
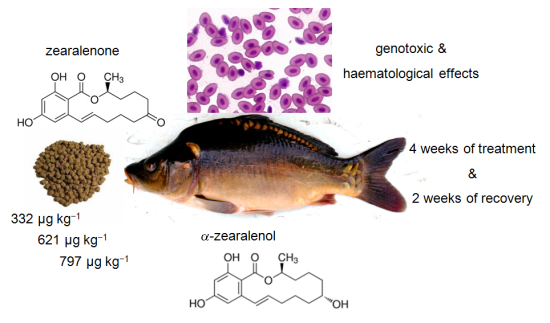
Effects of Dietary Exposure to Zearalenone (ZEN) on Carp (Cyprinus carpio L.)



Abstract
:
1. Introduction
2. Results and Discussion
2.1. Effects on Growth Performance

{kind=link}
| White Muscle Contamination | Basal feed | Low ZEN | Medium ZEN | High ZEN |
|---|---|---|---|---|
| After ZEN-treatment | ||||
| ZEN [ng g−1 dry weight] | n.d. | 0.13 ± 0.03 | 0.22 ± 0.18 | 0.15 ± 0.07 |
| α-ZEL [ng g−1 dry weight] | n.d. | 0.11 ± 0.03 | 0.16 ± 0.11 | 0.05 ± 0.07 |
| After Recovery | ||||
| ZEN [ng g−1 dry weight] | 0.04 ± 0.06 | 0.03 ± 0.03 | 0.03 ± 0.02 | 0.03 ± 0.03 |
| α-ZEL [ng g−1 dry weight] | n.d. | n.d. | n.d. | n.d. |
| Sampling Parameters | Basal feed | Low ZEN | Medium ZEN | High ZEN |
|---|---|---|---|---|
| Initial weight [g] | 30.0 ± 3.69 | 29.5 ± 3.45 | 28.8 ± 2.97 | 29.3 ± 3.55 |
| Final weight [g] | 44.0 ± 7.42 | 52.1 ± 7.21 | 41.8 ± 4.02 | 47.3 ± 5.88 |
| Weight gain [g fish−1 week−1] | 3.7 ± 0.68 | 5.7 ± 1.10 | 3.2 ± 0.35 | 4.7 ± 0.74 |
| Specific growth rate, SGR [% week−1] | 4.1 ± 0.06 | 4.2 ± 0.07 | 4.0 ± 0.04 | 4.2 ± 0.07 |
| FCR | 1.6 ± 0.41 | 1.0 ± 0.07 | 1.8 ± 0.28 | 1.2 ± 0.20 |
| Final total length [cm] | 15.1 ± 0.72 | 14.0 ± 0.75 | 13.7 ± 0.43 | 14.4 ± 0.59 |
| Final condition factor | 0.02 ± 0.000 | 0.02 ± 0.000 | 0.02 ± 0.001 | 0.02 ± 0.001 |
| Splenosomatic index | 0.2 ± 0.01 | 0.1 ± 0.02 * | 0.1 ± 0.03 | 0.4 ± 0.15 |
| Gonadosomatic index | 2.0 ± 0.60 | 1.1 ± 0.66 | 1.7 ± 0.52 | 1.4 ± 0.73 |
| Ratio males/females | 5/1 | 3/3 | 4/2 | 3/3 |
| Sampling Parameters | Basal feed | Low ZEN | Medium ZEN | High ZEN |
|---|---|---|---|---|
| Initial weight [g] | 26.5 ± 4.10 | 25.3 ± 2.87 | 23.0 ± 0.60 | 22.0 ± 3.48 |
| Final weight [g] | 52.3 ± 6.06 | 46.6 ± 2.51 | 45.1 ± 4.08 | 42.9 ± 5.02 |
| Weight gain [g fish−1 week−1] | 4.3 ± 0.76 | 3.5 ± 0.36 | 3.7 ± 0.59 | 3.5 ± 0.35 |
| Specific growth rate, SGR [% week−1] | 3.5 ± 0.10 | 3.5 ± 0.07 | 3.5 ± 0.07 | 3.5 ± 0.10 |
| FCR | 1.4 ± 0.25 | 1.6 ± 0.06 | 1.6 ± 0.42 | 1.7 ± 0.24 |
| Final total length [cm] | 14.7 ± 0.51 | 14.5 ± 0.27 | 14.2 ± 0.41 | 13.5 ± 0.58 |
| Final condition factor | 0.02 ± 0.000 | 0.02 ± 0.000 | 0.02 ± 0.000 | 0.02 ± 0.001 |
| Splenosomatic index | 0.1 ± 0.01 | 0.1 ± 0.03 | 0.2 ± 0.03 | 0.2 ± 0.06 |
| Gonadosomatic index | 2.0 ± 1.02 | 2.2 ± 0.84 | 1.0 ± 0.33 | 2.7 ± 1.04 |
