Characterization of Venom Components and Their Phylogenetic Properties in Some Aculeate Bumblebees and Wasps

 ,
,
Abstract
:1. Introduction
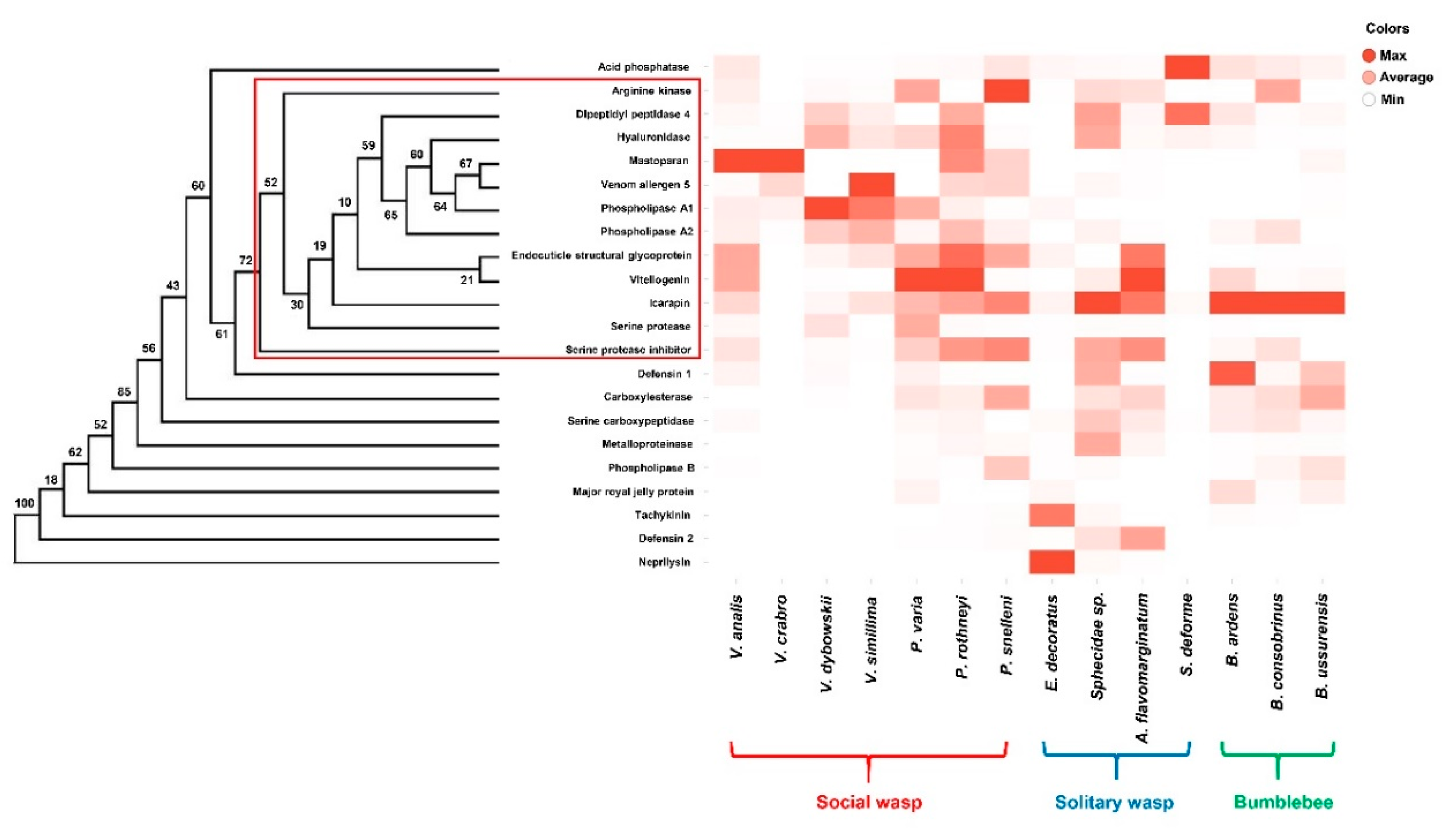
2. Results and Discussion
2.1. Differential Transcriptional Profiles of the Venom Glands of Some Aculeate Species
2.1.1. Venom Proteins Highly Expressed in Solitary Hunting Wasps
2.1.2. Venom Proteins Highly Expressed in Social Wasps
2.1.3. Venom Proteins Highly Expressed in Bumblebees
2.1.4. Expression Patterns of Other Venom Proteins
2.2. Identification of Putatively Novel Venom Components in Some Aculeate Bees and Wasps
Properties of the Six USPs (USP1–USP6) as Putative Allergenic Proteins
2.3. Evolutionary Patterns in the Venom of Some Aculeate Bees and Wasps
3. Conclusions
4. Materials and Methods
4.1. Wasp and Bumblebee Collection and Total RNA Purification
4.2. Construction of an in Silico cDNA Library and Computational Analysis
4.3. Ortholog Analysis of Venom Component Genes
4.4. Construction of a Phylogenetic Tree
4.5. Quantitative Real-Time PCR
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Piek, T. Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Gauld, I.D.; Bolton, B.; Huddleston, T.; Fitton, M.; Shaw, M.R.; Noyes, J.; Day, M.; Else, G.; Ferguson, N.; Ward, S. The hymenoptera. Hymenoptera 1988, 1988, 1–332. [Google Scholar]
- Dowton, M. Simultaneous analysis of 16S, 28S, COI and morphology in the Hymenoptera: Apocrita—Evolutionary transitions among parasitic wasps. Biol. J. Linn. Soc. 2001, 74, 87–111. [Google Scholar] [CrossRef]
- O’Neill, K.M. Solitary Wasps: Behavior and Natural History; Cornell University Press: Ithaca, NY, USA, 2001. [Google Scholar]
- Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps. Toxins 2016, 8, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, H.; Eberhard, M.J. The Wasps; University of Michigan Press: Ann Arbor, MI, USA, 1971. [Google Scholar]
- Schmidt, J.O. Insect Defenses: Adaptive Mechanisms and Strategies of Prey and Predators; SUNY Press: Albany, NY, USA, 1990. [Google Scholar]
- Akre, R.D. Nesting biology and behavior of the baldfaced hornet, Dolichovespula maculata (L.)(Hymenoptera: Vespidae) in the Pacific Northwest. Melanderia 1992, 48, 1–33. [Google Scholar]
- Jeanne, R.L.; Williams, N.M.; Yandell, B.S. Age polyethism and defense in a tropical social wasp (Hymenoptera: Vespidae). J. Insect Behav. 1992, 5, 211–227. [Google Scholar] [CrossRef] [Green Version]
- Spradbery, J.P. Wasps. In An Account of the Biology and Natural History of Social and Solitary Wasps, with Particular Reference to Those of the British Isles; Sidgwick & Jackson: London, UK, 1973. [Google Scholar]
- Richter, M.R. Social wasp (Hymenoptera: Vespidae) foraging behavior. Annu. Rev. Entomol. 2000, 45, 121–150. [Google Scholar] [CrossRef]
