Potential Effects of Awn Length Variation on Seed Yield and Components, Seed Dispersal and Germination Performance in Siberian Wildrye (Elymus sibiricus L.)

 ,
,
Abstract
:1. Introduction
2. Results
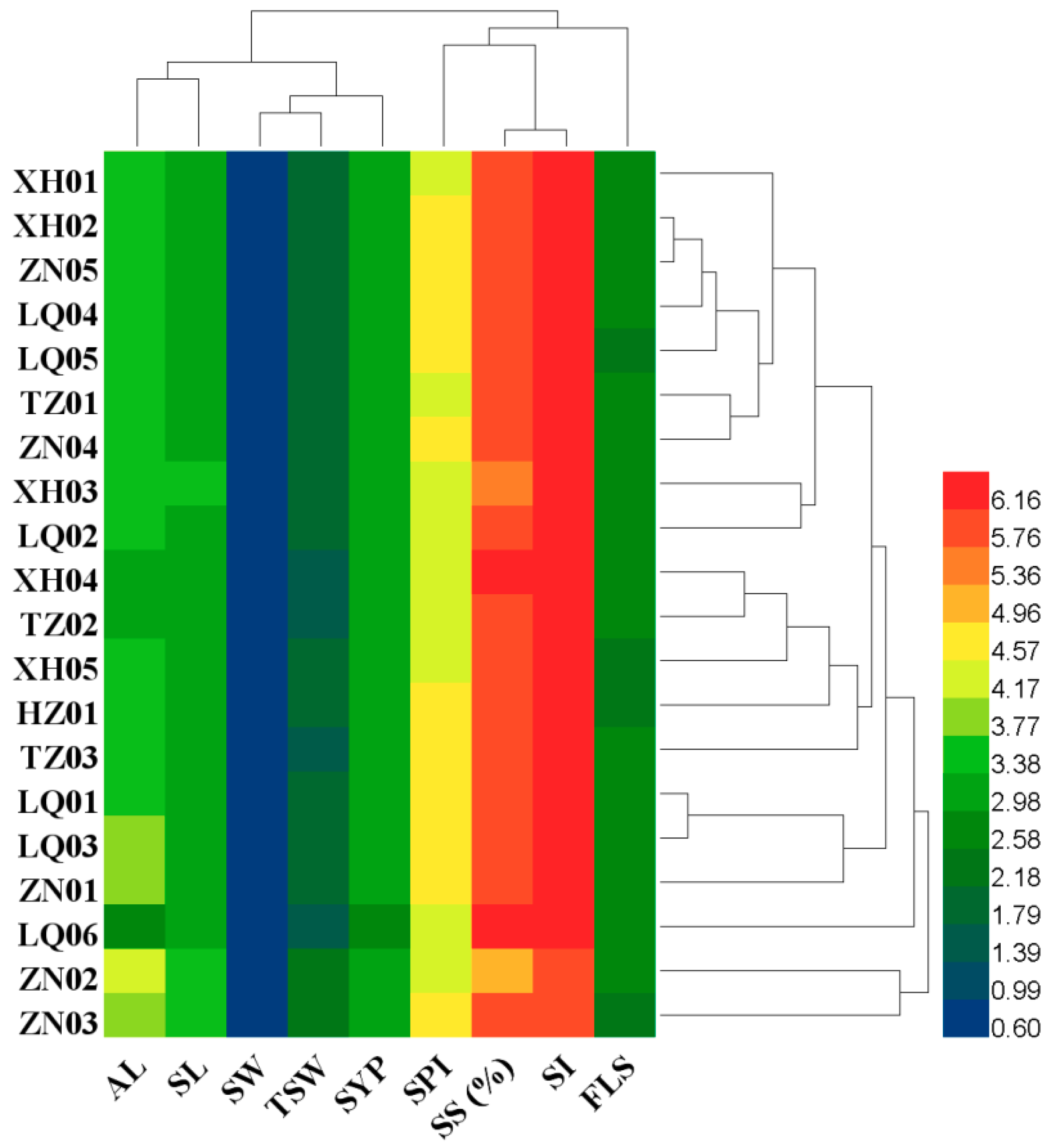
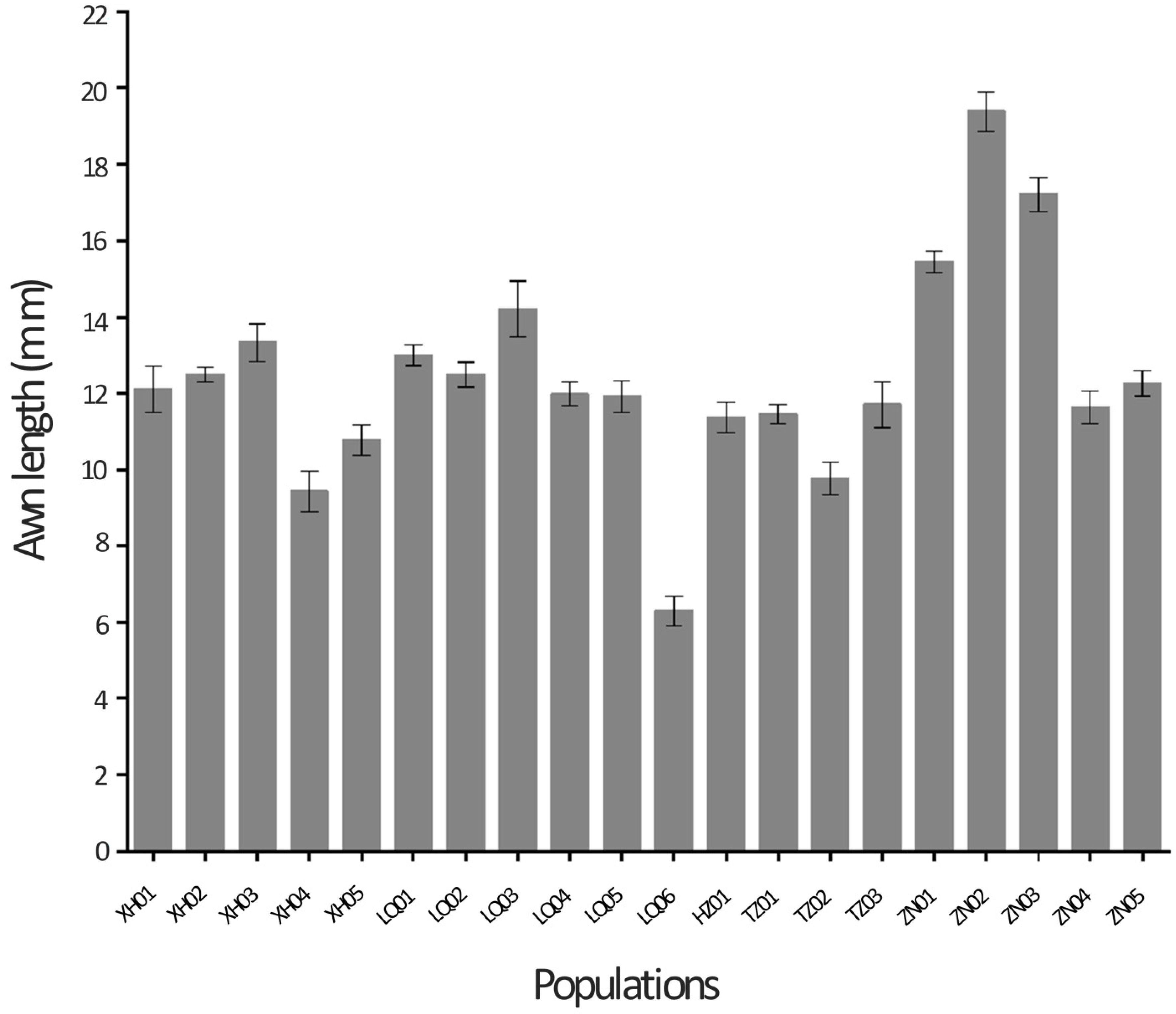
2.1. Variability in Awn Length, Yield Componentsand Seed Yield
2.2. Relationships Between Awn Length, Yield Componentsand Seed Yield
2.3. Impact of Geographical Distribution on Variation of Awn Length, Seed Yield Components and Seed Yield
2.4. Pattern of Seed Dispersal Distance and Germination Traits
2.5. Effects of Seed Traits on Seed Dispersal Distance and Germination Behavior
3. Discussion
3.1. Impact of Awn Length on Seed Yield and Components
3.2. Seed Yield and Components as Affected by Geographical Factors
3.3. The Impact of Seed Traits on Seed Dispersal Distance
3.4. The Impact of Seed Traits on Germination Indices
4. Material and Methods
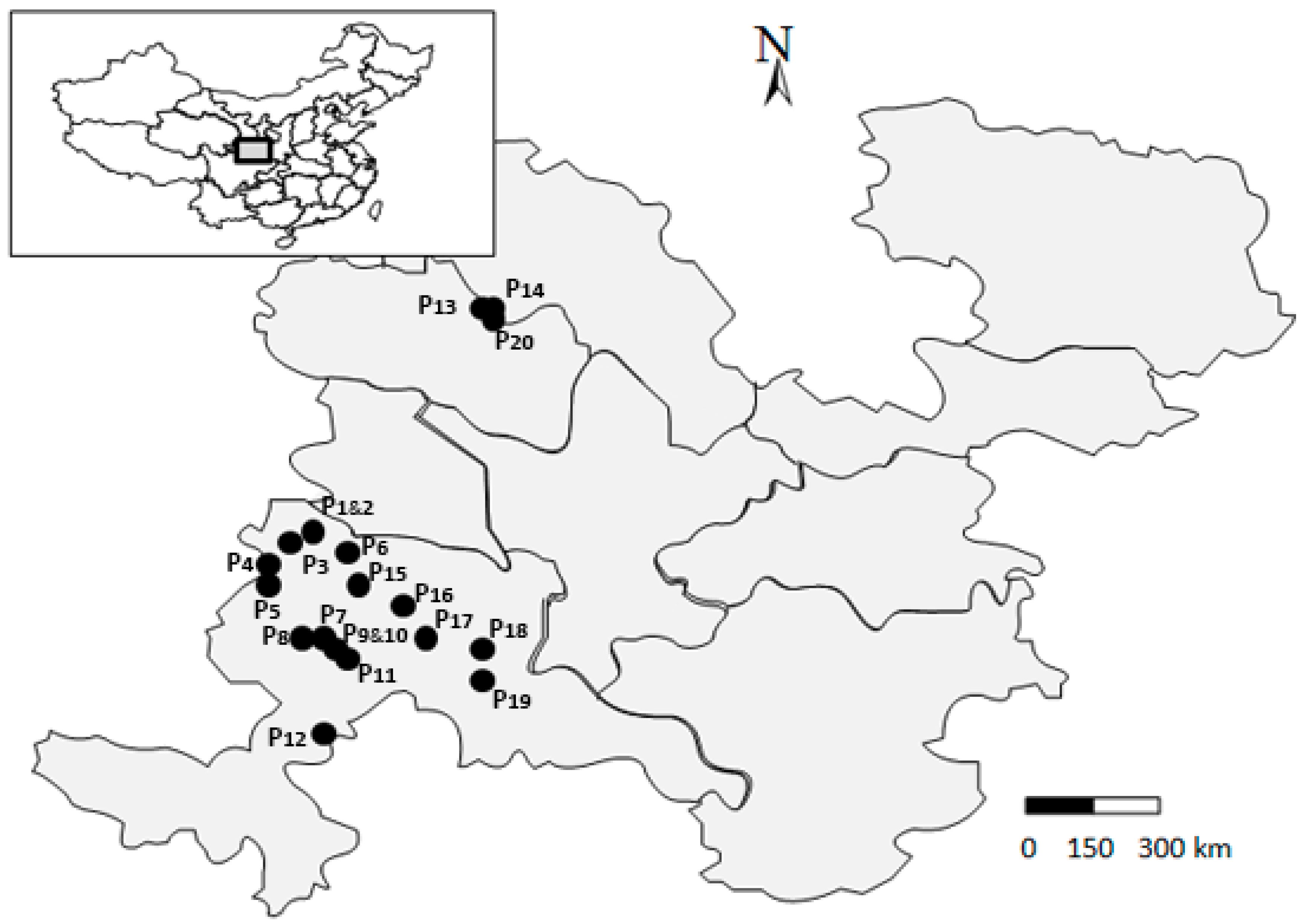
4.1. Study Area
4.2. Sampling and Measurement
4.2.1. Seed Yield Measurement
4.2.2. Evaluation of Awn Length and Seed Yield Components
4.3. Seed Dispersal Distance Measurement
4.4. Seed Germination Test
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.-Y.; Xie, W.-G.; Zhang, J.-C.; Zhao, X.-D.; Zhao, Y.-Q.; Wang, Y.-R. Phenotype-and SSR-Based Estimates of Genetic Variation between and within Two Important Elymus Species in Western and Northern China. Genes 2018, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-Y.; Hou, L.-Y.; Zhu, Y.-Q.; Zhang, Q.; Wang, H.; Xia, F.-S.; Chen, L.-L.; Mao, P.-S.; Hannaway, D.B. Siberian wildrye seed yield limited by assimilate source. Field Crop. Res. 2018, 218, 18–23. [Google Scholar] [CrossRef]
- Barkworth, M.E.; Campbell, J.J.N.; Salomon, B. Elymus. In Flora of North America North of Mexico; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2007; Volume 24, pp. 288–342. [Google Scholar]
- Yuo, T.; Yamashita, Y.; Kanamori, H.; Matsumoto, T.; Lundqvist, U.; Sato, K.; Ichii, M.; Jobling, S.A.; Taketa, S. A Short Internodes (SHI) family transcription factor gene regulates awn elongation and pistil morphology in barley. J. Exp. Bot. 2012, 63, 5223–5232. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.-G.; Zhang, J.-C.; Zhao, X.-H.; Zhang, Z.-Y.; Wang, Y.-R. Transcriptome profiling of Elymus sibiricus, an important forage grass in Qinghai-Tibet plateau, reveals novel insights into candidate genes that potentially connected to seed shattering. BMC Plant Biol. 2017, 17, 78. [Google Scholar] [CrossRef] [PubMed]
