Sustainable Land Use Management Needed to Conserve the Dragon’s Blood Tree of Socotra Island, a Vulnerable Endemic Umbrella Species

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
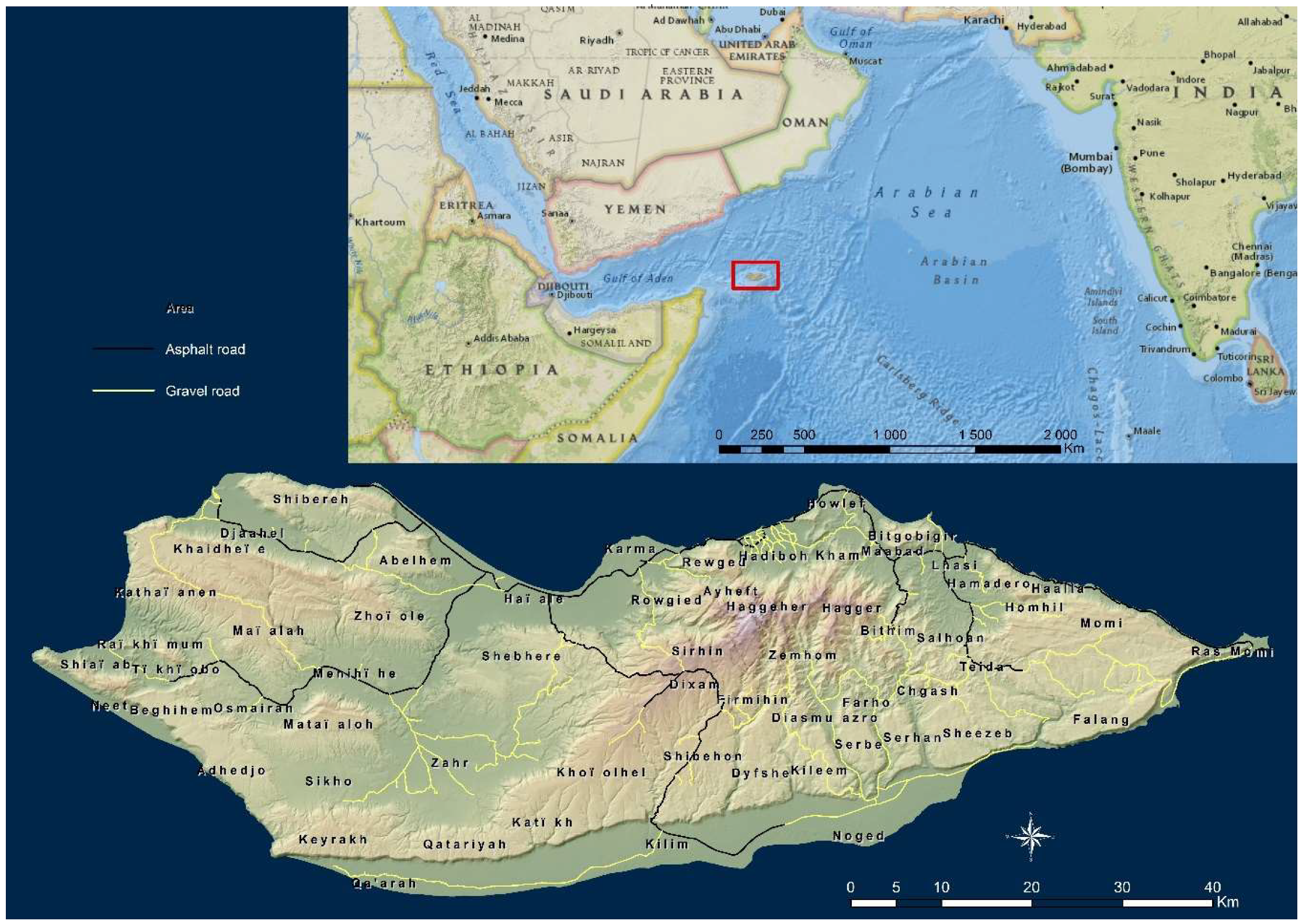
2.1. Detection of Dragon’s Blood Trees on Socotra Island
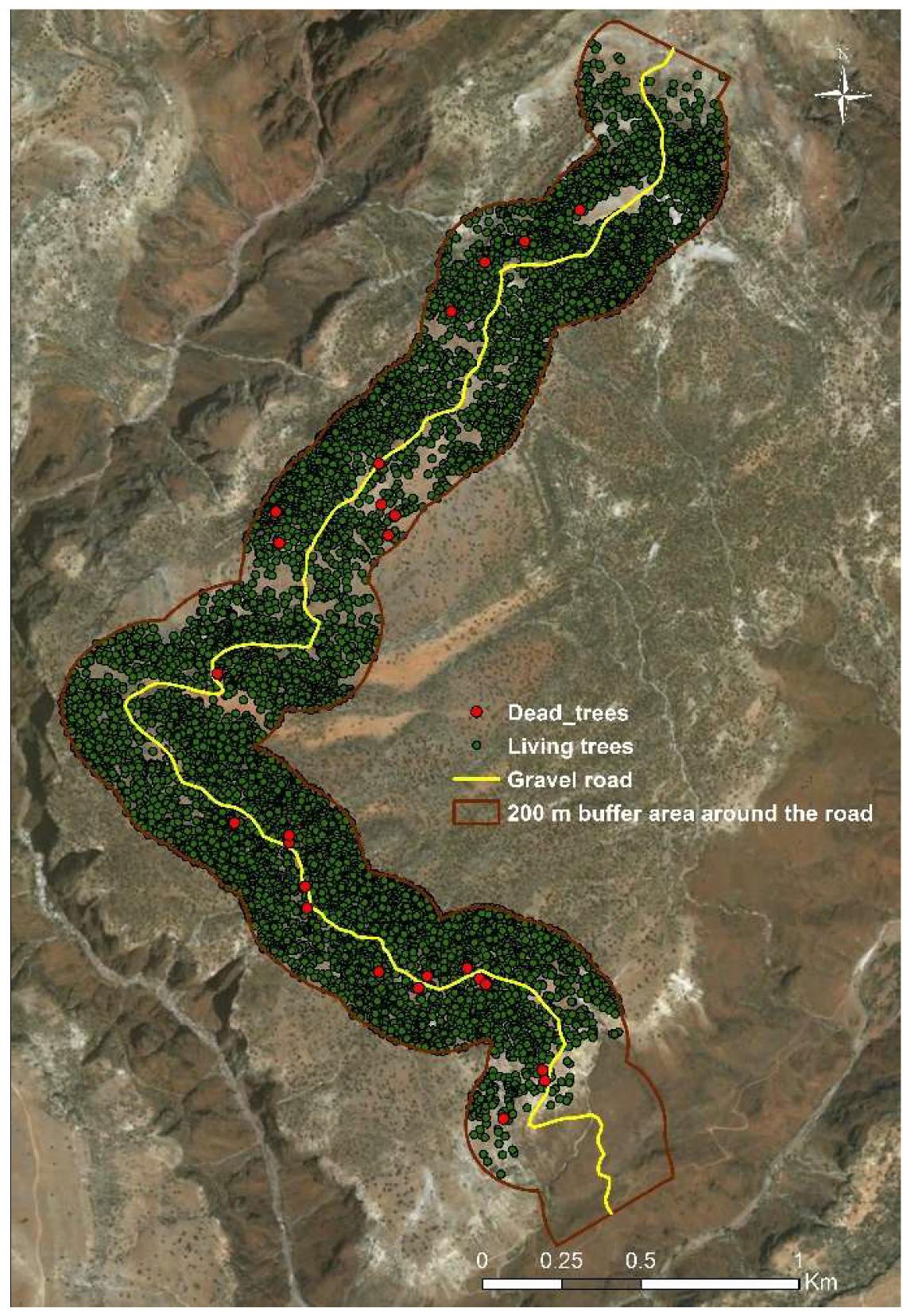
2.2. Accuracy of Abundance Estimation
2.3. Growth Characteristics and Age Estimation
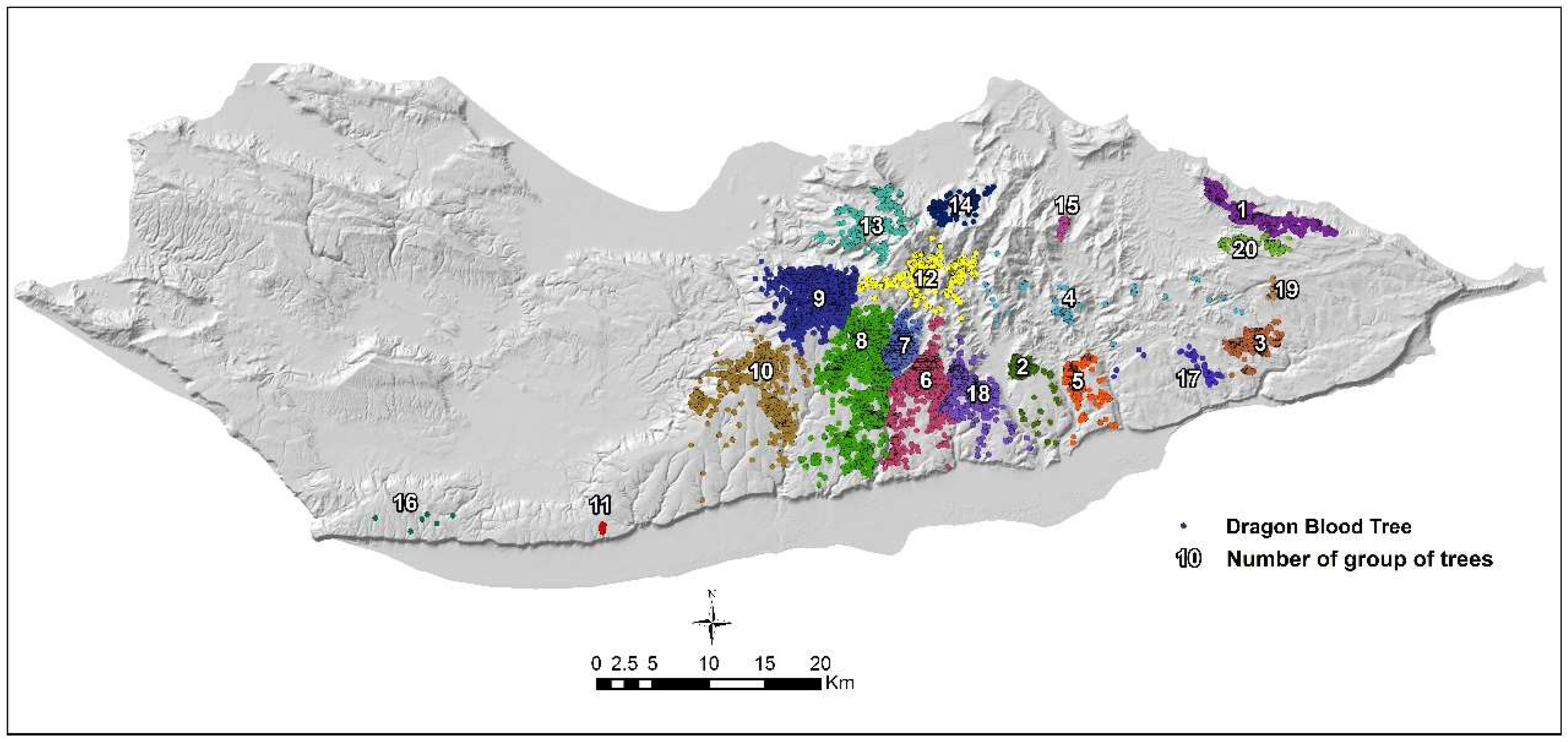
2.4. Sub-Population Delimitation
2.5. Modelling of Sub-Population Extinction Times
