A Systematic Review of Recent Advances in Equine Influenza Vaccination

Abstract
:1. Introduction
1.1. Introduction on Equine Influenza
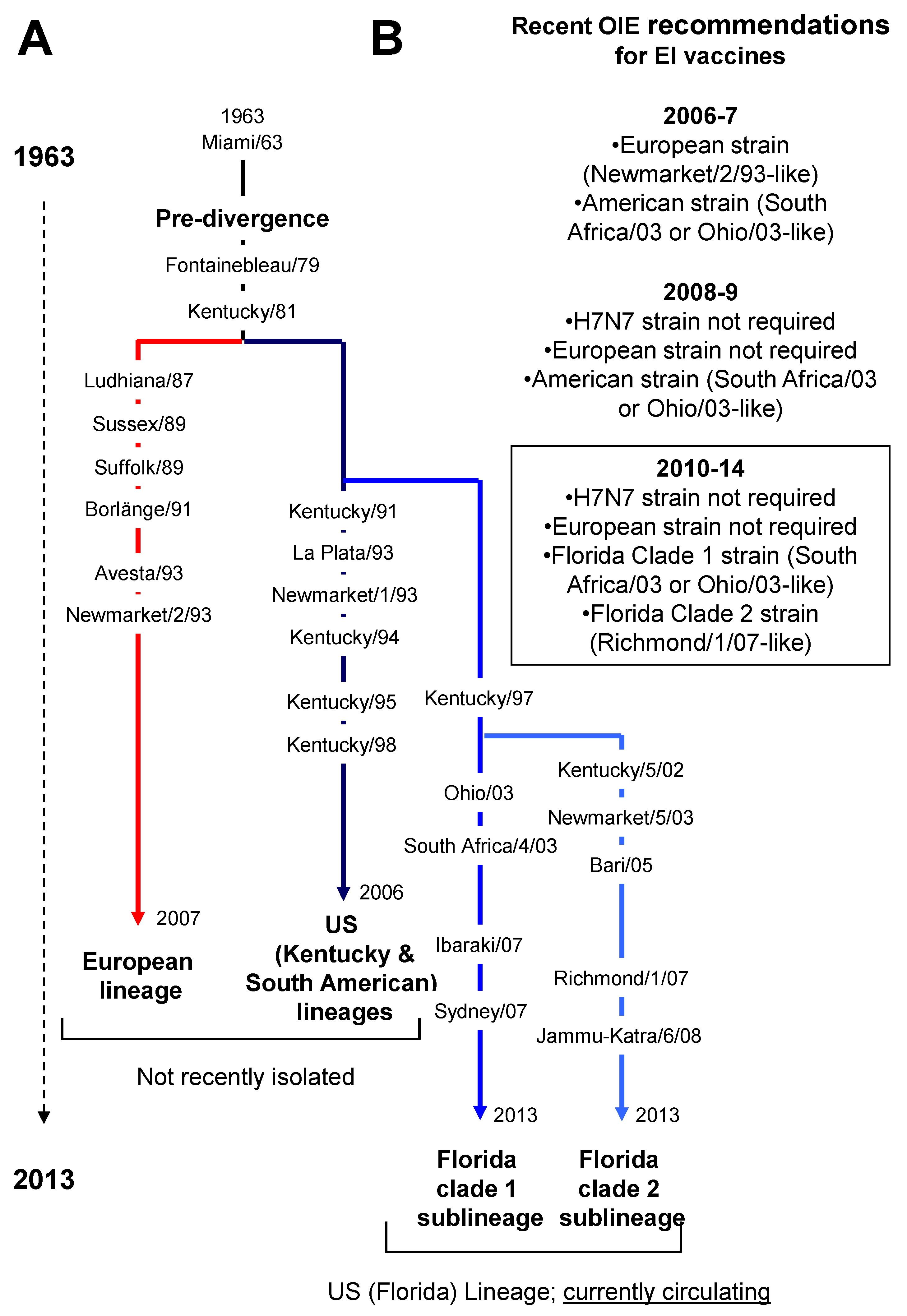
1.2. Equine Influenza Outbreaks in the Recent Years

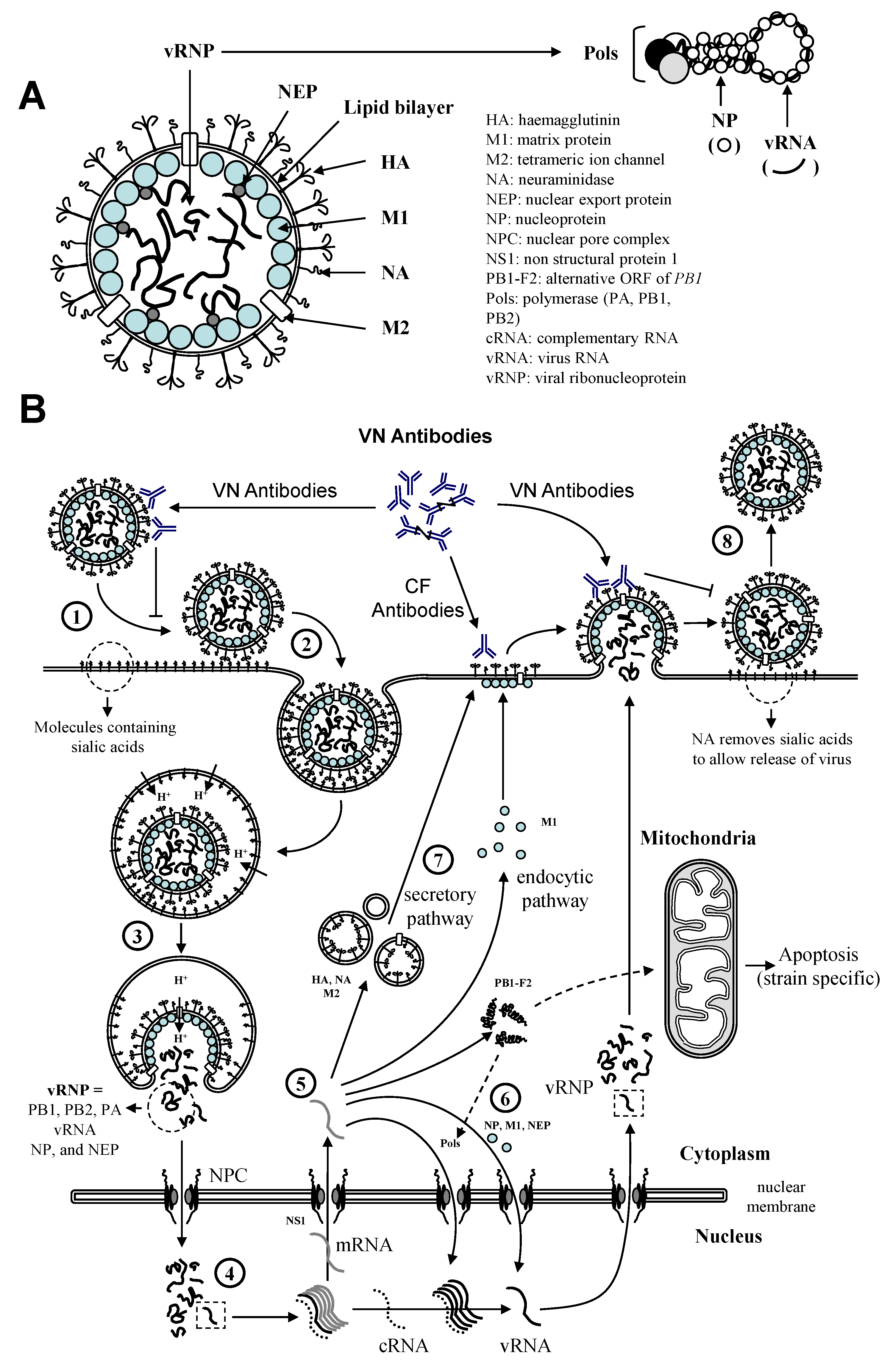
1.3. Protection against Equine influenza

2. Systematic Review: Methods
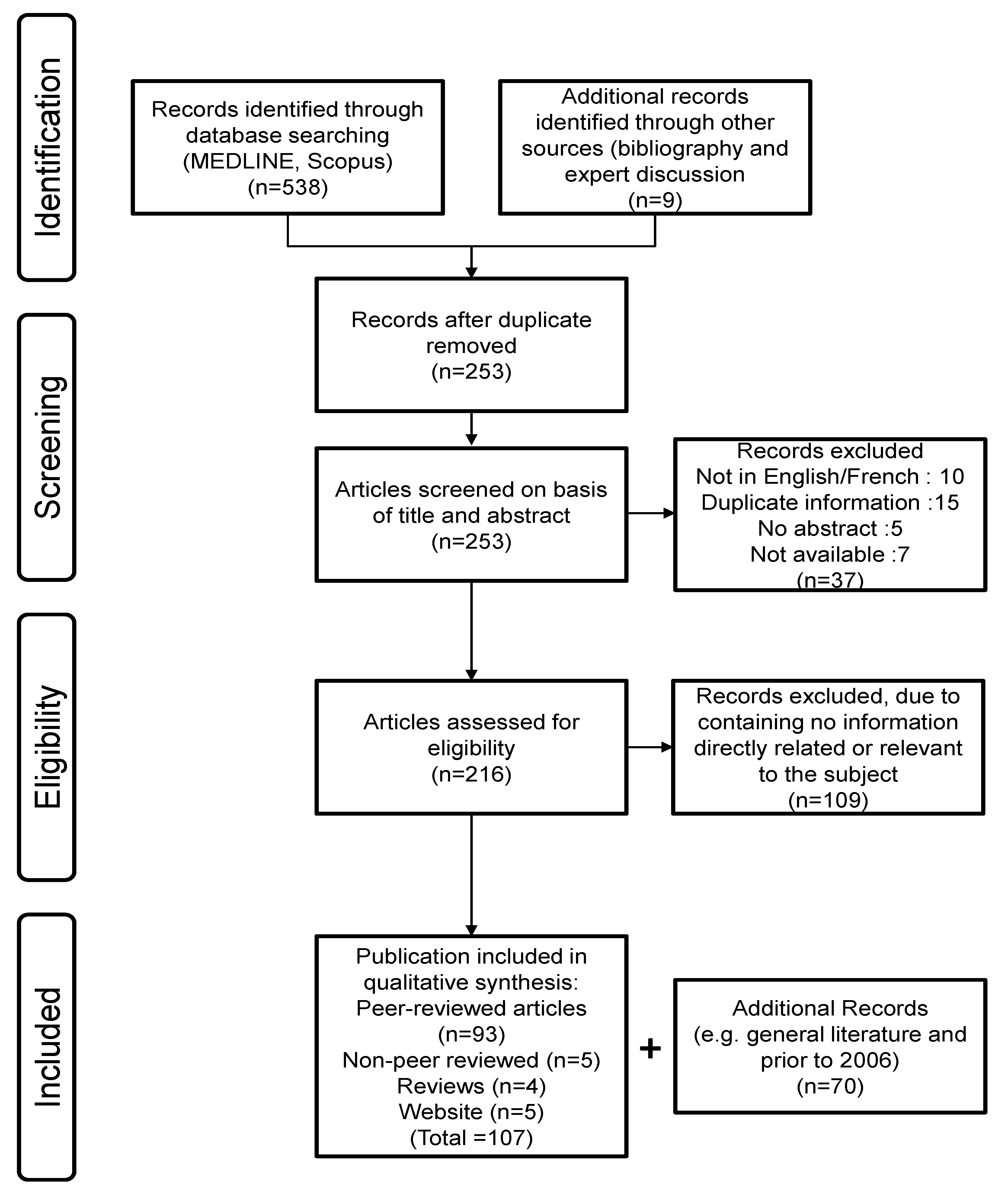
2.1. Search Strategy (Figure 3)

2.2. Publication Selection
2.3. Data Collection Process and Items
2.4. Risk of Bias in Individual Studies
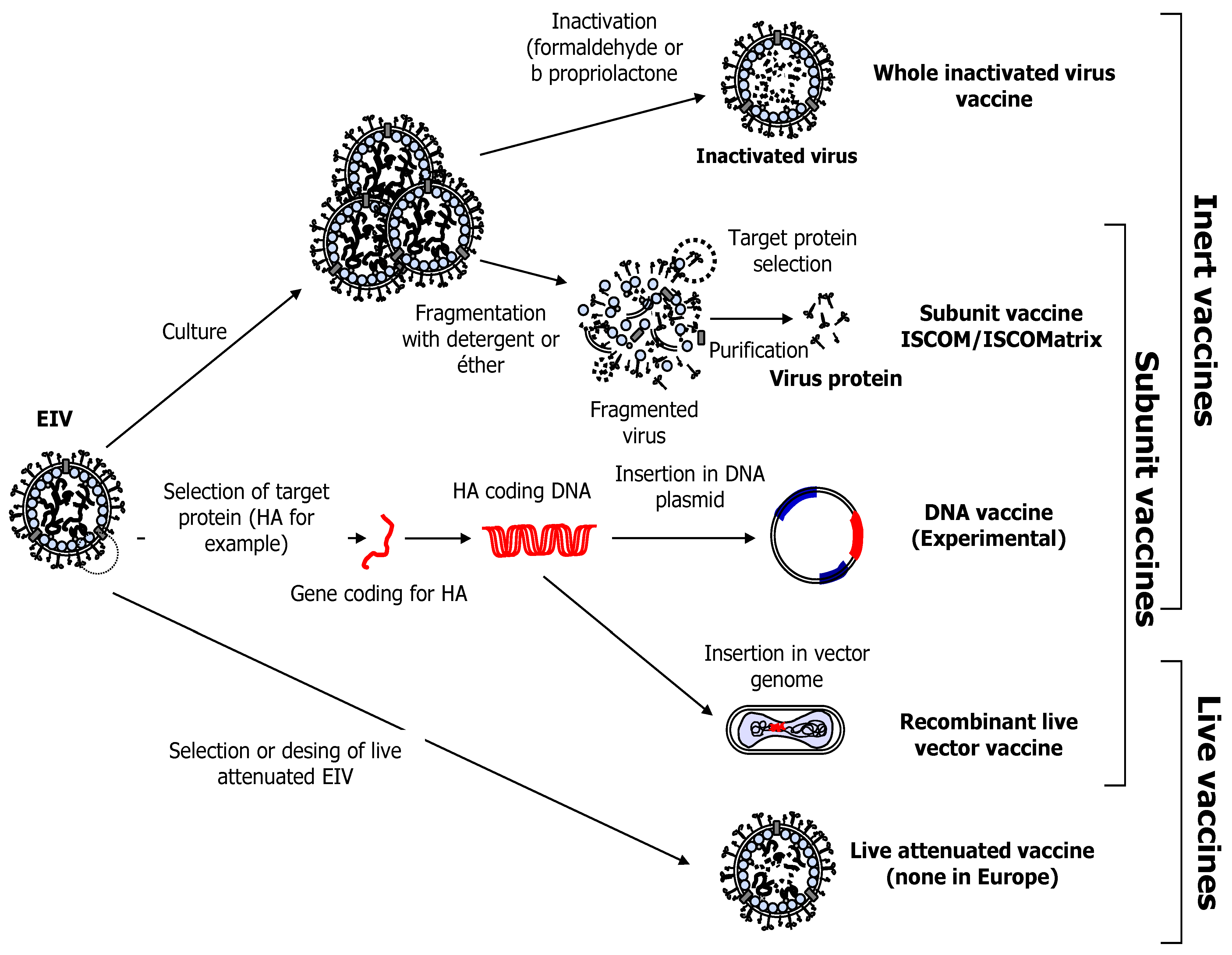
3. Equine Influenza Vaccination
- The safety of the product
- Protection: a significant reduction in clinical signs of disease (only slight signs recorded in vaccinated animals) and virus shedding is expected, when compared with unvaccinated controls animals. Neutralising immunity (i.e., neutralisation of virus leading to an absence of infection and subsequent seroconvertion) is rare.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technology | Example of Vaccine | Company | Adjuvant | Antigens | EIV Strains |
