Comparative Proteomics Reveals Cold Acclimation Machinery Through Enhanced Carbohydrate and Amino Acid Metabolism in Wucai (Brassica Campestris L.)

 , and
, and
Abstract
:1. Introduction
2. Results
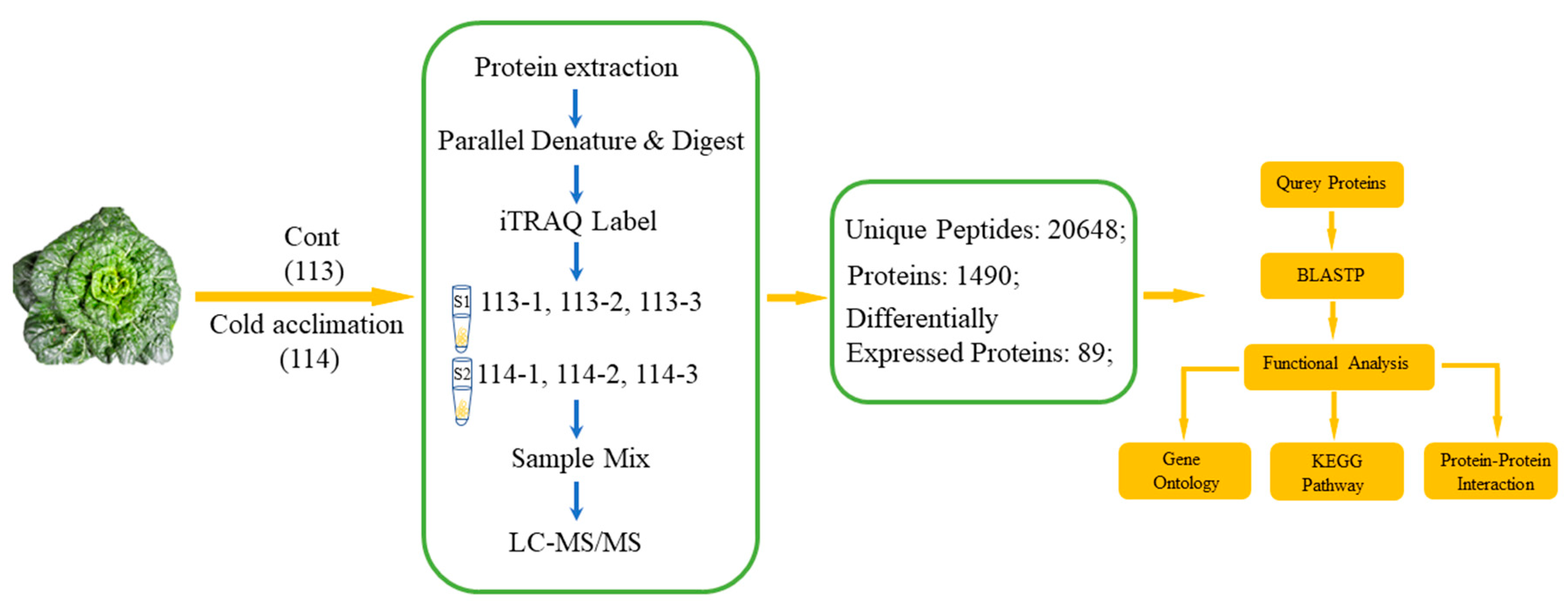
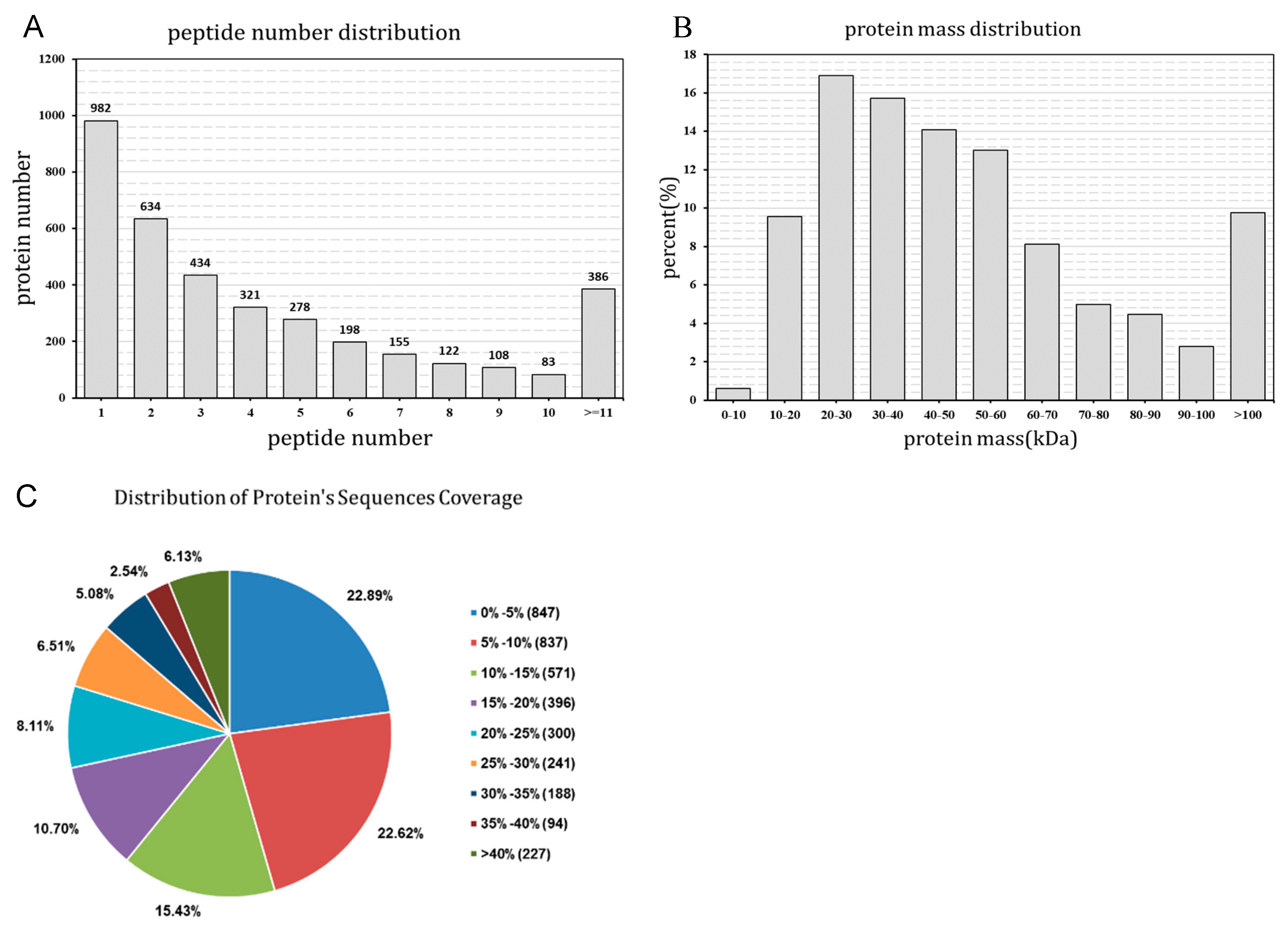
2.1. Primary Data Analysis and Protein Identification
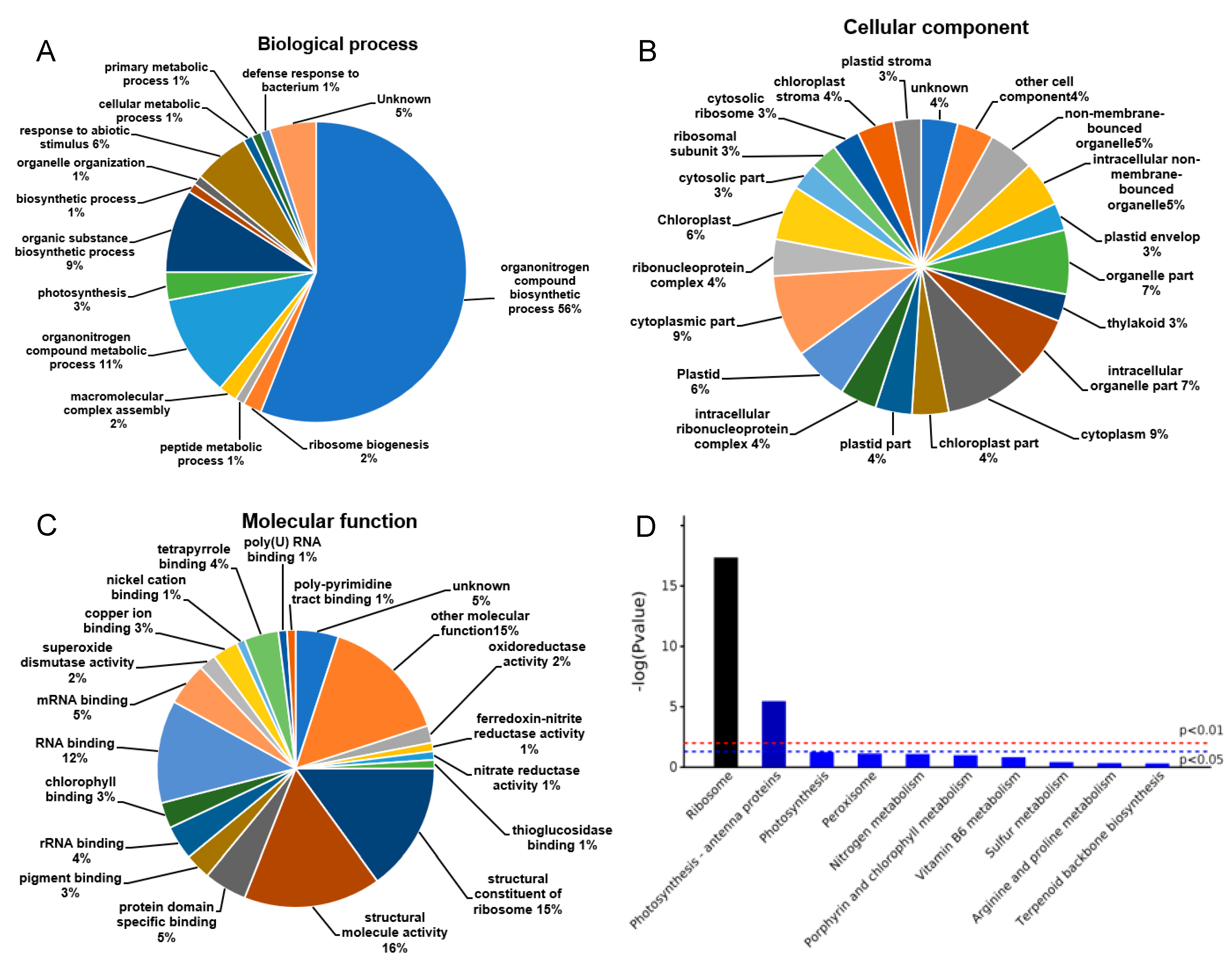
2.2. Functional Cataloging of DEPs under Low Temperature
2.3. DEPs Involved in Different Metabolism Processes
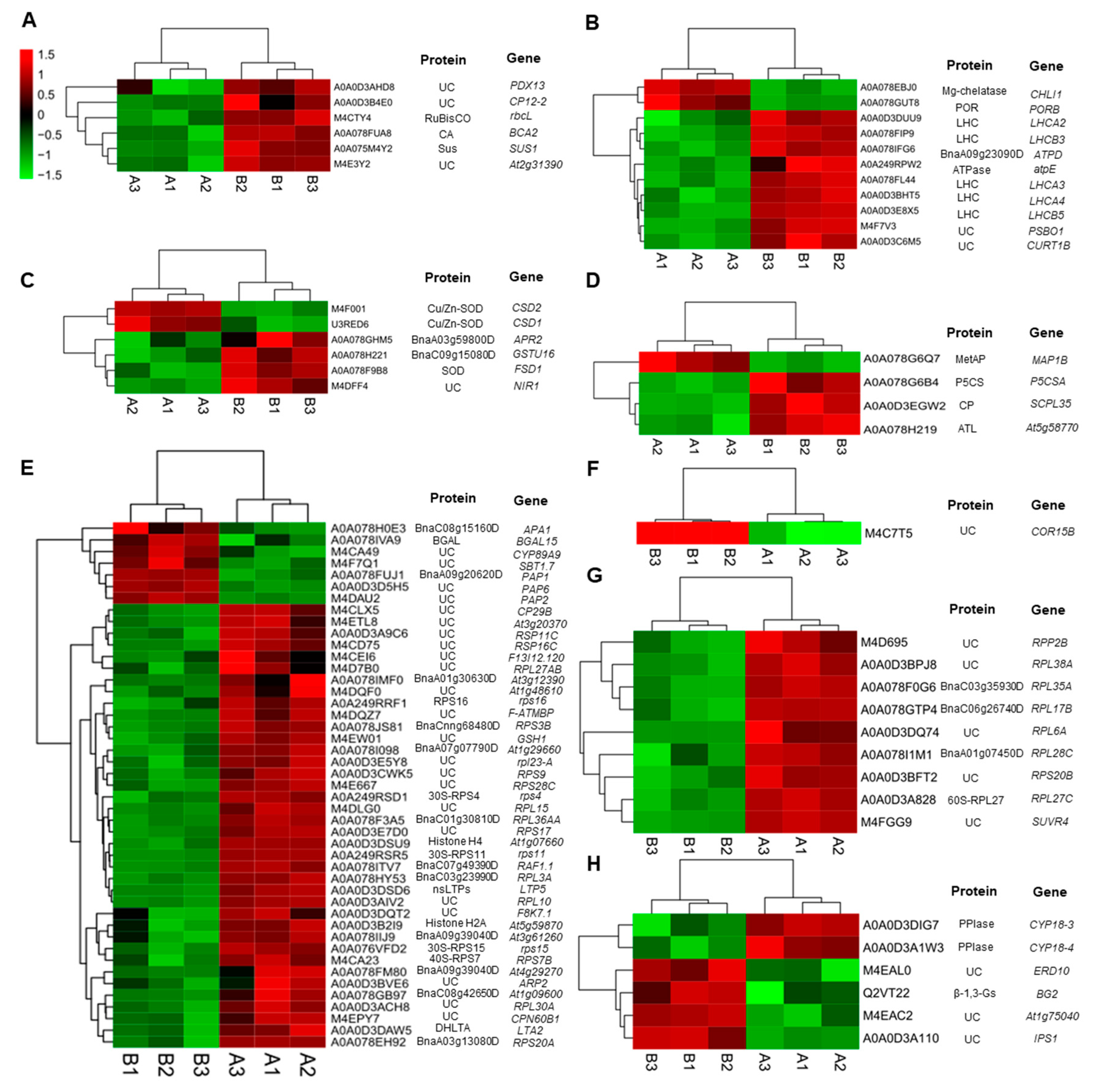
2.3.1. DEPs Involved in Carbohydrate Metabolism
2.3.2. DEPs Involved in Photosynthesis and Energy Metabolism
2.3.3. DEPs Involved in Oxidative Defense
2.3.4. DEPs Involved in Amino Acid Metabolism
2.3.5. Metabolism Related Proteins
2.3.6. Cold Regulated Protein
2.3.7. Proteins Involved in Methylation
2.3.8. Proteins Involved in Signal Transduction
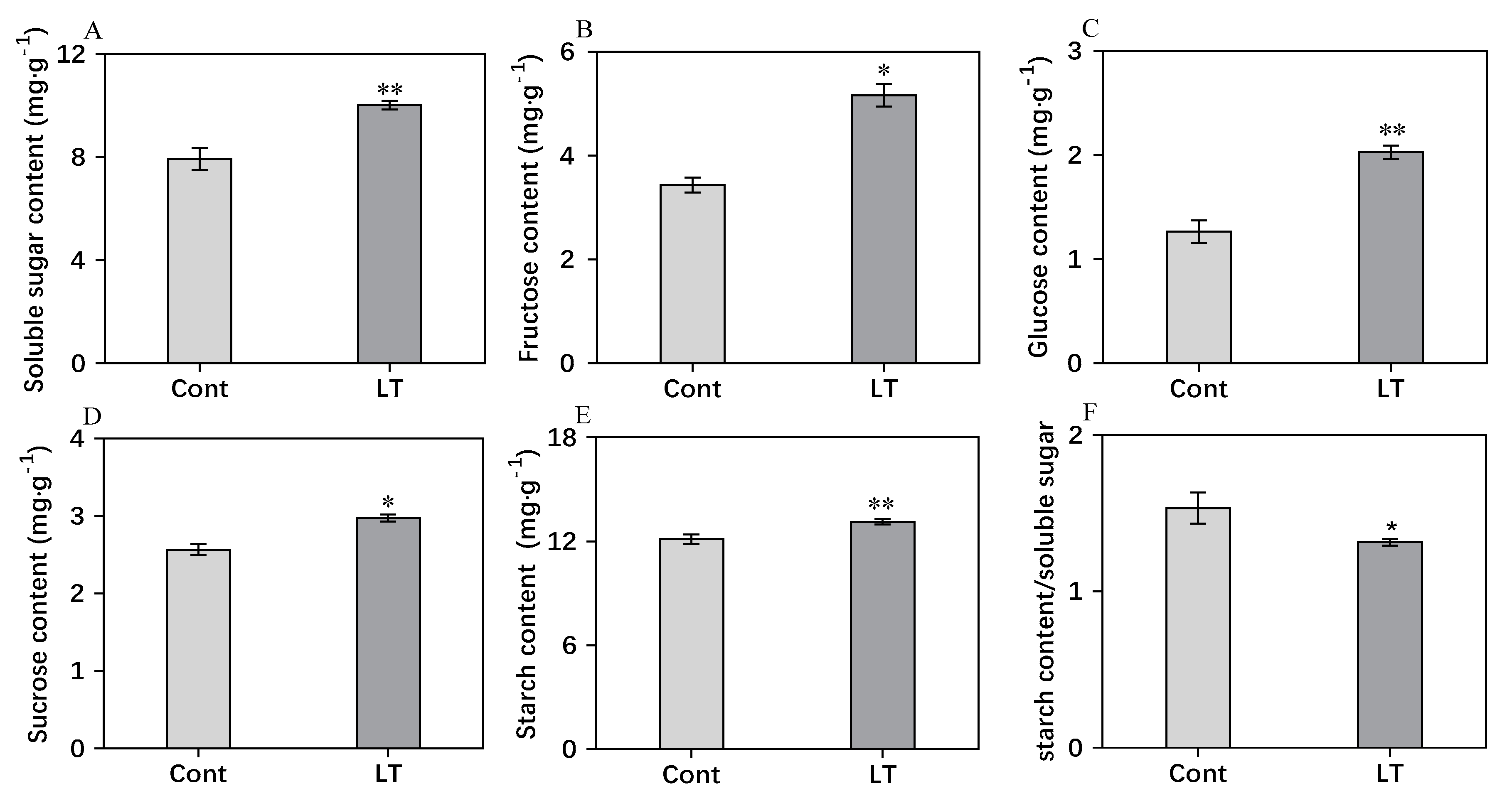
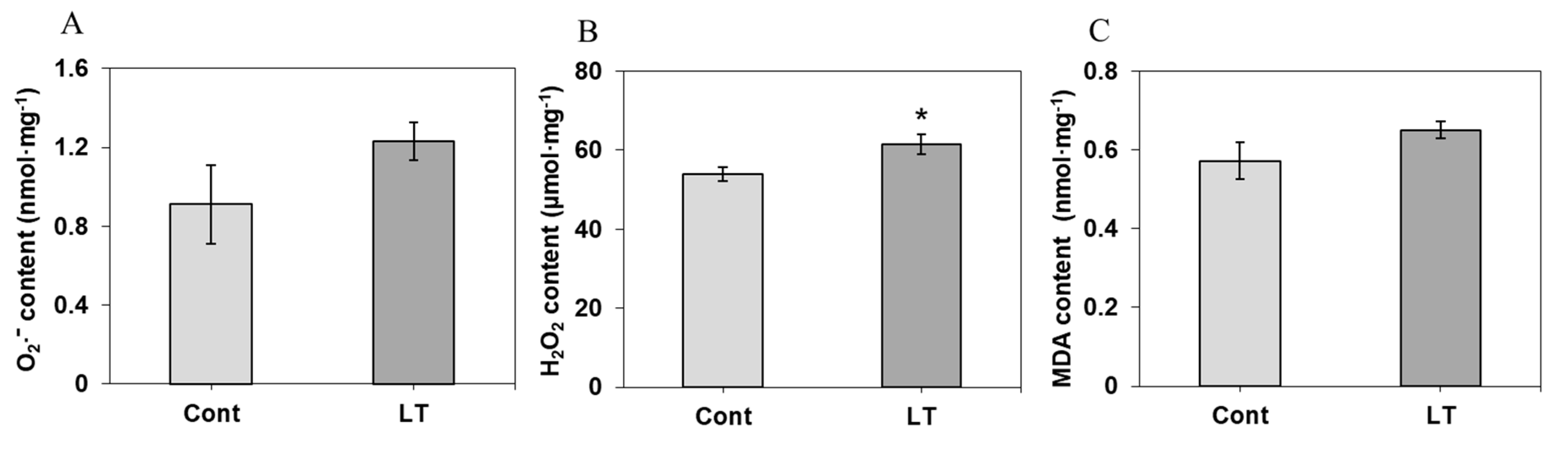
2.4. Analysis of Carbohydrate Content, O2− Content, H2O2 Content, and MDA Content
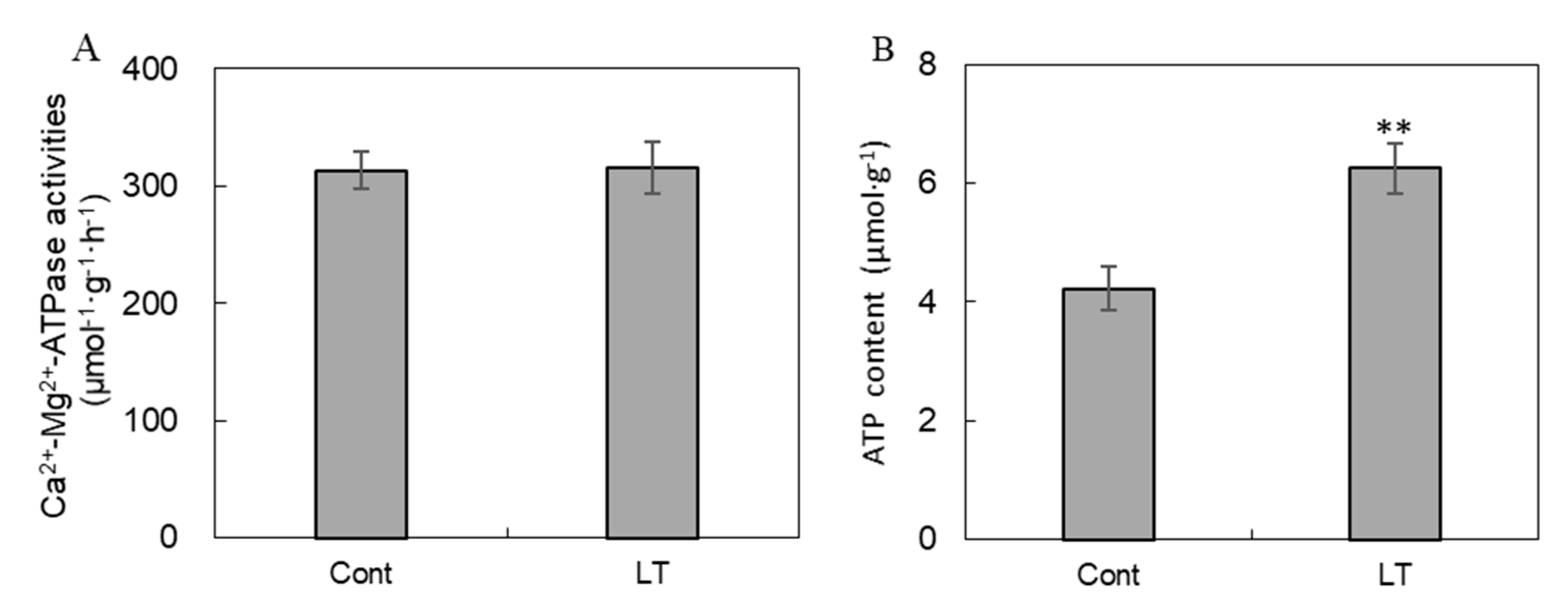
2.5. Analysis of ATPase Activity and ATP Content
2.6. Analysis of Chlorophyll (Chl) Content and Chlorophyll Fluorescence Parameters
2.7. Analysis of Free Amino Acids
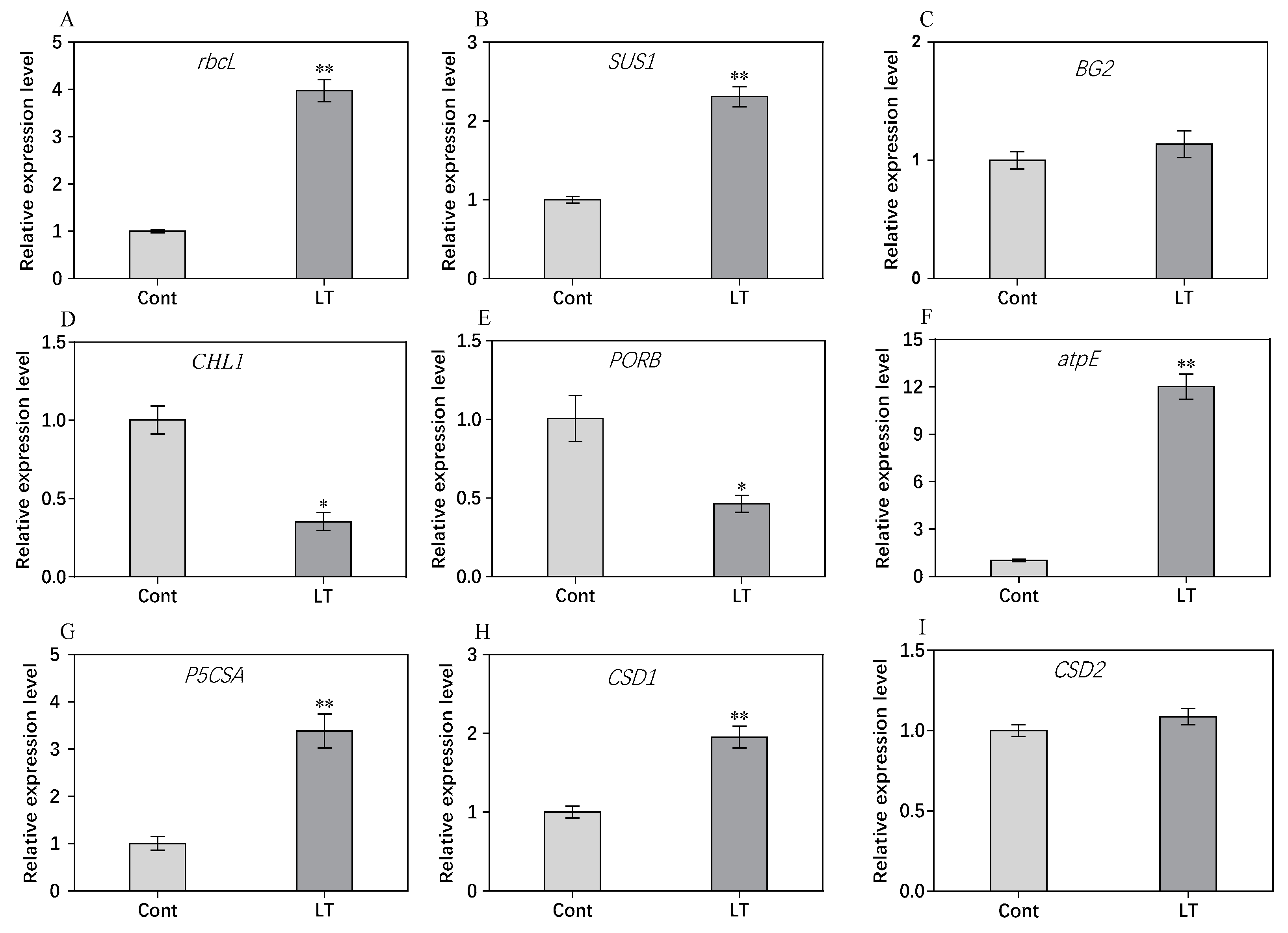
2.8. RT-PCR Analysis of Genes for Some DEPs
3. Discussion
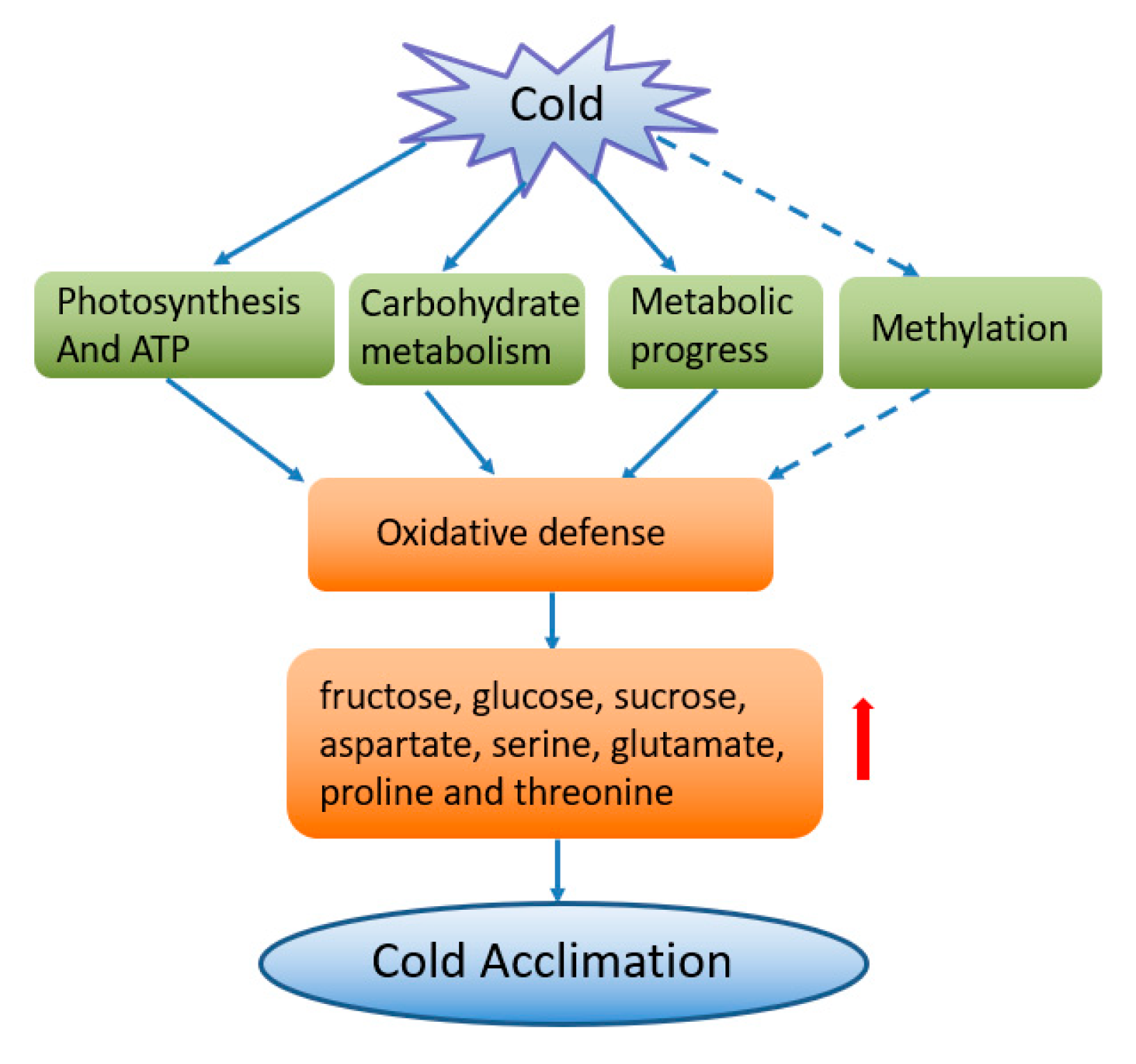
4. Conclusions
5. Materials and Methods
5.1. Growth Condition
5.2. Protein Preparation and iTRAQ Labeling
5.3. iTRAQ Analysis and Bioinformatic Analysis of Proteins
5.4. Analysis of Carbohydrate Content, H2O2 Content, O2− Produce Rate, and MDA Content
5.5. Analysis of ATPase Activity and ATP Content
5.6. Analysis of Chlorophyll Fluorescence Parameters
5.7. Determination of Free Amino Acids in Leaves
5.8. Analysis of the Expression Levels of Differentially Expressed Protein Related Genes
5.9. Statistical Treatment
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Anderson, J.A.; Taliaferro, C.M. Freeze tolerance of seed-producing turf bermudagrasses. Crop. Sci. 2002, 42, 190–192. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Pocock, T.H.; Hurry, V.; Savitch, L.V.; Huner, N.P.A. Susceptibility to low-temperature photoinhibition and the acquisition of freezing tolerance in winter and spring wheat: The role of growth temperature and irradiance. Physiol. Plant 2001, 113, 499–506. [Google Scholar] [CrossRef]
