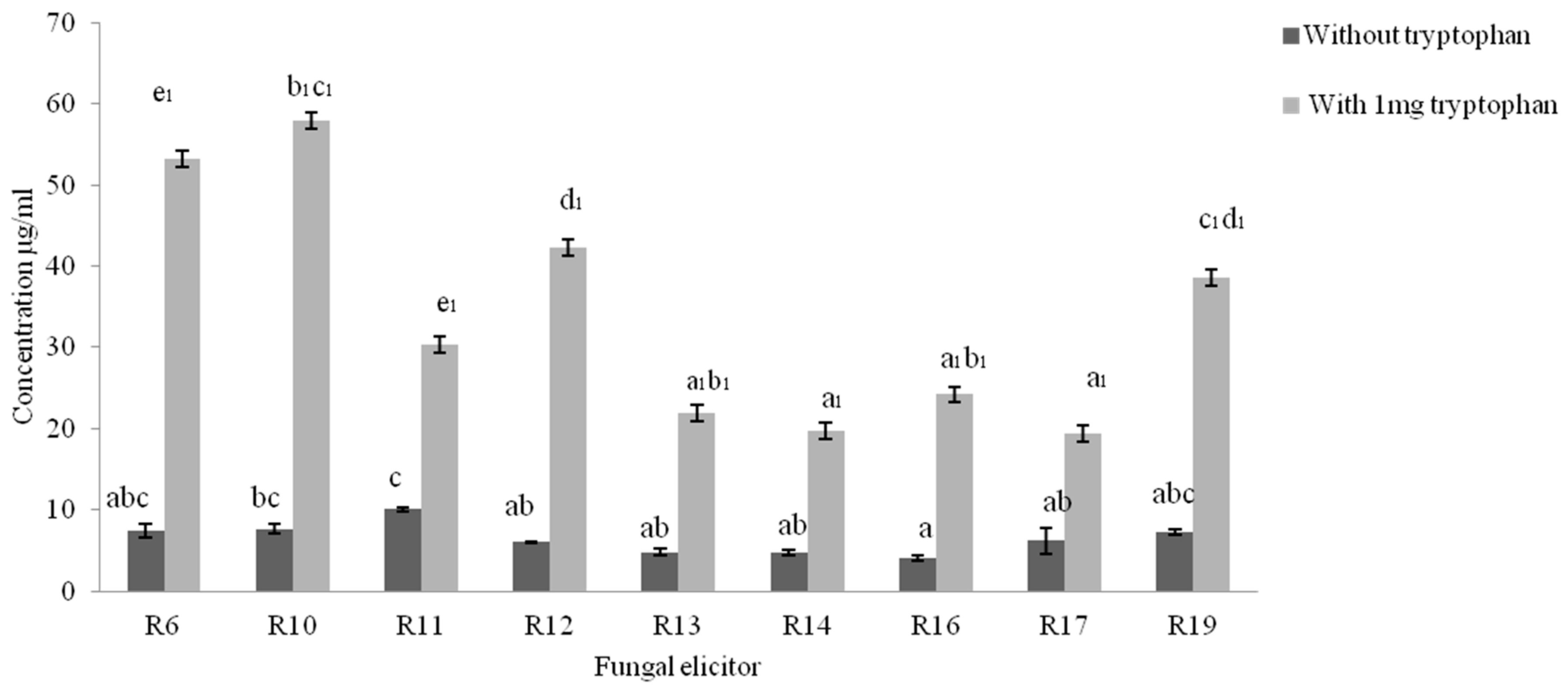
2.2. Quantification of Indole Acetic Acid (IAA) Synthesis by Isolated Fungi
2.3. Detection of Bioactive Compound by Gas Chromatography-Mass Spectroscopy (GC-MS) analysis
2.4. Plant Growth Assay with Fungal Elicitor Treatment
3. Discussion
4. Materials and Methods
4.1. Surface Sterilization of Roots
4.2. Endophytes Isolation and Axenically Culture
4.3. DNA Extraction, PCR Amplification and Sequencing
4.4. Biochemical Assays
4.5. Quantitative Estimation of IAA in Culture Extract
4.6. Identification of Compounds Present in Fungal Extract
4.7. Plant Growth Assay after Supplementation with Fungal Elicitor
4.8. Quantification of Chlorophyll Content in Fungal Elicitor Plantlet
4.9. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Supplementary File 1Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PDA | Potato dextrose agar |
| CDA | Czapek Dox agar |
| MS | Murashige and Skoog |
| GC-MS | Gas chromatography and mass spectrometry |
References
- Dearanley, J.W.D.; Perotto, S.; Selosse, M.-A. Structure and development of orchid mycorrhizas. In Molecular Mycorrhizal Symbiosis; Martin, F., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2016; pp. 63–86. [Google Scholar]
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Tesitelova, T.; Tesitel, J.; Rihova, G.; Jersakova, J.; Selosse, M.-A. Symbiotic germination capability of four Epipactis species (Orchidaceae) is broader than expected from adult ecology. Am. J. Bot. 2012, 99, 1020–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.A.; Martos, F.; Perry, B.A.; Padamsee, M.; Roy, M.; Pailler, T. Saprotrophic fungal mycorrhizal symbionts in achlorophyllous orchids: Finding treasures among the “molecular scraps”? Plant Signal. Behav. 2010, 5, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.-A.; Schneider-Maunoury, L.; Martos, F. Time to re-think fungal ecological niches? New Phytol. 2018, 217, 968–972. [Google Scholar] [CrossRef] [PubMed]
- Vujanovic, V.; Vujanovic, J. Mycovitality and mycoheterotrophy: Where lies dormancy in terrestrial orchid and plants with minute seeds. Symbiosis 2007, 44, 93–99. [Google Scholar]
- Vujanovic, V.; St-Arnaud, M.; Barabe, D.; Thibeault, G. Viability testing of orchid seed and the promotion of colouration and germination. Ann. Bot. 2000, 86, 79–86. [Google Scholar] [CrossRef]
- Zhang, F.S.; Lv, Y.L.; Zhao, Y.; Guo, S.X. Promoting role of an endophyte on the growth and contents of kinsenosides and flavonoids of Anoectochilus formosanus Hayata, a rare and threatened medicinal Orchidaceae plant. J. Zhejiang Univ. Sci. B 2013, 14, 785–792. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Botina, S.G.; Netrusov, A.I. Bacteria associated with orchid roots and microbial production of auxin. Microbiol. Res. 2007, 162, 69–76. [Google Scholar] [CrossRef]