- Eno, A.E. Pharmacological investigation of oedema induced by venom from the wasp Polistes fuscatus. Toxicon 1997, 35, 1691–1698. [Google Scholar] [CrossRef]
- Fisher, B.A.; Antonios, T.F. Atrial flutter following a wasp sting. J. Postgrad. Med. 2003, 49, 254–255. [Google Scholar]
- Ferrari, S.; Pietroiusti, A.; Galanti, A.; Compagnucci, M.; Fontana, L. Paroxysmal atrial fibrillation after insect sting. J. Allergy Clin. Immunol. 1996, 98, 759–761. [Google Scholar] [CrossRef]
- Sachdev, A.; Mahapatra, M.; D’Cruz, S.; Kumar, A.; Singh, R.; Lehl, S.S. Wasp sting induced neurological manifestations. Neurol. India 2002, 50, 319–321. [Google Scholar]
- Sharmila, R.R.; Chetan, G.; Narayanan, P.; Srinivasan, S. Multiple organ dysfunction syndrome following single wasp sting. Indian J. Pediatr. 2007, 74, 1111–1112. [Google Scholar] [CrossRef]
- Argiolas, A.; Pisano, J.J. Facilitation of Phospholipase-A2 Activity by Mastoparans, a New Class of Mast-Cell Degranulating Peptides from Wasp Venom. J. Biol. Chem. 1983, 258, 3697–3702. [Google Scholar]
- Krishnakumari, V.; Nagaraj, R. Antimicrobial and hemolytic activities of crabrolin, a 13-residue peptide from the venom of the European hornet, Vespa crabro, and its analogs. J. Pept. Res. 1997, 50, 88–93. [Google Scholar] [CrossRef]
- Bucher, C.; Korner, P.; Wüthrich, B. Allergy to bumblebee venom. Curr. Opin. Allergy Clin. Immunol. 2001, 1, 361–365. [Google Scholar] [CrossRef]
- Moreau, S.J.M.; Asgari, S. Venom Proteins from Parasitoid Wasps and Their Biological Functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.A.; Kim, K.; Nguyen, P.; Seo, J.B.; Park, Y.H.; Kim, K.G.; Seo, H.Y.; Koh, Y.H.; Lee, S.H. Comparative bioactivities of mastoparans from social hornets Vespa crabro and Vespa analis. J. Asia-Pac. Entomol. 2015, 18, 825–829. [Google Scholar] [CrossRef]
- Baek, J.H.; Lee, S.H. Identification and characterization of venom proteins of two solitary wasps, Eumenes pomiformis and Orancistrocerus drewseni. Toxicon 2010, 56, 554–562. [Google Scholar] [CrossRef]
- Sunagar, K.; Morgenstern, D.; Reitzel, A.M.; Moran, Y. Ecological venomics: How genomics, transcriptomics and proteomics can shed new light on the ecology and evolution of venom. J. Proteom. 2016, 135, 62–72. [Google Scholar] [CrossRef]
- Rokyta, D.R.; Margres, M.J.; Calvin, K. Post-transcriptional Mechanisms Contribute Little to Phenotypic Variation in Snake Venoms. G3-Genes Genom. Genet. 2015, 5, 2375–2382. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.Y.; Ye, G.Y.; Hu, C. Molecular cloning and characterization of acid phosphatase in venom of the endoparasitoid wasp Pteromalus puparum (Hymenoptera: Pteromalidae). Toxicon 2008, 51, 1391–1399. [Google Scholar] [CrossRef]
- Dani, M.P.; Edwards, J.P.; Richards, E.H. Hydrolase activity in the venom of the pupal endoparasitic wasp, Pimpla hypochondriaca. Comp. Biochem. Phys. B 2005, 141, 373–381. [Google Scholar] [CrossRef]