- Huang, B.R.; DaCosta, M.; Jiang, Y.W. Research advances in mechanisms of turfgrass tolerance to abiotic stresses: From physiology to molecular biology. Crit. Rev. Plant Sci. 2014, 33, 141–189. [Google Scholar] [CrossRef]
- Wang, K.; Bai, Z.; Liang, Q.; Liu, Q.; Zhang, L.; Pan, Y.; Liu, G.; Jiang, B.; Zhang, F.; Jia, Y. Transcriptome analysis of chrysanthemum (Dendranthema grandiflorum) in response to low temperature stress. BMC Genom. 2018, 19, 319. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, S.; Zhu, S.; Chen, G.; Liu, F.; Zou, M.; Wang, C. Comparative response of two wucai (Brassica campestris L.) genotype to heat stress on antioxidative system and cell ultrastructure in root. Acta Physiol. Plant 2016, 38, 223. [Google Scholar] [CrossRef]
- Yuan, L.; Yuan, Y.; Liu, S.; Wang, J.; Zhu, S.; Chen, G.; Hou, J.; Wang, C. Influence of high temperature on photosynthesis, antioxidative capacity of chloroplast, and carbon assimilation among heat-tolerant and heat-susceptible genotypes of non-heading Chinese cabbage. Hortscience 2017, 52, 1–7. [Google Scholar] [CrossRef]
- Hannah, M.A.; Heyer, A.G.; Hincha, D.K. A global survey of gene regulation during cold acclimation in Arabidopsis thaliana. PLoS Genet. 2005, 1, e26. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.K.; Sunkar, R. Gene regulation during cold stress acclimation in plants. Mol. Biol. 2010, 639, 39–55. [Google Scholar]
- Zhu, J.; Dong, C.; Zhu, J. Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr. Opin. Plant Biol. 2007, 10, 290–295. [Google Scholar] [CrossRef]
- Lissarre, M.; Ohta, M.; Sato, A.; Miura, K. Cold-responsive gene regulation during cold acclimation in plants. Plant Signal. Behav. 2010, 5, 948–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willick, I.R.; Takahashi, D.; Fowler, D.B.; Uemura, M.; Tanino, K.K. Tissue-specific changes in apoplastic proteins and cell wall structure during cold acclimation of winter wheat crowns. J. Exp. Bot. 2018, 69, 1221–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltész, A.; Smedley, M.; Vashegyi, I.; Galiba, G.; Harwood, W.; Vágújfalvi, A. Transgenic barley lines prove the involvement of TaCBF14 and TaCBF15 in the cold acclimation process and in frost tolerance. J. Exp. Bot. 2013, 64, 1849–1862. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant. Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Barrero-Gil, J.; Salinas, J. Post-translational regulation of cold acclimation response. Plant Sci. 2013, 205, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vitámvás, P.; Urban, M.O.; Prášil, I.T.; Renaut, J. Plant abiotic stress proteomics: The major factors determining alterations in cellular proteome. Front. Plant Sci. 2018, 9, 122. [Google Scholar] [CrossRef]
- Kupriyanova, E.; Pronina, N.; Los, D. Carbonic anhydrase—A universal enzyme of the carbon life. Photosynthetica 2017, 55, 3–19. [Google Scholar] [CrossRef]
- Jiang, C.; Tholen, D.; Mercedes Xu, J.J.; Xin, C.P.; Zhang, H.; Zhu, X.; Zhao, Y. Increases expression of mitochondria-localized carbonic anhydrase activity resulted in an increased biomass accumulation in Arabidopsis thaliana. J. Plant Biol. 2014, 57, 366–374. [Google Scholar] [CrossRef]
- Ramon, M.; Rolland, F.; Sheen, J. Sugar sensing and signaling in Arabidopsis. In The Arabidopsis Book; ASPB: Rockville, MD, USA, 2008; pp. 115–117. [Google Scholar]
- Burbulis, N.; Jonytienė, V.; Kuprienė, R.; Blinstrubienė, A.; Vytautas Liakas, V. Biochemical and physiological factors related to cold de-acclimation and by trehalose-6-phosphate. Plant Physiol. 2012, 149, 1860–1871. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plant. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Hu, C.M.; Hou, X.L. Relationship of major nutrient component with low temperature tolerance in non-heading Chinese cabbage. J. Nanjing Agric. Univ. 2010, 33, 37–41. [Google Scholar]
- Jansson, S. A guide to the Lhc genes and their relatives in Arabidopsis. Trends Plant Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Hazen, S.P.; Pathan, M.S.; Sanchez, A.; Baxter, I.; Dunn, M.; Estes, B.; Chang, H.S.; Zhu, T.; Kreps, J.A.; Nguyen, H.T. Expression profiling of rice segregating for drought tolerance QTLs using a rice genome array. Funct. Integr. Genom. 2005, 5, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Baum, M.; Grando, S.; Ceccarelli, S.; Bai, G.; Li, R.; Von Korff, M.; Varshney, R.K.; Graner, A.; Valkoun, J. Differentially expressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage. J. Exp. Bot. 2009, 12, 3531–3544. [Google Scholar] [CrossRef]