- Cameron, K.M.; Chase, M.W.; Whitten, W.M.; Kores, P.J.; Jarrell, D.C.; Albert, V.A.; Goldman, D.H. A phylogenetic analysis of the Orchidaceae: Evidence from rbcL nucleotide sequences. Am. J. Bot. 1999, 86, 208–224. [Google Scholar] [CrossRef] [PubMed]
- Rokaya, M.B.; Raskoti, B.B.; Timsina, B.; Münzbergová, Z. An annotated checklist of the orchids of Nepal. Nordic J. Bot. 2013, 31, 511–550. [Google Scholar] [CrossRef]
- Rajbhandari, K.R. Orchids of Nepal: Status, threat and conservation. In Proceedings of the National Workshop on NTFP/maps Sector Action Plant Development: Orchid, Department of Plant Resources, Ministry of Forest and Soil Conservation and Central Department of Botany, Tribhuvan University, Kathmandu, Nepal, 9 July 2014; pp. 1–40. [Google Scholar]
- Xing, Y.-M.; Chen, J.; Cui, J.-L.; Chen, X.-M.; Guo, S.-X. Antimicrobial Activity and Biodiversity of Endophytic Fungi in Dendrobium devonianum and Dendrobium thyrsiflorum from Vietnam. Curr. Microbiol. 2011, 62, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- Pant, B.; Paudel, M.; Chand, M.B.; Wagner, S.H. Treasure Troves of Orchids in Central Nepal; Central Department of Botany, Tribhuvan University: Kirtipur, Nepal, 2016. [Google Scholar]
- Baek, J.M.; Kim, J.-Y.; Ahn, S.-J.; Cheon, Y.-H.; Yang, M.; Oh, J.; Choi, M.K. Dendrobium moniliforme Exerts Inhibitory Effects on Both Receptor Activator of Nuclear Factor Kappa-B Bone Erosion in Vivo. Molecules 2016, 21, 295. [Google Scholar] [CrossRef] [PubMed]
- Brick, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid in situ assay for indole acetic acid production by bacteria immobilized on nitrocellulose membrane. Appl. Environ. Microb. 2004, 57, 535–538. [Google Scholar]
- Pandey, M.; Sharma, J.; Taylor, D.L.; Yadon, V.L. A narrowly endemic photosynthetic orchid is non-specific in its mycorrhizal associations. Mol. Ecol. 2013, 22, 2341–2354. [Google Scholar] [CrossRef] [PubMed]
- Lofgren, L.A.; LeBlanc, N.R.; Certano, A.K.; Nachtigall, J.; LaBine, K.M.; Riddle, J.; Broz, K.; Dong, Y.; Bethan, B.; Kafer, C.W. Fusarium graminearum: Pathogen or endophyte of North American grasses? New Phytol. 2018, 217, 1203–1212. [Google Scholar] [CrossRef]
- Gonzaga, L.L.; Costa, L.E.; Santos, T.T.; Araújo, E.F.; Queiroz, M.V. Endophytic fungi from the genus Colletotrichum are abundant in the Phaseolus vulgaris and have high genetic diversity. J. Appl. Microbiol. 2015, 118, 485–496. [Google Scholar] [CrossRef]
- Davis, E.C.; Franklin, J.B.; Shaw, A.J.; Vilgalys, R. Endophytic Xylaria (Xylariaceae) among liverworts and angiosperms: Phylogenetics, distribution, and symbiosis. Am. J. Bot. 2003, 90, 1661–1667. [Google Scholar] [CrossRef]