- Turner, A.J.; Isaac, R.E.; Coates, D. The neprilysin (NEP) family of zinc metalloendopeptidases: Genomics and function. Bioessays 2001, 23, 261–269. [Google Scholar] [CrossRef]
- Isaac, R.E. Neuropeptide-Degrading Endopeptidase Activity of Locust (Schistocerca-Gregaria) Synaptic-Membranes. Biochem. J. 1988, 255, 843–847. [Google Scholar] [CrossRef] [Green Version]
- Arvidson, R.; Kaiser, M.; Pan, S.Q.; Libersat, F.; Adams, M.E. Bioinformatic and Functional Analysis of Venom from the Jewel Wasp Ampulex compressa. Faseb J. 2016, 30, 819–821. [Google Scholar]
- Binder, M.; Mahler, V.; Hayek, B.; Sperr, W.R.; Scholler, M.; Prozell, S.; Wiedermann, G.; Valent, P.; Valenta, R.; Duchene, M. Molecular and immunological characterization of arginine kinase from the indianmeal moth, Plodia interpunctella, a novel cross-reactive invertebrate pan-allergen. J. Immunol. 2001, 167, 5470–5477. [Google Scholar] [CrossRef] [Green Version]
- Schiener, M.; Hilger, C.; Eberlein, B.; Pascal, M.; Kuehn, A.; Revets, D.; Planchon, S.; Pietsch, G.; Serrano, P.; Moreno-Aguilar, C.; et al. The high molecular weight dipeptidyl peptidase IV Pol d 3 is a major allergen of Polistes dominula venom. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Hino, K.; Yamamoto, S.; Sano, O.; Taniguchi, Y.; Kohno, K.; Usui, M.; Fukuda, S.; Hanzawa, H.; Haruyama, H.; Kurimoto, M. Carbohydrate Structures of the Glycoprotein Allergen Cry-J-I from Japanese Cedar (Cryptomeria-Japonica) Pollen. J. Biochem. 1995, 117, 289–295. [Google Scholar] [CrossRef]
- Borodina, I.; Jensen, B.M.; Wagner, T.; Hachem, M.A.; Sondergaard, I.; Poulsen, L.K. Expression of Enzymatically Inactive Wasp Venom Phospholipase A1 in Pichia pastoris. PLoS ONE 2011, 6, e21267. [Google Scholar] [CrossRef] [Green Version]
- Corthesy, B.; Jilek, S.; Barbey, C.; Spertini, F. Antigen-independent suppression of the allergic immune response to bee venom phospholipase A2 by DNA vaccination in CBA/J mice. J. Allergy Clin. Immunol. 2001, 107, S325. [Google Scholar]
- Hoffman, D.R.; Jacobson, R.S. Allergens in Hymenoptera venom. XXVII: Bumblebee venom allergy and allergens. J. Allergy Clin. Immunol. 1996, 97, 812–821. [Google Scholar] [CrossRef]
- Winningham, K.M.; Fitch, C.D.; Schmidt, M.; Hoffman, D.R. Hymenoptera venom protease allergens. J. Allergy Clin. Immunol. 2004, 114, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Golden, D.B.; Kelly, D.; Hamilton, R.G.; Craig, T.J. Venom immunotherapy reduces large local reactions to insect stings. J. Allergy Clin. Immunol. 2009, 123, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Michel, Y.; McIntyre, M.; Ginglinger, H.; Ollert, M.; Cifuentes, L.; Blank, S.; Spillner, E. The Putative Serine Protease Inhibitor Api m 6 From Apis mellifera Venom: Recombinant and Structural Evaluation. J. Investig. Allerg. Clin. 2012, 22, 476–484. [Google Scholar]