2.6. Sub-Population Conservation Importance Evaluation
3. Results
3.1. Abundance of Dragon’s Blood Tree Population on Socotra Island
3.2. Spatial Distribution of Dragon’s Blood Tree Population on Socotra Island
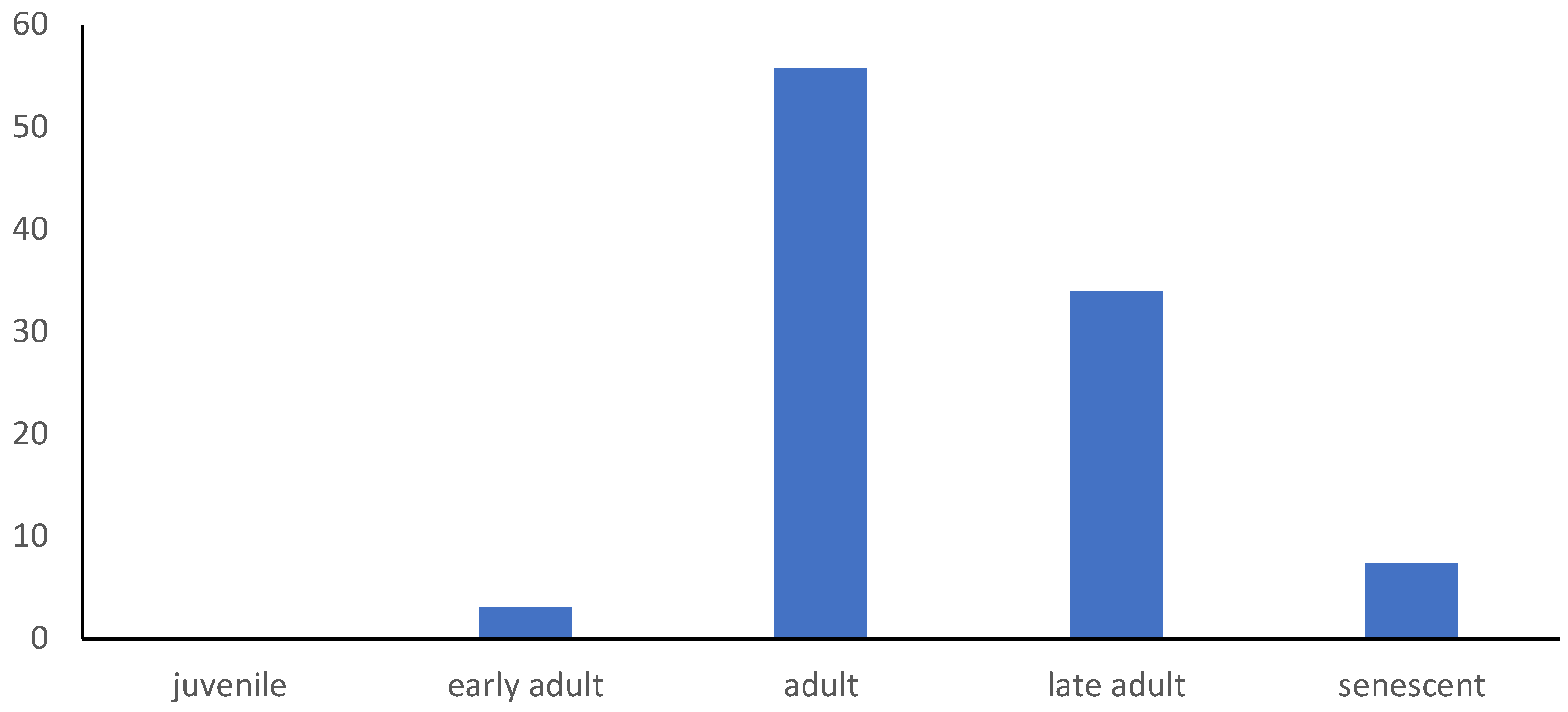
3.3. Age Structure of Dracaena cinnabari Population on Socotra Island
3.4. Prediction of Dragon’s Blood Tree Extinction
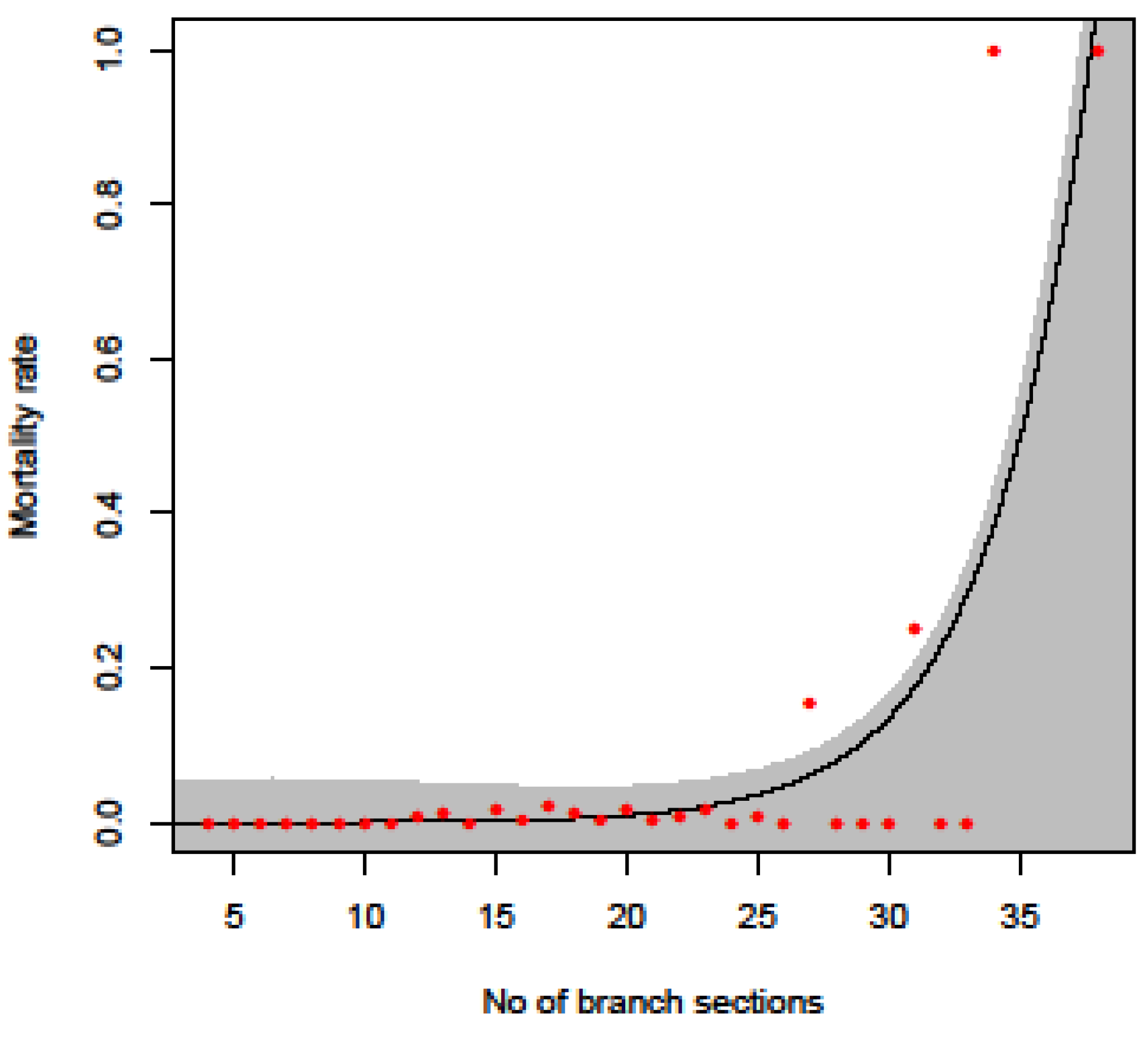
3.4.1. Mortality Curve
3.4.2. Extinction Time of Dragon’s Blood Tree Sub-Populations
3.5. The Conservation Importance of the Dragon’s Blood Tree Sub-Populations on Socotra Island
3.6. Sustainable Land Management Measures to Conserve Dragon’s Blood Tree Population
- a forest nursery to produce tree seedlings from local tree species populations;
- a trial demonstration focused on the presentation of the advantages of rotational pastures;
- a trial demonstration focused on reforestation using artificial regeneration (Figure S1);
- a trial agro-forestry demonstration focused on dragon’s blood tree natural regeneration together with Aloe perryi plantation.