- Abel, S.; Gislum, R.; Boelt, B. Path and correlation analysis of perennial ryegrass (Lolium perenne L.) seed yield components. J. Agron. Crop Sci. 2017, 203, 338–344. [Google Scholar] [CrossRef]
- Guo, Z.; Schnurbusch, T. Variation of floret fertility in hexaploid wheat revealed by tiller removal. J. Exp. Bot. 2015, 66, 5945–5958. [Google Scholar] [CrossRef]
- Capstaff, N.M.; Miller, A.J. Improving the yield and nutritional quality of forage crops. Front. Plant Sci. 2018, 9, 535. [Google Scholar] [CrossRef]
- Rebetzke, G.; Bonnett, D.; Reynolds, M. Awns reduce grain number to increase grain size and harvestable yield in irrigated and rainfed spring wheat. J. Exp. Bot. 2016, 67, 2573–2586. [Google Scholar] [CrossRef]
- Wang, J.; Xie, J.; Zhang, Y.; Gao, S.; Zhang, J.; Mu, C. Methods to improve seed yield of Leymus chinensis based on nitrogen application and precipitation analysis. Agron. J. 2010, 102, 277–281. [Google Scholar] [CrossRef]
- Liller, C.B.; Walla, A.; Boer, M.P.; Hedley, P.; Macaulay, M.; Effgen, S.; von Korff, M.; van Esse, G.W.; Koornneef, M. Fine mapping of a major QTL for awn length in barley using a multiparent mapping population. Theor. Appl. Genet. 2017, 130, 269–281. [Google Scholar] [CrossRef]
- Motzo, R.; Giunta, F. Awnedness affects grain yield and kernel weight in near-isogenic lines of durum wheat. Aust. J. Agric. Res. 2002, 53, 1285–1293. [Google Scholar] [CrossRef]
- Guo, Z.; Schnurbusch, T. Costs and benefits of awns. J. Exp. Bot. 2016, 67, 2533. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Bragado, R.; Molero, G.; Reynolds, M.P.; Araus, J.L. Photosynthetic contribution of the ear to grain filling in wheat: A comparison of different methodologies for evaluation. J. Exp. Bot. 2016, 67, 2787–2798. [Google Scholar] [CrossRef] [PubMed]
- Merah, O.; Monneveux, P. Contribution of different organs to grain filling in durum wheat under mediterranean conditions I. Contribution of post-anthesis photosynthesis and remobilization. J. Agron. Crop Sci. 2015, 201, 344–352. [Google Scholar] [CrossRef]
- Johnson, E.E.; Baruch, Z. Awn length variation and its effect on dispersal unit burial of Trachypogon spicatus (Poaceae). Rev. Biol. Trop. 2014, 62, 319–324. [Google Scholar] [CrossRef]
- Yoshioka, M.; Iehisa, J.C.; Ohno, R.; Kimura, T.; Enoki, H.; Nishimura, S.; Nasuda, S.; Takumi, S. Three dominant awnless genes in common wheat: Fine mapping, interaction and contribution to diversity in awn shape and length. PLoS ONE 2017, 12, e0176148. [Google Scholar] [CrossRef]