| Ratio males/females | 3/3 | 3/3 | 2/4 | 3/3 |
| Blood cell counts | Basal feed | Low ZEN | Medium ZEN | High ZEN |
|---|---|---|---|---|
| ZEN-treated | ||||
| leukocytes [% total blood cells] | 3.4 ± 0.20 | 3.7 ± 0.31 | 3.4 ± 0.35 | 3.1 ± 0.32 |
| lymphocytes [% all white blood cells] | 60.9 ± 2.06 | 58.1 ± 3.83 | 63.3 ± 4.87 | 60.9 ± 2.42 |
| thrombocytes [% all white blood cells] | 31.1 ± 2.92 | 32.1 ± 2.81 | 27.6 ± 4.37 | 30.0 ± 2.89 |
| monocytes [% all white blood cells] | 4.8 ± 0.49 | 4.2 ± 0.61 | 2.9 ± 0.40 * | 3.1 ± 0.30 * |
| granulocytes [% all white blood cells] | 3.2 ± 0.78 | 5.7 ± 1.69 | 6.2 ± 1.34 * | 6.1 ± 0.85 * |
| MN in erythrocytes [per 1000 cells] | 12.9 ± 5.13 | 43.5 ± 9.66 * | 29.9 ± 2.92 * | 36.54 ± 7.5 * |
| Recovery | ||||
| leukocytes [% total blood cells] | 3.6 ± 0.54 | 2.8 ± 0.32 | 4.9 ± 0.50 | 2.7 ± 0.36 |
| lymphocytes [% all white blood cells] | 59.6 ± 3.30 | 59.3 ± 1.56 | 65.6 ± 3.80 | 64.0 ± 4.92 |
| thrombocytes [% all white blood cells] | 34.3 ± 2.77 | 30.2 ± 2.41 | 29.8 ± 3.39 | 29.0 ± 4.95 |
| monocytes [% all white blood cells] | 2.6 ± 0.49 | 3.8 ± 0.97 | 2.8 ± 0.44 | 2.5 ± 0.40 |
| granulocytes [% all white blood cells] | 3.4 ± 0.50 | 6.7 ± 1.43 | 1.8 ± 0.34 | 4.7 ± 1.01 |
2.2. Hematology
| Haematological Parameters | Basal feed | Low ZEN | Medium ZEN | High ZEN |
|---|---|---|---|---|
| ZEN-treated | ||||
| Haematocrit [%] | 28.4 ± 1.51 | 32.6 ± 1.85 | 28.7 ± 0.75 | 32.9 ± 2.33 |
| Haemoglobin [mg dL−1] | 7.9 ± 0.46 | 9.1 ± 0.68 | 8.7 ± 0.50 | 8.9 ± 0.54 |
| Vitellogenin [ng mL−1] | 104 ± 3.5 | 112 ± 6.8 | 103 ± 1.9 | 108 ± 3.1 |
| Recovery | ||||
| Haematocrit [%] | 29.1 ± 0.97 | 29.3 ± 0.82 | 26.9 ± 0.79 | 30.5 ± 1.25 |
| Haemoglobin [mg dL−1] | 8.5 ± 0.24 | 8.4 ± 0.24 | 7.6 ± 0.28 * | 8.4 ± 0.35 |
| Vitellogenin [ng mL−1] | 102 ± 1.4 | 103 ± 3.1 | 110 ± 4.4 | 103 ± 1.7 |
3. Experimental Section
3.1. Chemicals
3.2. Preparation of Feeds and Husbandry
3.3. Analysis of ZEN in Experimental Diets and ZEN and Its Metabolites in Muscle Samples
3.4. Experimental Feeding Design
3.5. Determination of Haematological Parameters
3.6. Determination of Differential Blood Cell Counts and Micronuclei Counts
3.7. Analysis of Plasma Vitellogenin
3.8. Statistics