4. Discussion
4.1. Conservation Status
4.2. Driving Forces of Dragon’s Blood Tree Population Decline
4.3. Extinction Model for the Dragon’s Blood Tree
4.4. Suggestions for Sustainable Land Management Measures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Culek, M. Geological and morphological evolution of the Socotra archipelago (Yemen) from the biogeographical view. J. Landsc. Ecol. 2013, 6, 84–108. [Google Scholar] [CrossRef]
- Denk, T.; Güner, H.T.; Grimm, G.W. From mesic to arid: Leaf epidermal features suggest preadaptation in Miocene dragon trees (Dracaena). Rev. Paleobotany Palynol. 2014, 200, 211–228. [Google Scholar] [CrossRef]
- Scholte, P.; De Geest, P. The climate of Socotra Island (Yemen). A first-time assessment of the timing of the monsoon wind reversal and its influence on precipitation and vegetation patterns. J. Arid Environ. 2010, 74, 1507–1515. [Google Scholar] [CrossRef]
- Miller, A.G.; Morris, M.; Diccon, A.; Atkinson, R. Ethnoflora of the Soqotra Archipelago, 1st ed.; Royal Botanic Garden: Edinburgh, UK, 2004; p. 759. [Google Scholar]
- Brown, G.; Mies, B.A. Vegetation Ecology of Socotra, 1st ed.; Plant and Vegetation 7; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2012; p. 379. [Google Scholar]
- De Sanctis, M.; Adeeb, A.; Farcomeni, A.; Patriarca, C.; Saed, A.; Attorre, F. Classification and distribution patterns of plant communities on Socotra Island, Yemen. Appl. Veg. Sci. 2013, 16, 148–165. [Google Scholar] [CrossRef]
- Habrová, H.; Buček, A. Overview of biotope types of Socotra island. J. Landsc. Ecol. 2013, 6, 60–83. [Google Scholar] [CrossRef]
- Kürschner, H.; Hein, P.; Kilian, N.; Hubaishan, M.A. Diversity and zonation of the forests and woodlands of the mountains of northern Socotra, Yemen. Englera 2006, 28, 11–55. [Google Scholar] [CrossRef]
- Kilian, N.; Hein, P. New and noteworthy records for the vascular plant flora of Socotra Island, Yemen. Englera 2006, 28, 57–77. [Google Scholar] [CrossRef]
- Řepka, R.; Maděra, P.; Čermák, M.; Forrest, A. Carex socotrana, a New Endemic Species from Socotra Island. Novon 2017, 25, 467–472. [Google Scholar] [CrossRef]
- Marrero, A.; Almeida, S.R.; Martín-González, M. A new species of the wild Dragon Tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic Implications. Bot. J. Linn. Soc. 1998, 128, 291–314. [Google Scholar]
- Adolt, R.; Pavliš, J. Age structure and growth of Dracaena cinnabari populations on Socotra. Trees—Struct. Funct. 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Habrová, H.; Čermák, Z.; Pavliš, J. Dragon’s blood tree—Threatened by overmaturity, not by extinction: Dynamics of a Dracaena cinnabari woodland in the mountains of Soqotra. Biol. Conserv. 2009, 142, 772–778. [Google Scholar] [CrossRef]
- IUCN. IUCN Red List of Threatened Species. Version 2017.2. Available online: htttp://www.iucnredlist.org (accessed on 25 October 2017).