- Yang, W.; Liu, F.; Zhang, S.; An, S. Dispersal and germination syndromes of tree seeds in a seasonal evergreen monsoon rainforest on Hainan Island, China. Seed Sci. Res. 2013, 23, 41–55. [Google Scholar] [CrossRef]
- Thapliyal, R.C.; Phartyal, S.S. Dispersal and germination syndromes of tree seeds in a monsoonal forest in northern India. Seed Sci. Res. 2007, 15, 29–42. [Google Scholar] [CrossRef]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef]
- Martinez, S.A.; Godoy, J.; Huang, M.; Zhang, Z.; Carter, A.H.; Garland Campbell, K.A.; Steber, C.M. Genome-Wide Association Mapping for Tolerance to Preharvest Sprouting and Low Falling Numbers in Wheat. Front. Plant Sci. 2018, 9, 141. [Google Scholar] [CrossRef]
- Elbaum, R.; Zaltzman, L.; Burgert, I.; Fratzl, P. The role of wheat awns in the seed dispersal unit. Science 2007, 316, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.; Kim, W.; Kim, H.-Y. Self-burial mechanics of hygroscopically responsive awns. Integr. Comp. Biol. 2014, 54, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Vervelde, G. The agricultural value of awns in cereals. Neth. J. Agric. Sci. 1953, 1, 2–10. [Google Scholar]
- Chauhan, B.S.; Johnson, D.E. Weedy Rice (Oryza sativa) I. Grain Characteristics and Growth Response to Competition of Weedy Rice Variants from Five Asian Countries. Weed Sci. 2010, 58, 374–380. [Google Scholar] [CrossRef]
- Singh, P.; Ibrahim, H.M.; Flury, M.; Schillinger, W.F.; Knappenberger, T. Critical water potentials for germination of wheat cultivars in the dryland Northwest USA. Seed Sci. Res. 2013, 23, 189–198. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Poustini, K.; Siddique, K.H.; Palta, J.A. Photosynthesis of barley awns does not play a significant role in grain yield under terminal drought. Crop Pasture Sci. 2012, 63, 489–499. [Google Scholar] [CrossRef]
- Li, X.-J.; Wang, H.-G.; Li, H.-B.; Zhang, L.-Y.; Teng, N.-J.; Lin, Q.-Q.; Wang, J.; Kuang, T.-Y.; Li, Z.-S.; Li, B.; et al. Awns play a dominant role in carbohydrate production during the grain-filling stages in wheat (Triticum aestivum). Physiol. Plant. 2006, 127, 701–709. [Google Scholar] [CrossRef]
- Luo, J.-H.; Liu, H.; Zhou, T.-Y.; Gu, B.-G.; Huang, X.-H.; Shangguan, Y.-Y.; Zhu, J.-J.; Li, Y.; Zhao, Y.; Wang, Y.-C.; et al. An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice. Plant Cell 2013, 25, 3360–3376. [Google Scholar] [CrossRef]
- Merah, O.; Evon, P.; Monneveux, P. Participation of Green Organs to Grain Filling in Triticum turgidum var durum Grown under Mediterranean Conditions. Int. J. Mol. Sci. 2017, 19, 56. [Google Scholar] [CrossRef]