- King, T.P.; Spangfort, M.D. Structure and biology of stinging insect venom allergens. Int. Arch. Allergy Imm. 2000, 123, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Seismann, H.; McIntyre, M.; Ollert, M.; Wolf, S.; Bantleon, F.I.; Spillner, E. Vitellogenins Are New High Molecular Weight Components and Allergens (Api m 12 and Ves v 6) of Apis mellifera and Vespula vulgaris Venom. PLoS ONE 2013, 8, e62009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, S.; Seismann, H.; Bockisch, B.; Braren, I.; Cifuentes, L.; McIntyre, M.; Ruhl, D.; Ring, J.; Bredehorst, R.; Ollert, M.W.; et al. Identification, Recombinant Expression, and Characterization of the 100 kDa High Molecular Weight Hymenoptera Venom Allergens Api m 5 and Ves v 3. J. Immunol. 2010, 184, 5403–5413. [Google Scholar] [CrossRef] [Green Version]
- Peiren, N.; de Graaf, D.C.; Brunain, M.; Bridts, C.H.; Ebo, D.G.; Stevens, W.J.; Jacobs, F.J. Molecular cloning and expression of icarapin, a novel IgE-binding bee venom protein. FEBS Lett. 2006, 580, 4895–4899. [Google Scholar] [CrossRef] [Green Version]
- Blank, S.; Seismann, H.; Michel, Y.; McIntyre, M.; Cifuentes, L.; Braren, I.; Grunwald, T.; Darsow, U.; Ring, J.; Bredehorst, R.; et al. Api m 10, a genuine A. mellifera venom allergen, is clinically relevant but underrepresented in therapeutic extracts. Allergy 2011, 66, 1322–1329. [Google Scholar] [CrossRef]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta 2012, 1824, 164–176. [Google Scholar] [CrossRef]
- Orsolic, N. Bee venom in cancer therapy. Cancer Metastasis Rev. 2012, 31, 173–194. [Google Scholar] [CrossRef]
- Bernheimer, A.W.; Linder, R.; Weinstein, S.A.; Kim, K.S. Isolation and characterization of a phospholipase B from venom of Collett’s snake, Pseudechis colletti. Toxicon 1987, 25, 547–554. [Google Scholar] [CrossRef]
- Matysiak, J.; Hajduk, J.; Pietrzak, L.; Schmelzer, C.E.H.; Kokot, Z.J. Shotgun proteome analysis of honeybee venom using targeted enrichment strategies. Toxicon 2014, 90, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Bown, D.P.; Wilkinson, H.S.; Gatehouse, J.A. Midgut carboxypeptidase from Helicoverpa armigera (Lepidoptera: Noctuidae) larvae: Enzyme characterisation, cDNA cloning and expression. Insect Biochem. Mol. Biol. 1998, 28, 739–749. [Google Scholar] [CrossRef]
- Cho, W.L.; Deitsch, K.W.; Raikhel, A.S. An Extraovarian Protein Accumulated in Mosquito Oocytes Is a Carboxypeptidase Activated in Embryos. Proc. Natl. Acad. Sci. USA 1991, 88, 10821–10824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.Y.; Machleder, E.M.; Chenchik, A.; Li, R.; Siebert, P.D. Reverse transcriptase template switching: A SMART (TM) approach for full-length cDNA library construction. Biotechniques 2001, 30, 892–897. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Muller, U.; Helbling, A.; Berchtold, E. Immunotherapy with Honeybee Venom and Yellow Jacket Venom IS Different Regarding Efficacy and Safety. J. Allergy Clin. Immunol. 1992, 89, 529–535. [Google Scholar] [CrossRef]
