Construction of an Onion (Allium cepa L.) Genetic Linkage Map Using Genotyping-by-Sequencing Analysis with a Reference Gene Set and Identification of QTLs Controlling Anthocyanin Synthesis and Content



Abstract
:
1. Introduction
2. Results
2.1. SNP Detection and Genotyping Using GBS Analysis
2.2. Development of HRM Markers for Comparative Mapping
2.3. Construction of an Onion Genetic Linkage Map
2.4. Comparison of the SP3B×H6 and OH1×5225 Onion Genetic Linkage Maps
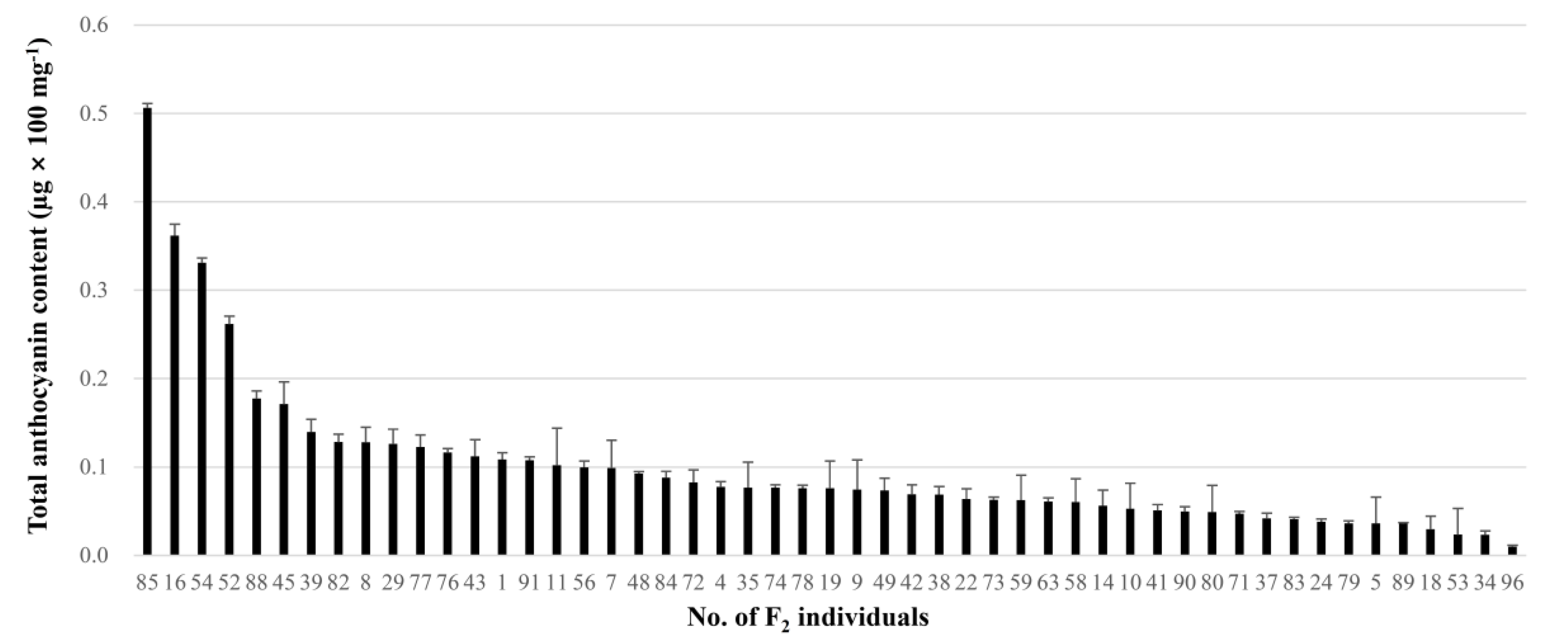
2.5. Identification of a Major QTL for Anthocyanin Synthesis of Onion Bulbs
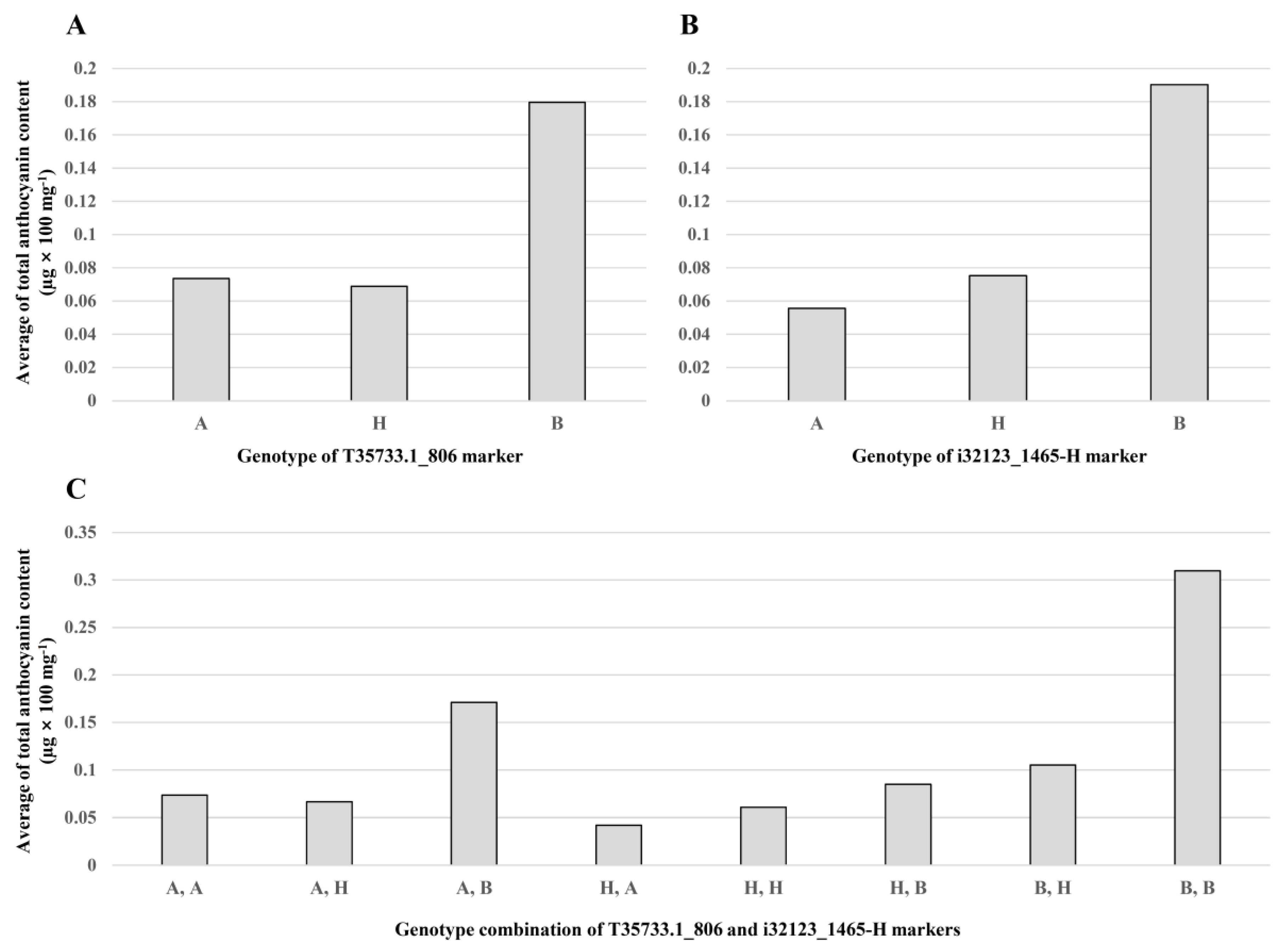
2.6. Identification of Two QTLs for Anthocyanin Content of Onion Bulbs
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Phenotyping of Bulb Color
4.3. Assessment of Anthocyanin Content
4.4. DNA Extraction
4.5. Genotyping-by-Sequencing Analysis
4.6. High-Resolution Melting Analysis
4.7. Genetic Linkage Mapping
4.8. Assignment of Linkage Groups to Onion Chromosomes
4.9. QTL Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AC | anthocyanin content |
| AFLP | amplified fragment length polymorphism |
| ANS | anthocyanidin synthase |
| AS | anthocyanin synthesis |
| CAPS | cleaved amplified polymorphic sequence |
| CIM | composite interval mapping |
| cM | centi Morgan |
| ddRAD-seq | double digest restriction site-associated DNA sequencing |
| DFR | dihydroflavonol 4-reductase |
| EST | expressed sequence tag |
| GBS | genotyping-by-sequencing |
| HRM | high-resolution melting |
| KASP | kompetitive allele specific PCR |
| LOD | logarithm of odds |
| MAS | marker-assisted selection |
| NGS | next-generation sequencing |
| QTL | quantitative trait loci |
| RAPD | random amplified polymorphic DNA |
| RFLP | restriction fragment length polymorphism |
| SNP | single-nucleotide polymorphism |
| SSCP | single-strand conformation polymorphism |
| SSR | simple sequence repeat |
References
- Khosa, J.S.; McCallum, J.; Dhatt, A.S.; Macknight, R.C. Enhancing onion breeding using molecular tools. Plant Breed. 2016, 135, 9–20. [Google Scholar] [CrossRef]
- Griffiths, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions—A global benefit to health. Phytother. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.D.; Wang, C.K.; Hu, D.G.; Hao, Y.J. How do anthocyanins paint our horticultural products? Sci. Hort. 2019, 249, 257–262. [Google Scholar] [CrossRef]
