The Tug-of-War between Plants and Viruses: Great Progress and Many Remaining Questions



1
College of Agriculture, Northeast Agricultural University, Harbin 150030, China
2
Centro Nacional de Biotecnología (CNB-CSIC), Campus Universidad Autónoma de Madrid, 28049 Madrid, Spain
3
State Key Laboratory of Rice Biology, Institute of Biotechnology, Zhejiang University, Hangzhou 310058, China
4
State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Viruses 2019, 11(3), 203; https://0-doi-org.brum.beds.ac.uk/10.3390/v11030203
Submission received: 17 January 2019
/
Revised: 18 February 2019
/
Accepted: 23 February 2019
/
Published: 28 February 2019
(This article belongs to the Special Issue Plant Immunity to Virus Infections)
{kind=link}
Abstract
:Plants are persistently challenged by various phytopathogens. To protect themselves, plants have evolved multilayered surveillance against all pathogens. For intracellular parasitic viruses, plants have developed innate immunity, RNA silencing, translation repression, ubiquitination-mediated and autophagy-mediated protein degradation, and other dominant resistance gene-mediated defenses. Plant viruses have also acquired diverse strategies to suppress and even exploit host defense machinery to ensure their survival. A better understanding of the defense and counter-defense between plants and viruses will obviously benefit from the development of efficient and broad-spectrum virus resistance for sustainable agriculture. In this review, we summarize the cutting edge of knowledge concerning the defense and counter-defense between plants and viruses, and highlight the unexploited areas that are especially worth investigating in the near future.
1. Introduction
Plant viruses are obligate intracellular parasites that rely almost exclusively on the host cell to accomplish their infection cycle, e.g., genomic information expression, genome replication, and intercellular movement. During the infection, a comprehensive and competitive protein–protein and protein–nucleic acid interaction network is formed between the plant and virus. In general, the interaction network includes mechanisms by which the plant forms antiviral responses and the virus hijacks host factors for proliferation and to cope with plant antiviral defenses. The elaborate balance between the mechanisms used by the plant and the virus determines the outcome of the interaction, whether a virus is pathogenic or not. Understanding the plant–virus interaction network is one of the key goals of virology research. Impressive progress has been made in the knowledge of plant–virus interplay in the past decade, which not only quickly filled in the gap between plant innate immunity against virus and other pathogens, but also revealed unique plant antiviral machinery, such as NUCLEAR SHUTTLE PROTEIN (NSP) INTERACTING KINASE 1 (NIK1)-mediated translation repression and atypical dominant resistance by inhibiting viral protein function. In this review, we present the cutting edge of knowledge concerning the defense–counter-defense between plants and viruses, and outline the areas that may be particularly fruitful for study in the coming years. Due to space limitations, how plant viruses hijack the host’s metabolism for multiplication and dissemination is not included in the review, and readers are referred to the recent reviews [1,2]. Recessive resistance as specific antiviral resistance machinery due to the loss of key host factors for virus proliferation is also not discussed here; instead, the readers are referred to reviews [3,4,5].
2. Plant Antiviral Defenses
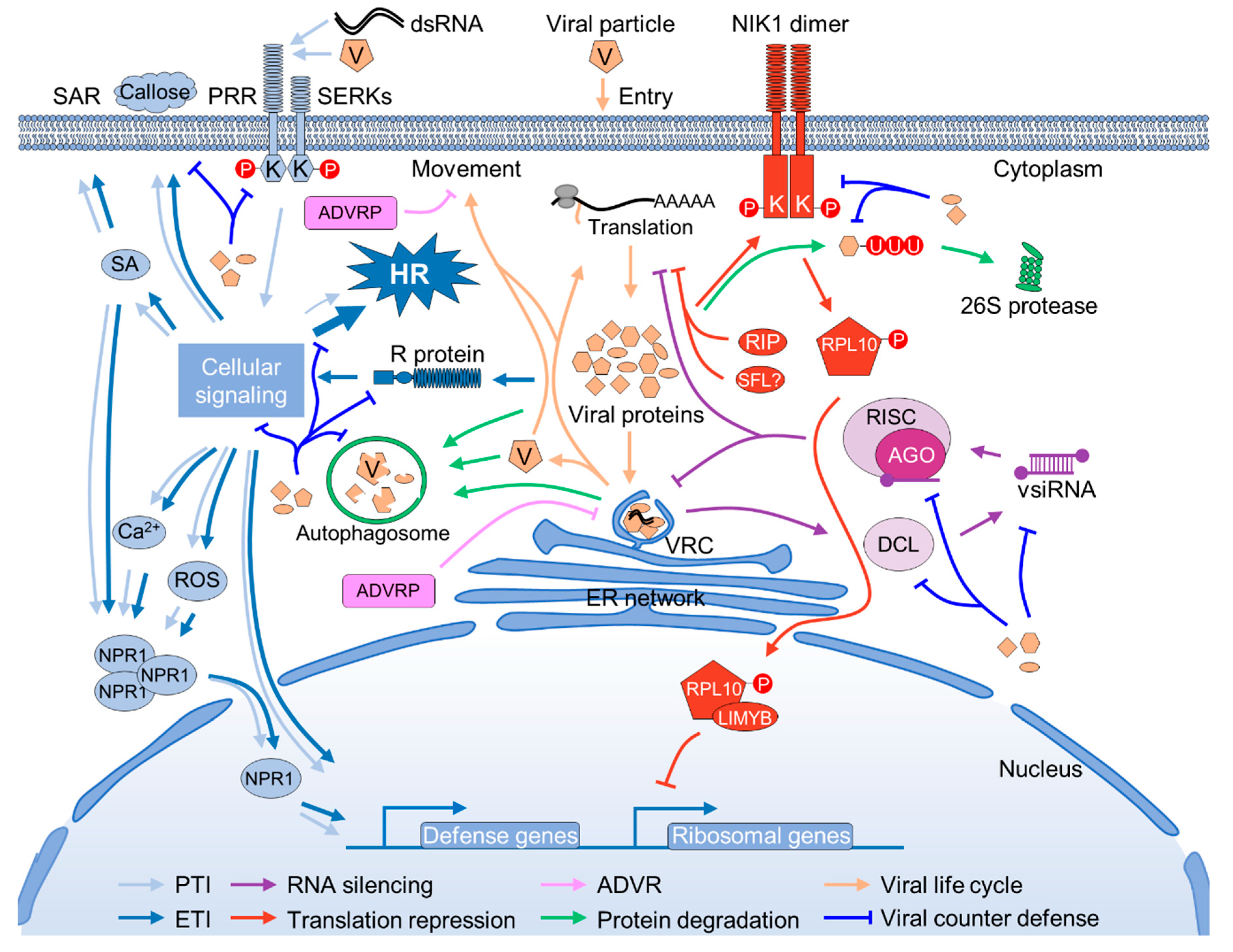
Plants are persistently challenged by varied pathogens, including viruses, bacteria, fungi, nematodes, and insects. Unlike the animal counterparts that possess specialized defense cells, plants rely on the capacity of every cell to perceive and defend against the “challengers”. During the millions of years of co-evolution with pathogens, plants have evolved multilayered surveillance mechanisms against pathogens that invade them [6,7]. Innate immunity, RNA silencing, translational repression, and ubiquitination-mediated and autophagy-mediated protein degradation are the major defense mechanisms against viruses in plants [8]. Exploration of the dominant resistance genes in the last decade also resulted in the identification of some antiviral proteins that restrict virus proliferation by directly interacting with and inhibiting viral protein functions (Figure 1).
2.1. Innate Antiviral Immunity
According to current plant immunity descriptions, there are two layers of plant immune responses against microbial pathogens. First, plant perception of the microorganism at the cell surface through the recognition of certain conserved pathogen-associated molecular patterns (PAMPs) by the extracellular surface pattern recognition receptor (PRR) initiates the so-called PAMP-triggered immunity (PTI) [9,10]. Plant PRRs fall into two major classes: the plasma membrane-localized receptor kinases (RKs) and the receptor-like proteins (RLPs) [11,12]. After the successful perception of PAMPs, PRRs immediately dimerize and associate with cofactors, such as SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE (SERKs) and SUPPRESSOR OF BIR1-1 (SOBIR1), and trigger a series of downstream intracellular signaling events including, but not limited to, oxidative burst, ion influx, the increased biosynthesis of defense hormones, and the activation of MITOGEN-ACTIVATED PROTEIN KINASES (MAPKs) that finally lead to resistance responses, e.g., the expression of pathogenesis-related (PR) genes, the synthesis and deposition of callose at the plasmodesmata (PD), and strengthening of the cell wall [13,14]. Occasionally, the activation of PTI also results in the hypersensitive response (HR), which is a specific type of programmed cell death (PCD) that occurs in the cells at the site of infection and causes discrete necrotic spots [15]. At present, a large number of plant PRRs and their respective PAMPs have been identified and characterized [12].
Despite the mass of knowledge on plant PTI against other phytopathogens, little is known about plant PTI against plant viruses, as they are historically viewed as non-PAMP coding pathogens [7]. Nevertheless, several lines of evidence suggest that PTI also plays a pivotal role in both incompatible and compatible plant–virus interaction. First, the Arabidopsis mutants in the PRR coreceptor kinases SERK3 (also known as BRASSINOSTEROID INSENSITIVE1 (BRI1)-ASSOCIATED RECEPTOR KINASE1, BAK1) and/or SERK4 (also called BAK1-LIKE, BKK1) exhibit increased susceptibility to different RNA viruses [16,17]. Similarly, knockdown of the expression of CaLecRK-S.5, which is a transmembrane RK possessing an L-type lectin domain at the N-terminus from pepper (Capsicum annuum L.), results in the enhanced susceptibility of pepper to several unrelated pathogens, including two plant viruses, tobacco mosaic virus (TMV, a tobamovirus), and pepper mild mottle virus (PMMoV, a tobamovirus), one bacterium (Xanthomonas campestris pv. vesicatoria), and one oomycete pathogen (Phytophthora capsici) [18]. Moreover, pre-treatment with β-Aminobutyric acid (BABA), which is a well-known immunity-priming chemical trigger [19], restores the disease resistance in CaLecRK-S.5-silenced plants [18]. Second, the exogenous application of double-strand RNA (dsRNA), which is a well-characterized PAMP in animal antiviral immunity [20,21], also triggers typical PTI responses in Arabidopsis that are dependent on the co-receptor SERK1, but independent of the RNA silencing pathway [22]. Third, the coat protein (CP) of TMV and potato virus X (PVX, a potexvirus) can trigger PTI-like responses in tobacco and Arabidopsis, respectively [23,24]. CP is the outermost component of all the non-enveloped viruses that should have contact with the plant cell surface during the invasion. Thus, it is possible that the PTI-like responses are triggered by unknown PAMPs derived from TMV and PVX CP proteins. Agreeing with the hypothesis, conserved local or overall three-dimensional folding was found in the CPs of viruses with similar particles, such as flexuous filamentous rods [25]. Nevertheless, viral PAMPs, the plant PRRs that perceive them, and cellular signaling components are still elusive at present.
Adapted pathogens can deliver specific proteins (effectors) into plant cells to compromise PTI defenses. To further counteract the effectors, plants have evolved an additional monitoring system that relies on particular intracellular receptors known as R proteins to directly or indirectly recognize the microbial effectors that are used to subvert PTI and trigger the so-called effector-triggered immunity (ETI) [26]. Over the past decade, a large number of R genes that mediate resistance against various plant viruses have been cloned, as they have apparent practical application value [27,28]. Functional R proteins typically contain an N-terminal Toll/Interleukin-1 receptor homology (TIR) domain or a coiled-coil (CC) domain, a central nucleotide-binding (NB) domain, and a C-terminal leucine-rich repeat (LRR) domain [6]. In addition, non-canonical domains have also been identified in some plant R proteins [29]. The NB-LRR domain plays a pivotal role in pathogen perception, although the non-canonical domain might also have a role in enhancing the recognition [30]. Similar to PTI, the successful perception of an effector by an R protein will trigger a series of downstream signaling events that lead to resistance responses. The direct consequence of defense in most R-mediated resistance responses is HR [31]. However, some R genes, such as the potato Rx1 gene, confer an extreme immunity that is not associated with HR [32]. Rx1 recognizes the PVX CP by the leucine-rich repeat domain, and it is then translocated to the nucleus by the RAN GTPase-ACTIVATING PROTEIN 2 RanGAP2 to activate the resistance against PVX, possibly via a Golden2-like transcription factor (NbGlk1) [33,34,35]. Thus, it was proposed that resistance and HR are separate responses in R-mediated resistance [36]. However, HR has an apparent function in restricting viral proliferation, as inhibiting or delaying the cell death via high-temperature treatment or the transgenic over-expression of HR suppressors, e.g., the baculovirus p35 protein and the endogenous plastid-localized protein DS9, results in the systemic infection of TMV on tobacco plants carrying the resistance N gene [37,38,39].
The components that are required for R-mediated resistance against virus infection largely overlap with those for other phytopathogens, suggesting the convergence of plant ETI [7]. For instance, SUPPRESSOR OF THE G2 ALLELE OF SKP1 (SGT1), REQUIRED FOR MLA12 RESISTANCE1 (RAR1), and HEAT SHOCK PROTEIN90 (HSP90) form the SGT1/RAR1/HSP90 complex that is required for both the N-mediated and Rx-mediated resistance against TMV and PVX, respectively, and immune responses against bacterial infections [40,41,42,43]. The complex formed by ENHANCED DISEASE SUSCEPTIBILITY1 (EDS1), PHYTOALEXIN DEFICIENT4 (PAD4), and SENESCENCE-ASSOCIATED GENE101 (SAG101) regulates HR TO TCV INFECTION (HRT)-mediated resistance against turnip crinkle virus (TCV), bacteria, fungi, and even cold tolerance [44,45,46,47,48].