- Wang, X.; Radwan, M.M.; Taráwneh, A.H.; Gao, J.; Wedge, D.E.; Rosa, L.H.; Cutler, H.G.; Cutler, S.J. Antifungal activity against plant pathogens of metabolites from the endophytic fungus Cladosporium cladosporioides. J. Agric. Food Chem. 2013, 61, 4551–4555. [Google Scholar] [CrossRef]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Narayan, O.P.; Verma, N.; Singh, A.K.; Oelmüller, R.; Kumar, M.; Prasad, D.; Kapoor, R.; Dua, M.; Johri, A.K. Antioxidant enzymes in chickpea colonized by Piriformospora indica participate in defense against the pathogen Botrytis cinerea. Sci. Rep. 2017, 7, 13553. [Google Scholar] [CrossRef] [PubMed]
- Dolatabad, H.K.; Goltapeh, E.M.; Safari, M.; Golafaie, T.P. Potential effect of Piriformospora indica on plant growth and essential oil yield in Mentha piperita. Plant Pathol. Quarantine 2017, 7, 96–104. [Google Scholar] [CrossRef]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltruschat, H.; Fodor, J.; Harrach, B.D.; Niemczyk, E.; Barna, B.; Gullner, G.; Janeczko, A.; Kogel, K.-H.; Schäfer, P.; Schwarczinger, I.; et al. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. New Phytol. 2008, 180, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; Riov, J.; Sharon, A. Indole-3-Acetic Acid Biosynthesis in Colletotrichum gloeosporioides f. sp. aeschynomene. Appl. Environ. Microbiol. 1998, 64, 5030–5032. [Google Scholar] [PubMed]
- Peters, N.K.; Verma, D.P.S. Phenolic compounds as regulators of gene expression in plant-microbe interactions. Mol. Plant-Microbe Interact. 1990, 3, 4–8. [Google Scholar] [CrossRef]
- Gallie, D.R. L-Ascorbic Acid: A Multifunctional Molecule Supporting Plant Growth and Development. Scientifica 2013, 2013, 24. [Google Scholar] [CrossRef]
- Noctor, G.; Veljovic-Jovanovic, S.; Foyer, C.H.; Grace, S. Peroxide processing in photosynthesis: Antioxidant coupling and redox signaling. Philos. Trans. R. Soc. B 2000, 355, 1465–1475. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Van, L.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef] [Green Version]
- Selosse, M.-A.; Bessis, A.; Pozo, M.-J. Microbial priming of plant and animal immunity: Symbionts as developmental signals. Trends Microbiol. 2014, 22, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.L.; Al-harrasi, A.; Al-rawahi, A.; Alzfarsi, Z.; Al, A.; Waqas, M.; Shin, J. Endophytic fungi from frankincense tree improves host growth and produces extracellular enzymes and Indole Acetic Acid. PLoS ONE 2016, 11, e0158207. [Google Scholar] [CrossRef] [PubMed]
- Séne, S.; Avril, R.; Chaintreuil, C.; Geoffroy, A.; Ndiaye, C.; Dieddhiou, A.G.; Sadio, O. Ectomycorrhizal fungal communities of Coccoloba uvifera L. mature trees and seedlings in the neotropical coastal forests of Guadeloupe (Lesser Antilles). Mycorrhizal 2015, 25, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T. Plant propagation through tissue culture. Ann. Rev. Plant Physiol. 1974, 25, 135–166. [Google Scholar] [CrossRef]