- Loukehaich, R.; Wang, T.; Ouyang, B.; Zaif, K.; Li, H.; Zhang, J.; Lu, Y.; Ye, Z. SpUSP, an annexin-interacting universal stress protein, enhances drought tolerance in tomato. J. Exp. Bot. 2012, 63, 5593–5606. [Google Scholar] [CrossRef] [Green Version]
- Roose, J.L.; Wegener, K.M.; Pakrasi, H. The extrinsic proteins of photosystem II. Photosynth. Res. 2007, 3, 369–387. [Google Scholar] [CrossRef]
- Debus, R.J. Amino acid residues that modulate the properties of tyrosine Y(Z) and the manganese cluster in the water oxidizing complex of photosystem II. Biochim. Biophys. Acta 2001, 1503, 164–186. [Google Scholar] [CrossRef]
- Pospíšil, P. Production of reactive oxygen species by photosystem II as a response to light and temperature stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef]
- Mathur, S.; Jajoo, A. Alterations in photochemical efficiency of photosystem II in wheat plant on hot summer day. Physiol. Mol. Biol. Plants 2014, 20, 527–531. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Shan, Y.; Kochian, L.; Strasser, R.J.; Chen, G. Photochemical properties in flag leaves of a super-high-yielding hybrid and a traditional hybrid rice (Oryza sativa L.) probed by chlorophyll a fluorescence transient. Photosynth. Res. 2015, 126, 275–284. [Google Scholar] [CrossRef]
- Stirbet, A.; Govindjee, G. On the relation between the kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and application of the OJIP fluorescence transient. J. Photoch. Photobio. B 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Strasserf, R.J.; Srivastava, A. Polyphasic chlorophyll-a fluorescence transients in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Van Heerden, P.D.; Tsimilli-Michael, M.; Krüger, G.H.; Strasser, R.J. Dark chilling effects on soybean genotypes during vegetative development: Parallel studies of CO2 assimilation, chlorophyll-a fluorescence kinetics O-J-I-P and nitrogen fixation. Physiol. Plant 2003, 117, 476–491. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.M.F. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Kaur, G.; Kumar, S.; Thakur, P.; Malik, J.A.; Bhandhari, K.; Sharma, K.; Nayyar, H. Involvement of proline in response of chickpea (Cicer arietinum L.) to chilling stress at reproductive stage. Sci. Hortic. 2011, 128, 174–181. [Google Scholar] [CrossRef]