Both PTI and ETI can result in the establishment of resistance in the distal, non-infected tissues, which is a phenomenon called systemic acquired resistance (SAR) [49]. Intriguingly, the resistance is not only limited to the previous pathogens, but also to other unrelated pathogens, e.g., viruses, bacteria, oomycetes, and fungi [50]. Moreover, SAR can even be passed onto progeny through epigenetic regulation [51]. Salicylic acid (SA) is believed to be the key plant hormone for establishing SAR [52]. NON-EXPRESSOR OF PR1 (NPR1), an ankyrin domain-containing protein, is the core component responding to SA signaling and establishing SAR [49,53]. Besides NPR1, the model plant Arabidopsis encodes other two NPR1-like homologs, namely, NPR3 and NPR4 [54]. It was proposed that the local innate immune responses cause increments of SA concentration in the non-infected tissues that disrupt NPR1–NPR3, but promote interaction between NPR1–NPR4, and finally induce the expression of PATHOGEN-RELATED (PR) genes [55]. However, this hypothesis is challenged by a recent study, which provides genetic evidence supporting NPR3 and NPR4 as redundant immune repressors [56]. Nevertheless, NPR1 is not only required for SAR, but is also necessary for robust immune responses in both PTI and ETI, as the knockout of NPR1 attenuates PR gene expression [53]. Thus, NPR1 might be in the core position of plant innate immunity that allows plants to efficiently and dynamically balance immune responses and normal development. Indeed, NPR1 is heavily modified at the post-translational level to ensure the proper regulation of the immune responses [53].
2.2. RNA Silencing
RNA silencing, which is also called RNA interference (RNAi), is an evolutionary conserved and sequence-specific mechanism for regulating endogenous gene expression and fighting against foreign nucleic acids such as transposable elements and viruses [57]. The virus-derived double-stranded RNA (dsRNA), which is the key trigger of antiviral RNA silencing, is recognized and processed by plant type III endoribonucleases, the DICER-LIKE (DCL) proteins, into small 20 to 24-nucleotide (nt) RNA duplexes termed virus-derived short-interfering small RNAs (vsiRNAs) [58,59]. The vsiRNAs are incorporated into ARGONAUTE (AGO) proteins that form the core component of the RNA-induced silencing complex (RISC), which is able to directly cleave homologous viral RNAs and/or suppress viral protein translation [60,61,62]. RNA silencing has been recognized as one of the primary antiviral defense mechanisms in the compatible plant–virus interactions, as knockout of the viral suppressor of RNA silencing (VSR) usually results in the loss-of-infectivity of the virus. For instance, a turnip mosaic virus (TuMV, a potyvirus) mutant lacking the major VSR (HcPro) cannot infect wild-type Arabidopsis; however, it regains infectivity on a mutant plant in which the RNA silencing pathway is also impaired [63]. Besides, the phenomenon of symptom recovery is closely associated with enhanced RNA silencing and the dysfunction of VSR activity [64,65]. In fact, it is believed that almost all plant viruses have evolved one or more proteins that are able to interrupt RNA silencing [66]. RNA silencing can also determine the incompatibility of a plant–virus interaction. For instance, Arabidopsis is normally recognized as non-host for PVX. However, PVX can infect Arabidopsis in the presence of pepper ringspot virus (PepRSV, a potyvirus), or with the assistance of the VSR of PepRSV [67]. Further research showed that the inhibition of the replication of PVX in Arabidopsis is mainly dependent on DCL2 and DCL4, and the cooperative activity of AGO2 and AGO5 [67,68,69].
Similar to innate immunity, RNA silencing also (1) perceives extraneous dsRNA molecules in a nucleotide sequence-independent manner; (2) conducts a signal amplification process; and (3) establishes resistance against homologous virus infection on non-infected tissues via the cell-to-cell and systemic movement of vsiRNA duplexes [70,71]. However, unlike PTI, RNA silencing does not initiate typical innate immunity responses, such as PR gene expression and HR development, and is independent of small molecules, e.g., SA, reactive oxygen species (ROS), or calcium ions, as secondary messengers for signal amplification.
2.3. Translation Repression
The cellular protein biosynthesis is tightly regulated in response to nutrient starvation or stresses. It is believed that GENERAL CONTROL NON-DEREPRESSIBLE-2 (GCN2), which is a serine/threonine-protein kinase, is the key regulator of protein biosynthesis in plants. GCN2 functions by targeting and phosphorylating EUKARYOTIC TRANSLATION INITIATION FACTOR-2A (eIF2α), which binds Met-tRNA and transfers it to the 40S ribosomal subunit to initiate the synthesis of a protein upon bonding to guanosine triphosphate (GTP) [72,73]. The phosphorylation of eIF2α at serine 52 inhibits the recycling of eIF2α-bound GDP to GTP, thereby prohibiting protein synthesis [74]. The activation of GCN2 itself is regulated by an ATP-binding cassette (ABC) family protein, GCN1 [75,76]. The GCN1–GCN2–eIF2α pathway is involved in plant response to many biotic and abiotic stresses, such as wounding and cold stress [76,77]. A direct role for GCN2-mediated protein translation repression in antiviral defense has not been reported, and the infection of wild-type or GCN2-knockout Arabidopsis by turnip yellow mosaic virus (TYMV, a tymovirus) or TCV was shown not to cause the phosphorylation of eIF2α [72]. These data suggest that either GCN2 does not play a role in the plant response to viral infection or, if it does, its role has been co-opted by these two adapted viruses. Further studies are needed in order to fully illustrate these possibilities [78]. Plant viruses thoroughly rely on host protein translation machinery for protein biosynthesis; as a result, loss-of-function of the component(s) of translation machinery that are recruited by plant virus for viral protein synthesis usually begets loss-of-susceptibility to the virus, which is a phenomenon known as recessive resistance [79].
Ribosome-inactivating proteins (RIPs) form a family of proteins that is ubiquitously distributed in plants, mushrooms, and bacteria and can inhibit protein synthesis by depurinating the sarcin/ricin loop (SRL) of rRNA [80,81]. Research demonstrated that many RIPs have potent antiviral, antifungal, and insecticidal activities. The best-characterized RIP with antiviral activity is the pokeweed antiviral protein (PAP) from pokeweed (Phytolacca americana). PAP has been demonstrated to be able to reduce the propagation of many plant viruses, e.g., cucumber mosaic virus (CMV, a cucumovirus), PVX, tobacco etch virus (TEV, a potyvirus), alfalfa mosaic virus (AMV, an alfamovirus), potato virus Y (PVY, a potyvirus), brome mosaic virus (BMV, a bromovirus), African cassava mosaic virus (ACMV, a begomovirus), cauliflower mosaic virus (CaMV, a caulimovirus) [82,83,84,85,86], and many animal viruses as well [87]. Moreover, the expression of RIPs in plants is increased under stress conditions, e.g., virus attacking [88], suggesting that they are parts of a plant antiviral mechanism via global translation repression. Indeed, there was a burst of research that sought to utilize RIPs to achieve broad-spectrum antiviral resistance around 1990–2000s; however, their application in agricultural practice is hampered by the potent cytoplasmic toxicity to the plant itself and the animals that consume it [85,89]. How to reduce the cytoplasmic toxicity while maintaining its antiviral activities is a challenge for future research [84].
Recently, another antiviral mechanism that functions by suppression of the global cellular protein synthesis was discovered [90]. NUCLEAR SHUTTLE PROTEIN (NSP) INTERACTING KINASE 1 (NIK1), the key component of this antiviral machinery, was first identified as a geminiviral NSP-interacting host factor [91]. NIK1 is a plasma membrane-anchored LRR protein carrying a functional serine/threonine (Ser/Thr) kinase domain resembling the PRR co-receptor BAK1 [92]. Two additional homologs (NIK2 and NIK3) were identified in Arabidopsis, which together with NIK1 form a separate cluster belonging to group I of the LRR-RK superfamily [93]. Unlike BAK1-mediated resistance, NIK1-mediated antiviral responses do not induce typical innate immune reactions, e.g., the expression of PR genes; instead, NIK1 phosphorylates the cytoplasmic RIBOSOMAL PROTEIN 10 (RPL10) and results in the translocation of RPL10 from the cytoplasm to the nucleus [94]. In the nucleus, RPL10 interacts with an myeloblastosis (MYB) domain-containing transcriptional repressor, L10-INTERACTING MYB DOMAIN-CONTAINING PROTEIN (LIMYB), to repress the expression of ribosomal protein genes, and finally leads to the shutdown of global cellular and viral protein synthesis [90].
Besides global translation repression, plants also have developed specific translation repression machinery. One such example is small RNA-related translation repression. As addressed in the above content, the binding of miRNA-associated and short interfering RNA (siRNA)-associated RISCs to cellular or viral RNA, in addition to endonucleolytic cleavage, also causes direct translation inhibition [61,62,95,96]. Similar to RNA silencing, the specificity of small RNA-related translation repression is determined by the sequence of small RNA; thus, the overall protein synthesis is largely not affected [97]. Small RNA-related translation repression can play an important role in plant–virus interaction, and even determines the outcome of the battle. For instance, symptom recovery in tomato ringspot virus (ToRSV, a nepovirus)-infected plants is associated with the AGO1-dependent translation repression of viral RNA2 [61]. Recently, SHIFTLESS (SFL), a broad-spectrum inhibitor of –1 ribosomal frameshifting, was isolated from Human T cell leukemia (MT4 cell) [98]. Ribosomal frameshifting is also a prevailing protein translation mechanism in plant viruses; thus, a similar ribosomal frameshifting inhibition mechanism might also exist in plants.
2.4. Atypical Dominant Viral Resistances
The exploration of viral resistance in the last decade also resulted in the discovery of some dominant resistance genes that are independent of the typical innate immunity signaling cassette for function. The products of these dominant resistance genes are structurally different from each other and from the typical R proteins, and, thus cannot be integrated into the conventional plant innate immunity. Based on the current understanding, most of these dominant resistance genes likely function by directly interacting with viral proteins to inhibit their activity. Hereafter, we referred to the products of these dominant resistance genes as ATYPICAL DOMINANT VIRAL RESISTANCE PROTEINs (ADVRPs).
The major ADVRPs discovered so far are members of the lectin protein family. Lectins, which are also known as agglutinins or jacalins, are a group of the carbohydrate-binding domain (CBD)-containing proteins that bind reversibly to specific monosaccharides or oligosaccharides [99]. Besides the CBD, many lectins also possess additional catalytic domains, such as glucanase, glycosidase, kinase, and RIP as well. Some kinase domain-containing lectins, such as DOES NOT RESPOND TO NUCLEOTIDES 1 (DORN1) and LIPOOLIGOSACCHARIDE-SPECIFIC REDUCED ELICITATION (LORE) from Arabidopsis and I-3 from tomato, function as the PRR receptors in PTI [100,101,102]. RESTRICTED TEV MOVEMENT (RTM) 1 is a lectin-like protein in Arabidopsis that specifically confers resistance to several potyviruses, e.g., TEV, lettuce mosaic virus (LMV), and plum pox virus (PPV), by restricting their long-distance movement [103,104]. The RTM1-mediated resistance is independent of HR, and neither involves SA nor induces SAR; instead, it is thought to form a complex in the phloem with at least two additional proteins, RTM2 (a small heat shock protein) and RTM3 (a meprin with the tumor necrosis factor receptor associated factor (TRAF) homology domain) and other host factors, as the products of the RTM4 and RTM5 genes, which were only genetically characterized [103,105,106,107,108,109]. The loss-of-function of either member in the complex results in the evanishment of the resistance, suggesting that RTM1-mediated resistance is dependent on the protein complex [108]. This protein complex likely functions by targeting the potyviral viral particle or CP-containing ribonucleoprotein complex in the phloem, as CP is involved in the RTM-mediated resistance breaking [110]. JACALIN-TYPE LECTIN REQUIRED FOR POTEXVIRUS RESISTANCE1 (JAX1) is another lectin-like ADVRP of Arabidopsis that confers broad-spectrum resistance to potexviruses at the early infection stage through recognizing viral RNA-dependent RNA polymerase (RdRp) and inhibiting its activity [111,112,113]. Other examples including a lectin-like protein (CIP-29) isolated from Cyamopsis tetragonoloba (L.) that is able to confer tobacco with resistance against the Sunn-hemp mosaic virus (SHMV, a tobamovirus) and a lectin from Musa paradisiaca (BanLec-1) that binds to the CP of TMV and prevents viral infection [114,115].
Other ADVRPs, such as the tomato Tm-1, which confers resistance to tomato mosaic virus (ToMV, a tobamovirus) [116] and VIOLAXANTHIN DEEPOXIDASE (ZmVDE) and h-type thioredoxin (ZmTrxh), which bestow maize with sugarcane mosaic virus (SCMV, a potyvirus) resistance [117,118], are structurally different from each other and cannot be classified into a typical protein family. Tm-1 is a T-cell immunoglobulin and mucin (TIM)-barrel-like domain-containing protein that is able to directly bind the replication proteins of ToMV and inhibit its activity [116]. ZmVDE functions by interacting with helper component-proteinase (HcPro), the VSR of SCMV, and attenuates its silencing suppression activity [118], whereas ZmTrxh inhibits viral RNA accumulation in the cytoplasm [117]. There are also several ADVRPs that probably do not function by interacting directly with viral proteins. For instance, STV11, a sulfotransferase from rice, confers rice with RSV resistance by catalyzing the conversion of SA into sulfonated SA [119]. Allelic Ty-1 and Ty-3 encode a γ-class RdRp that confers resistance to tomato yellow leaf curl virus (TYLCV, a begomovirus), and the hypermethylation of the genomic DNA has been hypothesized as the possible source of the resistance [120,121].