- Li, X.; Yin, X.; Yang, S.-H.; Yang, Y.-Q.; Qian, M.; Zhou, Y.-L.; Zhang, C.; Du, Y.; Yang, Y.-P. Variations in seed characteristics among and within Stipa purpurea populations on the Qinghai—Tibet Plateau. Botany 2015, 93, 651–662. [Google Scholar] [CrossRef]
- Furuta, T.; Komeda, N.; Asano, K.; Uehara, K.; Gamuyao, R.; Angeles-Shim, R.B.; Nagai, K.; Doi, K.; Wang, D.R.; Yasui, H.; et al. Convergent loss of awn in two cultivated rice species Oryza sativa and Oryza glaberrima is caused by mutations in different loci. G3 Genes Genomes Genet. 2015, 5, 2267–2274. [Google Scholar] [CrossRef]
- Hebblethwaite, P.D.; Ivins, J.D. Nitrogen studies in Lolium perenne grown for seed I. Level of application. Grass Forage Sci. 1977, 32, 195–204. [Google Scholar] [CrossRef]
- Maydup, M.L.; Antonietta, M.; Graciano, C.; Guiamet, J.J.; Tambussi, E.A. The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: Responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field Crop. Res. 2014, 167, 102–111. [Google Scholar] [CrossRef]
- Li, Z.-Q.; Lu, W.; Yang, L.; Kong, X.-H.; Deng, X.-W. Seed weight and germination behavior of the submerged plant Potamogeton pectinatus in the arid zone of northwest China. Ecol. Evol. 2015, 5, 1504–1512. [Google Scholar] [CrossRef]
- Assefa, Y.; Bajjalieh, N.; Archontoulis, S.; Casteel, S.; Davidson, D.; Kovács, P.; Naeve, S.; Ciampitti, I.A. Spatial Characterization of Soybean Yield and Quality (Amino Acids, Oil, and Protein) for United States. Sci. Rep. 2018, 8, 14653. [Google Scholar] [CrossRef] [Green Version]
- Garnier, L.K.; Dajoz, I. Evolutionary significance of awn length variation in a clonal grass of fire-prone savannas. Ecology 2001, 82, 1720–1733. [Google Scholar] [CrossRef]
- Steinbrecher, T.; Leubner-Metzger, G. The biomechanics of seed germination. J. Exp. Bot. 2016, 68, 765–783. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Bai, Y.; Lamb, E.G.; Simpson, D.; Liu, G.; Wei, Y.; Wang, D.; McKenney, D.W.; Papadopol, P. Patterns of cross-continental variation in tree seed mass in the Canadian boreal forest. PLoS ONE 2013, 8, e61060. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Allaby, R. Seed dispersal and crop domestication: Shattering, germination and seasonality in evolution under cultivation. Annu. Plant Rev. Fruit Dev. Seed Dispersal 2009, 38, 238–295. [Google Scholar]
- Javaid, M.M.; Florentine, S.; Ali, H.H.; Weller, S. Effect of environmental factors on the germination and emergence of Salvia verbenaca L. cultivars (verbenaca and vernalis): An invasive species in semi-arid and arid rangeland regions. PLoS ONE 2018, 13, e0194319. [Google Scholar] [CrossRef]