|---|---|---|---|---|---|
| Whole inactivated Sub-unit I SCOM/ISCOM-Matrix | DuvaxynTm IE Plus | Elanco | Carbopol | whole virus | Newmarket/1/93 (H3N8) Suffolk/89 (H3N8)Prague/56 (H7N7) |
| Calvenza®-03 EIV | Boehringer Ingelheim Animal Health | Carbopol | whole virus | Newmarket/2/93 (H3N8)Kentucky/2/95 (H3N8)Oiho/03 (H3N8) | |
| Equilis Prequenza (updated 2013) | MSD Animal Health | ISCOM-Matrix | whole virus | Newmarket/2/93 (H3N8)South Africa/4/03 (H3N8) | |
| Equilis Prequenza | MSD Animal Health | ISCOM-Matrix | Sub-unitHA | Prague/56 (H7N7)Newmarket/1/93 (H3N8)Newmarket/2/93 (H3N8) | |
| EquipTM F | Pfizer Ltd. | Self adjuvanting (ISCOM) | Sub-unit mainly HA and NA | Newmarket/77 (H7N7)Borlänge/91 (H3N8)Kentucky/98 (H3N8) | |
| Modified live EIV | Flu Avert® I.N. | Intervet/Schering-Plough Animal Health (US) | na | whole virus | Attenuated, cold adapted EIV: Kentucky/91 (H3N8) |
| Viral-vector based | PROTEQ FLU™ | Merial Animal Health Ltd. | Carbomer | HA | Ohio/03 (H3N8)Newmarket/2/93 (H3N8) |