- Yoon, K.A.; Kim, K.; Nguyen, P.; Seo, J.B.; Park, Y.H.; Kim, K.G.; Seo, H.Y.; Koh, Y.H.; Lee, S.H. Comparative functional venomics of social hornets Vespa crabro and Vespa analis. J. Asia-Pac. Entomol. 2015, 18, 815–823. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Kall, L.; Krogh, A.; Sonnhammer, E.L.L. A combined transmembrane topology and signal peptide prediction method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Stoeckert, C.J.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Mistry, J.; Schuster-Bockler, B.; Griffiths-Jones, S.; Hollich, V.; Lassmann, T.; Moxon, S.; Marshall, M.; Khanna, A.; Durbin, R.; et al. Pfam: Clans, web tools and services. Nucleic Acids Res. 2006, 34, D247–D251. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.K.; Zhang, S.; Luo, J.Y.; Wang, C.Y.; Lu, L.M.; Zhang, L.J.; Zhu, X.Z.; Wang, L.; Lu, H.; Cui, J.J. Comprehensive evaluation of candidate reference genes for gene expression studies in Lysiphlebia japonica (Hymenoptera: Aphidiidae) using RT-qPCR. Gene 2017, 637, 211–218. [Google Scholar] [CrossRef]
- Takezaki, N.; Nei, M.; Tamura, K. POPTREE2: Software for Constructing Population Trees from Allele Frequency Data and Computing Other Population Statistics with Windows Interface. Mol. Biol. Evol. 2010, 27, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Moore, A.M.; Ahmadi, S.; Patel, S.; Gibson, M.K.; Wang, B.; Ndao, I.M.; Deych, E.; Shannon, W.; Tarr, P.I.; Warner, B.B.; et al. Gut resistome development in healthy twin pairs in the first year of life (vol 3, 27, 2015). Microbiome 2015, 3. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Species | Total Number of Reads | Total Base Pairs | Trimmed Reads | Total Number of Assembled Transcripts | Total Number of Annotated Transcripts |
|---|---|---|---|---|---|
| E. decorates | 58,988,410 | 5,957,829,410 | 55,681,360 | 18,062 | 16,357 |
| Sphecidae sp. | 63,954,192 | 6,459,373,392 | 60,641,236 | 31,134 | 27,423 |
| A. flavomarginatum | 71,380,878 | 7,209,468,678 | 67,788,990 | 21,512 | 19,033 |
| S. deforme | 57,404,468 | 5,797,851,268 | 52,911,920 | 24,483 | 20,097 |
| V. crabro | 62,246,264 | 5,756,774,997 | 56,318,830 | 11,097 | 10,321 |
| V. analis | 61,648,078 | 5,686,981,519 | 55,419,968 | 12,531 | 11,284 |
| V. dybowskii | 113,343,134 | 11,447,656,534 | 103,185,332 | 21,227 | 19,627 |
| V. simillima | 114,592,946 | 11,573,887,546 | 89,279,802 | 29,150 | 27,065 |
| P. varia | 82,025,386 | 8,284,563,986 | 75,702,936 | 20,378 | 18,539 |
| P. snelleni | 80,798,216 | 8,160,649,816 | 75,374,540 | 27,742 | 25,879 |