- Zhao, R.; Cheng, Z.; Lu, W.-F.; Lu, B.-R. Estimating genetic diversity and sampling strategy for a wild soybean (Glycine soja) population based on different molecular markers. Sci. Bull. 2006, 51, 1219–1227. [Google Scholar] [CrossRef]
- Wang, M.-Y.; Hou, L.Y.; Zhang, Q.; Yu, X.-N.; Zhao, L.; Lu, J.-X.; Mao, P.-S.; Hannaway, D.B. Influence of Row Spacing and P and N Applications on Seed Yield Components and Seed Yield of Siberian Wildrye (Elymus sibiricus L.). Crop Sci. 2017, 57, 2205–2212. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671. [Google Scholar] [CrossRef]
- Deng, W.-K.; Wang, Y.-B.; Liu, Z.-X.; Cheng, H.; Xue, Y. HemI: A toolkit for illustrating heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Traits | Minimum | Maximum | Mean | SD | CV (%) |
|---|---|---|---|---|---|
| AL (mm) | 6.30 | 19.39 | 12.41 | 2.76 | 22.26 |
| SL (mm) | 8.37 | 10.48 | 9.40 | 0.69 | 7.32 |
| SW (mm) | 1.51 | 1.76 | 1.62 | 0.07 | 4.42 |
| SPI | 19.34 | 27.77 | 23.82 | 2.23 | 9.35 |
| FLS | 5.80 | 6.47 | 6.10 | 0.16 | 2.55 |
| SS (%) | 38.50 | 73.09 | 60.76 | 8.04 | 13.24 |
| SI | 55.25 | 93.86 | 86.76 | 10.41 | 11.99 |
| TSW (g) | 3.05 | 4.70 | 3.89 | 0.43 | 10.95 |
| SYP (g) | 7.60 | 9.99 | 8.76 | 0.56 | 6.43 |
| Source of Variance | df | AL | SL | SW | SPI | FLS | SS (%) | SI | TSW | SYP |
|---|---|---|---|---|---|---|---|---|---|---|
| Population (P) | 19 | *** | *** | *** | *** | ns | *** | *** | *** | *** |
| Spikelet position (Sp) | 2 | ** | ns | * | ns | ns | ns | ns | ns | ns |
| P × Sp | 38 | *** | ns | ** | ns | ns | ns | ns | ns | ns |
| Populations | AL (mm) | SL (mm) | SW (mm) | SPI | FLS | SS (%) | SI | TSW (g) | SYP (g) |
|---|---|---|---|---|---|---|---|---|---|
| XH01 | 12.05ef | 9.73a–e | 1.65a–e | 22.69e–h | 6.47a | 58.19b–e | 86.44ab | 4.00a–e | 8.96bc |
| XH02 | 12.51ef | 9.78a–d | 1.70a–d | 25.73a–d | 6.16a | 58.07c–e | 92.28a | 4.01a–d | 9.50b |
| XH03 | 13.33c–e | 10.41ab | 1.66a–e | 20.64hi | 6.34a | 53.36e | 72.56c | 4.03a–e | 8.25fg |
| XH04 | 9.43h | 8.61fg | 1.57b–e | 21.45g–i | 6.02a | 72.74ab | 90.00a | 3.32ef | 7.96c–f |
| XH05 | 10.78f–h | 8.97d–g | 1.56b–e | 22.25f–h | 5.88a | 71.24a–c | 92.71a | 3.51c–f | 8.67b–e |
| LQ01 | 13.00ef | 8.37g | 1.59a–e | 26.83a–c | 6.08a | 56.65de | 91.83a | 3.91b–e | 8.80gh |
| LQ02 | 12.49ef | 9.00d–g | 1.54de | 21.70g–i | 6.14a | 55.06de | 84.80ab | 3.82b–e | 7.88bc |
| LQ03 | 14.20cd | 8.47fg | 1.65a–e | 27.26ab | 6.16a | 55.83de | 93.19a | 4.12a–d | 9.13b–d |