- Terahara, N.; Yamaguchi, M.; Honda, T. Malonylated anthocyanins from bulbs of red onion, Allium cepa L. Biosci. Biotech. Biochem. 1994, 58, 1324–1325. [Google Scholar] [CrossRef]
- Donner, H.; Gao, L.; Mazza, G. Separation and characterization of simple and malonylated anthocyanins in red onions, Allium cepa L. Food Res. Int. 1997, 30, 637–643. [Google Scholar] [CrossRef]
- Clarke, A.E.; Jones, H.A.; Little, T.M. Inheritance of bulb color in the onion. Genetics 1944, 29, 569–575. [Google Scholar]
- El-Shafie, M.W.; Davis, G.N. Inheritance of bulb color in the onion (Allium cepa L.). Hilgardia 1967, 38, 607–622. [Google Scholar] [CrossRef] [Green Version]
- Khar, A.; Jakse, J.; Havey, M.J. Segregations for onion-bulb colors reveal that red is controlled by at least three loci. J. Am. Soc. Hortic. Sci. 2008, 133, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Masuzaki, S.; Shigyo, M.; Yamauchi, N. Complete assignment of structural genes involved in flavonoid biosynthesis influencing bulb color to individual chromosomes of the shallot (Allium cepa L.). Genes Genet. Syst. 2006, 81, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Binzel, M.L.; Park, S.; Yoo, K.S.; Pike, L.M. Inactivation of DFR (Dihydroflavonol 4-reductase) gene transcription results in blockage of anthocyanin production in yellow onions (Allium cepa). Mol. Breed. 2004, 14, 253–263. [Google Scholar] [CrossRef]
- Kim, S.; Yoo, K.S.; Pike, L.M. Development of a PCR-based marker utilizing a deletion mutation in the dihydroflavonol 4-reductase (DFR) gene responsible for the lack of anthocyanin production in yellow onions (Allium cepa). Theor. Appl. Genet. 2005, 110, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Baek, D.; Cho, D.Y.; Lee, E.T.; Yoon, M.K. Identification of two novel inactive DFR-A alleles responsible for failure to produce anthocyanin and development of a simple PCR-based molecular marker for bulb color selection in onion (Allium cepa L.). Theor. Appl. Genet. 2009, 118, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Kim, C.W.; Moon, J.S.; Kim, S. At least nine independent natural mutations of the DFR-A gene are responsible for appearance of yellow onions (Allium cepa L.) from red progenitors. Mol. Breed. 2014, 33, 173–186. [Google Scholar] [CrossRef]