As research progresses, the number of ADVRPs will continue expanding. However, the origin of these ADVRPs is still a mystery at the present. A likely possibility is that these proteins have primary functions that are other than antiviral, and incidentally acquired the ability to inhibit virus proliferation [122]. For instance, Rsp5p, a yeast Nedd4 family E3 ubiquitin ligase that is normally involved in endocytosis [123], is able to interact with the replication proteins (p33 and p92) of tomato bushy stunt virus (TBSV, a tombusvirus), and mediates their degradation [124]. However, it is also possible that the resistance mediated by some ADVRPs is a conserved antiviral mechanism. For instance, lectins have been associated with antiviral activity since their discovery [99].
2.5. Ubiquitination and Autophagy-Mediated Protein Degradation
Ubiquitination-mediated protein degradation is the major protein turnover machinery in the plant. Ubiquitination is a multistep enzymatic reaction that is able to covalently attach ubiquitin (Ub) to the target proteins [125]. First, Ub precursor is proteolyzed by the Ub-activating enzymes (E1) and forms an E1–Ub intermediate in which the C-terminal glycine of Ub is linked via a thioester bond to a cysteine in E1. Next, the activated Ub is transferred to Ub-conjugating enzymes (E2). Finally, the Ub is delivered from the E2–Ub intermediate to target proteins with the help of Ub ligases (E3). Then, the ubiquitinated proteins are sent to the 26S proteasome for degradation. Ubiquitination-mediated protein degradation is pivotal for cellular protein homeostasis, and it is involved in nearly all cellular processes; therefore, it is not surprising that it is involved in almost all plant antiviral defense mechanisms. For instance, the full induction of PR genes’ expression in plant innate immunity involves the proteasome-mediated turnover of the transcription co-activator NPR1 [126]. Furthermore, the abundance of chitin receptor LYSIN MOTIF RECEPTOR KINASE5 (LYK5) is regulated by Arabidopsis E3 ubiquitin ligase PLANT U-BOX13 (PUB13) [127]. Unfolded protein response and endoplasmic reticulum (ER) stress, a complex and multifaceted intracellular signal pathway that is essential for reestablishing ER homeostasis, constitutes another example of plant response to viral infection regulated by ubiquitination-mediated protein degradation [128].
As the specificity of ubiquitination-mediated protein degradation is determined largely by E3, both plants and animals encode only a few E1s and E2s, and a large number of E3s. For instance, the model plant Arabidopsis encodes two E1s, 37 E2s, and more than 1400 E3s [129]. The masses of E3s allow plant cells to target, in addition to numerous endogenous proteins, exogenous proteins from invading pathogens. Hitherto, a group of E3 ligases was found to be able to specifically interact with typical viral proteins and mediate their degradation [130]. For instance, the tobacco really interesting new gene (RING) class E3 ligase Nicotiana tabacum RING FINGER PROTEIN 1 (NtRFP1) interacts with the βC1 protein encoded by the tomato yellow leaf curl China virus (TYLCCNV)-associated betasatellite, which is a viral pathogenesis factor, and VSR [131,132,133], to mediate its ubiquitination and degradation to attenuate virus proliferation [134]. Besides, the ubiquitination-mediated protein degradation is also involved in countering the RdRp of TYMV in Arabidopsis [135,136], and the CELL-DIVISION-CYCLE protein48 (CDC48), a conserved chaperone controlling protein fate in eukaryotes, extracts the movement protein (MP) of TMV from the ER to the cytosol for ubiquitination-mediated degradation [137,138]. However, whether the degradation of these viral proteins via ubiquitination is a defense mechanism of the plant or is the normal physiological homeostasis of the viral proteins needs to be carefully discriminated.
Autophagy is conserved machinery for transporting unwanted or misfolded protein aggerates or damaged organelles to the vacuole for degradation and recycling. In plants, it is pivotal for maintaining cellular homeostasis under normal conditions and resisting abiotic and biotic stresses [139]. Similar to ubiquitination, autophagy is also related to almost all aspects of cell physiological processes, including plant immunity [140,141]. Autophagy is involved in plant immunity in many ways, such as through balancing the homeostasis of immunity signaling components, degrading defense-related plant proteins, and regulating HR [140,142]. Plants also utilize selective autophagy to degrade virus-encoded protein aggregates, ribonucleoprotein, and even particles, which suggests that autophagy functions as a separate antiviral mechanism. Recently, several reports have highlighted the importance of this pathway in retarding virus proliferation: AUTOPHAGY-RELATED GENE6 (ATG6, also called BECLIN1), the core component of autophagy, interacts with NUCLEAR INCLUSION PROTEIN B (NIb), the RdRp of TuMV, to inhibit virus replication [143]. AUTOPHAGY-RELATED GENE8 (ATG8) specifically interacts with the βC1 of cotton leaf curl Multan virus (CLCuMuV)-associated betasatellite for degradation to impede its replication [144]. The autophagy cargo receptor, NEIGHBOR OF BRCA1 (NBR1), targets both unassembled CP and virus particles of CaMV to mediate their autophagy-dependent degradation, thereby restricting the establishment of CaMV infection [145]. Besides, NBR1 also suppresses TuMV accumulation by targeting HcPro, which is presumably in association with virus-induced RNA granules [146,147].
2.6. Cross-Talking between Different Antiviral Defenses
Plant antiviral mechanisms do not function separately; in contrast, there is vigorous cross-talking between different defense programs. In general, there are three types of cross-talking. Firstly, the component of one antiviral pathway is always regulated by other antiviral pathways. For instance, the transcripts of R genes are regulated by microRNAs, e.g., miR472, and further targeted by the trans-acting siRNAs (ta-siRNAs) or siRNAs produced from their transcripts by plant RNA-DEPENDENT RNA POLYMERASE 6 (RDR6), and the expression of the components of the RNA silencing pathway are also regulated by innate immunity responses [148,149,150]. Secondly, one mechanism needs the function of other pathways; for example, the HR of plant innate immune responses are regulated by autophagy and ER stress [140,151,152]. Finally, but most importantly, different types of mechanisms work cooperatively against plant viruses. For instance, the establishment of SAR involves SA-mediated innate immunity, RNA silencing, and even the protein secretory pathway [49,153]. SA is also involved in plant growth regulation by controlling gibberellin biosynthesis via endogenous small RNAs in the antiviral response to a potyvirus [154]. A calmodulin-like protein, REGULATOR OF GENE SILENCING CALMODULIN-LIKE PROTEIN (rgs-CaM), counterattacks intracellular VSRs by binding to their dsRNA-binding domains, mediates their degradation via the autophagy pathway, and also initiates SA-associated antiviral responses [155,156,157]. This sophisticated and dynamic cross-talk between different types of defense mechanisms allows the plant to respond quickly and efficiently to the infection of viruses and minimize the trade-off between defenses and normal development.
3. Strike Back from Viruses
Plant viruses have also gained multiple strategies to suppress and even exploit host defenses to ensure their successful infection [1]. Despite their small genome size (less than 20 kilobases) and that they only encode a few proteins, plant viruses have been endowed with great genome flexibility and multifunctional proteins to fight against the host’s sophisticated defense mechanisms. In general, plant viruses are able to strike back on every aspect of plant defense (Figure 1).
3.1. Mutation as a Master Mechanism for Escaping Host Antiviral Mechanisms
Due to the lack of proofreading viral RdRp, both plant and animal RNA viruses have a much higher mutation rate than that of their host cell, which is replicated via the DNA polymerase [158,159]. Plant DNA viruses, such as geminiviruses and nanoviruses, can evolve as quickly as their RNA counterparts [160,161]. As addressed in the previous section, plant antiviral mechanisms including PTI, ETI, and ADVRP-mediated resistance, are almost exclusively triggered by the recognition of a particular short sequence within the viral protein by PRRs, R proteins, or ADVRPs. As a result, it is easy for plant viruses to alter the amino acids responding to the perception to escape host immune response, which is a phenomenon called resistance breakdown. There are numerous such examples, for instance: one amino acid alteration in the VIRAL PROTEIN GENOME-LINKED (VPg) protein of Rice yellow mottle virus (RYMV, a sobemovirus) results in the loss of resistance of the Oryza glaberrima Tog7291 line carrying the RYMV2 resistance gene [162]. The breakdown of sugar beet Rz1-mediated resistance against Beet necrotic yellow vein virus (BNYVV, a benyvirus) infection can be caused by a single mutation in its p25 coding sequence [163]. A single amino acid in the RdRp of PVX is responsible for JAX1-and Tm-1-mediated resistance breakdown [112,164]. Achieving long-term resistance through dominant resistance genes is a big challenge in crop breeding.
3.2. Inhibition of Innate Immunity
Despite the wealth of knowledge on how other phytopathogens and animal viruses suppress host immune responses [31,165], how plant viruses suppress plant innate immunity is just starting to be uncovered. Recently, several viral proteins, including the CP of PPV, the MP of CMV, and the P6 of CaMV, have been shown to be able to interfere with plant PTI signaling, including ROS production and SA accumulation, and eventually increase the susceptibility of the host to other pathogens [166,167,168]. However, the intracellular target of these viral proteins and how they suppress PTI is still elusive. More recently, we found that the NIb protein of TuMV can suppress host immune responses as well [169]. Moreover, we identified its role in the process of a host factor, SMALL UBIQUITIN-LIKE MODIFIER 3 (SUMO3), which can interact with and sumoylate NIb via a SUMO-interacting motif (SIM) at the C-terminal domain of the viral protein. Interestingly, suppression of host immune responses by NIb is dependent on SUMO3-mediated sumoylation [169]. SUMO3 is strongly and widely induced by SA and the defense elicitor Flg22 [170], and directly participates in plant innate immunity through fine-tune a regulating NPR1 function [171]. These results thus suggested that NIb functions through the disruption of the SUMO3 function to suppress plant antiviral immunity.
3.3. Suppression and Exploitation of Host RNA Silencing
Antiviral RNA silencing mainly occurs in the cytoplasm, where the replication of most plant RNA viruses takes place. To encounter host RNA silencing, the replication of many positive-sense single-strand RNA (+ssRNA) viruses are concealed in membranous inclusion bodies, vesicles, multivesicular bodies, or spherules that are remodeled from the plant cell endogenous membranes by viral proteins [172,173,174,175]. These membrane-sheltered virus replication factories allow the minimization of the disclosure of dsRNA that is generated by viral RdRps and is targeted by DCL proteins to trigger RNA silencing. All endogenous membranes, e.g., the ER, the membrane of chloroplasts, mitochondria, peroxisomes, and vacuole, have been exploited by plant +ssRNA viruses for replication. It is noteworthy that there is no obvious conservation in utilizing typical endomembranes among plant +ssRNA viruses of the same family or even of the same genus, and plant +ssRNA viruses from different families might use the same endomembrane to replicate. For instance, the carnation Italian ringspot virus (CIRV) and tomato bushy stunt virus (TBSV), which are both from the genus Tombusvirus within the family Tombusviridae, have very a similar genome structure, particle morphology, and replication strategy. However, these two viruses utilize the outer membrane of mitochondria and peroxisomes for replication, respectively [176,177]. Meanwhile, the TuMV (family Potyviridae), TMV (family Virgaviridae), and BMV (family Bromoviridae) hold replication vesicles or organelle-like structures derived from the ER [178,179,180]. Thereby, the endomembrane is selected “randomly” by the viruses to evade the evolutionary pressure imposed by host RNA silencing. The disturbance of plant endomembranes, especially the ER, by viruses always triggers ER stress responses, which are also implicated in plant reaction against other biotic and abiotic challenges and in normal development [181]. Interestingly, studies showed that the ER stress responses actually promote viral proliferation [182,183,184,185,186], suggesting that the ER stress signal pathway has been exploited by plant viruses for their own benefits.
Membranous virus replication factories are not sufficient to support robust viral replication, as abnormally abundant viral RNA in the plant cell can also trigger RNA silencing through the de novo synthesis of dsRNA by the host RdRp, which in turn inhibits viral protein synthesis and viral particle assembly. For instance, a CMV mutant that lacks the protein suppressing host RNA silencing (2b), can only replicate and cause systemic infection with extremely low efficiency [187,188]. Remarkably, most plant viruses, including RNA and DNA viruses, have evolved one or more VSRs to directly block the host RNA silencing mechanism [189]. A large number of VSRs encoded by varied plant viruses have been characterized. It is beyond the scope of this review to describe the mechanisms of action of each VSR in detail [189,190,191]. However, it is worth highlighting that the VSRs encoded by different viruses share no obvious amino acid sequence or structure similarity, suggesting they have independent origins. Nevertheless, VSRs can largely be divided into two classes based on their function mechanisms. The first class consists of VSRs functioning by sequestering viral dsRNA or vsiRNAs and/or by disrupting their biogenesis, such as the p19 of tombusviruses, NS3 of tenuiviruses, or the small replicase subunit (p122) of tobamoviruses. The second class consists of those functioning by subverting the components of the host RNA silencing pathway, such as the VPg of potyviruses, ßC1 of the betasatellite of begomoviruses, TGBp1 (p25) of potexviruses, and P0 of poleroviruses, although some VSRs, such as the HcPro of potyviruses and 2b of cucumoviruses, can both sequester vsiRNAs/dsRNA and disrupt RNA silencing components [190,191,192].