| LQ04 | 11.98d–g | 9.04c–g | 1.55c–e | 24.74b–f | 6.16a | 60.87a–e | 92.54a | 3.83b–e | 8.82b–d |
| LQ05 | 11.92d–g | 9.28c–g | 1.61a–e | 24.61b–f | 5.84a | 64.39a–e | 92.03a | 3.84b–e | 8.91h |
| LQ06 | 6.30i | 8.62fg | 1.60a–e | 22.48f–h | 6.06a | 73.09a | 92.30a | 3.05f | 7.60b–d |
| HZ01 | 11.37e–h | 8.75e–g | 1.67a–e | 25.02a–f | 5.80a | 68.34a–d | 93.86a | 4.21a–c | 8.85e–g |
| TZ01 | 11.45e–h | 9.41b–f | 1.51e | 23.63d–g | 6.12a | 63.99a–e | 92.14a | 3.94b–e | 8.27d–g |
| TZ02 | 9.76gh | 8.87d–g | 1.56b–e | 22.28f–h | 6.16a | 66.36a–e | 90.14a | 3.37d–f | 8.38bc |
| TZ03 | 11.70e–h | 10.07a–c | 1.59a–e | 24.16c–g | 6.10a | 63.11a–e | 91.87a | 3.29ef | 9.06bcd |
| ZN01 | 15.45bc | 10.04a–c | 1.76a | 27.77a | 6.18a | 53.94e | 77.57bc | 4.41ab | 8.96bc |
| ZN02 | 19.39a | 10.48a | 1.72a–c | 19.34i | 6.12a | 38.50f | 55.25d | 4.70a | 9.26b |
| ZN03 | 17.20ab | 10.41ab | 1.73ab | 24.53b–f | 5.96a | 56.30de | 68.09c | 4.57ab | 9.99a |
| ZN04 | 11.62e–h | 9.79a–d | 1.56b–e | 23.99d–g | 6.14a | 63.18a–e | 92.76a | 3.83b–e | 8.99bc |
| ZN05 | 12.26ef | 9.82a–d | 1.69a–e | 25.31a–e | 6.02a | 60.90a–e | 92.92a | 3.96a–e | 9.22b |
| Traits | Lat | Long | Alt (m) | SL (mm) | SW (mm) | SPI | FLS | SS (%) | SI | TSW (g) | SYP (g) | AL (mm) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SL (mm) | 0.047 ns | −0.663 ** | −0.344 ns | 1.000 | ||||||||
| SW (mm) | −0.196 ns | −0.331 ns | 0.074 ns | 0.428 ** | 1.000 | |||||||
| SPI | −0.056 ns | −0.127 ns | 0.087 ns | −0.106 ns | 0.111 ns | 1.000 | ||||||
| FLS | 0.196 ns | −0.135 ns | −0.364 ns | 0.144 * | 0.132 ns | −0.011 ns | 1.000 | |||||
| SS (%) | 0.121 ns | 0.299 ns | −0.061 ns | −0.369 ** | −0.206 ** | 0.011 ns | −0.473 ** | 1.000 | ||||
| SI | 0.167 ns | 0.403 ns | −0.076 ns | −0.447 ** | −0.278 ** | 0.312 ** | −0.062 ns | 0.519 ** | 1.000 | |||
| TSW (g) | −0.182 ns | −0.385 ns | 0.041 ns | 0.466 ** | 0.604 ** | 0.094 ns | 0.059 ns | −0.418 ** | −0.493 ** | 1.000 | ||
| SYP (g) | −0.128 ns | −0.487 * | −0.161 ns | 0.487 ** | 0.448 ** | 0.371 ** | −0.025 ns | −0.231 ** | −0.117 ns | 0.526 ** | 1.000 | |
| AL (mm) | −0.189 ns | −0.400 ns | 0.094 ns | 0.474 ** | 0.345 ** | 0.069 ns | 0.054 ns | −0.677 ** | −0.722 ** | 0.721 ** | 0.544 ** | 1.000 |
| Eigenanalysis | Characteristic | Principal components | ||
|---|---|---|---|---|
| PC1 | PC2 | PC3 | ||
| Variation explained | Eigenvalue | 3.814 | 1.554 | 1.170 |
| Contribution rate (%) | 42.379 | 17.270 | 12.999 | |
| Cumulative rate (%) | 42.379 | 59.648 | 72.648 | |