- Kim, B.; Cho, Y.; Kim, S. Identification of a novel DFR-A mutant allele determining the bulb color difference between red and yellow onions (Allium cepa L.). Plant Breed. Biotechnol. 2017, 5, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Binzel, M.L.; Yoo, K.S.; Park, S.; Pike, L.M. Pink (P), a new locus responsible for a pink trait in onions (Allium cepa) resulting from natural mutations of anthocyanidin synthase. Mol. Genet. Genom. 2004, 272, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jones, R.; Yoo, K.S.; Pike, L.M. The L locus, one of complementary genes required for anthocyanin production in onions (Allium cepa), encodes anthocyanidin synthase. Theor. Appl. Genet. 2005, 111, 120–127. [Google Scholar] [CrossRef]
- Kim, S.; Yoo, K.S.; Pike, L.M. Development of a codominant PCR-based marker for allelic selection of the pink trait in onions (Allium cepa), based on the insertion mutation in the promoter of the anthocyanidin synthase gene. Theor. Appl. Genet. 2005, 110, 573–578. [Google Scholar] [CrossRef]
- Kim, E.Y.; Kim, C.W.; Kim, S. Identification of two novel mutant ANS alleles responsible for inactivation of anthocyanidin synthase and failure of anthocyanin production in onion (Allium cepa L.). Euphytica 2016, 212, 427–437. [Google Scholar] [CrossRef]
- Chen, F.; Dong, W.; Zhang, J.; Guo, X.; Chen, J.; Wang, Z.; Lin, Z.; Tang, H.; Zhang, L. The sequenced angiosperm genomes and genome databases. Front. Plant Sci. 2018, 9, 418. [Google Scholar] [CrossRef] [Green Version]
- King, J.J.; Bradeen, J.M.; Bark, O.; McCallum, J.A.; Havey, M.J. A low-density genetic map of onion reveals a role for tandem duplication in the evolution of an extremely large diploid genome. Theor. Appl. Genet. 1998, 96, 52–62. [Google Scholar] [CrossRef]
- Van Heusden, A.W.; van Ooijen, J.W.; Vrielink-van Ginkel, R.; Verbeek, W.H.J.; Wietsma, W.A.; Kik, C. A genetic map of an interspecific cross in Allium based on amplified fragment length polymorphism (AFLPTM) markers. Theor. Appl. Genet. 2000, 100, 118–126. [Google Scholar] [CrossRef]
- Van Heusden, A.W.; Shigyo, M.; Tashiro, Y.; Vrielink-van Ginkel, R.; Kik, C. AFLP linkage group assignment to the chromosomes of Allium cepa L. via monosomic addition lines. Theor. Appl. Genet. 2000, 100, 480–486. [Google Scholar] [CrossRef]
- McCallum, J.; Leite, D.; Pither-Joyce, M.; Havey, M.J. Expressed sequence markers for genetic analysis of bulb onion (Allium cepa L.). Theor. Appl. Genet. 2001, 103, 979–991. [Google Scholar] [CrossRef]