Plant viruses can also exploit the host RNA silencing system to promote their replication. Plant viruses exploit host RNA silencing mainly in two ways: one is by regulating endogenous miRNA expression, and another is by suppressing endogenous gene expression through viral-derived small-interfering RNAs (vsiRNAs). One such example is the infection of cymbidum ringspot virus (CymRSV, a tombusvirus), whose VSR (p19) can specifically upregulate the expression of miR168 to downregulate the transcripts of AGO1, which is a target of miR168 [193]. A similar phenomenon is involved in the breakdown of the RSV1-mediated resistance of the soybean cultivar PI96983 of the strain G7 of soybean mosaic virus (SMV, a potyvirus) [194]. Recently, Yang et al. showed that the intergenic siRNAs of TYLCV target a host long non-coding RNA to modulate disease symptoms [195]. Indeed, comparative analyses revealed that vsiRNAs have a wide-ranging and significant influence on the host transcriptome [196,197,198,199,200,201]. A more detailed outlook concerning how plant viruses utilize vsiRNAs to regulate endogenous gene expression can be found in these reviews [59,202].
3.4. Subversion of Translation Repression
A substantial amount of evidence suggests that animal viruses are able to subtly manipulate host protein synthesis machinery and adopt numerous unconventional translation mechanisms for a productive proliferation [203,204]. Plant viruses have also evolved numerous unconventional strategies that are both cap-dependent, and cap-independent to recruit the cellular translational machinery for the efficient synthesis of their proteins [205]. Thus, it is reasonable to believe that plant viruses are also able to “shut off” host protein biosynthesis, even though a direct link between the GCN1-GCN2-eIF2α regulatory pathway and plant virus infection has not been revealed yet.
NSP is the geminivirus-encoded membrane protein that counteracts NIK1-mediated translation repression [206]. NSP directly binds to the kinase domain of NIK1 and prevents its activation and the consequent downstream signaling by interrupting the phosphorylation of the key threonine residue at the position 474 [206]. The NSP–NIK interaction is conserved among geminivirus NSPs and NIK homologs from different hosts [206], suggesting that NIK-mediated translation suppression is a general plant antiviral defense that is successfully overcome by geminiviruses. Thus, NIK could be an excellent candidate for the development of broad-spectrum resistance against geminiviruses [207]. However, whether NIK-mediated translation repression is activated during RNA virus infection and has an antiviral effect against these viruses is still unknown at the present.
3.5. Prevention and Exploitation of Host Ubiquitination and Autophagy Pathways
Plant viruses also have developed several strategies to subvert host ubiquitination-mediated and autophagy-mediated antiviral machinery, e.g., by encoding deubiquitinases that target different endogenous and viral proteins [130]. For instances, the ubiquitination-mediated degradation of TYMV p66 can be counteracted by the interaction with viral 98K RdRp, which is also a functional deubiquitinating enzyme [208]. On the other hand, the βC1 protein disrupts the plant ubiquitination pathway by interacting with SPK1, and enhances geminivirus infection probably by subverting its own NtRFP1-mediated ubiquitination and degradation [209]. NBR1 suppresses TuMV infection by targeting HcPro protein for degradation [146]. However, NBR1-mediated HcPro degradation can be blocked by other viral proteins, such as VPg and 6K2 [146]. P6 protein, the major CaMV pathogenicity factor, suppresses SA-dependent autophagy [168], indicating that the autophagy-mediated restriction of CaMV proliferation could be diminished by P6.
Plant viruses are able to exploit ubiquitination and autophagy to promote their proliferation by encoding ubiquitin ligases or molecular adaptor proteins that target specific host factors for degradation. A large number of such examples have been reported, including the P0 protein of poleroviruses and enamoviruses, an F-box domain-containing protein, and PVX TGBp1 target AGO proteins for degradation via ubiquitin-mediated proteasome and autophagy pathways [210,211,212,213,214]; CLINK, another F-box domain-containing protein of faba bean necrotic yellows virus (FBNYV, a nanovirus), binds to SKP1 and the cell cycle protein pRB to promote virus replication [215,216]; rice stripe virus (RSV, a tenuivirus) MP (NSvc4) interferes with the S-acylation of remorin and induces its autophagic degradation to facilitate virus infection [217]; the geminivirus-encoded TRANSCRIPTIONAL ACTIVATOR PROTEIN (TrAP) protein, which is also known as C2, L2, AC2, or AL2, co-opts SKP1-CULLIN F-BOX PROTEIN (SCF)-mediated ubiquitination [218]; the VPg protein of potyviruses targets SUPPRESSOR OF GENE SILENCING 3 (SGS3) for degradation via both ubiquitination and autophagy pathways [219]. Recently, Yang et al. showed that the γb protein encoded by barley stripe mosaic virus (BSMV, a hordeivirus) can subvert host autophagy by disrupting the interaction between ATG7 and ATG8, which are two key regulators of the process [220], to promote viral infection [221]. Many animal viruses utilize autophagy machinery to assemble the membrane-bonded virus replication-associated structures [222,223,224,225,226]; however, a similar phenomenon has not been reported in plant +ssRNA viruses, even though the replication of most plant (+) RNA viruses are also associated with endomembranes [172].
4. Perspectives
Compared to the plant defense against other extracellular micropathogens, e.g., fungi and bacteria, in some aspects, plant antiviral defense could be more complex. This is apparent due to the intracellular parasitism of plant viruses, in which all genetic materials are directly explored by and connected with plant intracellular factors. Although this direct interaction allows the plant to evolve new defense mechanisms targeting viral factors, plant viruses can also take advantage of the interaction to explore the weak points of plant antiviral barriers and exploit their error-prone polymerases and multiple functional proteins to quickly escape host defenses and win the arms race. The ultimate goal of plant–virus interaction research is to establish sustainable virus resistance strategies to ensure food safety for the expanding human population. Indeed, considerable successes have been made in the management of viral diseases for varied crops in the last decade. For instance, R gene-directed breeding slowed down the ravages of many destructive plant viruses, such as tospoviruses, potyviruses, and begomoviruses [227,228]. Growing knowledge is also allowing researchers to develop engineered virus resistances, such as pathogen-derived resistance, RNA silencing-based resistance, pathogen-targeted resistance through zinc finger nucleases (ZiF), TRANSCRIPTION ACTIVATOR-LIKE EFFECTOR NUCLEASE (TALEN), and the CLUSTERED REGULARLY INTERSPACED PALINDROMIC REPEAT (CRISPR)/Cas9 system [229,230]. Small RNA-based genetic engineering has been commercially applied in engineering viral resistance for a number of crops, such as papaya, plum, squash, potato, pepper, and tomato [231]. Although multiple virus resistances have been achieved in several recent studies [232,233,234,235,236,237], preventing or retarding the virus resistance breakdown is still a major challenge in agriculture practice. Recently, several novel approaches have been proposed to this end, e.g., by stacking or pyramiding resistance genes, by altering known R proteins specificities and/or expanding resistance, by the exogenous application of dsRNA or siRNAs, and by the utilization of PRRs [238,239,240,241,242,243,244,245]. Nevertheless, how to develop novel antiviral strategies with broad-spectrum, efficient, sustaining, and environment-friendly resistances is still an open question.
Despite signs of progress in the last decade, many mysteries still need to be addressed, e.g., what molecular mechanisms are involved in incompatible interactions between plants and viruses, what virus-derived PAMPs and their respective PRRs exist, and what mechanisms underlie the mixed infection-associated resistance breakdown. For instance, the pre-infection of tomato cultivars that carry the Sw-5 gene (which confers broad-spectrum resistance to tospoviruses through the recognition of a conserved 21-amino acid viral epitope [28]) with tomato chlorosis virus (ToCV, a crinivirus) results in susceptibility to tomato spotted wilt virus (TSWV, a tospovirus) [246]. Thus, these unexploited areas are especially worth investigating in the future.
Funding
This study is financially supported by the National Natural Science Foundation of China (grant no: 31671998), the Natural Science Foundation of Heilongjiang Province (grant no: ZD2018002), and the Scientific Research Foundation of Department of Education of Heilongjiang Province for the Returned Overseas Chinese Scholars (grant no: 2018QD0002). J.A.G. and A.V. were supported by grants BIO2016-80572-R and BIO2015-73900-JIN and the FEDER program.
Acknowledgments
We apologize to the authors whose relevant work on the defense and counter-defense between plant and virus has not been cited owing to length constraints. We would like to express our special appreciation to Aiming Wang from the London Research and Development Centre of Agriculture and Agri-Food Canada for his constructive suggestions to the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wang, A. Dissecting the molecular network of virus-plant interactions: The complex roles of host factors. Annu. Rev. Phytopathol. 2015, 53, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J. The dependence of viral RNA replication on co-opted host factors. Nat. Rev. Microbiol. 2012, 10, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pendon, J.A.; Truniger, V.; Nieto, C.; Garcia-Mas, J.; Bendahmane, A.; Aranda, M.A. Advances in understanding recessive resistance to plant viruses. Mol. Plant Pathol. 2004, 5, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Krishnaswamy, S. Eukaryotic translation initiation factor 4E-mediated recessive resistance to plant viruses and its utility in crop improvement. Mol. Plant Pathol. 2012, 13, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Robaglia, C.; Caranta, C. Translation initiation factors: A weak link in plant RNA virus infection. Trends Plant Sci. 2006, 11, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, K.K.; Scholthof, K.-B.G. Plant immune responses against viruses: How does a virus cause disease? Plant Cell 2013, 25, 1489–1505. [Google Scholar] [CrossRef] [PubMed]
- Soosaar, J.L.M.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of plant resistance to viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Schwessinger, B.; Zipfel, C. News from the frontline: Recent insights into PAMP-triggered immunity in plants. Curr. Opin. Plant Biol. 2008, 11, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Early molecular events in PAMP-triggered immunity. Curr. Opin. Plant Biol. 2009, 12, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Boutrot, F.; Zipfel, C. Function, discovery, and exploitation of plant pattern recognition receptors for broad-spectrum disease resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.M.; Tang, D.; Wang, G. Receptor kinases in plant pathogen interactions: More than pattern recognition. Plant Cell 2017, 29, 618–637. [Google Scholar]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kørner, C.J.; Klauser, D.; Niehl, A.; Domínguez-Ferreras, A.; Chinchilla, D.; Boller, T.; Heinlein, M.; Hann, D.R. The immunity regulator BAK1 contributes to resistance against diverse RNA viruses. Mol. Plant Microbe Interact. 2013, 26, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Gou, X.; He, K.; Xi, D.; Du, J.; Lin, H.; Li, J. BAK1 and BKK1 in Arabidopsis thaliana confer reduced susceptibility to turnip crinkle virus. Eur. J. Plant Pathol. 2010, 127, 149–156. [Google Scholar] [CrossRef]
- Woo, J.Y.; Jeong, K.J.; Kim, Y.J.; Paek, K.H. CaLecRK-S.5, a pepper L-type lectin receptor kinase gene, confers broad-spectrum resistance by activating priming. J. Exp. Bot. 2016, 67, 5725–5741. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Singh, P.; Chen, M.C.; Zimmerli, L. L-Glutamine inhibits beta-aminobutyric acid-induced stress resistance and priming in Arabidopsis. J. Exp. Bot. 2010, 61, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. New Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, A.C.; Lapidot, M.; Culver, J.N.; Fluhr, R. An early tobacco mosaic virus-induced oxidative burst in tobacco indicates extracellular perception of the virus coat protein. Plant Physiol. 2001, 126, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Perraki, A.; Gronnier, J.; Gouguet, P.; Boudsocq, M.; Deroubaix, A.F.; Simon, V.; German-Retana, S.; Legrand, A.; Habenstein, B.; Zipfel, C.; et al. REM1.3’s phospho-status defines its plasma membrane nanodomain organization and activity in restricting PVX cell-to-cell movement. PLoS Pathog. 2018, 14, e1007378. [Google Scholar] [CrossRef] [PubMed]