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Måge, A.; Julshamn, K.; Lunestad, B.T. Overvåkningsprogram for förvarer til fisk og andre akvatiske dyr e Årsrapport 2008 og 2009; NIFES: Bergen, Norway, 2009. [Google Scholar]
- Santos, G.A.; Rodrigues, I.; Naehrer, K.; Encarnacao, P. Mycotoxins in aquaculture: occurrence in feed components and impact on animal performance. Aquac. Eur. 2010, 35, 6–10. [Google Scholar]
- Pietsch, C.; Kersten, S.; Burkhardt-Holm, P.; Valenta, H.; Dänicke, S. Occurrence of deoxynivalenol and zearalenone in commercial fish feed—An initial study. Toxins 2013, 5, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Coppock, R.W.; Mostrom, M.S.; Sparling, C.G.; Jacobsen, B.; Ross, S.C. Apparent zearalenone intoxication in a dairy herd from feeding spoiled acid treated corn. Vet. Hum. Toxicol. 1990, 32, 246–248. [Google Scholar] [PubMed]
- Veldman, A.; Borggreve, G.J.; Mulders, E.J.; Van de Lagemaat, D. Occurence of the mycotoxins ochratoxin A, zearalenone and deoxynivalenol in feed components. Food Add. Contam. 1992, 9, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Charmley, L.L.; Rosenberg, A.; Trenholm, H.L. Factors responsible for economic losses due to Fusarium mycotoxin contamination of grains, foods and feedstuffs. In Mycotoxins in Grain-Compounds Other than Aflatoxin; Miller, J.D., Trenholm, H.L., Eds.; Eagan Press: St. Paul, MN, USA, 1994; pp. 471–486. [Google Scholar]
- Whitlow, L.W.; Hagler, W.M., Jr. Mycotoxins in feeds. Feedstuffs 2005, 77, 69–79. [Google Scholar]
- Mankeviciene, A.; Butkute, B.; Dabkevicius, Z.; Suproniene, S. Fusarium mycotoxins in lithuanian cereals from the 2004e05 harvests. Ann. Agric. Environ. Med. 2007, 14, 103–107. [Google Scholar] [PubMed]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of mycotoxins in feedstuffs of dairy cows and estimation of total dietary intakes. J. Dairy Sci. 2008, 91, 4261–4271. [Google Scholar] [CrossRef] [PubMed]
- Maaroufi, K.; Chekir, L.; Creppy, E.E.; Ellouz, F.; Bacha, H. Zearalenone induces modifications of haematological and biochemical parameters in rats. Toxicon 1996, 34, 535–540. [Google Scholar] [CrossRef]
- Obremski, K.; Zielonka, L.; Zaluska, G.; Zwierzchowski, W.; Pirus, K.; Gajecki, M. The influence of low doses of zearalenone on liver enzyme activities in gilts. In Proceedings of the X Conference “Microscopic Fungi—Plant Pathogens and their Metabolites”, Pznan, Poland, 29 April 1999; p. 66.