| PROTEQ FLU™(updated 2014) | Merial Animal Health Ltd. | Carbomer | HA | Ohio/03 (H3N8)Richmond/1/07 (H3N8) |
3.1. Vaccine Strain Mismatch and Recent EI Outbreaks
3.2. Whole Inactivated and Sub-Unit EIV Vaccines
3.2.1. Whole Inactivated EI Vaccine and Immune Response
3.2.2. Sub-Unit EI Vaccine, ISCOM/ISCOM-Matrix Adjuvanted

3.3. DNA Vaccination against EIV
3.4. Modified Live EIV Vaccine
3.5. Non-Influenza Viruses as Vaccine Vector
3.5.1. Canarypox-Based Vaccine
3.5.2. Herpes Virus as Vaccine Vectors
3.6. Adverse Event to EI Immunisation
3.7. Vaccination in the Face of an Outbreak
4. Inefficient or Suboptimal Response of Horses to Vaccination
4.1. Immunity Gap and Interference of Pre-Existing Immunity
4.2. Immunisation with Multiple Different Types of Vaccines
4.3. Vaccination with Multiple Different Virus Strains—Effect of Antigenic Distance
4.4. Effect of Age and Maternally Derived Antibody (MDA) on the Immune Response to EI Vaccination
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Landolt, G.; Townsend, H.G.; Lunn, D.P. Equine influenza infection. In Equine Infectious Diseases; Sellon, D.C., Long, M.T., Eds.; Saunders Elsevier: St Louis, MO, USA, 2007; pp. 124–134. [Google Scholar]