- Zamora, M.; Méndez-López, E.; Agirrezabala, X.; Cuesta, R.; Lavín, J.L.; Sánchez-Pina, M.A.; Aranda, M.A.; Valle, M. Potyvirus virion structure shows conserved protein fold and RNA binding site in ssRNA viruses. Sci. Adv. 2017, 3, eaao2182. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, B.; Calil, I.; Machado, J.P.; Santos, A.; Fontes, E. Immune receptors and co-receptors in antiviral innate immunity in plants. Front. Microbiol. 2016, 7, 2139. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Jiang, L.; Bai, B.; Zhao, W.; Chen, X.; Li, J.; Liu, Y.; Chen, Z.; Wang, B.; Wang, C.; et al. The intracellular immune receptor Sw-5b confers broad-spectrum resistance to tospoviruses through recognition of a conserved 21-amino acid viral effector epitope. Plant Cell 2017, 29, 2214–2232. [Google Scholar] [CrossRef] [PubMed]
- Sarris, P.F.; Cevik, V.; Dagdas, G.; Jones, J.D.; Krasileva, K.V. Comparative analysis of plant immune receptor architectures uncovers host proteins likely targeted by pathogens. BMC Biol. 2016, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, H.; Zhu, M.; Huang, S.; Zhang, W.; Dinesh-Kumar, S.P.; Tao, X. A plant immune receptor adopts a two-step recognition mechanism to enhance viral effector perception. Mol. Plant. 2019. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, A.; Kanyuka, K.; Baulcombe, D.C. The Rx gene from potato controls separate virus resistance and cell death responses. Plant Cell 1999, 11, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Townsend, P.D.; Dixon, C.H.; Slootweg, E.J.; Sukarta, O.C.A.; Yang, A.W.H.; Hughes, T.R.; Sharples, G.J.; Palsson, L.O.; Takken, F.L.W.; Goverse, A.; et al. The intracellular immune receptor Rx1 regulates the DNA-binding activity of a Golden2-like transcription factor. J. Biol. Chem. 2018, 293, 3218–3233. [Google Scholar] [CrossRef] [PubMed]
- Tameling, W.I.; Nooijen, C.; Ludwig, N.; Boter, M.; Slootweg, E.; Goverse, A.; Shirasu, K.; Joosten, M.H. RanGAP2 mediates nucleocytoplasmic partitioning of the NB-LRR immune receptor Rx in the Solanaceae, thereby dictating Rx function. Plant Cell 2010, 22, 4176–4194. [Google Scholar] [CrossRef] [PubMed]
- Slootweg, E.; Roosien, J.; Spiridon, L.N.; Petrescu, A.J.; Tameling, W.; Joosten, M.; Pomp, R.; van Schaik, C.; Dees, R.; Borst, J.W.; et al. Nucleocytoplasmic distribution is required for activation of resistance by the potato NB-LRR receptor Rx1 and is balanced by its functional domains. Plant Cell 2010, 22, 4195–4215. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, P.; Yoshioka, K.; Shah, J.; Dooner, H.K.; Klessig, D.F. Resistance to turnip crinkle virus in Arabidopsis is regulated by two host genes and is salicylic acid dependent but NPR1, ethylene, and jasmonate independent. Plant Cell 2000, 12, 677–690. [Google Scholar] [CrossRef] [PubMed]
- del Pozo, O.; Lam, E. Expression of the baculovirus p35 protein in tobacco affects cell death progression and compromises N gene-mediated disease resistance response to tobacco mosaic virus. Mol. Plant Microbe Interact. 2003, 16, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Okamoto, M.; Iwai, T.; Iwano, M.; Fukui, K.; Isogai, A.; Nakajima, N.; Ohashi, Y. Reduced levels of chloroplast FtsH protein in tobacco mosaic virus-infected tobacco leaves accelerate the hypersensitive reaction. Plant Cell 2000, 12, 917–932. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.M.; Duncan, G.H.; Pradel, K.S.; Carr, F.; Wood, S.; Oparka, K.J.; Cruz, S.S. Analysis of the N gene hypersensitive response induced by a fluorescently tagged tobacco mosaic virus. Plant Physiol. 2000, 123, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, Y.; Takenaka, S.; Hase, S.; Kubota, M.; Ichinose, Y.; Kanayama, Y.; Nakaho, K.; Klessig, D.F.; Takahashi, H. Enhanced defense responses in Arabidopsis induced by the cell wall protein fractions from Pythium oligandrum require SGT1, RAR1, NPR1 and JAR1. Plant Cell Physiol. 2009, 50, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirasu, K. The HSP90-SGT1 chaperone complex for NLR immune sensors. Annu. Rev. Plant Biol. 2009, 60, 139–164. [Google Scholar] [CrossRef] [PubMed]
- Boter, M.; Amigues, B.; Peart, J.; Breuer, C.; Kadota, Y.; Casais, C.; Moore, G.; Kleanthous, C.; Ochsenbein, F.; Shirasu, K.; et al. Structural and functional analysis of SGT1 reveals that its interaction with HSP90 is required for the accumulation of Rx, an R protein involved in plant immunity. Plant Cell 2007, 19, 3791–3804. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Jeong, R.D.; Venugopal, S.C.; Lapchyk, L.; Navarre, D.; Kachroo, A.; Kachroo, P. SAG101 forms a ternary complex with EDS1 and PAD4 and is required for resistance signaling against turnip crinkle virus. PLoS Pathog. 2011, 7, e1002318. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Stuttmann, J.; Rietz, S.; Guerois, R.; Brunstein, E.; Bautor, J.; Niefind, K.; Parker, J.E. Structural basis for signaling by exclusive EDS1 heteromeric complexes with SAG101 or PAD4 in plant innate immunity. Cell Host Microbe 2013, 14, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.F.; Xu, L.; Tan, W.J.; Chen, L.; Qi, H.; Xie, L.J.; Chen, M.X.; Liu, B.Y.; Yu, L.J.; Yao, N.; et al. Disruption of the Arabidopsis defense regulator genes SAG101, EDS1, and PAD4 confers enhanced freezing tolerance. Mol. Plant 2015, 8, 1536–1549. [Google Scholar] [CrossRef] [PubMed]
- Feys, B.J.; Moisan, L.J.; Newman, M.A.; Parker, J.E. Direct interaction between the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4. EMBO J. 2001, 20, 5400–5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Gobbato, E.; Kracher, B.; Qiu, J.; Bautor, J.; Parker, J.E. A core function of EDS1 with PAD4 is to protect the salicylic acid defense sector in Arabidopsis immunity. New Phytol. 2017, 213, 1802–1817. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U. Systemic acquired resistance. Plant Signal Behav. 2006, 1, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.; Bruce, T.J.A.; Roberts, M.R.; Flors, V.; Ton, J. Next-generation systemic acquired resistance. Plant Physiol. 2012, 158, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Glazebrook, J.; Clarke, J.D.; Volko, S.; Dong, X. The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell 1997, 88, 57–63. [Google Scholar] [CrossRef]
- Withers, J.; Dong, X. Posttranslational modifications of NPR1: A single protein playing multiple roles in plant immunity and physiology. PLoS Pathog. 2016, 12, e1005707. [Google Scholar] [CrossRef] [PubMed]
- Dong, X. NPR1, all things considered. Curr. Opin. Plant Biol. 2004, 7, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cheng, Y.T.; Qu, N.; Zhao, Q.; Bi, D.; Li, X. Negative regulation of defense responses in Arabidopsis by two NPR1 paralogs. Plant J. 2006, 48, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Schauer, S.E.; Jacobsen, S.E.; Meinke, D.W.; Ray, A. DICER-LIKE1: Blind men and elephants in Arabidopsis development. Trends Plant Sci. 2002, 7, 487–491. [Google Scholar] [CrossRef]
- Llave, C. Virus-derived small interfering RNAs at the core of plant–virus interactions. Trends Plant Sci. 2010, 15, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Pantaleo, V.; Szittya, G.; Burgyan, J. Molecular bases of viral RNA targeting by viral small interfering RNA-programmed RISC. J. Virol. 2007, 81, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, B.; Sanfacon, H. Temperature-dependent symptom recovery in Nicotiana benthamiana plants infected with tomato ringspot virus is associated with reduced translation of viral RNA2 and requires ARGONAUTE 1. Virology 2014, 456–457, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; Zamora, A.; Azhar, M.T.; Sacco, M.A.; Lambert, L.H.; Moffett, P. Virus resistance induced by NB-LRR proteins involves Argonaute4-dependent translational control. Plant J. 2009, 58, 940–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-dependent RNA polymerases and dicer-like proteins in antiviral defense and small interfering RNA biogenesis during turnip mosaic virus infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Kørner, C.J.; Pitzalis, N.; Peña, E.J.; Erhardt, M.; Vazquez, F.; Heinlein, M. Crosstalk between PTGS and TGS pathways in natural antiviral immunity and disease recovery. Nat. Plants 2018, 4, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, F.; Harrison, B.D.; Baulcombe, D.C. A similarity between viral defense and gene silencing in plants. Science 1997, 276, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Ding, S.W. Viral suppressors of RNA silencing. Curr. Opin. Biotechnol. 2001, 12, 150–154. [Google Scholar] [CrossRef]
- Jaubert, M.; Bhattacharjee, S.; Mello, A.F.S.; Perry, K.L.; Moffett, P. ARGONAUTE2 mediates RNA-silencing antiviral defenses against potato virus X in Arabidopsis. Plant Physiol. 2011, 156, 1556–1564. [Google Scholar] [CrossRef] [PubMed]
- Andika, I.B.; Maruyama, K.; Sun, L.; Kondo, H.; Tamada, T.; Suzuki, N. Differential contributions of plant Dicer-like proteins to antiviral defences against potato virus X in leaves and roots. Plant J. 2015, 81, 781–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosseau, C.; Moffett, P. Functional and genetic analysis identify a role for Arabidopsis ARGONAUTE5 in antiviral RNA silencing. Plant Cell 2015, 27, 1742–1754. [Google Scholar] [CrossRef] [PubMed]
- Molnar, A.; Melnyk, C.W.; Bassett, A.; Hardcastle, T.J.; Dunn, R.; Baulcombe, D.C. Small silencing RNAs in plants are mobile and direct epigenetic modification in recipient cells. Science 2010, 328, 872–875. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, K.S.; Masuta, C. Interaction between viral RNA silencing suppressors and host factors in plant immunity. Curr. Opin. Plant Biol. 2014, 20, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, Y.; Kanyuka, K.; Parry, M.A.; Powers, S.J.; Halford, N.G. GCN2-dependent phosphorylation of eukaryotic translation initiation factor-2alpha in Arabidopsis. J. Exp. Bot. 2008, 59, 3131–3141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dickinson, J.R.; Paul, M.J.; Halford, N.G. Molecular cloning of an arabidopsis homologue of GCN2, a protein kinase involved in co-ordinated response to amino acid starvation. Planta 2003, 217, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Browning, K.S. The plant translational apparatus. Plant Mol. Biol. 1996, 32, 107–144. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, Y.; Kulasekaran, S.; Benito, P.; López, B.; Marcos, R.; Cascón, T.; Hamberg, M.; Castresana, C. Arabidopsis nonresponding to oxylipins locus NOXY7 encodes a yeast GCN1 homolog that mediates noncanonical translation regulation and stress adaptation. Plant Cell Environ. 2018, 41, 1438–1452. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, H.; Zhao, C.; Li, S.; Kong, L.; Wu, W.; Kong, W.; Liu, Y.; Wei, Y.; Zhu, J.K.; et al. The inhibition of protein translation mediated by AtGCN1 is essential for cold tolerance in Arabidopsis thaliana. Plant Cell Environ. 2017, 40, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Lageix, S.; Lanet, E.; Pouch-Pélissier, M.-N.; Espagnol, M.-C.; Robaglia, C.; Deragon, J.-M.; Pélissier, T. ArabidopsiseIF2α kinase GCN2 is essential for growth in stress conditions and is activated by wounding. BMC Plant Biol. 2008, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Echevarria-Zomeno, S.; Yanguez, E.; Fernandez-Bautista, N.; Castro-Sanz, A.B.; Ferrando, A.; Castellano, M.M. Regulation of translation initiation under biotic and abiotic stresses. Int. J. Mol. Sci. 2013, 14, 4670–4683. [Google Scholar] [CrossRef] [PubMed]
- Truniger, V.; Aranda, M.A. Recessive resistance to plant viruses. In Advances in Virus Research; Gad, L., John, P.C., Eds.; Academic Press: New York, NY, USA, 2009; Volume 75, pp. 119–231. [Google Scholar]
- Endo, Y.; Tsurugi, K.; Lambert, J.M. The site of action of six different ribosome-inactivating proteins from plants on eukaryotic ribosomes: The RNA N-glycosidase activity of the proteins. Biochem. Biophys. Res. Commun. 1988, 150, 1032–1036. [Google Scholar] [CrossRef]
- Stirpe, F. Ribosome-inactivating proteins: From toxins to useful proteins. Toxicon 2013, 67, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Domashevskiy, A.V.; Williams, S.; Kluge, C.; Cheng, S.-Y. Plant translation initiation complex eIFiso4F directs pokeweed antiviral protein to selectively depurinate uncapped tobacco etch virus RNA. Biochemistry 2017, 56, 5980–5990. [Google Scholar] [CrossRef] [PubMed]
- Picard, D.; Kao, C.C.; Hudak, K.A. Pokeweed antiviral protein inhibits brome mosaic virus replication in plant cells. J. Biol. Chem. 2005, 280, 20069–20075. [Google Scholar] [CrossRef] [PubMed]
- Tumer, N.E.; Hwang, D.J.; Bonness, M. C-terminal deletion mutant of pokeweed antiviral protein inhibits viral infection but does not depurinate host ribosomes. Proc. Natl. Acad. Sci. USA 1997, 94, 3866–3871. [Google Scholar] [CrossRef] [PubMed]
- Lodge, J.K.; Kaniewski, W.K.; Tumer, N.E. Broad-spectrum virus resistance in transgenic plants expressing pokeweed antiviral protein. Proc. Natl. Acad. Sci. USA 1993, 90, 7089–7093. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.C.; White, R.F.; Antoniw, J.F.; Lin, Q. Effect of pokeweed antiviral protein (PAP) on the infection of plant viruses. Plant Pathol. 1991, 40, 612–620. [Google Scholar] [CrossRef]
- Bijal, A.P.; Nilgun, E.T. Antiviral activity of ribosome inactivating proteins in medicine. Mini Rev. Med. Chem. 2004, 4, 523–543. [Google Scholar]
- Stirpe, F.; Gilabert-Oriol, R. Ribosome-inactivating proteins: An overview. In Plant Toxins; Carlini, C.R., Ligabue-Braun, R., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 153–182. [Google Scholar]