- Conkova, E.; Laciakova, A.; Pastorova, B.; Seidel, H.; Kovac, G. The effect of zearalenone on some enzymatic parameters in rabbits. Toxicol. Lett. 2001, 121, 145–149. [Google Scholar] [CrossRef]
- Marin, L.; Murtha, J.; Dong, W.; Pestka, J.J. Effects of mycotoxins on cytokine production and proliferation in EL-4 thymoma cells. J. Toxicol. Environ. Health 1996, 48, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Berek, L.; Petri, I.B.; Mesterhazy, A.; Teren, J.; Molnar, J. Effects of mycotoxins on human immune functions in vitro. Toxicol. In Vitro 2001, 15, 25–30. [Google Scholar] [CrossRef]
- Ouanes, Z.; Abid, S.; Ayed, I.; Anane, R.; Mobio, T.; Creppy, E.; Bacha, H. Induction of micronuclei by zearalenone in Vero monkey kidney cells and in bone marrow cells of mice: protective effect of vitamine E. Mutat. Res. 2003, 538, 63–70. [Google Scholar] [CrossRef]
- Abid-Essefi, S.; Ouanes, Z.; Hassen, W.; Baudrimont, I.; Creppy, E.E.; Bacha, H. Cytotoxicity, inhibition of DNA and protein syntheses and oxidative damage in cultured cells exposed to zearalenone. Toxicol. In Vitro 2004, 18, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Lioi, M.B.; Santoro, A.; Barbieri, R.; Salzano, S.; Ursini, M.V. Ochratoxin and zearalenone: A comparative study on genotoxic effects and cell death induced in bovine lymphocytes. Mutat. Res. 2004, 557, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Döll, S.; Valenta, H.; Baardsen, G.; Möller, P.; Koppe, W.; Stubhaug, I.; Dänicke, S. Effects of increasing concentrations of deoxynivalenol, zearalenone and ochratoxin A in diets for Atlantic salmon (Salmo salar) on performance, health and toxin residues. In Proceedings of the Abstracts of the 33rd Mycotoxin Workshop, Freising, Germany, 30 May–1 June 2011.
- Pietsch, C. Impact of natural toxins on carp. In Biology and Ecology of Carp; Pietsch, C., Hirsch, P.E., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2015; pp. 334–346. [Google Scholar]
- Woźny, M.; Brzuzan, P.; Gusiatin, M.; Jakimiuk, E.; Dobosz, S.; Kuźmiński, H. Influence of zearalenone on selected biochemical parameters in juvenile rainbow trout (Oncorynchus mykiss). Polish J. Vet. Sci. 2012, 15, 221–225. [Google Scholar]
- Chattopadhyay, P.; Upadhyay, A.; Agnihotri, A.; Karmakar, S.; Ghoyary, D.; Veer, V. Comparative hematoxicity of Fusirium mycotoxin in experimental Sprague-Dawley rats. Toxicol. Int. 2013, 20, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Vlata, Z.; Porichis, F.; Tzanakakis, G.; Tsatsakis, A.; Krambovitis, E. A study of zearalenone cytotoxicity on human peripheral blood mononuclear cells. Toxicol Lett. 2006, 165, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Jilani, K.; Lang, F. Ca2+-dependent suicidal erythrocyte death following zearalenone exposure. Arch. Toxicol. 2013, 87, 1821–1828. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Chang, W.P.; Kirsch, V.M.; Holland, N.; Bonassi, S.; Zeiger, E. HUMN project: detailed description of the scoring criteria for the cytokinesis-block micronucleus assay using isolated human lymphocyte cultures. Mutat. Res. 2003, 534, 65–75. [Google Scholar] [CrossRef]
- Heddle, J.A.; Cimino, M.C.; Hayashi, M.; Romagna, F.; Shelby, M.D.; Tucker, J.D.; Vanparys, P.; MacGregor, J.T. Micronuclei as an index of cytogenetic damage: past, present, and future. Environ. Mol. Mutagen. 1991, 18, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Ayed-Boussema, I.; Ouanes, Z.; Bacha, H.; Abid, S. Toxicities induced in cultured cells exposed to zearalenone: Apoptosis or mutagenesis? J. Biochem. Mol. Toxicol. 2007, 21, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Noser, J.; Wettstein, F.E.; Burkhardt-Holm, P. Unravelling the mechanisms involved in zearalenone-mediated toxicity in permanent fish cell cultures. Toxicon 2014, 88, 44–61. [Google Scholar] [CrossRef] [PubMed]