- Yang, S.; Lan, S.; Deng, F.; Gong, M. Effects of calcium and calmodulin antagonists on chilling stress-induced proline accumulation in Jatropha curcas L. J. Plant Grow. Regul. 2016, 35, 815–826. [Google Scholar] [CrossRef]
- Vítámvás, P.; Prášil, I.T.; Kosová, K.; Planchon, S.; Renaut, J. Analysis of proteome and frost tolerance in chromosome 5A and 5B reciprocal substitution lines between two winter wheats during long-term cold acclimation. Proteomics 2012, 12, 68–85. [Google Scholar] [CrossRef]
- Araújo, W.L.; Tohge, T.; Ishizaki, K.; Leaver, C.J.; Fernie, A.R. Protein degradation-an alternative respiratory substrate for stressed plants. Trends Plant Sci. 2011, 16, 489–498. [Google Scholar] [CrossRef]
- Snowden, C.J.; Thomas, B.; Baxter, C.J.; Smith, J.A.C.; Sweetlove, L. A tonoplast Glu/Asp/GABA exchanger that affects tomato fruit amino acid composition. Plant J. 2015, 81, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Marček, T.; Tkalec, M.; Vidaković-Cifrek, Ž.; Ježić, M.; Ćurković-Perica, M. Effect of NaCl stress on dihaploid tobacco lines tolerant to Potato virus Y. Acta Physiol. Plant 2014, 36, 1739–1747. [Google Scholar] [CrossRef]
- Dietz, K.J.; Jacob, S.; Oelze, M.L.; Laxa, M.; Tognetti, V.; Marina, S.; Miranda, N.D.; Baier, M.; Finkemeier, I. The function of peroxiredoxins in plant organelle redox metabolism. J. Exp. Bot. 2006, 57, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- Ferreyra, M.L.F.; Pezza, A.; Biarc, J.; Burlingame, A.L.; Casati, P. Plant L10 ribosomal proteins have different roles during development and translation under ultraviolet-B stress. Plant Physiol. 2010, 153, 1878–1894. [Google Scholar] [CrossRef] [PubMed]
- Hulm, J.L.; McIntosh, K.B. Variation in transcript abundance among the four members of the Arabidopsis thaliana RIBOSOMAL PROTEIN S15a gene family. Plant Sci. 2005, 169, 267–278. [Google Scholar] [CrossRef]
- Parankusam, S.; Bhatnagar-Mathur, P.; Sharma, K.K. Heat responsive proteome changes reveal molecular mechanisms underlying heat tolerance in chickpea. Environ. Exp. Bot. 2017, 141, 132–144. [Google Scholar] [CrossRef]
- Wu, Z.; Burns, J.K. A β-galactosidase gene is expressed during mature fruit abscission of Valencia orange (Citrus sinensis). J. Exp. Bot. 2004, 55, 1483–1490. [Google Scholar] [CrossRef]
- Chandrasekar, B.; Van der Hoorn, R.A.L. Beta galactosidases in Arabidopsis and tomato—A mini review. Biochem. Soc. Trans. 2016, 44, 150–158. [Google Scholar] [CrossRef]