- Wilson, W.D. Equine influenza. Vet. Clin. North Am. Equine Pract. 1993, 9, 257–282. [Google Scholar] [PubMed]
- Hannant, D.; Mumford, J.A. Equine influenza. In Virus Infections of Vertebrates; Studdert, M.J., Horzinek, M.C., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1996; Volume Virus Infections of Equines; pp. 285–293. [Google Scholar]
- Miller, W.C. Equine influenza. Further observations on the “Coughing” Outbreak, 1965. Vet. Rec. 1965, 77, 455–456. [Google Scholar] [PubMed]
- Peek, S.F.; Landolt, G.; Karasin, A.I.; Slack, J.A.; Steinberg, H.; Semrad, S.D.; Olsen, C.W. Acute respiratory distress syndrome and fatal interstitial pneumonia associated with equine influenza in a neonatal foal. J. Vet. Intern. Med. 2004, 18, 132–134. [Google Scholar] [CrossRef] [PubMed]
- Patterson-Kane, J.C.; Carrick, J.B.; Axon, J.E.; Wilkie, I.; Begg, A.P. The pathology of bronchointerstitial pneumonia in young foals associated with the first outbreak of equine influenza in Australia. Equine Vet. J. 2008, 40, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Sovinova, O.; Tumova, B.; Pouska, F.; Nemec, J. Isolation of a virus causing respiratory disease in horses. Acta Virol. 1958, 2, 52–61. [Google Scholar] [PubMed]
- Singh, G. Characterization of A/eq-1 virus isolated during the equine influenza epidemic in India. Acta Virol. 1994, 38, 25–26. [Google Scholar] [PubMed]
- Ismail, T.M.; Sami, A.M.; Youssef, H.M.; Abou Zaid, A.A. An outbreak of equine influenza type 1 in Egypt in 1989. Vet. Med. J. Giza 1990, 38, 195–206. [Google Scholar]
- Waddell, G.H.; Teigland, M.B.; Sigel, M.M. A new influenza virus associated with equine respiratory disease. J. Am. Vet. Med. Assoc. 1963, 143, 587–590. [Google Scholar] [PubMed]
- World Organisation for Animal Health (OIE). OIE expert surveillance panel on equine influenza vaccine composition. Available online: http://www.oie.int/fileadmin/Home/eng/Publications_%26_Documentation/docs/pdf/bulletin/Bull_2013-2-ENG.pdf (accessed on 1 April 2014).