- Van Damme, K.; Banfield, L. Past and present human impacts on the biodiversity of Socotra Island (Yemen): Implications for future conservation. Zool. Middle East 2011, 54, 31–88. [Google Scholar] [CrossRef]
- Rejžek, M.; Svátek, M.; Šebesta, J.; Adolt, R.; Maděra, P.; Matula, R. Loss of a single tree species will lead to an overall decline in plant diversity: Effect of Dracaena cinnabari Balf. f. on the vegetation of Socotra Island. Biol. Conserv. 2016, 196, 165–172. [Google Scholar] [CrossRef]
- García, C.; Vasconcelos, R. The beauty and the beast: Endemic mutualistic interactions promote community-based conservation on Socotra Island (Yemen). J. Nat. Conserv. 2017, 35, 20–23. [Google Scholar] [CrossRef]
- González-Castro, A.; Pérez-Pérez, D.; Romero, J.; Nogales, M. Unraveling the Seed Dispersal System of an Insular “Ghost” Dragon Tree (Dracaena draco) in the Wild. Front. Ecol. Evol. 2019, 7, 39. [Google Scholar] [CrossRef]
- Batelka, J. Socotra Archipelago—A lifeboat in the sea of changes: Advancement in Socotran insect biodiversity survey. Acta Entomol. Musei Natl. Pragae 2012, 52 (Suppl. 2), 1–26. [Google Scholar]
- Nadezhdina, N.; Al-Okaishi, A.; Madera, P. Sap Flow Measurements in a Socotra Dragon’s Blood Tree (Dracaena cinnabari) in its Area of Origin. Trop. Plant Biol. 2018, 11, 107–118. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Eltahir, E.A.B. Forest on the edge: Seasonal cloud forest in Oman creates its own ecological niche. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef]
- Attorre, F.; Francesconi, F.; Taleb, N.; Scholte, P.; Saed, A.; Alfo, M.; Bruno, F. Will dragonblood survive the next period of climate change? Current and future potential distribution of Dracaena cinnabari (Socotra, Yemen). Biol. Conserv. 2007, 138, 430–439. [Google Scholar] [CrossRef]
- Hubálková, I.; Maděra, P.; Volařík, D. Growth dynamics of Dracaena cinnabari under controlled conditions as the most effective way to protect endangered species. Saudi J. Biol. Sci. 2017, 24, 1445–1452. [Google Scholar] [CrossRef]
- Habrová, H.; Pavliš, J. Dynamic response of woody vegetation on fencing protection in semi-arid areas; Case study: Pilot exclosure on the Firmihin Plateau, Socotra island. Saudi J. Biol. Sci. 2017, 24, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.D.; Yokoya, N.; Bui, D.T.; Yoshino, K.; Friess, D.A. Remote Sensing Approaches for Monitoring Mangrove Species, Structure, and Biomass: Opportunities and Challenges. Remote Sens. 2019, 11, 230. [Google Scholar] [CrossRef]
- Pettorelli, N.; Schulte to Bühne, H.; Tulloch, A.; Dubois, G.; Macinnis-Ng, C.; Queirós, A.M.; Keith, D.A.; Wegmann, M.; Schrodt, F.; Stellmes, M.; et al. Satellite remote sensing of ecosystem functions: Opportunities, challenges and way forward. Remote Sens. Ecol. Conserv. 2018, 4, 71–93. [Google Scholar] [CrossRef]
- Gougeon, F.A.; Leckie, D.G. Forest Information Extraction from High Spatial Resolution Images Using an Individual Tree Crown Approach, 1st ed.; Canadian Forest Service: Victoria, BC, Canada, 2003; p. 27. [Google Scholar]
- Gomes, M.F.; Maillard, P. Detection of Tree Crowns in Very High Spatial Resolution Images. In Environmental Applications of Remote Sensing; Marghany, M., Ed.; InTech: London, UK, 2016; pp. 41–71. [Google Scholar] [CrossRef]
- Attorre, F.; Issa, A.; Malatesta, L.; Adeeb, A.; De Sanctis, M.; Vitale, M.; Farcomeni, A. Analysing the relationship between land units and plant communities: The case of Socotra Island (Yemen). Plant Biosyst. 2014, 148, 529–539. [Google Scholar] [CrossRef]
- Malatesta, L.; Attorre, F.; Altobelli, A.; Adeeb, A.; De Sanctis, M.; Taleb, N.M.; Scholte, P.T.; Vitale, M. Vegetation mapping from high-resolution satellite images in the heterogeneous arid environments of Socotra Island (Yemen). J. Appl. Remote Sens. 2013, 7, 073527. [Google Scholar] [CrossRef]
- Habrová, H. Geobiocoenological differentiation as a tool for sustainable land-use of Socotra Island. Ekológia (Bratislava) 2004, 23, 47–57. [Google Scholar]
- Král, K.; Pavliš, J. The first detailed land cover map of Socotra Island by Landsat /ETM+ data. Int. J. Remote Sens. 2006, 27, 3239–3250. [Google Scholar] [CrossRef]
- Adolt, R.; Maděra, P.; Abraham, J.; Čupa, P.; Svátek, M.; Matula, R.; Šebesta, J.; Čermák, M.; Volařík, D.; Koutecký, T.; et al. Field survey of Dracaena cinnabari populations in Firmihin, Socotra island: Methodology and preliminary results. J. Landsc. Ecol. 2013, 6, 7–34. [Google Scholar] [CrossRef]
- Adolt, R.; Habrová, H.; Maděra, P. Crown age estimation of a monocotyledonous tree species Dracaena cinnabari using logistic regression. Trees—Struct. Funct. 2012, 26, 1287–1298. [Google Scholar] [CrossRef]
- Hubálková, I. Prediction of Dragon’s Blood Tree (Dracaena cinnabari Balf.) stand sample density on Soqotra Island. J. Landsc. Ecol. 2011, 4, 5–17. [Google Scholar] [CrossRef]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Hubálková, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Naděždina, N.; Němec, P.; Rosenthal, J.; et al. Growth dynamics of endemic Dracaena cinnabari of Socotra Island suggest essential elements for a conservation strategy. Biologia 2019, 74, 339–349. [Google Scholar] [CrossRef]
- Zheng, D.J.; Xie, L.S.; Zhu, J.H.; Zhang, Z.L. Low genetic diversity and local adaptive divergence of Dracaena cambodiana (Liliaceae) populations associated with historical population bottlenecks and natural selection: An endangered long-lived tree endemic to Hainan Island, China. Plant Biol. 2012, 14, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.L.; Zhang, L.; Dayanandan, S.; Nagaraju, S.; Liu, D.M.; Li, Q.M. Tertiary origin and pleistocene diversification of dragon blood tree (Dracaena cambodiana-Asparagaceae) populations in the Asian tropical forests. PLoS ONE 2013, 8, e60102. [Google Scholar] [CrossRef]
- Kamel, M.; Ghazaly, U.M.; Callmander, M.W. Conservation status of the Endangered Nubian dragon tree Dracaena ombet in Gebel Elba National Park, Egypt. Oryx 2015, 49, 704–709. [Google Scholar] [CrossRef]
- Elnoby, S.K.; Moustafa, A.A.; Mansour, S.R. Impact of climate change on the endangered Nubian dragon tree (Dracaena ombet) in the South Eastern of Egypt. Catrina 2017, 16, 23–28. [Google Scholar]
- Almeida Pérez, R.S. Censo, distribución, habitat y estado de conservation de Dracaena tamaranae A.Marrero, R.S.González-Martín. Gran Canaria, Islas Canarias. Botanica Macarónesica 2003, 24, 39–56. [Google Scholar]
- Almeida Pérez, R.S. Dracaena draco (L.) L. In Atlas y Libro Rojo de la Flora Vascular Amenazada de España, 2nd ed.; Bañares, A., Blanca, G., Güemes, J., Moreno, J.C., Ortiz, S., Eds.; Publicaciones de O.A.P.N.: Madrid, Spain, 2004; pp. 680–681. [Google Scholar]
- Wilkin, P.; Suksathan, P.; Keeratikiat, K.; Van Welzen, P.; Wiland-Szymanska, J. A new species from Thailand and Burma, Dracaena kaweesakii Wilkin & Suksathan (Asparagaceae subfamily Nolinoideae). PhytoKeys 2013, 26, 101–112. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 25 December 2018).