- Thalhammer, A.; Hundertmark, M.; Popova, A.V.; Seckler, R.; Hincha, D.K. Interaction of two intrinsically disordered plant stress proteins (COR15A and COR15B) with lipid membranes in the dry state. BBA-Biomembranes 2010, 1798, 1812–1820. [Google Scholar] [CrossRef] [Green Version]
- Renaut, J.; Hausman, J.F.; Wisniewski, M. Proteomics and low temperature studies: Bridging the gap between gene expression and metabolism. Physiol. Plant 2006, 126, 97–109. [Google Scholar] [CrossRef]
- Hoffmann-Sommergruber, K. Plant allergens and pathogenesis-related proteins. What do they have in common? Int. Arch. Allergy Immunol. 2000, 122, 155–166. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plant 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Fu, R.; Zhang, M.; Ding, Z.; Chang, L.; Zhu, X.; Wang, Y.; Fan, B.; Ye, W.; Yuan, Y. Analysis of methylation-sensitive amplified polymorphism in different cotton accessions under salt stress based on capillary electrophoresis. Genes Genom. 2015, 37, 713–724. [Google Scholar] [CrossRef]
- Fan, H.H.; Wei, J.; Li, T.C.; Li, Z.P.; Guo, N.; Cai, Y.P.; Lin, Y. DNA methylation alterations of upland cotton (Gossypium hirsutum) in response to cold stress. Acta. Physiol. Plant 2013, 35, 2445–2453. [Google Scholar] [CrossRef]
- Liu, T.; Li, Y.; Duan, W.; Huang, F.; Hou, X. Cold acclimation alters DNA methylation patterns and confers tolerance to heat and increases growth rate in Brassica rapa. J. Exp. Bot. 2017, 68, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Thorstensen, T.; Fischer, A.; Sandvik, S.V.; Johnsen, S.S.; Grini, P.E.; Reuter, G.; Aalen, R.B. The Arabidopsis SUVR4 protein is a nucleolar histone methyltransferase with preference for monomethylated H3K9. Nucleic Acids Res. 2006, 34, 5461–5470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, L.R.; Shi, C.N.; Zhao, L.; Cui, D.Q.; Chen, F. Identification of proteins using iTRAQ and virus-induced gene silencing reveals three bread wheat proteins involved in the response to combined osmotic-cold stress. J. Proteome Res. 2018, 17, 2256–2281. [Google Scholar] [CrossRef]
- Yang, L.; Qi, Y.; Lu, Y.; Guo, P.; Sang, W.; Feng, H.; Zhang, H.; Chen, L. iTRAQ protein profile analysis of Citrus sinensis roots in response to long-term boron-deficiency. J. Proteom. 2013, 93, 179–206. [Google Scholar] [CrossRef]