- Bucheli, T.D.; Wettstein, F.E.; Hartmann, N.; Erbs, M.; Vogelsang, S.; Forrer, H.-R.; Schwarzenbach, R.P. Fusarium mycotoxins: Overlooked aquatic micropollutants. J. Agric. Food Chem. 2008, 56, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Hoerger, C.; Schenzel, J.; Strobel, B.; Bucheli, T. Analysis of selected phytotoxins and mycotoxins in environmental samples. Anal. Bioanal. Chem. 2009, 395, 1261–1289. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, N.; Erbs, M.; Forrer, H.-R.; Vogelsang, S.; Wettstein, F.E.; Schwarzenbach, R.P.; Bucheli, T.D. Occurrence of zearalenone on Fusarium graminearum infected wheat and maize field in crop organs, soil, and drainage water. Environ. Sci. Technol. 2008, 42, 5455–5460. [Google Scholar] [CrossRef] [PubMed]
- Bucheli, T.D.; Erbs, M.; Hartmann, N.; Vogelsang, S.; Wettstein, F.E.; Forrer, H.R. Estrogenic mycotoxins in the environment. Mitt. Lebensm. Hyg. 2005, 96, 386–403. [Google Scholar]
- Molina-Molina, J.-M.; Real, M.; Jimenez-Diaz, I.; Belhassen, H.; Hedhili, A.; Tornè, P.; Fernàndez, M.F.; Olea, N. Assessment of estrogenic and anti-androgenic activities of the mycotoxin zearalenone and its metabolites using in vitro receptor-specific bioassays. Food Chem. Toxicol. 2014, 74, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Benzoni, E.; Minervini, F.; Giannoccaro, A.; Fornelli, F.; Vigo, D.; Visconti, A. Influence of in vitro exposure to mycotoxin zearalenone and its derivatives on swine sperm quality. Reprod. Toxicol. 2008, 25, 461–467. [Google Scholar] [CrossRef] [PubMed]
- De Andrés, F.; Zougagh, M.; Castañeda, G.; Ríos, A. Determination of zearalenone and its metabolites in urine samples by liquid chromatography with electrochemical detection using a carbon nanotube-modified electrode. J. Chromatogr. A 2008, 1212, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Filannino, A.; Stout, T.; Gadella, B.; Sostaric, E.; Pizzi, F.; Colenbrander, B.; Dell’Aquila, M.; Minervini, F. Dose-response effects of estrogenic mycotoxins (zearalenone, alpha- and beta-zearalenol) on motility, hyperactivation and the acrosome reaction of stallion sperm. Reprod. Biol. Endocrinol. 2011, 9, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Uhlig, S.; Miles, C.O.; Verhaegen, S.; Elliott, C.T.; Eriksen, G.S.; Sorlie, M.; Ropstad, E.; Connolly, L. Biotransformation of zearalenone and zearalenols to their major glucuronide metabolites reduces estrogenic activity. Toxicol. In Vitro 2015, 29, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Johns, S.M.; Denslow, N.D.; Kane, M.D.; Watanabe, K.H.; Orlando, E.F.; Sepulveda, M.S. Effects of estrogens and antiestrogens on gene expression of fathead minnow (Pimephales promelas) early life stages. Environ. Toxicol. 2009, 26, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, P.; Thorpe, K.L.; Bucheli, T.D.; Wettstein, F.E.; Burkhardt-Holm, P. Short-term exposure to the environmentally relevant estrogenic mycotoxin zearalenone impairs reproduction in fish. Sci. Tot. Environ. 2010, 409, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Bakos, K.; Kovács, R.; Staszny, Á.; Sipos, D.K.; Urbányi, B.; Müller, F.; Csenki, Z.; Kovács, B. Developmental toxicity and estrogenic potency of zearalenone in zebrafish (Danio rerio). Aquat. Toxicol. 2013, 136, 13–21. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- European Commission. Commission Recommendation (2006/576/EC) of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- Lagana, A.; Faberi, A.; Fago, G.; Marino, A.; Pastorini, E.; Samperi, R. Application of an innovative matrix solid-phase dispersionesolidphase extractioneliquid chromatographyetandem mass spectrometry analytical methodology to the study of the metabolism of the estrogenic mycotoxin zearalenone in rainbow trout liver and muscular tissue. Int. J. Environ. Anal. Chem. 2004, 84, 1009–1016. [Google Scholar]