- Caswell, H. Matrix Population Models: Construction, Analysis, and Interpretation, 2nd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001; p. 722. [Google Scholar]
- FAO. Farmer Field School Guidance Document. Planning for Quality Programmes; Food and Agriculture Organization of the United Nations: Roma, Italy, 2016; p. 112. [Google Scholar]
- Scholte, P.; Miller, A.; Shamsan, A.R.; Suleiman, A.S.; Taleb, N.; Millroy, T.; Attorre, F.; Porter, R.; Carugati, C.; Pella, F. Goats: Part of the Problem or the Solution to Biodiversity Conservation on SOCOTRA? Technical Report; SCDP: Hadibo, Socotra, Republic of Yemen, 2007; p. 11. [Google Scholar]
- Edward, H.G.M.; de Oliveira, L.F.C.; Quye, A. Raman spectroscopy of coloured resins used in antiquity: dragon’s blood and related substances. Spectrochim. Acta Part A 2001, 57, 2831–2842. [Google Scholar] [CrossRef]
- Miller, A. Dracaena cinnabari. Available online: http://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2004.RLTS.T30428A9548491.en (accessed on 27 December 2018).
- Lavranos, J.J. A new, arborescent subspecies of Dracaena from Saudi Arabia. Cactus Succul. J. 2017, 89, 148–152. [Google Scholar] [CrossRef]
- Van Rampelbergh, M.; Fleitmann, D.; Verheyden, S.; Cheng, H.; Edwards, L.; De Geest, P.; De Vleeschouwer, D.; Burns, J.S.; Matter, A.; Claeys, P.; et al. Mid- to late holocene Indian ocean monsoon variability recorded in four speleothems from Socotra island, Yemen. Quat. Sci. Rev. 2013, 65, 129–142. [Google Scholar] [CrossRef]
- Akçakaya, H.R.; Sjögren-Gulve, P. Population viability analysis in conservation planning: An overview. Ecol. Bull. 2000, 48, 9–21. [Google Scholar]
- Kappler, R.H.; Knight, K.S.; Root, K.V. Evaluating the population viability of green ash trees (Fraxinus pennsylvanica) before and after the emerald ash borer beetle (Agrilus planipennis) invasion. Ecol. Model. 2019, 400, 53–59. [Google Scholar] [CrossRef]
- Davelos, A.L.; Jarosz, A.M. Demography of american chestnut populations: Effects of a pathogen and a hyperparasite. J. Ecol. 2004, 92, 675–685. [Google Scholar] [CrossRef]
- Dhar, A.; Ruprecht, H.; Vacik, H. Population viability risk management (PVRM) for in situ management of endangered tree species—a case study on a Taxus baccata L. population. For. Ecol. Manag. 2008, 255, 2835–2845. [Google Scholar] [CrossRef]
- Grogan, J.; Landis, R.M.; Free, C.M.; Schulze, M.D.; Lentini, M.; Ashton, M.S. Bigleaf mahogany Swietenia macrophylla population dynamics and implications for sustainable management. J. Appl. Ecol. 2014, 51, 664–674. [Google Scholar] [CrossRef]
- Van Rensburg, J.J.; Hopper, K. Incense and imagery: Mapping agricultural and water management systems on the island of Socotra, Yemen. Proc. Semin. Arab. Stud. 2017, 47, 129–138. [Google Scholar]
- Attorre, F.; Taleb, N.; De Sanctis, M.; Farcomeni, A.; Guillet, A.; Vitale, M. Developing conservation strategies for endemic tree species when faced with time and data constraints: Boswellia spp. on Socotra (Yemen). Biodivers. Conserv. 2011, 20, 1483–1499. [Google Scholar] [CrossRef]
- Lvončík, S.; Maděra, P.; Volařík, D.; Vrškový, B.; Habrová, H. First proposal of seed regions for frankincense trees (Boswellia spp.) on Socotra Island. J. Landsc. Ecol. 2013, 6, 35–45. [Google Scholar] [CrossRef]
- Maděra, P.; Paschová, Z.; Ansorgová, A.; Vrškový, B.; Lvončík, S.; Habrová, H. Volatile compounds in oleo-gum resin of Socotran species of Burseraceae. Acta Universitatis Agriculturae Et Silviculturae Mendelianae Brunensis 2017, 65, 73–90. [Google Scholar] [CrossRef]
- Ceccolini, L. The homegardens of Soqotra island, Yemen: An example of agroforestry approach to multiple land-use in an isolated location. Agrofor. Syst. 2002, 56, 107–115. [Google Scholar] [CrossRef]
- Nováková, P. Evaluation of Success of Native Trees Plantation in Home-gardens on Socotra. Bachelor’s Thesis, Mendel University, Brno, Czech Republic, 2015; p. 83. [Google Scholar]
- Gauld, R. Maintaining Centralized Control in Community-based Forestry: Policy Construction in the Philippines. Dev. Chang. 2000, 31, 229–254. [Google Scholar] [CrossRef]