- Buysse, J.; Merckx, R. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar] [CrossRef]
- Hu, Z.; Weijian, L.; Yali, F.; Huiquan, L. Gibberellic acid enhances postharvest toon sprout tolerance to chilling stress by increasing the antioxidant capacity during the short-term cold storage. Sci. Horticult. 2018, 237, 184–191. [Google Scholar] [CrossRef]
- Yao, R.; Fang, S.; Shang, X.; Yang, W. Cytochemical localization of ATPase and sub-cellular variation in mesophyll cell of Cyclocarya paliurus seedlings under iso-osmotic stress and calcium regulation. J. For. Res. 2009, 20, 343–348. [Google Scholar] [CrossRef]
- Stewart, J.M.D.; Guinn, G. Chilling injury and changes in adenosine triphosphate of cotton seedlings. Plant Physiol. 1969, 44, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Guha, A.; Sengupta, D.; Reddy, A.R. Polyphasic chlorophyll a fluorescence kinetics and leaf protein analyses to track dynamics of photosynthetic performance in mulberry during progressive drought. J. Photochem. Photobiol. B 2013, 119, 71–83. [Google Scholar] [CrossRef]
- Aurisano, N.; Bertani, A.; Reggiani, R. Involvement of calcium and calmodulin in protein and amino acid metabolism in rice roots under anoxia. Plant Cell Phys. 1995, 36, 1087–1088. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAP | Chl Content (mg∙g−1 FW) | Chl a/b Ratio | PIabs | PItotal |
|---|---|---|---|---|
| Cont | 2.84 ± 0.05 a | 1.693 ± 0.08 a | 14.68 ± 0.07 b | 18.6 ± 0.21 b |
| LT | 2.92 ± 0.53 a | 1.701 ± 0.15 a | 15.72 ± 0.22 a | 20.70 ± 0.13 a |
| ABS/RC | TRo/RC | DIo/RC | ETo/RC | |
| Cont | 0.9484 ± 0.0157 b | 0.7648 ± 0.0043 a | 0.1607 ± 0.0011 b | 0.6273 ± 0.0140 a |
| LT | 1.0542 ± 0.0181 a | 0.7886 ± 0.0253 a | 0.1705 ± 0.0036 a | 0.5150 ± 0.0055 b |
| Amino Acid (μg∙g−1) | Cont | LT |
|---|---|---|
| Nonessential amino acids | ||
| Aspartate | 301.77 ± 5.32 b | 376.12 ± 12.65 a |
| Serine | 113.52 ± 1.74 b | 167.00 ± 5.24 a |
| Glutamate | 24.33 ± 0.66 b | 90.35 ± 3.78 a |
| Glycine | 26.70 ± 0.62 a | 24.64 ± 1.36 b |
| Alanine | 180.84 ± 3.31 a | 183.79 ± 7.14 a |
| Tyrosine | 175.18 ± 3.28 a | 119.03 ± 9.92 b |
| Histidine | 26.87 ± 0.45 a | 26.13 ±1.35 a |
| Arginine | 222.83 ± 3.64 a | 121.06 ± 2.75 b |
| Proline | 16.94 ± 1.25 b | 72.32 ± 4.88 a |