- Görschen, E.; Dunaeva, M.; Hause, B.; Reeh, I.; Wasternack, C.; Parthier, B. Expression of the ribosome-inactivating protein JIP60 from barley in transgenic tobacco leads to an abnormal phenotype and alterations on the level of translation. Planta 1997, 202, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Zorzatto, C.; Machado, J.P.B.; Lopes, K.V.G.; Nascimento, K.J.T.; Pereira, W.A.; Brustolini, O.J.B.; Reis, P.A.B.; Calil, I.P.; Deguchi, M.; Sachetto-Martins, G.; et al. NIK1-mediated translation suppression functions as a plant antiviral immunity mechanism. Nature 2015, 520, 679–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariano, A.C.; Andrade, M.O.; Santos, A.A.; Carolino, S.M.; Oliveira, M.L.; Baracat-Pereira, M.C.; Brommonshenkel, S.H.; Fontes, E.P. Identification of a novel receptor-like protein kinase that interacts with a geminivirus nuclear shuttle protein. Virology 2004, 318, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontes, E.P.B.; Santos, A.A.; Luz, D.F.; Waclawovsky, A.J.; Chory, J. The geminivirus nuclear shuttle protein is a virulence factor that suppresses transmembrane receptor kinase activity. Genes Dev. 2004, 18, 2545–2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.S.; Choi, J.H.; Heinz, J.; Chetty, C.S. Domain-specific positive selection contributes to the evolution of Arabidopsis leucine-rich repeat receptor-like kinase (LRR RLK) genes. J. Mol. Evol. 2006, 63, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.M.; Santos, A.A.; Pires, S.R.; Rocha, C.S.; Saraiva, D.I.; Machado, J.P.B.; Mattos, E.C.; Fietto, L.G.; Fontes, E.P.B. Regulated nuclear trafficking of rpL10A mediated by NIK1 represents a defense strategy of plant cells against virus. PLoS Pathog. 2008, 4, e1000247. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, P.; Sakvarelidze-Achard, L.; Bruun-Rasmussen, M.; Dunoyer, P.; Yamamoto, Y.Y.; Sieburth, L.; Voinnet, O. Widespread translational inhibition by plant miRNAs and siRNAs. Science 2008, 320, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Nicole, M.C.; Meteignier, L.V.; Hong, N.; Wang, G.; Moffett, P. Different roles for RNA silencing and RNA processing components in virus recovery and virus-induced gene silencing in plants. J. Exp. Bot. 2015, 66, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Sanchez, M.A.; Liu, J.; Hannon, G.J.; Parker, R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev. 2006, 20, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Xuan, Y.; Han, Y.; Ding, X.; Ye, K.; Yang, F.; Gao, P.; Goff, S.P.; Gao, G. Regulation of HIV-1 Gag-Pol expression by shiftless, an inhibitor of programmed -1 ribosomal frameshifting. Cell 2019, 176, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Peumans, W.J.; Van Damme, E.J. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Tanaka, K.; Cao, Y.; Qi, Y.; Qiu, J.; Liang, Y.; Lee, S.Y.; Stacey, G. Identification of a plant receptor for extracellular ATP. Science 2014, 343, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Catanzariti, A.M.; Lim, G.T.; Jones, D.A. The tomato I-3 gene: A novel gene for resistance to Fusarium wilt disease. New Phytol. 2015, 207, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Ranf, S.; Gisch, N.; Schaffer, M.; Illig, T.; Westphal, L.; Knirel, Y.A.; Sanchez-Carballo, P.M.; Zahringer, U.; Huckelhoven, R.; Lee, J.; et al. A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat. Immunol. 2015, 16, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Mahajan, S.K.; Whitham, S.A.; Yamamoto, M.L.; Carrington, J.C. Cloning of the Arabidopsis RTM1 gene, which controls restriction of long-distance movement of tobacco etch virus. Proc. Natl. Acad. Sci. USA 2000, 97, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decroocq, V.; Sicard, O.; Alamillo, J.M.; Lansac, M.; Eyquard, J.P.; García, J.A.; Candresse, T.; Le Gall, O.; Revers, F. Multiple resistance traits control plum pox virus infection in Arabidopsis thaliana. Mol. Plant Microbe Interact. 2006, 19, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Whitham, S.A.; Anderberg, R.J.; Chisholm, S.T.; Carrington, J.C. Arabidopsis RTM2 gene is necessary for specific restriction of tobacco etch virus and encodes an unusual small heat shock-like protein. Plant Cell 2000, 12, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Parra, M.A.; Anderberg, R.J.; Carrington, J.C. Arabidopsis RTM1 and RTM2 genes function in phloem to restrict long-distance movement of tobacco etch virus. Plant Physiol. 2001, 127, 1667–1675. [Google Scholar] [CrossRef] [PubMed]
- Cosson, P.; Sofer, L.; Le, Q.H.; Leger, V.; Schurdi-Levraud, V.; Whitham, S.A.; Yamamoto, M.L.; Gopalan, S.; Le Gall, O.; Candresse, T.; et al. RTM3, which controls long-distance movement of potyviruses, is a member of a new plant gene family encoding a meprin and TRAF homology domain-containing protein. Plant Physiol. 2010, 154, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Cosson, P.; Schurdi-Levraud, V.; Le, Q.H.; Sicard, O.; Caballero, M.; Roux, F.; Le Gall, O.; Candresse, T.; Revers, F. The RTM resistance to potyviruses in Arabidopsis thaliana: Natural variation of the RTM genes and evidence for the implication of additional genes. PLoS ONE 2012, 7, e39169. [Google Scholar] [CrossRef] [PubMed]
- Sofer, L.; Cabanillas, D.G.; Gayral, M.; Téplier, R.; Pouzoulet, J.; Ducousso, M.; Dufin, L.; Bréhélin, C.; Ziegler-Graff, V.; Brault, V.; et al. Identification of host factors potentially involved in RTM-mediated resistance during potyvirus long distance movement. Arch. Virol. 2017, 162, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Decroocq, V.; Salvador, B.; Sicard, O.; Glasa, M.; Cosson, P.; Svanella-Dumas, L.; Revers, F.; García, J.A.; Candresse, T. The determinant of potyvirus ability to overcome the RTM resistance of Arabidopsis thaliana maps to the N-terminal region of the coat protein. Mol. Plant Microbe Interact. 2009, 22, 1302–1311. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, Y.; Maejima, K.; Komatsu, K.; Shiraishi, T.; Okano, Y.; Himeno, M.; Sugawara, K.; Neriya, Y.; Minato, N.; Miura, C.; et al. Lectin-mediated resistance impairs plant virus infection at the cellular level. Plant Cell 2012, 24, 778–793. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Shiraishi, T.; Yoshida, T.; Fujita, N.; Netsu, O.; Yamaji, Y.; Namba, S. A replicase of potato virus X acts as the resistance-breaking determinant for JAX1-mediated resistance. Mol. Plant Microbe Interact. 2013, 26, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Shiraishi, T.; Hagiwara-Komoda, Y.; Komatsu, K.; Maejima, K.; Okano, Y.; Fujimoto, Y.; Yusa, A.; Yamaji, Y.; Namba, S. The plant non-canonical antiviral resistance protein JAX1 inhibits potexviral replication by targeting the viral RNA-dependent RNA polymerase. J. Virol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Prasad, V.; Mishra, S.K.; Srivastava, S.; Srivastava, A. A virus inhibitory protein isolated from Cyamopsis tetragonoloba (L.) Taub. upon induction of systemic antiviral resistance shares partial amino acid sequence homology with a lectin. Plant Cell Rep. 2014, 33, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Li, H.; Zhang, W. The lectin from Musa paradisiaca binds with the capsid protein of tobacco mosaic virus and prevents viral infection. Biotechnol. Biotechnol. Equip. 2014, 28, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Masuda, K.; Naito, S.; Meshi, T.; Ishikawa, M. An inhibitor of viral RNA replication is encoded by a plant resistance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 13833–13838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Yang, Q.; Ye, J.; Lai, J.; Wu, J.; et al. An atypical thioredoxin imparts early resistance to sugarcane mosaic virus in maize. Mol. Plant 2017, 10, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yan, Z.; Xia, Z.; Cheng, Y.; Jiao, Z.; Sun, B.; Zhou, T.; Fan, Z. A violaxanthin de-epoxidase interacts with a viral suppressor of RNA silencing to inhibit virus amplification. Plant Physiol. 2017, 175, 1774–1794. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, Y.; He, J.; Zheng, X.; Hu, J.; Liu, Y.; Dai, H.; Zhang, Y.; Wang, B.; Wu, W.; et al. STV11 encodes a sulphotransferase and confers durable resistance to rice stripe virus. Nat. Commun. 2014, 5, 4768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlaan, M.G.; Hutton, S.F.; Ibrahem, R.M.; Kormelink, R.; Visser, R.G.F.; Scott, J.W.; Edwards, J.D.; Bai, Y. The tomato yellow leaf curl virus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD-class RNA–dependent RNA polymerases. PLoS Genet. 2013, 9, e1003399. [Google Scholar] [CrossRef] [PubMed]
- Caro, M.; Verlaan, M.G.; Julian, O.; Finkers, R.; Wolters, A.A.; Hutton, S.F.; Scott, J.W.; Kormelink, R.; Visser, R.G.F.; Diez, M.J.; et al. Assessing the genetic variation of Ty-1 and Ty-3 alleles conferring resistance to tomato yellow leaf curl virus in a broad tomato germplasm. Mol. Breed. 2015, 35, 132. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Mawatari, N.; Miyashita, S.; Kishino, H.; Meshi, T.; Ishikawa, M. Coevolution and hierarchical interactions of tomato mosaic virus and the resistance gene Tm-1. PLoS Pathog. 2012, 8, e1002975. [Google Scholar] [CrossRef] [PubMed]
- Galan, J.M.; Moreau, V.; Andre, B.; Volland, C.; Haguenauer-Tsapis, R. Ubiquitination mediated by the Npi1p/Rsp5p ubiquitin-protein ligase is required for endocytosis of the yeast uracil permease. J. Biol. Chem. 1996, 271, 10946–10952. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Li, Z.; Nagy, P.D. The Nedd4-type Rsp5p ubiquitin ligase inhibits tombusvirus replication by regulating degradation of the p92 replication protein and decreasing the activity of the tombusvirus replicase. J. Virol. 2009, 83, 11751–11764. [Google Scholar] [CrossRef] [PubMed]
- Kerscher, O.; Felberbaum, R.; Hochstrasser, M. Modification of proteins by ubiquitin and ubiquitin-like proteins. Annu. Rev. Cell Dev. Biol. 2006, 22, 159–180. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Mou, Z.; Tada, Y.; Spivey, N.W.; Genschik, P.; Dong, X. Proteasome-mediated turnover of the transcription coactivator NPR1 plays dual roles in regulating plant immunity. Cell 2009, 137, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Cao, Y.; Sun, X.; Espinoza, C.; Nguyen, C.T.; Liang, Y.; Stacey, G. Arabidopsis E3 ubiquitin ligase PLANT U-BOX13 (PUB13) regulates chitin receptor LYSIN MOTIF RECEPTOR KINASE5 (LYK5) protein abundance. New Phytol. 2017, 214, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Verchot, J. The ER quality control and ER associated degradation machineries are vital for viral pathogenesis. Front. Plant Sci. 2014, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Vierstra, R.D. The ubiquitin–26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 2009, 10, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Alcaide-Loridan, C.; Jupin, I. Ubiquitin and plant viruses, let’s play together! Plant Physiol. 2012, 160, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Tao, X.; Xie, Y.; Fauquet, C.M.; Zhou, X. A DNAβ associated with tomato yellow leaf curl China virus is required for symptom induction. J. Virol. 2004, 78, 13966–13974. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, X.; Wu, J.; Briddon, R.W.; Zhou, X. βC1 encoded by tomato yellow leaf curl China betasatellite forms multimeric complexes in vitro and in vivo. Virology 2011, 409, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Li, G.; Wang, D.; Hu, D.; Zhou, X. A begomovirus DNAβ-encoded protein binds DNA, functions as a suppressor of RNA silencing, and targets the cell nucleus. J. Virol. 2005, 79, 10764–10775. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Hu, T.; Bao, M.; Cao, L.; Zhang, H.; Song, F.; Xie, Q.; Zhou, X. Tobacco RING E3 ligase NtRFP1 mediates ubiquitination and proteasomal degradation of a geminivirus-encoded betaC1. Mol. Plant 2016, 9, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Camborde, L.; Planchais, S.; Tournier, V.; Jakubiec, A.; Drugeon, G.; Lacassagne, E.; Pflieger, S.; Chenon, M.; Jupin, I. The ubiquitin-proteasome system regulates the accumulation of turnip yellow mosaic virus RNA-dependent RNA polymerase during viral infection. Plant Cell 2010, 22, 3142–3152. [Google Scholar] [CrossRef] [PubMed]
- Jakubiec, A.; Drugeon, G.; Camborde, L.; Jupin, I. Proteolytic processing of turnip yellow mosaic virus replication proteins and functional impact on infectivity. J. Virol. 2007, 81, 11402–11412. [Google Scholar] [CrossRef] [PubMed]
- Reichel, C.; Beachy, R.N. Degradation of tobacco mosaicvirus movement protein by the 26S proteasome. J. Virol. 2000, 74, 3330–3337. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Amari, K.; Gereige, D.; Brandner, K.; Mély, Y.; Heinlein, M. Control of tobacco mosaic virus movement protein fate by CELL-DIVISION-CYCLE protein 48. Plant Physiol. 2012, 160, 2093–2108. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Bassham, D.C. New insight into the mechanism and function of autophagy in plant cells. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, UK, 2015; Volume 320, pp. 1–40. [Google Scholar]