| P. rothneyi | 81,060,754 | 8,187,136,154 | 76,096,056 | 25,436 | 23,475 |
| B. ardens | 76,622,404 | 7,738,862,804 | 71,427,168 | 31,366 | 26,158 |
| B. consobrinus | 109,962,550 | 11,106,217,550 | 72,432,174 | 21,625 | 20,050 |
| B. ussurensis | 106,973,794 | 10,804,353,194 | 99,138,794 | 26,425 | 22,619 |
| Description | TPM Value of Top 100 Highly Expressed Genes a | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V. an | V. cr | V. dy | V. si | P. va | P. ro | P. sn | E. de | Sph | A. fl | S. de | B. ar | B. co | B. us | |
| Defensin 1 | - | - | 1652 | - | - | - | - | - | - | 1163 | - | - | ||
| Endocuticle structural glycoprotein | 6209 | - | 1132 | 4404 | 3779 | - | - | 4847 | - | - | - | - | - | |
| Hyaluronidase | 10,229 | 4566 | 1409 | - | 1275 | - | 8511 | - | - | - | - | - | - | |
| Icarapin | 1495 | 1167 | 725 | - | - | - | - | 2857 | 2081 | - | 2039 | 4623 | 6943 | |
| Neprilysin | - | - | - | - | - | 4174 | - | - | 1895 | - | - | - | ||
| Phospholipase A1 | 49,149 | 106,395 | 29,423 | 6440 | 1246 | - | - | - | - | - | - | - | - | |
| Phospholipase A2 | 4722 | 1547 | 952 | - | - | - | - | - | - | - | - | - | 1157 | |
| Serine protease | 1301 | - | 1737 | 947 | - | 22,355 | - | - | - | - | - | - | ||
| Venom allergen 5 | 150,567 | 65,462 | 32,241 | 13,212 | - | - | - | - | - | - | - | - | - | |
| Uncharacterized protein 1 | 2677 | - | 29,700 | 4756 | - | 874 | - | - | - | - | - | 6955 | - | |
| Uncharacterized protein 2 | 17,769 | 1199 | - | 2611 | - | - | - | - | 865 | - | - | - | - | |
| Uncharacterized protein 3 | 2125 | - | - | - | - | - | - | - | - | - | 1805 | 1537 | ||
| Uncharacterized protein 4 | - | - | - | - | - | - | - | - | - | 1800 | - | 1741 | ||
| Uncharacterized protein 5 | 1584 | - | - | - | - | - | - | 1944 | - | - | - | - | ||
| Uncharacterized protein 6 | - | - | 2439 | - | - | - | - | 1322 | - | - | - | - | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, K.A.; Kim, K.; Kim, W.-J.; Bang, W.Y.; Ahn, N.-H.; Bae, C.-H.; Yeo, J.-H.; Lee, S.H. Characterization of Venom Components and Their Phylogenetic Properties in Some Aculeate Bumblebees and Wasps. Toxins 2020, 12, 47. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010047
Yoon KA, Kim K, Kim W-J, Bang WY, Ahn N-H, Bae C-H, Yeo J-H, Lee SH. Characterization of Venom Components and Their Phylogenetic Properties in Some Aculeate Bumblebees and Wasps. Toxins. 2020; 12(1):47. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010047
Chicago/Turabian StyleYoon, Kyungjae Andrew, Kyungmun Kim, Woo-Jin Kim, Woo Young Bang, Neung-Ho Ahn, Chang-Hwan Bae, Joo-Hong Yeo, and Si Hyeock Lee. 2020. "Characterization of Venom Components and Their Phylogenetic Properties in Some Aculeate Bumblebees and Wasps" Toxins 12, no. 1: 47. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12010047