- Martin, W.J.; McCallum, J.; Shigyo, M.; Jakse, J.; Kuhl, J.C.; Yamane, N.; Pither-Joyce, M.; Gokce, A.F.; Sink, K.C.; Town, C.D.; et al. Genetic mapping of expressed sequences in onion and in silico comparisons with rice show scant colinearity. Mol. Genet. Genom. 2005, 274, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Jakše, J.; Martin, W.; McCallum, J.; Havey, M.J. Single nucleotide polymorphisms, indels and simple sequence repeats for onion (Allium cepa L.) cultivar identification. J. Am. Soc. Hortic. Sci. 2005, 130, 912–917. [Google Scholar] [CrossRef]
- McCallum, J.; Thomson, S.; Pitcher-Joyce, M.; Kenel, F.; Clarke, A.; Havey, M.J. Genetic diversity analysis and single nucleotide polymorphism marker development in cultivated bulb onion based on expressed sequence tag-simple sequence repeats markers. J. Am. Soc. Hortic. Sci. 2008, 113, 810–818. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.; Revanna, R.; Thomson, S.; Pither-Joyce, M.; Wright, K.; Crowhurst, R.; Fiers, M.; Chen, L.; Macknight, R.; McCallum, J. A toolkit for bulk PCR-based marker design from next-generation sequence data: Application for development of a framework linkage map in bulb onion (Allium cepa L.). BMC Genom. 2012, 13, 637. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.; Pither-Joyce, M.; Wright, K.; Chen, L.; McCallum, J. Development of robust genomic simple sequence repeat markers for estimation of genetic diversity within and among bulb onion (Allium cepa L.) populations. Mol. Breed. 2012, 30, 1401–1411. [Google Scholar] [CrossRef]
- Duangjit, J.; Bohanec, B.; Chan, A.P.; Town, C.D.; Havey, M.J. Transcriptome sequencing to produce SNP-based genetic maps of onion. Theor. Appl. Genet. 2013, 126, 2093–2101. [Google Scholar] [CrossRef]
- Kim, S.; Kim, M.S.; Kim, Y.M.; Yeom, S.I.; Cheong, K.; Kim, K.T.; Jeon, J.; Kim, S.; Kim, D.S.; Sohn, S.H.; et al. Integrative structural annotation of de novo RNA-Seq provides an accurate reference gene set of the enormous genome of the onion (Allium cepa L.). DNA Res. 2015, 22, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.H.; Ahn, Y.K.; Lee, T.H.; Lee, J.E.; Jeong, M.H.; Seo, C.H.; Chandra, R.; Kwon, Y.S.; Kim, C.W.; Kim, D.S.; et al. Construction of a draft reference transcripts of onion (Allium cepa) using long-read sequencing. Plant Biotechnol. Rep. 2016, 10, 383–390. [Google Scholar] [CrossRef]
- Scholten, O.E.; van Kaauwen, M.P.W.; Shahin, A.; Hendrickx, P.M.; Keizer, L.C.P.; Burger, K.; van Heusden, A.W.; van der Linden, C.G.; Vosman, B. SNP-markers in Allium species to facilitate introgression breeding in onion. BMC Plant Biol. 2016, 16, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, J.; Purushotham, P.M.; Han, K.; Lee, H.R.; Nah, G.; Kang, B.C. Development of a genetic map for onion (Allium cepa L.) using reference-free genotyping-by-sequencing and SNP assays. Front. Plant Sci. 2017, 8, 1606. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Natarajan, S.; Biswas, M.K.; Shirasawa, K.; Isobe, S.; Kim, H.T.; Park, J.I.; Seong, C.N.; Nou, I.S. SNP discovery of Korean short day onion inbred lines using double digest restriction site-associated DNA sequencing. PLoS ONE 2018, 13, e0201229. [Google Scholar] [CrossRef] [Green Version]
- Duangjit, J.; Welsh, K.; Wise, M.L.; Bohanec, B.; Havey, M.J. Genetic analyses of anthocyanin concentrations and intensity of red bulb color among segregating haploid progenies of onion. Mol. Breed. 2014, 34, 75–85. [Google Scholar] [CrossRef]
- Shin, J.; Park, E.; Choi, G. PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. Plant J. 2007, 49, 981–994. [Google Scholar] [CrossRef]
- Mancinelli, A.L.; Schwartz, O.M. The photoregulation of anthocyanin synthesis IX. The photosensitivity of the response in dark and light-grown tomato seedlings. Plant Cell Physiol. 1984, 25, 93–105. [Google Scholar]
- Lee, Y.R.; Kim, J.; Lee, S.Y.; Lee, J. Diallelic SNP marker development and genetic linkage map construction in octoploid strawberry (Fragaria × ananassa) through next-generation resequencing and high-resolution melting analysis. Hortic. Environ. Biotechnol. 2020, 61, 371–383. [Google Scholar] [CrossRef]
- Eun, M.H.; Han, J.H.; Yoon, J.B.; Lee, J. QTL mapping of resistance to the Cucumber mosaic virus P1 strain in pepper using a genotyping-by-sequencing analysis. Hortic. Environ. Biotechnol. 2016, 57, 589–597. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, K.; Choi, D.; Lee, J. Fine mapping of the genic male-sterile ms1 gene in Capsicum annuum L. Theor. Appl. Genet. 2018, 131, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Kosambi, D.D. The estimation of map distance from recombination value. Ann. Eugen. 1944, 12, 172–175. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2007. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summary of Illumina Sequencing | Data |
|---|---|
| Number of plants for multiplexing | 96 |
| Total number of raw reads generated | 1,008,750,538 (100%) |
| Total base number of raw reads (bp) | 101,883,804,338 (101.9 Gbp) |
| Total number of demultiplexed reads | 967,290,922 (95.9%) |
| Total number of trimmed reads | 869,413,090 (86.2%) |
| Total number of mapped reads | 358,301,156 (35.5%) |
| Total number of mapped regions | 1,604,901 |