- Liu, Y.; Schiff, M.; Czymmek, K.; Tallóczy, Z.; Levine, B.; Dinesh-Kumar, S.P. Autophagy regulates programmed cell death during the plant innate immune response. Cell 2005, 121, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Yu, B.; Wang, Y.; Liu, Y. Role of plant autophagy in stress response. Protein Cell 2011, 2, 784–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, K.; Jikumaru, Y.; Kamiya, Y.; Kusano, M.; Consonni, C.; Panstruga, R.; Ohsumi, Y.; Shirasu, K. Autophagy negatively regulates cell death by controlling NPR1-dependent salicylic acid signaling during senescence and the innate immune response in Arabidopsis. Plant Cell 2009, 21, 2914–2927. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, C.; Li, Y.; Wu, G.; Hou, X.; Zhou, X.; Wang, A. Beclin1 restricts RNA virus infection in plants through suppression and degradation of the viral polymerase. Nat. Commun. 2018, 9, 1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haxim, Y.; Ismayil, A.; Jia, Q.; Wang, Y.; Zheng, X.; Chen, T.; Qian, L.; Liu, N.; Wang, Y.; Han, S.; et al. Autophagy functions as an antiviral mechanism against geminiviruses in plants. eLife 2017, 6, e23897. [Google Scholar] [CrossRef] [PubMed]
- Hafrén, A.; Macia, J.L.; Love, A.J.; Milner, J.J.; Drucker, M.; Hofius, D. Selective autophagy limits cauliflower mosaic virus infection by NBR1-mediated targeting of viral capsid protein and particles. Proc. Natl. Acad. Sci. USA 2017, 114, E2026–E2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafrén, A.; Üstün, S.; Hochmuth, A.; Svenning, S.; Johansen, T.; Hofius, D. Turnip mosaic virus counteracts selective autophagy of the viral silencing suppressor HCpro. Plant Physiol. 2018, 176, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Hafrén, A.; Lõhmus, A.; Mäkinen, K. Formation of potato virus A-induced RNA granules and viral translation are interrelated processes required for optimal virus accumulation. PLoS Pathog. 2015, 11, e1005314. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Jeong, D.H.; De Paoli, E.; Park, S.; Rosen, B.D.; Li, Y.; González, A.J.; Yan, Z.; Kitto, S.L.; Grusak, M.A.; et al. MicroRNAs as master regulators of the plant NB-LRR defense gene family via the production of phased, trans-acting siRNAs. Genes Dev. 2011, 25, 2540–2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccara, M.; Sarazin, A.; Thiébeauld, O.; Jay, F.; Voinnet, O.; Navarro, L.; Colot, V. The Arabidopsis miR472-RDR6 silencing pathway modulates PAMP- and effector-triggered immunity through the post-transcriptional control of disease resistance genes. PLoS Pathog. 2014, 10, e1003883. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, J.; Tung, J.; Liu, D.; Zhou, Y.; He, S.; Du, Y.; Baker, B.; Li, F. A role for small RNA in regulating innate immunity during plant growth. PLoS Pathog. 2018, 14, e1006756. [Google Scholar] [CrossRef] [PubMed]
- Hatsugai, N.; Kuroyanagi, M.; Yamada, K.; Meshi, T.; Tsuda, S.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. A plant vacuolar protease, VPE, mediates virus-induced hypersensitive cell death. Science 2004, 305, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Caplan, J.L.; Zhu, X.; Mamillapalli, P.; Marathe, R.; Anandalakshmi, R.; Dinesh-Kumar, S.P. Induced ER chaperones regulate a receptor-like kinase to mediate antiviral innate immune response in plants. Cell Host Microbe 2009, 6, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Weaver, N.D.; Kesarwani, M.; Dong, X. Induction of protein secretory pathway is required for systemic acquired resistance. Science 2005, 308, 1036–1040. [Google Scholar] [CrossRef] [PubMed]
- Križnik, M.; Petek, M.; Dobnik, D.; Ramšak, Ž.; Baebler, Š.; Pollmann, S.; Kreuze, J.F.; Žel, J.; Gruden, K. Salicylic acid perturbs sRNA-gibberellin regulatory network in immune response of potato to potato virus Y infection. Front. Plant Sci. 2017, 8, 2192. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, K.S.; Masuta, C.; Yamada, S.; Shimura, H.; Kashihara, Y.; Wada, T.S.; Meguro, A.; Goto, K.; Tadamura, K.; Sueda, K.; et al. Tobacco calmodulin-like protein provides secondary defense by binding to and directing degradation of virus RNA silencing suppressors. Proc. Natl. Acad. Sci. USA 2012, 109, 10113–10118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, E.J.; Tadamura, K.; Murakami, T.; Inaba, J.-I.; Kim, B.M.; Sato, M.; Atsumi, G.; Kuchitsu, K.; Masuta, C.; Nakahara, K.S. rgs-CaM detects and counteracts viral RNA silencing suppressors in plant immune priming. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Anandalakshmi, R.; Marathe, R.; Ge, X.; Herr, J.M., Jr.; Mau, C.; Mallory, A.; Pruss, G.; Bowman, L.; Vance, V.B. A calmodulin-related protein that suppresses posttranscriptional gene silencing in plants. Science 2000, 290, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, D.A.; Holland, J.J. Rapid evolution of RNA viruses. Annu. Rev. Microbiol. 1987, 41, 409–431. [Google Scholar] [CrossRef] [PubMed]
- Pita, J.S.; Roossinck, M.J. Virus populations, mutation rates and frequencies. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 109–121. [Google Scholar]
- Ge, L.; Zhang, J.; Zhou, X.; Li, H. Genetic structure and population variability of tomato yellow leaf curl China virus. J. Virol. 2007, 81, 5902–5907. [Google Scholar] [CrossRef] [PubMed]
- Grigoras, I.; Timchenko, T.; Grande-Pérez, A.; Katul, L.; Vetten, H.-J.; Gronenborn, B. High variability and rapid evolution of a nanovirus. J. Virol. 2010, 84, 9105–9117. [Google Scholar] [CrossRef] [PubMed]
- Pinel-Galzi, A.; Dubreuil-Tranchant, C.; Hebrard, E.; Mariac, C.; Ghesquiere, A.; Albar, L. Mutations in rice yellow mottle virus polyprotein P2a involved in RYMV2 gene resistance breakdown. Front. Plant Sci. 2016, 7, 1779. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Leal, R.; Bryan, B.K.; Smith, J.T.; Rush, C.M. Breakdown of host resistance by independent evolutionary lineages of beet necrotic yellow vein virus involves a parallel c/u mutation in its p25 gene. Phytopathology 2010, 100, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Naito, S.; Meshi, T.; Ishikawa, M. An inhibitory interaction between viral and cellular proteins underlies the resistance of tomato to nonadapted tobamoviruses. Proc. Natl. Acad. Sci. USA 2009, 106, 8778–8783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Nicaise, V.; Candresse, T. Plum pox virus capsid protein suppresses plant pathogen-associated molecular pattern (PAMP)-triggered immunity. Mol. Plant Pathol. 2016, 18, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Wei, M.; Li, G.; Lei, R.; Qiu, Y.; Wang, C.; Li, Z.-H.; Zhu, S. The cucumber mosaic virus movement protein suppresses PAMP-triggered immune responses in Arabidopsis and tobacco. Biochem. Biophys. Res. Commun. 2018, 498, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, A.S.; Golyaev, V.; Turco, S.; Gubaeva, E.G.; Rajeswaran, R.; Schepetilnikov, M.V.; Srour, O.; Ryabova, L.A.; Boller, T.; Pooggin, M.M. Viral protein suppresses oxidative burst and salicylic acid-dependent autophagy and facilitates bacterial growth on virus-infected plants. New Phytol. 2016, 211, 1020–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Xiong, R.; Li, Y.; Li, F.; Zhou, X.; Wang, A. Sumoylation of turnip mosaic virus RNA polymerase promotes viral infection by counteracting the host NPR1-mediated immune response. Plant Cell 2017, 29, 508–525. [Google Scholar] [CrossRef] [PubMed]
- van den Burg, H.A.; Kini, R.K.; Schuurink, R.C.; Takken, F.L.W. Arabidopsis small ubiquitin-like modifier paralogs have distinct functions in development and defense. Plant Cell 2010, 22, 1998–2016. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Withers, J.; Mohan, R.; Marqués, J.; Gu, Y.; Yan, S.; Zavaliev, R.; Nomoto, M.; Tada, Y.; Dong, X. Posttranslational modifications of the master transcriptional regulator NPR1 enable dynamic but tight control of plant immune responses. Cell Host Microbe 2015, 18, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Cao, X.; Wang, X.; Jiang, J.; Wan, J.; Laliberté, J.F.; Zhang, Y. Three-dimensional architecture and biogenesis of membrane structures associated with plant virus replication. Front. Plant Sci. 2018, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Harak, C.; Lohmann, V. Ultrastructure of the replication sites of positive-strand RNA viruses. Virology 2015, 479–480, 418–433. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Nagy, P.D. Expanding use of multi-origin subcellular membranes by positive-strand RNA viruses during replication. Curr. Opin. Virol. 2014, 9, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Laliberté, J.F.; Zheng, H. Viral manipulation of plant host membranes. Annu. Rev. Virol. 2014, 1, 237–259. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.; McCartney, A.; Gidda, S.; Mullen, R. Localization of the carnation Italian ringspot virus replication protein p36 to the mitochondrial outer membrane is mediated by an internal targeting signal and the TOM complex. BMC Cell Biol. 2008, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Jiang, Y.; Nagy, P.D. A unique role for the host ESCRT proteins in replication of tomato bushy stunt virus. PLoS Pathog. 2009, 5, e1000705. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Hartwig, M.A.; Ahlquist, P. Brome mosaic virus helicase- and polymerase-like proteins colocalize on the endoplasmic reticulum at sites of viral RNA synthesis. J. Virol. 1996, 70, 8908–8916. [Google Scholar] [PubMed]
- Cotton, S.; Grangeon, R.; Thivierge, K.; Mathieu, I.; Ide, C.; Wei, T.; Wang, A.; Laliberté, J.F. Turnip mosaic virus RNA replication complex vesicles are mobile, align with microfilaments, and are each derived from a single viral genome. J. Virol. 2009, 83, 10460–10471. [Google Scholar] [CrossRef] [PubMed]
- Más, P.; Beachy, R.N. Replication of tobacco mosaic virus on endoplasmic reticulum and role of the cytoskeleton and virus movement protein in intracellular distribution of viral RNA. J. Cell Biol. 1999, 147, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Howell, S.H. The unfolded protein response supports plant development and defense as well as responses to abiotic stress. Front. Plant Sci. 2017, 8, 344. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Dickman, M.B.; Whitham, S.A.; Payton, M.; Verchot, J. The unfolded protein response is triggered by a plant viral movement protein. Plant Physiol. 2011, 156, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.M.; Chen, S.; Payton, M.; Dickman, M.B.; Verchot, J. TGBp3 triggers the unfolded protein response and SKP1-dependent programmed cell death. Mol. Plant Pathol. 2013, 14, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, H.; Brandizzi, F.; Verchot, J.; Wang, A. The UPR branch IRE1-bZIP60 in plants plays an essential role in viral infection and is complementary to the only UPR pathway in yeast. PLoS Genet. 2015, 11, e1005164. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Yang, D.; Xie, L.; Sun, L.; Zhang, S.; Zhu, Q.; Li, J.; Wang, X.; Chen, J. Rice black-streaked dwarf virus P10 induces membranous structures at the ER and elicits the unfolded protein response in Nicotiana benthamiana. Virology 2013, 447, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, H.; Shine, M.B.; Cui, X.; Chen, X.; Ma, N.; Kachroo, P.; Zhi, H.; Kachroo, A. The potyviral P3 protein targets eukaryotic elongation factor 1A to promote the unfolded protein response and viral pathogenesis. Plant Physiol. 2016, 172, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pendon, J.A.; Li, F.; Li, W.X.; Ding, S.W. Suppression of antiviral silencing by cucumber mosaic virus 2b protein in Arabidopsis is associated with drastically reduced accumulation of three classes of viral small interfering RNAs. Plant Cell 2007, 19, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Hong, J.-S.; Tabayashi, N.; Ito, A.; Masuta, C.; Matsumura, T. Development of cucumber mosaic virus as a vector modifiable for different host species to produce therapeutic proteins. Planta 2007, 225, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, X.; Ding, S.W. Viral suppressors of RNA-based viral immunity: Host targets. Cell Host Microbe 2010, 8, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Burgyán, J.; Havelda, Z. Viral suppressors of RNA silencing. Trends Plant Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Valli, A.A.; Gallo, A.; Rodamilans, B.; López-Moya, J.J.; García, J.A. The HcPro from the Potyviridae family: An enviable multitasking helper component that every virus would like to have. Mol. Plant Pathol. 2018, 19, 744–763. [Google Scholar] [CrossRef] [PubMed]
- Várallyay, É.; Válóczi, A.; Ágyi, Á.; Burgyán, J.; Havelda, Z. Plant virus-mediated induction of miR168 is associated with repression of ARGONAUTE1 accumulation. EMBO J. 2010, 29, 3507–3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zhang, L.; Yu, K.; Wang, A. Pathogenesis of soybean mosaic virus in soybean carrying Rsv1 gene is associated with miRNA and siRNA pathways, and breakdown of AGO1 homeostasis. Virology 2015, 476, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, T.; Shen, D.; Wang, J.; Ling, X.; Hu, Z.; Chen, T.; Hu, J.; Huang, J.; Yu, W.; et al. Tomato yellow leaf curl virus intergenic siRNAs target a host long noncoding RNA to modulate disease symptoms. PLoS Pathog. 2019, 15, e1007534. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Lin, L.; Wang, S.; Guo, Q.; Zhou, H.; Rong, L.; Li, J.; Peng, J.; Lu, Y.; Zheng, H.; et al. Identification and regulation of host genes related to rice stripe virus symptom production. New Phytol. 2016, 209, 1106–1119. [Google Scholar] [CrossRef] [PubMed]