- Daly, J.M.; Lai, A.C.; Binns, M.M.; Chambers, T.M.; Barrandeguy, M.; Mumford, J.A. Antigenic and genetic evolution of equine H3N8 influenza A viruses. J. Gen. Virol. 1996, 77, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Murcia, P.R.; Wood, J.L.; Holmes, E.C. Genome-scale evolution and phylodynamics of equine H3N8 influenza a virus. J. Virol. 2011, 85, 5312–5322. [Google Scholar] [CrossRef] [PubMed]
- Lai, A.C.; Chambers, T.M.; Holland, R.E., Jr.; Morley, P.S.; Haines, D.M.; Townsend, H.G.; Barrandeguy, M. Diverged evolution of recent equine-2 influenza (H3N8) viruses in the western hemisphere. Arch. Virol. 2001, 146, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health (OIE). Conclusion and recommendations from the expert surveillance panel on equine influenza vaccines. Available online: http://www.oie.int/fileadmin/Home/eng/Publications_%26_Documentation/docs/pdf/bulletin/Bull_2008-2-ENG.pdf (accessed on 1 April 2014).
- Bryant, N.A.; Rash, A.S.; Russell, C.A.; Ross, J.; Cooke, A.; Bowman, S.; Macrae, S.; Lewis, N.S.; Paillot, R.; Zanoni, R.; et al. Antigenic and genetic variations in European and north American equine influenza virus strains (H3N8) isolated from 2006 to 2007. Vet. Microbiol. 2009, 138, 41–52. [Google Scholar] [CrossRef]
- Bryant, N.A.; Rash, A.S.; Woodward, A.L.; Medcalf, E.; Helwegen, M.; Wohlfender, F.; Cruz, F.; Herrmann, C.; Borchers, K.; Tiwari, A.; et al. Isolation and characterisation of equine influenza viruses (H3N8) from Europe and north America from 2008 to 2009. Vet. Microbiol. 2010, 147, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Macrae, S.; Newton, J.R.; Wattrang, E.; Elton, D.M. Equine influenza: A review of an unpredictable virus. Vet. J. 2010. [Google Scholar] [CrossRef]
- Elton, D.; Bryant, N. Facing the threat of equine influenza. Equine Vet. J. 2011, 43, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Cullinane, A.; Elton, D.; Mumford, J. Equine influenza—Surveillance and control. Influenza Other Respir. Viruses 2010, 4, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Cullinane, A.; Newton, J.R. Equine influenza—A global perspective. Vet. Microbiol. 2013, 167, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, C.A.; Elahi, S.M.; Tremblay, D.; Lavoie, J.P.; Bryant, N.A.; Elton, D.M.; Carman, S.; Elsener, J. Genetic relatedness of recent Canadian equine influenza virus isolates with vaccine strains used in the field. Can. Vet. J. 2007, 48, 1028–1030. [Google Scholar] [PubMed]
- Gildea, S.; Arkins, S.; Cullinane, A. Management and environmental factors involved in equine influenza outbreaks in Ireland 2007–2010. Equine Vet. J. 2011, 43, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Gröndahl, G.; Berndtsson, L.T. The Swedish Outbreak of Equine Influenza 2007; Merial European Symposium on Equine Infectious Diseases: Lyon, France, 2008; pp. 20–21. [Google Scholar]
- Yamanaka, T.; Niwa, H.; Tsujimura, K.; Kondo, T.; Matsumura, T. Epidemic of equine influenza among vaccinated racehorses in Japan in 2007. J. Vet. Med. Sci. 2008, 70, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Nagai, M.; Hayakawa, Y.; Komae, H.; Murakami, N.; Yotsuya, S.; Asakura, S.; Sakoda, Y.; Kida, H. Genetic analyses of an H3N8 influenza virus isolate, causative strain of the outbreak of equine influenza at the Kanazawa racecourse in Japan in 2007. J. Vet. Med. Sci. 2008, 70, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Equine disease surveillance, April to June 2008: Summary of the Australian equine influenza outbreak. Vet. Rec. 2008, 163, 377–380.