- Olsen, M.; Pettersson, H.; Kiessling, K.H. Reduction of zearalenone to zearalenol in female rat liver by 3 alpha-hydroxysteroid dehydrogenase. Acta Pharmacol. Toxicol. 1981, 48, 157–161. [Google Scholar] [CrossRef]
- Schaut, A.; De Saeger, S.; Sergent, T.; Schneider, Y.-J.; Larondelle, Y.; Pussemier, L.; Van Peteghem, C. Study of the gastrointestinal biotransformation of zearalenone in a Caco-2 cell culture system with liquid chromatographic methods. J. Appl. Toxicol. 2008, 28, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Videmann, B.; Mazallon, M.; Tep, J.; Lecoeur, S. Metabolism and transfer of the mycotoxin zearalenone in human intestinal Caco-2 cells. Food Chem. Toxicol. 2008, 46, 3279–3286. [Google Scholar] [CrossRef] [PubMed]
- Becci, P.J.; Voss, K.A.; Hess, F.G.; Gallo, M.A.; Parent, R.A.; Stevens, K.R.; Taylor, J.M. Long-term carcinogenicity and toxicity study of zearalenone in the rat. J. Appl. Toxicol. 1982, 2, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, K.H. The effect of zearalenone on growth rate, organ weight and muscle fibre composition in growing rats. Acta Pharmacol. Toxicol. (Copenh) 1982, 51, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Kanora, A.; Maes, D. The role of mycotoxins in pig reproduction: a review. Vet. Med. 2009, 54, 565–576. [Google Scholar]
- Allen, N.K.; Mirocha, C.J.; Weaver, G.; Aakhus-Allen, S.; Bates, F. Effects of dietary zearalenone on finishing broiler chickens and young turkey poults. Poultry Sci. 1981, 60, 124–131. [Google Scholar] [CrossRef]
- Houston, A.H.; Roberts, W.C.; Kennington, J.A. Hematological response in fish: pronephric and splenic involvements in the goldfish, Carassius auratus L. Fish Physiol. Biochem. 1996, 15, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Houston, A.H.; Murad, A. Erythrodynamics in goldfish, Carassius auratus L.: Temperature effects. Physiol. Zool. 1992, 65, 55–76. [Google Scholar]
- Fange, R. Physiology of hemopoiesis. In Fish physiology: recent advances; Nilsson, S., Holgren, S., Eds.; Croom Helm: London, UK, 1986; pp. 1–23. [Google Scholar]
- Lang, E.; Lang, F. Triggers, inhibitors, mechanisms, and significance of eryptosis: The suicidal erythrocyte death. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Al-Rudainy, A.J.; Kadhim, M.H. Hematological and neurotoxic effects of endosulfan pesticide on common carp Cyprinus carpio. The Iraqi J. Vet. Med. 2012, 36, 58–67. [Google Scholar]
- Tun, N.; Houston, A.H. Temperature, oxygen, photoperiod and the hemoglobin system of the rainbow trout, Salmo gairdneri. Can. J. Zool. 1986, 64, 1883–1888. [Google Scholar] [CrossRef]
- Warth, B.; Sulyok, M.; Berthiller, F.; Schuhmacher, R.; Krska, R. New insights into the human metabolism of the Fusarium mycotoxins deoxynivalenol and zearalenone. Toxicol. Lett. 2013, 220, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Kostro, K.; Gajecka, M.; Lisiecka, U.; Majer-Dziedzic, B.; Obremski, K.; Zielonka, L.; Gajecki, M. Subpopulation of lymphocytes CD4+ and CD8+ in peripheral blood of sheep with zearalenone mycotoxicosis. Bull. Vet. Inst. Pulawy 2011, 55, 241–246. [Google Scholar]
- Pietsch, C.; Junge, R.; Burkhardt-Holm, P. Immunomodulation by zearalenone (ZEN) in carp (Cyprinus carpio L.). BioMed Research Int. 2015. submitted. [Google Scholar]
- Abbès, S.; Ben Salah-Abbès, J.; Sharafi, H.; Noghabi, K.A.; Oueslati, R. Interaction of Lactobacillus plantarum MON03 with Tunisian Montmorillonite clay and ability of the composite to immobilize Zearalenone in vitro and counteract immunotoxicity in vivo. Immunopharmacol. Immunotoxicol. 2012, 34, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gao, R.; Meng, Q.; Zhang, Y.; Bi, C.; Shan, A. Toxic effects of maternal zearalenone exposure on intestinal oxidative stress, barrier function, immunological and morphological changes in rats. PLoS ONE 2014, 9, e106412. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Tudor, D.S. Effects of zearalenone and its derivatives on the innate immune response of swine. Toxicon 2010, 56, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Manda, G.; Motiu, M.; Neagoe, I.; Dragomir, C.; Stancu, M.; Calin, L. Effects of zearalenone and its derivatives on porcine immune response. Toxicol. In Vitro 2011, 25, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Deng, J.L.; Xu, S.W.; Peng, X.; Zuo, Z.C.; Cui, H.M.; Wang, Y.; Ren, Z.H. Effects of zearalenone on IL-2, IL-6, and IFN-gamma mRNA levels in the splenic lymphocytes of chickens. Sci. World J. 2012. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.-K.; Cho, J.-H.; Jeong, S.-H.; Shin, H.-S.; Son, S.-W.; Yeo, Y.-K.; Kang, H.-G. Zearalenone affects immune-related parameters in lymphoid organs and serum of rats vaccinated with porcine parvovirus vaccine. Toxicol. Res. 2012, 28, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Pistol, G.C.; Braicu, C.; Motiu, M.; Gras, M.A.; Marin, D.E.; Stancu, M.; Calin, L.; Israel-Roming, F.; Berindan-Neagoe, I.; Taranu, I. Zearalenone mycotoxin affects immune mediators, MAPK signalling molecules, nuclear receptors and genome-wide gene expression in pig spleen. PLoS ONE 2015, 10, e0127503. [Google Scholar] [CrossRef] [PubMed]
- Obremski, K. Changes in Th1 and Th2 cytokine concentrations in ileal Peyer’s patches in gilts exposed to zearalenone. Polish J. Vet. Sci. 2014, 17, 53–59. [Google Scholar] [CrossRef]
- Wu, Q.; Li, M.; Gao, X.; Giesy, J.P.; Cui, Y.; Yang, L.; Kong, Z. Genotoxicity of crude extracts of cyanobacteria from Taihu Lake on carp (Cyprinus carpio). Ecotoxicol. 2011, 20, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Duca, R.-C.; Bravin, F.; Delaforge, M. Study of zearalenone contaminated feedstuffs on the detoxification enzymes. Archiva Zootechnica 2006, 9, 19–27. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Fish; National Academy Press: Washington, DC, USA, 1993; p. 114. [Google Scholar]
- VDLUFA-Methodenbuch III. 6. Ergänzung 2006, Zearalenon 16.9.2; VDLUFA-Verlag: Darmstadt, Germany, 2006.
- Brezina, U.; Valenta, H.; Rempe, I.; Kersten, S.; Humpf, H.-U.; Dänicke, S. Development of a liquid chromatography tandem mass spectrometry method for the simultaneous determination of zearalenone, deoxynivalenol and their metabolites in pig serum. Mycotox. Res. 2014, 30, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Valenta, H.; Dänicke, S.; Döll, S. Analysis of deoxynivalenol and de-epoxy-deoxynivalenol in animal tissues by liquid chromatography after clean-up with an immunoaffinity column. Mycotox. Res. 2003, 19, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Drabkin, D.L.; Austin, J.H. Spectrophotometric studies. II. Preparations from washed blood cells; nitric oxide hemoglobin and sulfhemoglobin. J. Biol. Chem. 1935, 112, 51–65. [Google Scholar]
- Al-Sabti, K.; Metcalfe, C.D. Fish micronuclei for assessing genotoxicity in water. Mutat. Res. 1995, 343, 121–135. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietsch, C.; Kersten, S.; Valenta, H.; Dänicke, S.; Schulz, C.; Burkhardt-Holm, P.; Junge, R. Effects of Dietary Exposure to Zearalenone (ZEN) on Carp (Cyprinus carpio L.). Toxins 2015, 7, 3465-3480. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7093465
Pietsch C, Kersten S, Valenta H, Dänicke S, Schulz C, Burkhardt-Holm P, Junge R. Effects of Dietary Exposure to Zearalenone (ZEN) on Carp (Cyprinus carpio L.). Toxins. 2015; 7(9):3465-3480. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7093465
Chicago/Turabian StylePietsch, Constanze, Susanne Kersten, Hana Valenta, Sven Dänicke, Carsten Schulz, Patricia Burkhardt-Holm, and Ranka Junge. 2015. "Effects of Dietary Exposure to Zearalenone (ZEN) on Carp (Cyprinus carpio L.)" Toxins 7, no. 9: 3465-3480. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7093465