- Moyo, L.; Ramesh, S.V.; Kappagantu, M.; Mitter, N.; Sathuvalli, V.; Pappu, H.R. The effects of potato virus Y-derived virus small interfering RNAs of three biologically distinct strains on potato (Solanum tuberosum) transcriptome. Virol. J. 2017, 14, 129. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shibuya, M.; Taneda, A.; Kurauchi, T.; Senda, M.; Owens, R.A.; Sano, T. Accumulation of potato spindle tuber viroid-specific small RNAs is accompanied by specific changes in gene expression in two tomato cultivars. Virology 2011, 413, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhou, G. Characteristics of siRNAs derived from southern rice black-streaked dwarf virus in infected rice and their potential role in host gene regulation. Virol. J. 2017, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, Y.; Ding, B.; Fei, Z. Comprehensive transcriptome analyses reveal that potato spindle tuber viroid triggers genome-wide changes in alternative splicing, inducible trans-acting activity of phased secondary small interfering RNAs, and immune responses. J. Virol. 2017, 91, e00247-17. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, K.K.; Scholthof, K.-B.G. Genome-wide analysis of alternative splicing landscapes modulated during plant-virus interactions in Brachypodium distachyon. Plant Cell 2015, 27, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yang, M.; Lu, L.; Zhang, X. Diverse functions of small RNAs in different plant-pathogen communications. Front. Microbiol. 2016, 7, 1552. [Google Scholar] [CrossRef] [PubMed]
- He, B. Viruses, endoplasmic reticulum stress, and interferon responses. Cell Death Differ. 2006, 13, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toribio, R.; Ventoso, I. Inhibition of host translation by virus infection in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 9837–9842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miras, M.; Miller, W.A.; Truniger, V.; Aranda, M.A. Non-canonical translation in plant RNA viruses. Front. Plant Sci. 2017, 8, 494. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.A.; Lopes, K.V.G.; Apfata, J.A.C.; Fontes, E.P.B. NSP-interacting kinase, NIK: A transducer of plant defence signalling. J. Exp. Bot. 2010, 61, 3839–3845. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.P.B.; Brustolini, O.J.B.; Mendes, G.C.; Santos, A.A.; Fontes, E.P.B. NIK1, a host factor specialized in antiviral defense or a novel general regulator of plant immunity? BioEssays 2015, 37, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Chenon, M.; Camborde, L.; Cheminant, S.; Jupin, I. A viral deubiquitylating enzyme targets viral RNA-dependent RNA polymerase and affects viral infectivity. EMBO J. 2012, 31, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Liu, N.; Xie, K.; Dai, Y.; Han, S.; Zhao, X.; Qian, L.; Wang, Y.; Zhao, J.; Gorovits, R.; et al. CLCuMuB βC1 subverts ubiquitination by interacting with NbSKP1s to enhance geminivirus infection in Nicotiana benthamiana. PLoS Pathog. 2016, 12, e1005668. [Google Scholar] [CrossRef] [PubMed]
- Chiu, M.H.; Chen, I.H.; Baulcombe, D.C.; Tsai, C.H. The silencing suppressor P25 of potato virus X interacts with Argonaute1 and mediates its degradation through the proteasome pathway. Mol. Plant Pathol. 2010, 11, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, A.F.; Correa, R.L.; Nakasugi, K.; Jackson, C.; Kawchuk, L.; Vaslin, M.F.S.; Waterhouse, P.M. The enamovirus P0 protein is a silencing suppressor which inhibits local and systemic RNA silencing through AGO1 degradation. Virology 2012, 426, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Lozsa, R.; Hutvagner, G.; Burgyan, J. Polerovirus protein P0 prevents the assembly of small RNA-containing RISC complexes and leads to degradation of ARGONAUTE1. Plant J. 2010, 62, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumberger, N.; Tsai, C.-H.; Lie, M.; Havecker, E.; Baulcombe, D.C. The polerovirus silencing suppressor P0 targets Argonaute proteins for degradation. Curr. Biol. 2007, 17, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Derrien, B.; Baumberger, N.; Schepetilnikov, M.; Viotti, C.; De Cillia, J.; Ziegler-Graff, V.; Isono, E.; Schumacher, K.; Genschik, P. Degradation of the antiviral component ARGONAUTE1 by the autophagy pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 15942–15946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronson, M.N.; Meyer, A.D.; Györgyey, J.; Katul, L.; Vetten, H.J.; Gronenborn, B.; Timchenko, T. Clink, a nanovirus-encoded protein, binds both pRB and SKP1. J. Virol. 2000, 74, 2967–2972. [Google Scholar] [PubMed]
- Aronson, M.N.; Complainville, A.; Clérot, D.; Alcalde, H.; Katul, L.; Vetten, H.J.; Gronenborn, B.; Timchenko, T. In planta protein-protein interactions assessed using a nanovirus-based replication and expression system. Plant J. 2002, 31, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Xu, Y.; Li, C.; Li, Y.; Wu, J.; Zhou, X. Rice stripe virus interferes with S-acylation of remorin and induces its autophagic degradation to facilitate virus infection. Mol. Plant 2018, 11, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Duran, R.; Bejarano, E.R. Geminivirus C2 protein might be the key player for geminiviral co-option of SCF-mediated ubiquitination. Plant Signal Behav. 2011, 6, 999–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Wang, A. The potyviral silencing suppressor protein VPg mediates degradation of SGS3 via ubiquitination and autophagy pathways. J. Virol. 2017, 91, e01478-16. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.B.; Kim, B.-W.; Lee, K.E.; Kim, S.W.; Jeon, H.; Kim, J.; Song, H.K. Insights into noncanonical E1 enzyme activation from the structure of autophagic E1 Atg7 with Atg8. Nat. Struct. Mol. Biol. 2011, 18, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, Y.; Xie, X.; Yue, N.; Li, J.; Wang, X.B.; Han, C.G.; Yu, J.L.; Liu, Y.; Li, D. Barley stripe mosaic virus γb protein subverts autophagy to promote viral infection by disrupting the ATG7-ATG8 interaction. Plant Cell 2018, 30, 1582–1595. [Google Scholar] [CrossRef] [PubMed]
- Wileman, T. Aggresomes and autophagy generate sites for virus replication. Science 2006, 312, 875–878. [Google Scholar] [CrossRef] [PubMed]
- Suhy, D.A.; Giddings, T.H.; Kirkegaard, K. Remodeling the endoplasmic reticulum by poliovirus infection and by individual viral proteins: An autophagy-like origin for virus-induced vesicles. J. Virol. 2000, 74, 8953–8965. [Google Scholar] [CrossRef] [PubMed]
- Dreux, M.; Gastaminza, P.; Wieland, S.F.; Chisari, F.V. The autophagy machinery is required to initiate hepatitis C virus replication. Proc. Natl. Acad. Sci. USA 2009, 106, 14046–14051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Brey, I.; Bartenschlager, R. Membranous replication factories induced by plus-strand RNA viruses. Viruses 2014, 6, 2826. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Luo, H. Interplay between the cellular autophagy machinery and positive-stranded RNA viruses. Acta Biochim. Biophys. Sin. 2012, 44, 375–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boiteux, L.S.; de Noronha Fonseca, M.E.; Vieira, J.V.; de Cássia Pereira-Carvalho, R. Breeding for resistance to viral diseases. In Plant Breeding for Biotic Stress Resistance; Fritsche-Neto, R., Borém, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 57–79. [Google Scholar]
- Rey, C.; Vanderschuren, H. Cassava mosaic and brown streak diseases: Current perspectives and beyond. Annu. Rev. Virol. 2017, 4, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Singh, S.P.; Kianian, S.F. Engineering resistance to plant viruses: Present status and future prospects. In Current Developments in Biotechnology and Bioengineering; Larroche, C., Sanroman, M., Du, G., Pandey, A., Eds.; Elsevier: London, UK, 2017; pp. 75–100. [Google Scholar]
- Galvez, L.C.; Banerjee, J.; Pinar, H.; Mitra, A. Engineered plant virus resistance. Plant Sci. 2014, 228, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.; Zhang, Q.; Yasir, M.; Li, F. Small RNA based genetic engineering for plant viral resistance: Application in crop protection. Front. Microbiol. 2017, 8, 43. [Google Scholar] [PubMed]
- Chen, L.; Cheng, X.; Cai, J.; Zhan, L.; Wu, X.; Liu, Q.; Wu, X. Multiple virus resistance using artificial trans-acting siRNAs. J. Virol. Methods 2016, 228, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, F.; Cai, J.; Chen, W.; Zhao, N.; Sun, Y.; Guo, Y.; Yang, X.; Wu, X. Artificial TALE as a convenient protein platform for engineering broad-spectrum resistance to begomoviruses. Viruses 2015, 7, 4772–4782. [Google Scholar] [PubMed]
- Lin, C.Y.; Ku, H.M.; Tsai, W.S.; Green, S.K.; Jan, F.J. Resistance to a DNA and a RNA virus in transgenic plants by using a single chimeric transgene construct. Transgenic Res. 2011, 20, 261–270. [Google Scholar] [PubMed]
- Chung, B.N.; Palukaitis, P. Resistance to multiple viruses in transgenic tobacco expressing fused, tandem repeat, virus-derived double-stranded RNAs. Virus Genes 2011, 43, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Bucher, E.; Lohuis, D.; van Poppel, P.M.; Geerts-Dimitriadou, C.; Goldbach, R.; Prins, M. Multiple virus resistance at a high frequency using a single transgene construct. J. Gen. Virol. 2006, 87, 3697–3701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, T.; Zhang, L.; Gao, Z.; Zhu, C.X.; Guo, X. Highly efficient virus resistance mediated by artificial microRNAs that target the suppressor of PVX and PVY in plants. Plant Biol. (Stuttg.) 2011, 13, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, M. Pyramiding resistance-conferring gene sequences in crops. Curr. Opin. Virol. 2017, 26, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.-T.; Widyasari, K.; Park, J.Y.; Kim, K.-H. Engineering an auto-activated R protein that is in vivo activated by a viral protease. Virology 2017, 510, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Qi, D.; Ashfield, T.; Helm, M.; Innes, R.W. Using decoys to expand the recognition specificity of a plant disease resistance protein. Science 2016, 351, 684–687. [Google Scholar] [CrossRef] [PubMed]
- Römer, P.; Recht, S.; Lahaye, T. A single plant resistance gene promoter engineered to recognize multiple TAL effectors from disparate pathogens. Proc. Natl. Acad. Sci. USA 2009, 106, 20526–20531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacombe, S.; Rougon-Cardoso, A.; Sherwood, E.; Peeters, N.; Dahlbeck, D.; van Esse, H.P.; Smoker, M.; Rallapalli, G.; Thomma, B.P.H.J.; Staskawicz, B.; et al. Interfamily transfer of a plant pattern-recognition receptor confers broad-spectrum bacterial resistance. Nat. Biotechnol. 2010, 28, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Soininen, M.; Poranen, M.M.; Heinlein, M. Synthetic biology approach for plant protection using dsRNA. Plant Biotechnol. J. 2018, 16, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef] [PubMed]
- García-Cano, E.; Resende, R.O.; Fernández-Muñoz, R.; Moriones, E. Synergistic interaction between tomato chlorosis virus and tomato spotted wilt virus results in breakdown of resistance in tomato. Phytopathology 2006, 96, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic overview of plant antiviral pathways and viral counter-defenses. Pathogen-associated molecular patterns (PAMP)-triggered immunity (PTI), effector-triggered immunity (ETI), RNA silencing, ATYPICAL DOMINANT VIRAL RESISTANCE PROTEINs (ADVR), translation repression, and ubiquitination-mediated and autophagy-mediated protein degradation are indicated by light blue, dark blue, purple, red, pink and green arrows, or lines with bars, respectively. Virus infection circles are indicated by yellow arrows, and viral counter-defenses are indicated by blue lines with bars. Red circles with “P” and “U” letter indicate phosphorylation and ubiquitination, respectively. Unknown or putative paradigms are indicated as“?”.
Figure 1.
Schematic overview of plant antiviral pathways and viral counter-defenses. Pathogen-associated molecular patterns (PAMP)-triggered immunity (PTI), effector-triggered immunity (ETI), RNA silencing, ATYPICAL DOMINANT VIRAL RESISTANCE PROTEINs (ADVR), translation repression, and ubiquitination-mediated and autophagy-mediated protein degradation are indicated by light blue, dark blue, purple, red, pink and green arrows, or lines with bars, respectively. Virus infection circles are indicated by yellow arrows, and viral counter-defenses are indicated by blue lines with bars. Red circles with “P” and “U” letter indicate phosphorylation and ubiquitination, respectively. Unknown or putative paradigms are indicated as“?”.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, X.; Valli, A.; García, J.A.; Zhou, X.; Cheng, X. The Tug-of-War between Plants and Viruses: Great Progress and Many Remaining Questions. Viruses 2019, 11, 203. https://0-doi-org.brum.beds.ac.uk/10.3390/v11030203
AMA Style
Wu X, Valli A, García JA, Zhou X, Cheng X. The Tug-of-War between Plants and Viruses: Great Progress and Many Remaining Questions. Viruses. 2019; 11(3):203. https://0-doi-org.brum.beds.ac.uk/10.3390/v11030203
Chicago/Turabian StyleWu, Xiaoyun, Adrian Valli, Juan Antonio García, Xueping Zhou, and Xiaofei Cheng. 2019. "The Tug-of-War between Plants and Viruses: Great Progress and Many Remaining Questions" Viruses 11, no. 3: 203. https://0-doi-org.brum.beds.ac.uk/10.3390/v11030203
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.