- Maryudi, A.; Devkota, R.R.; Schusser, C.; Yufanyi, C.; Salla, M. Back to basics: Considerations in evaluating the outcomes of community forestry. For. Policy Econ. 2012, 14, 1–5. [Google Scholar] [CrossRef]
- Baynes, J.; Herbohn, J.; Smith, C.; Fisher, R.; Braye, D. Key factors which influence the success of community forestry in developing countries. Glob. Environ. Chang. 2015, 35, 226–238. [Google Scholar] [CrossRef]
- Rametsteiner, E.; Simula, M. Forest certification—An instrument to promote sustainable forest management? J. Environ. Manag. 2003, 67, 87–98. [Google Scholar] [CrossRef]
- Gullison, R.E. Does forest certification conserve biodiversity? Oryx 2003, 37, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Aynekulu, E. Forest Diversity in Fragmented Landscapes of Northern Ethiopia and Implications for Conservation. Ph.D. Thesis, University of Bonn, Bonn, Germany, 2011; p. 132. [Google Scholar]
- Aynekulu, E.; Aerts, R.; Moonen, P.; Denich, M.; Gebrehiwot, K.; Vagen, T.G.; Mekuria, W.; Boehmer, H.J. Altitudinal variation and conservation priorities of vegetation along the Great Rift Valley escarpment, northern Ethiopia. Biodivers. Conserv. 2012, 21, 2691–2707. [Google Scholar] [CrossRef]
- Marrero, A.; Almeida, R.S.; Roca, A. Diáspora y rescate: Acciones para la conservación del Drago de Gran Canaria, Dracaena tamaranae. In Proceedings of the V Congreso de Biología de la Conservación de Plantas, Menorca, Es Mercadal, Spain, 28 September–1 October 2011. [Google Scholar]
- Al Hosni, A.; Oliver, I.; Al Jabri, I.; Al Saidi, A.; Al Rawahi, A.; Al Hinai, H. Ex situ conservation of Dracaena serrulata in Dhofar province, southern Oman. Acta Hortic. 2018, 1190, 9–14. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landcover class. | CN |
|---|---|
| Dracaena forest | 1.0 |
| Frankincense forest | 1.0 |
| Montane forest | 1.0 |
| Dracaena woodland | 0.6 |
| Frankincense woodland | 0.6 |
| Montane mosaic | 0.6 |
| Submontane shrubland | 0.4 |
| High shrubland with succulents | 0.4 |
| Low Croton-Jatropha shrubland | 0.3 |
| Sparse dwarf shrubland | 0.2 |
| Submontane grassland | 0.2 |
| Montane grassland | 0.2 |
| Limestone rocks | 1.0 |
| Basement rocks | 1.0 |
| Wadi | 0.1 |
| Date palm plantations | 0.0 |
| Sub-Pop. No | Area [ha] | No. of Trees | Tree Density [tree per ha] | Mean Elevation [m a.s.l.] |
|---|---|---|---|---|
| 1 | 1635.73 | 3553 | 2.17 | 482 |
| 2 | 1675.31 | 1392 | 0.83 | 476 |
| 3 | 923.91 | 1872 | 2.03 | 358 |
| 4 | 4873.31 | 223 | 0.05 | 465 |
| 5 | 1900.13 | 2867 | 1.51 | 427 |
| 6 | 4383.35 | 10 890 | 2.48 | 530 |
| 7 | 1089.69 | 32 195 | 29.55 | 638 |
| 8 | 7952.07 | 9343 | 1.17 | 611 |
| 9 | 4487.63 | 7005 | 1.56 | 900 |
| 10 | 10 408.80 | 1252 | 0.12 | 745 |
| 11 | 11.19 | 22 | 1.97 | 513 |
| 12 | 3105.48 | 1089 | 0.35 | 744 |
| 13 | 2632.81 | 719 | 0.27 | 771 |
| 14 | 1162.84 | 311 | 0.27 | 630 |
| 15 | 77.21 | 111 | 1.44 | 479 |
| 16 | 462.07 | 15 | 0.03 | 549 |
| 17 | 871.86 | 81 | 0.09 | 403 |
| 18 | 3714.82 | 6551 | 1.76 | 445 |
| 19 | 48.45 | 76 | 1.57 | 495 |
| 20 | 546.52 | 565 | 1.03 | 475 |
| Landcover Class | No. of Trees | Area [ha] | Tree Density [tree per ha] |
|---|---|---|---|
| Dracaena forests | 16,979 | 240.00 | 70.75 |
| Dracaena woodland | 26,869 | 4883.54 | 5.50 |
| Limestone rocks | 59 | 19.96 | 2.96 |
| Submontane grassland | 17,043 | 15,413.97 | 1.11 |
| Submontane shrubland | 11,483 | 14,829.53 | 0.77 |
| High shrubland with succulents | 4596 | 7899.90 | 0.58 |
| Montane forest | 119 | 214.87 | 0.55 |
| Frankincense woodland | 1762 | 4049.95 | 0.44 |
| Frankincense forest | 356 | 927.21 | 0.38 |
| Montane mosaic | 518 | 1603.33 | 0.32 |
| Montane grassland | 82 | 329.00 | 0.25 |
| Sparse dwarf shrubland | 25 | 103.14 | 0.24 |
| Low Croton-Jatropha shrubland | 233 | 1211.76 | 0.19 |
| Wadi | 9 | 78.55 | 0.11 |
| Basement rocks | 6 | 67.28 | 0.09 |
| Date palm plantations | 0 | 72.36 | 0.00 |