- Callinan, I. Equine influenza, the August 2007 outbreak in Australia. Available online: http://www.daff.gov.au/about/publications/eiinquiry (accessed on 01 October 2014).
- Watson, J.; Daniels, P.; Kirkland, P.; Carroll, A.; Jeggo, M. The 2007 outbreak of equine influenza in Australia: Lessons learned for international trade in horses. Rev. Sci. Tech. 2011, 30, 87–93. [Google Scholar]
- Virmani, N.; Bera, B.C.; Gulati, B.R.; Karuppusamy, S.; Singh, B.K.; Vaid, R.K.; Kumar, S.; Kumar, R.; Malik, P.; Khurana, S.K.; et al. Descriptive epidemiology of equine influenza in India (2008–2009): Temporal and spatial trends. Vet. Ital. 2010, 46, 449–458. [Google Scholar]
- Yondon, M.; Heil, G.L.; Burks, J.P.; Zayat, B.; Waltzek, T.B.; Jamiyan, B.O.; McKenzie, P.P.; Krueger, W.S.; Friary, J.A.; Gray, G.C. Isolation and characterization of H3N8 equine influenza A virus associated with the 2011 epizootic in Mongolia. Influenza Other Respir. Viruses 2013, 7, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Xue-Feng, L.; Yan, Y.; Ying-Yuan, W.; Ling-Li, D.; Li-Ping, Z.; Wen-Hua, X.; Jian-Hua, Z. Equine influenza viruses isolated during outbreaks in China in 2007 and 2008. Vet. Rec. 2010, 167, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Guo, W.; Huang, W.; Dai, L.; Zhao, L.; Li, H.; Li, X.; Zhang, X.; Wang, Y.; Yan, Y.; et al. Isolation and genetic characterization of H3N8 equine influenza virus from donkeys in China. Vet. Microbiol. 2010. [Google Scholar] [CrossRef]
- Virmani, N.; Bera, B.C.; Singh, B.K.; Shanmugasundaram, K.; Gulati, B.R.; Barua, S.; Vaid, R.K.; Gupta, A.K.; Singh, R.K. Equine influenza outbreak in India (2008–09): Virus isolation, sero-epidemiology and phylogenetic analysis of HA gene. Vet. Microbiol. 2010. [Google Scholar] [CrossRef]
- OIE Animal health information. Available online: http://www.oie.int/wahis/public.php (accessed on 1 October 2014).
- AHT International collating centre. Available online: http://www.aht.org.uk/icc/linksicc.html (accessed on 1 October 2014).
- Legrand, L.J.; Pitel, P.H.; Marcillaud-Pitel, C.J.; Cullinane, A.A.; Courouce, A.M.; Fortier, G.D.; Freymuth, F.L.; Pronost, S.L. Surveillance of equine influenza viruses through the RESPE network in France from November 2005 to October 2010. Equine Vet. J. 2013. [Google Scholar] [CrossRef]
- Gildea, S.; Arkins, S.; Cullinane, A. A comparative antibody study of the potential susceptibility of thoroughbred and non-thoroughbred horse populations in Ireland to equine influenza virus. Influenza Other Respir. Viruses 2010, 4, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.G. Nasopharyngeal, tracheobronchial, and systemic immune responses to vaccination and aerosol infection with equine-2 influenza a virus (H3N8). In Equine Infectious Diseases V, Proceedings of the Fifth International Conference, Lexington, KY, 1987; University Press of Kentucky: Lexington, KY, USA, 1987; pp. 66–73. [Google Scholar]
- Bryant, N.A.; Paillot, R.; Rash, A.S.; Medcalf, E.; Montesso, F.; Ross, J.; Watson, J.; Jeggo, M.; Lewis, N.S.; Newton, J.R.; et al. Comparison of two modern vaccines and previous influenza infection against challenge with an equine influenza virus from the Australian 2007 outbreak. Vet. Res. 2010, 41. [Google Scholar] [CrossRef]
- Mumford, J.A.; Wood, J. Establishing an acceptability threshold for equine influenza vaccines. Dev. Biol. Stand. 1992, 79, 137–146. [Google Scholar]
- Mumford, J.A.; Jessett, D.M.; Dunleavy, U.; Wood, J.L.N.; Hannant, D.; Sundquist, B.; Cook, R.F. Antigenicity and immunogenicity of experimental equine influenza ISCOM vaccines. Vaccine 1994, 12, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Mumford, J.A.; Jessett, D.M.; Rollinson, E.A.; Hannant, D.; Draper, M.E. Duration of protective efficacy of equine influenza immunostimulating complex/tetanus vaccines. Vet. Rec. 1994, 134, 158–162. [Google Scholar] [CrossRef]