| Eigenvectors | Traits | |||
| AL (mm) | 0.886 | −0.059 | −0.099 | |
| TSW (g) | 0.832 | 0.169 | −0.134 | |
| SL | 0.706 | −0.031 | −0.113 | |
| SI | −0.702 | 0.491 | 0.295 | |
| SS (%) | −0.696 | 0.364 | −0418 | |
| SYP (g) | 0.641 | 0.593 | −0.002 | |
| SW (mm) | 0.630 | 0.298 | −0.025 | |
| SPI | 0.051 | 0.773 | 0.399 | |
| FLS | 0.227 | −0.331 | 0.841 | |
| Populations | SDD (m) | GP (%) | GI | GDU (days) | GDE (days) |
|---|---|---|---|---|---|
| XH01 | 1.33a–c | 90.00ab | 2.98ab | 3.80a | 3.00c |
| XH02 | 1.36a–c | 92.00a | 3.02ab | 4.00a | 3.00c |
| XH03 | 1.23bc | 85.00ab | 2.75a–d | 4.00a | 3.30a–c |
| XH04 | 1.06c | 95.00a | 2.71a–d | 4.30a | 3.00c |
| XH05 | 1.34a–c | 96.00a | 2.48b–d | 4.80a | 3.10bc |
| LQ01 | 1.23bc | 96.00a | 2.46b–d | 4.70a | 3.40a–c |
| LQ02 | 1.28bc | 96.00a | 2.49b–d | 4.90a | 3.20bc |
| LQ03 | 1.47ab | 94.00a | 2.34cd | 4.40a | 3.30a–c |
| LQ04 | 1.23bc | 90.00ab | 2.84a–c | 4.20a | 3.00c |
| LQ05 | 1.24bc | 86.00ab | 2.47b–d | 5.10a | 3.20bc |
| LQ06 | 1.23bc | 97.00a | 2.24d | 4.20a | 3.80a |
| HZ01 | 1.68a | 95.00a | 2.21d | 5.00a | 3.60ab |
| TZ01 | 1.30a–c | 90.00ab | 3.10a | 4.20a | 3.00c |
| TZ02 | 1.07c | 95.00a | 2.87a–c | 4.00a | 3.00c |
| TZ03 | 1.40a–c | 93.00a | 3.01ab | 3.80a | 3.20bc |
| ZN01 | 1.35a–c | 93.00a | 2.50b–d | 5.20a | 3.20bc |
| ZN02 | 1.02c | 78.00b | 2.36cd | 5.00a | 3.30a–c |
| ZN03 | 1.51ab | 87.00ab | 2.48b–d | 4.40a | 3.00c |
| ZN04 | 1.20bc | 86.00ab | 2.57a–d | 5.00a | 3.10bc |
| ZN05 | 1.24bc | 95.00a | 2.95ab | 4.70a | 3.00c |
| Seed Traits | GP (%) | GI | GDU (days) | GDE (days) | SDD (m) |
|---|---|---|---|---|---|
| AL (mm) | −0.357 ** | −0.080 ns | 0.075 ns | −0.057 ns | 0.059 ns |
| SL (mm) | −0.313 ** | 0.132 ns | −0.059 ns | −0.105 ns | 0.034 ns |
| SW (mm) | −0.140 * | −0.241 ** | 0.050 ns | 0.139 * | 0.121 ns |
| TSW (g) | −0.241 ** | −0.084 ns | 0.088 ns | 0.031 ns | 0.108 ns |
| Eigenanalysis | Characteristic | Principal Components | ||
|---|---|---|---|---|
| PC1 | PC2 | PC3 | ||
| Variation explained | Eigenvalue | 2.762 | 1.69 | 1.102 |
| Contribution rate (%) | 30.685 | 18.78 | 12.249 | |
| Cumulative rate (%) | 30.685 | 49.466 | 61.714 | |
| Eigenvectors | Traits | |||
| TSW (g) | 0.858 | 0.118 | 0.110 | |
| AL (mm) | 0.803 | 0.174 | −0.132 | |
| SW (mm) | 0.717 | −0.069 | 0.327 | |
| SL (mm) | 0.682 | 0.396 | −0.043 | |
| GP (%) | −0.516 | 0.136 | 0.479 | |
| GI | −0.276 | 0.821 | 0.059 | |
| GDE (days) | 0.072 | −0.644 | 0.358 | |
| GDU (days) | 0.168 | −0.605 | −0.390 | |
| SDD (m) | 0.161 | −0.104 | 0.672 | |