| Sub-Pop. No | Mean Age [years] | SD | SE | Min [years] | Max [years] | Juve-Nile [%] | Early Adult [%] | Adult [%] | Late Adult [%] | Sene- Scent [%] | Extinction Time [years] |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 299 | 87.01 | 1.46 | 0 | 545 | 0.03 | 4.28 | 69.43 | 22.63 | 3.63 | 509 |
| 2 | 307 | 86.00 | 2.31 | 56 | 529 | 0.00 | 3.52 | 65.88 | 26.51 | 4.09 | 451 |
| 3 | 335 | 80.44 | 1.86 | 56 | 513 | 0.00 | 2.08 | 54.01 | 38.14 | 5.77 | 442 |
| 4 | 200 | 89.95 | 6.02 | 56 | 428 | 0.00 | 29.15 | 65.47 | 5.38 | 0.00 | 470 |
| 5 | 332 | 81.65 | 1.52 | 56 | 521 | 0.00 | 1.95 | 55.14 | 37.32 | 5.58 | 461 |
| 6 | 346 | 87.46 | 0.84 | 56 | 552 | 0.00 | 2.18 | 47.14 | 39.70 | 10.98 | 506 |
| 7 | 327 | 82.27 | 0.46 | 56 | 559 | 0.00 | 2.45 | 58.11 | 34.48 | 4.96 | 546 |
| 8 | 352 | 87.98 | 0.91 | 27 | 545 | 0.00 | 1.51 | 47.00 | 36.68 | 14.81 | 495 |
| 9 | 282 | 89.35 | 1.07 | 27 | 529 | 0.00 | 7.35 | 71.83 | 18.93 | 1.88 | 564 |
| 10 | 355 | 89.38 | 2.53 | 56 | 545 | 0.00 | 1.36 | 47.04 | 34.27 | 17.33 | 395 |
| 11 | 355 | 46.08 | 9.82 | 238 | 438 | 0.00 | 0.00 | 45.45 | 54.55 | 0.00 | 117 |
| 12 | 230 | 109.66 | 3.32 | 27 | 521 | 0.00 | 24.33 | 60.33 | 11.75 | 3.58 | 545 |
| 13 | 314 | 97.18 | 3.62 | 56 | 529 | 0.00 | 6.82 | 57.02 | 28.23 | 7.93 | 428 |
| 14 | 364 | 85.28 | 4.84 | 27 | 529 | 0.00 | 1.93 | 41.16 | 39.87 | 17.04 | 313 |
| 15 | 343 | 70.33 | 6.68 | 108 | 504 | 0.00 | 0.90 | 54.05 | 37.84 | 7.21 | 226 |
| 16 | 466 | 61.20 | 15.80 | 353 | 559 | 0.00 | 0.00 | 7.14 | 21.43 | 71.43 | 31 |
| 17 | 388 | 63.43 | 7.05 | 198 | 504 | 0.00 | 0.00 | 22.22 | 59.26 | 18.52 | 162 |
| 18 | 350 | 78.35 | 0.97 | 27 | 545 | 0.00 | 0.72 | 48.05 | 41.87 | 9.36 | 461 |
| 19 | 363 | 71.67 | 8.22 | 177 | 504 | 0.00 | 0.00 | 40.79 | 51.32 | 7.89 | 194 |
| 20 | 365 | 74.76 | 3.15 | 27 | 537 | 0.00 | 0.35 | 42.48 | 43.54 | 13.63 | 311 |
| Sub-Pop. No | HD | AM | DT | SP | TN | CI | CI rel |
|---|---|---|---|---|---|---|---|
| 7 | 0.58 | 0.52 | 0.59 | 0.10 | 1.00 | 2.79 | 55.90 |
| 4 | 0.42 | 1.00 | 0.00 | 0.47 | 0.01 | 1.90 | 37.97 |
| 8 | 0.38 | 0.43 | 0.02 | 0.76 | 0.29 | 1.88 | 37.66 |
| 10 | 0.40 | 0.42 | 0.00 | 1.00 | 0.04 | 1.86 | 37.22 |
| 9 | 0.44 | 0.69 | 0.03 | 0.43 | 0.22 | 1.81 | 36.28 |
| 12 | 0.55 | 0.89 | 0.01 | 0.30 | 0.03 | 1.77 | 35.48 |
| 6 | 0.37 | 0.45 | 0.05 | 0.42 | 0.34 | 1.63 | 32.66 |
| 13 | 0.66 | 0.57 | 0.01 | 0.25 | 0.02 | 1.51 | 30.19 |
| 18 | 0.37 | 0.44 | 0.04 | 0.36 | 0.20 | 1.40 | 28.04 |
| 1 | 0.39 | 0.63 | 0.04 | 0.16 | 0.11 | 1.32 | 26.47 |
| 14 | 0.66 | 0.38 | 0.01 | 0.11 | 0.01 | 1.16 | 23.30 |
| 5 | 0.36 | 0.50 | 0.03 | 0.18 | 0.09 | 1.16 | 23.26 |
| 2 | 0.33 | 0.60 | 0.02 | 0.16 | 0.04 | 1.15 | 23.03 |
| 3 | 0.37 | 0.49 | 0.04 | 0.09 | 0.06 | 1.05 | 21.03 |
| 20 | 0.43 | 0.38 | 0.02 | 0.05 | 0.02 | 0.90 | 18.02 |
| 15 | 0.38 | 0.46 | 0.03 | 0.01 | 0.00 | 0.88 | 17.66 |
| 19 | 0.38 | 0.39 | 0.03 | 0.00 | 0.00 | 0.80 | 16.00 |
| 11 | 0.30 | 0.42 | 0.04 | 0.00 | 0.00 | 0.76 | 15.13 |
| 17 | 0.32 | 0.29 | 0.00 | 0.08 | 0.00 | 0.70 | 14.07 |
| 16 | 0.30 | 0.00 | 0.00 | 0.04 | 0.00 | 0.35 | 6.93 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable Land Use Management Needed to Conserve the Dragon’s Blood Tree of Socotra Island, a Vulnerable Endemic Umbrella Species. Sustainability 2019, 11, 3557. https://0-doi-org.brum.beds.ac.uk/10.3390/su11133557
Maděra P, Volařík D, Patočka Z, Kalivodová H, Divín J, Rejžek M, Vybíral J, Lvončík S, Jeník D, Hanáček P, et al. Sustainable Land Use Management Needed to Conserve the Dragon’s Blood Tree of Socotra Island, a Vulnerable Endemic Umbrella Species. Sustainability. 2019; 11(13):3557. https://0-doi-org.brum.beds.ac.uk/10.3390/su11133557
Chicago/Turabian StyleMaděra, Petr, Daniel Volařík, Zdeněk Patočka, Hana Kalivodová, Josef Divín, Martin Rejžek, Jan Vybíral, Samuel Lvončík, David Jeník, Pavel Hanáček, and et al. 2019. "Sustainable Land Use Management Needed to Conserve the Dragon’s Blood Tree of Socotra Island, a Vulnerable Endemic Umbrella Species" Sustainability 11, no. 13: 3557. https://0-doi-org.brum.beds.ac.uk/10.3390/su11133557