| Essential amino acids | ||
| Cysteine | 9.06 ± 0.85 a | 6.58 ± 3.32 a |
| Valine | 123.81 ± 2.80 a | 91.33 ± 13.01 b |
| Methionine | 42.01 ± 1.41 a | 19.18 ± 9.02 b |
| Threonine | 228.50 ± 3.48 b | 439.02 ± 5.08 a |
| Isoleucine | 99.21 ± 1.67 a | 76.31 ± 13.84 a |
| Leucine | 184.95 ± 3.13 a | 145.08 ± 9.68 b |
| Phenylalanine | 193.94 ± 3.11 a | 149.24 ±10.16 b |
| Lysine | 135.25 ± 2.21 a | 123.84 ± 4.47 b |
| Association No. | Gene Name | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|---|
| XM_009127097.2 | ß-actin | TGGGTTTGCTGGTGACGAT | TGCCTAGGACGACCAACAATACT |
| AY167977.1 | rbcl | TATGCCTGCTTTGACCGAGA | GCAAGATCACGTCCCTCA |
| XM_009113306.2 | SUS1 | TTGGTGGAGAGTGGAGAGAAG | CTGGTACTGAAGCTCTGCCT |
| XM_009118181.2 | BG2 | GACGACCCATACTCTTACACA | TTCCAACGACCCTCCGCCTGAT |
| XM_009133530.2 | CHLI1 | CCACAGAAATCAACTCCAC | CATTATCATCACACCACCG |
| XM_009139412.2 | PORB | TACACGGTGATGCATTTGGAC | GTCATCAAGCAACAACCTCGA |
| XM_018654732.1 | atpE | AATGCTCTGGTGGTTAAGGGT | TCAACAGGTTCCCCAAGTACA |
| XM_009143589.2 | P5CSA | AAGCAAGGTCGTTCAAG | TATTCCCACCTCAGCACCAA |
| XM_009149890.2 | CSD1 | TGCTGGCGATCTAGGAAACA | AGCCCTGAAGACCAATAATGC |
| XM_009142600.2 | CSD2 | CCGACAAAAGTGAGTGTTCGT | GGCAATAATGTTTCCCAGGTC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, L.; Xie, S.; Nie, L.; Zheng, Y.; Wang, J.; Huang, J.; Zhao, M.; Zhu, S.; Hou, J.; Chen, G.; et al. Comparative Proteomics Reveals Cold Acclimation Machinery Through Enhanced Carbohydrate and Amino Acid Metabolism in Wucai (Brassica Campestris L.). Plants 2019, 8, 474. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110474
Yuan L, Xie S, Nie L, Zheng Y, Wang J, Huang J, Zhao M, Zhu S, Hou J, Chen G, et al. Comparative Proteomics Reveals Cold Acclimation Machinery Through Enhanced Carbohydrate and Amino Acid Metabolism in Wucai (Brassica Campestris L.). Plants. 2019; 8(11):474. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110474
Chicago/Turabian StyleYuan, Lingyun, Shilei Xie, Libing Nie, Yushan Zheng, Jie Wang, Ju Huang, Mengru Zhao, Shidong Zhu, Jinfeng Hou, Guohu Chen, and et al. 2019. "Comparative Proteomics Reveals Cold Acclimation Machinery Through Enhanced Carbohydrate and Amino Acid Metabolism in Wucai (Brassica Campestris L.)" Plants 8, no. 11: 474. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8110474