| Code | Populations | Location | Latitude (N) | Longitude (E) | Altitude (m) |
|---|---|---|---|---|---|
| P1 | XH01 | Xiahe, Gansu, China | 35°13.267′ | 102°49.067′ | 2550 |
| P2 | XH02 | Xiahe, Gansu, China | 35°13.267′ | 102°49.067′ | 2550 |
| P3 | XH03 | Xiahe, Gansu, China | 35°12.392′ | 102°40.479′ | 2770 |
| P4 | XH04 | Xiahe, Gansu, China | 35°11.270′ | 102°30.313′ | 2960 |
| P5 | XH05 | Xiahe, Gansu, China | 35°04.138′ | 102°22.895′ | 3140 |
| P6 | LQ01 | Lintan, Gansu, China | 35°06.351′ | 102°24.655′ | 3080 |
| P7 | LQ02 | Lintan, Gansu, China | 34°35.167′ | 102°29.535′ | 3110 |
| P8 | LQ03 | Lintan, Gansu, China | 34°33.793′ | 102°34.437′ | 3080 |
| P9 | LQ04 | Lintan, Gansu, China | 34°32.108′ | 102°37.887′ | 3050 |
| P10 | LQ05 | Lintan, Gansu, China | 34°32.108′ | 102°37.887′ | 3050 |
| P11 | LQ06 | Lintan, Gansu, China | 34°29.782′ | 102°40.725′ | 3010 |
| P12 | HZ01 | Hezuo, Gansu, China | 34°05.662′ | 102°37.927′ | 3380 |
| P13 | TZ01 | Tianzhu, Gansu, China | 36°57.456′ | 103°07.778′ | 2370 |
| P14 | TZ02 | Tianzhu, Gansu, China | 36°57.703′ | 103°08.896′ | 2430 |
| P15 | TZ03 | Tianzhu, Gansu, China | 34°56.489′ | 102°54.985′ | 2960 |
| P16 | ZN01 | Zhuoni, Gansu, China | 34°50.216′ | 103°09.503′ | 3200 |
| P17 | ZN02 | Zhuoni, Gansu, China | 34°41.015′ | 103°14.835′ | 3160 |
| P18 | ZN03 | Zhuoni, Gansu, China | 34°34.037′ | 103°31.393′ | 2530 |
| P19 | ZN04 | Zhuoni, Gansu, China | 34°25.071′ | 103°35.153′ | 2540 |
| P20 | ZN05 | Zhuoni, Gansu, China | 36°57.357′ | 103°08.708′ | 2380 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntakirutimana, F.; Xiao, B.; Xie, W.; Zhang, J.; Zhang, Z.; Wang, N.; Yan, J. Potential Effects of Awn Length Variation on Seed Yield and Components, Seed Dispersal and Germination Performance in Siberian Wildrye (Elymus sibiricus L.). Plants 2019, 8, 561. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120561
Ntakirutimana F, Xiao B, Xie W, Zhang J, Zhang Z, Wang N, Yan J. Potential Effects of Awn Length Variation on Seed Yield and Components, Seed Dispersal and Germination Performance in Siberian Wildrye (Elymus sibiricus L.). Plants. 2019; 8(12):561. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120561
Chicago/Turabian StyleNtakirutimana, Fabrice, Bowen Xiao, Wengang Xie, Junchao Zhang, Zongyu Zhang, Na Wang, and Jiajun Yan. 2019. "Potential Effects of Awn Length Variation on Seed Yield and Components, Seed Dispersal and Germination Performance in Siberian Wildrye (Elymus sibiricus L.)" Plants 8, no. 12: 561. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120561