| Average depth of mapped region | 81.42 |
| Total length of mapped regions (bp) | 1,855,555 (1.9 Mbp) |
| Total length of the reference gene set (bp) | 202,991,716 (203.0 Mbp) |
| Coverage of the reference gene set | 0.9141% |
| Total number of SNPs detected | 8431 |
| Total number of SNPs filtered | 1625 |
| Number of Assembled Contigs | Total Length (bp) | Minimum Length (bp) | Maximum Length (bp) | Average Length (bp) | N50 Length (bp) |
|---|---|---|---|---|---|
| 165,179 | 202,991,716 | 200 | 16,504 | 1228 | 1756 |
| No. | SP3B×H6 Map | OH1×5225 Map z | Transcript ID y | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chr. No. x | Position (cM) | Marker Name | Forward Primer | Reverse Primer | H6 | SP3B | Chr. No. x | Position (cM) | AC.Combine.Assembly.v.1.0 | |
| 1 | 1 | 23.1 | i34152_369-HRM | TCCACATATCTCATATTGCGCTCA | CTTTGGCTTAACTTACCCGATTAC | G | A | 1 | 8.7 | AC.Combine.Locus_5700 |
| 2 | 1 | 63.6 | i37206_320-HRM | CCGGTTGTGGTTGGTCGAA | ACAAGTTAGTGGCACGTTACAAAA | G | T | 1 | 59.0 | AC.Combine.Locus_9298 |
| 3 | 2 | 39.6 | i26238_573-HRM | ACAAACCTTATGCAGATACACTCA | GCAACATCAAAAGCTCCCCATC | T | C | 2 | 67.3 | AC.Combine.Locus_14118 |
| 4 | 2 | 55.4 | i26198_779-HRM | TTCTATTACCGGAGCTGTAGTTGG | CAAATGCAATATCTCCAAGGGCTT | G | A | 2 | 95.5 | AC.Combine.Locus_9799 |
| 5 | 2 | 82.8 | i30225_1161-HRM | GAAGGGACAGTTCAAGGTAGTAGG | TCTCAAATTCCTTCTCCAACTTCA | G | A | 2 | 154.5 | AC.Combine.Locus_7495 |
| 6 | 2 | 96.9 | i32865_1404-HRM | TAGTCAGAATCTTCCTCTCCTGGT | AGTGGAGGAAGATGAAGAAGTTGA | A | G | 2 | 193.2 | AC.Combine.Locus_19254 |
| 7 | 2 | 100.7 | i32416_685-HRM | AGCAATGAAGTACGATTTACAGCA | TGAAGAAGAACCCTCCAACGTTAT | T | C | 2 | 203.3 | AC.Combine.Locus_17664 |
| 8 | 2 | 116.5 | i33538_1298-HRM | AATCGCCATTAGAAAGCTTTACCG | TACACTAAACCCTACAAACGTCGA | C | G | 2 | 217.9 | AC.Combine.Locus_250 |
| 9 | 2 | 126.2 | i26131_2020-HRM | GCTTCTTTGGCCCCATATTCAAG | CATTTGCATAATGTGAGAAAGCGC | T | C | 2 | 227.0 | AC.Combine.Locus_1772 |
| 10 | 3 | 0 | i35099_237-HRM | GAAGGATGCTGGTAAGAGGTCTAC | ATTATCCAAACCTGTACCCGTGAA | C | T | 3 | 0.0 | AC.Combine.Locus_31460 |
| 11 | 3 | 20.2 | i39498_201-HRM | AAGAGTTGGGTGTGAAAGGAGATT | CCTGTGTTGAGATTTGGGGATTTC | T | C | 3 | 12.7 | AC.Combine.Locus_89589 |
| 12 | 3 | 25.8 | i33531_1155-HRM | CCTTATGCAGATTCACCATGGAAG | CGGATCTCGTTTAACAGTGGAAAG | T | C | 3 | 12.7 | AC.Combine.Locus_14239 |
| 13 | 3 | 39.2 | i35038_601-HRM | GACTTGGAGTGCAGTTGAGAC | AATCATCGGGCCTCAATGTTCAA | G | A | 3 | 25.1 | AC.Combine.Locus_14173 |