- Baldi, A.; Srivastava, A.K.; Bisaria, V.S. Fungal Elicitors for Enhanced Production of Secondary Metabolites in Plant Cell Suspension Cultures. In Symbiotic Fungi; Varma, A., Kharkwal, A.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 373–380. [Google Scholar]
- Parry, C.; Blonquist, J.M.; Bugbee, B. The optical/absolute chlorophyll relationship. Plant Cell Environ. 2014, 37, 2508–2520. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphotype | Tentative Affiliation | No. Isolates | Query Coverage | Percentage of Identity | Accession Code |
|---|---|---|---|---|---|
| R6 | Hypoxylon sp. | 4 | 99% | 88% | MH532318 |
| R10 | Colletotrichum alatae | 7 | 100% | 100% | MH532312 |
| R11 | Leptosphaerulina chartarum | 3 | 100% | 100% | MH532310 |
| R12 | Fusarium sp. | 6 | 100% | 100% | MH532313 |
| R13 | Fusarium sp. | 7 | 100% | 84% | MH532317 |
| R14 | Cladosporium tenuissimum | 2 | 100% | 100% | MH532316 |
| R16 | Fusarium equiseti | 5 | 100% | 100% | MH532315 |
| R17 | Cylindrocarpon sp. | 4 | 95% | 100% | MH532314 |
| R19 | Trichoderma harzianum | 6 | 100% | 100% | MH532311 |
| Peak | Retention Time | Area% | Compound Identified | Fungal Extract | Base m/z |
|---|---|---|---|---|---|
| 1 | 5.381 | 1.24 | 7-Methyl-1 H-indole | R11 | 130.15 |
| 2 | 6.247 | 37.30 | Phenol, 2,4-bis (1,1-dimethylethyl) | R11 | 191.15 |
| 3 | 6.289 | 0.47 | Phenol, 2,4-bis (1,1-dimethylethyl)- | R13 | 191.15 |
| 4 | 6.840 | 4.91 | 3-Octadecene, (E)- | R13 | 55.05 |
| 5 | 7.916 | 0.38 | Benzoic acid, 2,4-dihydroxy-3,6-dimethyl-, methyl | R13 | 136.10 |
| 6 | 8.336 | 33.86 | 9-Eicosene, (E)- | R13 | 55.05 |
| 7 | 9.221 | 2.61 | Hexadecanoic acid, methyl ester | R11 | 74.05 |
| 8 | 9.253 | 4.37 | Hexadecanoic acid, methyl ester | R13 | 74.05 |
| 9 | 9.521 | 3.61 | l-(+)-Ascorbic acid 2,6-dihexadecanoate | R13 | 43.10 |
| 10 | 9.590 | 1.98 | Dibutyl phthalate | R13 | 149.05 |
| 11 | 9.696 | 29.59 | 3-Eicosene, (E)- | R11 | 43.10 |
| 12 | 10.383 | 4.00 | 10-Octadecenoic acid, methyl ester | R11 | 55.10 |
| 13 | 10.531 | 2.35 | Hexadecanoic acid, 15-methyl-, methyl ester | R11 | 74.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, S.; Shrestha, R.; Maharjan, S.; Selosse, M.-A.; Pant, B. Isolation and Characterization of Plant Growth-Promoting Endophytic Fungi from the Roots of Dendrobium moniliforme. Plants 2019, 8, 5. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8010005
Shah S, Shrestha R, Maharjan S, Selosse M-A, Pant B. Isolation and Characterization of Plant Growth-Promoting Endophytic Fungi from the Roots of Dendrobium moniliforme. Plants. 2019; 8(1):5. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8010005
Chicago/Turabian StyleShah, Sujit, Roshani Shrestha, Sabitri Maharjan, Marc-Andre Selosse, and Bijaya Pant. 2019. "Isolation and Characterization of Plant Growth-Promoting Endophytic Fungi from the Roots of Dendrobium moniliforme" Plants 8, no. 1: 5. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8010005