| 14 | 3 | 100.9 | i26005_1583-HRM | CAGAGATCTCAACTTGTTCCCTGA | ATTGCATACCTCGAATCGCCTTTA | A | G | 3 | 174.0 | AC.Combine.Locus_9082 |
| 15 | 4 | 0 | i29163_2080-HRM | TTCAGTAAACAAAAGATCGGCTGA | AAATCGGCCATCTTATTGTCTCCA | G | A | 4B | 0.0 | AC.Combine.Locus_8343 |
| 16 | 4 | 26.2 | i26442_1225-HRM | ACATTCTTCAAAGCGGTAACAACC | CAGTCATATACACCTTTATGCAAGT | A | G | 4A | 28.3 | AC.Combine.Locus_1490 |
| 17 | 4 | 54.3 | i26526_748-HRM | AGGAGGTAATGCACTGATTATTTGT | TGCACAATTGAGAGAAGGTGTTTT | A | G | 4B | 52.4 | AC.Combine.Locus_10803 |
| 18 | 4 | 62.0 | i32123_1465-HRM | CACGAATCCATAAGAGTTATCGCA | TGATCAGGGCTAGGAAAGTTTGAT | T | C | 4B | 52.4 | AC.Combine.Locus_5789 |
| 19 | 5 | 68.6 | i25881_1343-2-HRM | TTCTGACAATTTGACCGGTTGAAG | CGCGGTTACTCAAGGTTTAAGATT | T | C | 5 | 59.0 | AC.Combine.Locus_3681 |
| 20 | 5 | 69.1 | i25881_1343-1-HRM | CCATCCTGAACACGATAAACCTTC | GATTAGGAGTTTGGCTTTGCTGTG | C | T | 5 | 59.0 | AC.Combine.Locus_3681 |
| 21 | 5 | 85.6 | i29728_1131-HRM | CACAAAGGGGAATCAATAATCGCA | GCCTGCTCTTGGAACTGATAAAAT | T | C | 5 | 119.4 | AC.Combine.Locus_2597 |
| 22 | 5 | 112.1 | i30593_868-HRM | TAAAGACCACAACAGACTCGTTCA | TTGGTTAAGGGAGTCTATGTGAGC | T | C | 5 | 178.6 | AC.Combine.Locus_70708 |
| 23 | 5 | 115.2 | i36364_683-HRM | GAACCCGCCTAAGAACCAGAA | TTCATCCTCGGACTGTCTACTAGA | G | A | 5 | 176.4 | AC.Combine.Locus_24909 |
| 24 | 5 | 118.4 | i29592_700-HRM | CTTCTAGAGTTGGTGTTGTGTCCA | ACTCTATGCAAACTTCACCTGAGA | G | C | 5 | 178.6 | AC.Combine.Locus_4560 |
| 25 | 6 | 54.8 | i30880_1388-HRM | CGTTGGAAGATTATGTTCATCGCA | TTGGCTGCAGTGAAGTAGGTATAG | C | A | 6B | 9.3 | AC.Combine.Locus_8405 |
| 26 | 6 | 73.3 | i35768_1013-HRM | GACATGCCGCAATCCAAGATTAG | CGGTAGATGGTGAAATTTGTGTCA | T | C | 6B | 30.2 | AC.Combine.Locus_37095 |
| 27 | 6 | 91.1 | i32739_152-HRM | AAACGGCCATCTTGAAGCAATAGA | GCAAAACTTGGTCAGATAGAGAGC | G | A | 6B | 41.0 | AC.Combine.Locus_12004 |
| 28 | 6 | 91.7 | i36782_698-HRM | GCATGTTGATAGGAATTCGAATGC | GTGTTGTCTTGTTCTCGTGGTTC | A | T | 6B | 44.3 | AC.Combine.Locus_15991 |
| 29 | 7 | 15.6 | i39918_357-HRM | ATAACCTCTTCTCAATTCGAACTTC | TCCGATCCTCAATGACGACAATAA | C | G | 7 | 39.1 | AC.Combine.Locus_48105 |
| 30 | 7 | 38.8 | i29101_1894-HRM | CATACCAACCTGCACACTTAAACA | GTACCATAGCGACATCCTATAGCC | A | G | 7 | 92.6 | AC.Combine.Locus_854 |
| 31 | 7 | 43.4 | i28923_2628-HRM | TACTATGGGAATTAGCTACGATGC | AACCGTCTATCCTGGAACCCTA | C | T | 7 | 94.8 | AC.Combine.Locus_51 |
| 32 | 8 | 18.2 | i31126_1315-HRM | ACTCTACTTGATGTTCAGTGTGGC | CTTGTCATCATCTTTCCCTAGGCT | T | C | 8 | 18.2 | AC.Combine.Locus_2785 |
| 33 | 8 | 22.0 | i30907_420-HRM | TGGCTCTACTGGGGATTTGTTAAA | CACTCGGCAAATATCCCTGGTAG | C | T | 8 | 15.4 | AC.Combine.Locus_65044 |
| 34 | 8 | 66.9 | i31261_1350-HRM | GTCCCCTAGAAACAGATCTCCAAC | CGACTGTGACTTTTCGGGAATTTA | A | C | 8 | 69.4 | AC.Combine.Locus_815 |
| Chromosome No. | Length of Linkage Maps (cM) | Total Number of Markers (A+B) | Number of SNPs Resulting from GBS (A) z | Number of HRM Markers (B) y |
|---|---|---|---|---|
| 1 | 122.5 | 36 | 34 | 2 |
| 2 | 127.9 | 53 | 46 | 7 |
| 3 | 134.1 | 42 | 37 | 5 |
| 4 | 94.5 | 38 | 34 | 4 |
| 5 | 142.8 | 45 | 39 | 6 |
| 6 | 111.3 | 64 | 60 | 4 |
| 7 | 73.9 | 38 | 35 | 3 |
| 8 | 74.3 | 37 | 34 | 3 |
| Total | 881.4 | 353 | 319 | 34 |
| Average | 110.2 | 44 | 40 | 4 |
| Population | Generation | Number of Onion Plants | Expected Ratio | X2 Value | p Value | ||
|---|---|---|---|---|---|---|---|
| Red Bulb | Yellow Bulb | Total | |||||
| SP3B×H6 | F2 | 51 | 18 | 69 | 3:1 | 0.0435 | 0.835 |
| Trait | QTL | Chr. | Marker Interval | QTL Peak Position (cM) | Additive Effect | Dominance Effect | R2 z (%) | LOD y Value | LOD Threshold x |
|---|---|---|---|---|---|---|---|---|---|
| Anthocyanin synthesis | qAS7.1 | 7 | T25488.1_1462-i39918_357-HRM | 13.8 | −0.9573 | 0.2401 | 87.61 | 9.19 | 5.3 |
| Anthocyanin content | qAC4.1 | 4 | T57513.1_314-T53764.1_356 | 7.0 | −0.0299 | −0.0513 | 19.43 | 3.26 | 3.0 |
| qAC4.2 | 4 | T84695.1_220-i32123_1465-HRM | 62.0 | −0.0399 | −0.0335 | 26.28 | 3.03 | 3.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.; Kim, S.; Lee, J. Construction of an Onion (Allium cepa L.) Genetic Linkage Map Using Genotyping-by-Sequencing Analysis with a Reference Gene Set and Identification of QTLs Controlling Anthocyanin Synthesis and Content. Plants 2020, 9, 616. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050616
Choi Y, Kim S, Lee J. Construction of an Onion (Allium cepa L.) Genetic Linkage Map Using Genotyping-by-Sequencing Analysis with a Reference Gene Set and Identification of QTLs Controlling Anthocyanin Synthesis and Content. Plants. 2020; 9(5):616. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050616
Chicago/Turabian StyleChoi, Yousoo, Sunggil Kim, and Jundae Lee. 2020. "Construction of an Onion (Allium cepa L.) Genetic Linkage Map Using Genotyping-by-Sequencing Analysis with a Reference Gene Set and Identification of QTLs Controlling Anthocyanin Synthesis and Content" Plants 9, no. 5: 616. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050616