- Plotkin, S.A. Complex correlates of protection after vaccination. Clin. Infect. Dis. 2013, 56, 1458–1465. [Google Scholar] [CrossRef] [PubMed]
- Gildea, S.; Fitzpatrick, D.A.; Cullinane, A. Epidemiological and virological investigations of equine influenza outbreaks in Ireland (2010–2012). Influenza Other Respir. Viruses 2013, 7, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Mumford, J.; Wood, J.M.; Scott, A.M.; Folkers, C.; Schild, G.C. Studies with inactivated equine influenza vaccine. 2. Protection against experimental infection with influenza virus a/equine/newmarket/79 (H3N8). J. Hyg. (Lond.) 1983, 90, 385–395. [Google Scholar] [CrossRef]
- Mumford, J.A.; Wood, J.M.; Folkers, C.; Schild, G.C. Protection against experimental infection with influenza virus a/equine/miami/63 (H3N8) provided by inactivated whole virus vaccines containing homologous virus. Epidemiol. Infect. 1988, 100, 501–510. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health (OIE). OIE expert surveillance panel on equine influenza vaccine composition. Available online: http://www.oie.int/fileadmin/Home/eng/Publications_%26_Documentation/docs/pdf/bulletin/Bull_2011-2-ENG.pdf (accessed on 1 April 2014).
- Hannant, D.; Mumford, J.A.; Jessett, D.M. Duration of circulating antibody and immunity following infection with equine influenza virus. Vet. Rec. 1988, 122, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Fouchier, R.A.; Osterhaus, A.D. Influenza virus-specific cytotoxic T lymphocytes: A correlate of protection and a basis for vaccine development. Curr. Opin. Biotechnol. 2007, 18, 529–536. [Google Scholar] [CrossRef]
- Hillaire, M.L.; Osterhaus, A.D.; Rimmelzwaan, G.F. Induction of virus-specific cytotoxic T lymphocytes as a basis for the development of broadly protective influenza vaccines. J. Biomed. Biotechnol. 2011. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar]
- Nishiura, H.; Satou, K. Potential effectiveness of public health interventions during the equine influenza outbreak in racehorse facilities in Japan, 2007. Transbound. Emerg. Dis. 2010, 57, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Firestone, S.M.; Schemann, K.A.; Toribio, J.A.; Ward, M.P.; Dhand, N.K. A case-control study of risk factors for equine influenza spread onto horse premises during the 2007 epidemic in Australia. Prev. Vet. Med. 2011, 100, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Hannant, D.; Kydd, J.H.; Daly, J.M. Vaccination against equine influenza: Quid novi? Vaccine 2006, 24, 4047–4061. [Google Scholar] [CrossRef]
- Equine influenza vaccine (inactivated). In European Pharmacopoeia 7.0; European Directorate for the Quality of Medicine and Healthcare (EDQM) Publication, Council of Europe: Strasbourg, France, 2010; pp. 893–895.
- Woodland, R.M. Influenza vaccine strains: Licensing perspectives. Equine Vet. J. 2013, 45, 772–773. [Google Scholar] [CrossRef] [PubMed]
- EquiFluNet Global surveillance network for equine influenza. Vaccination regulations and dosing regimes. Available online: http://www.equiflunet.org.uk/ (accessed on 1 October 2014).
- Barbic, L.; Madic, J.; Turk, N.; Daly, J. Vaccine failure caused an outbreak of equine influenza in Croatia. Vet. Microbiol. 2009, 133, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Newton, J.R.; Daly, J.M.; Spencer, L.; Mumford, J.A. Description of the outbreak of equine influenza (H3N8) in the united kingdom in 2003, during which recently vaccinated horses in Newmarket developed respiratory disease. Vet. Rec. 2006, 158, 185–192. [Google Scholar] [CrossRef]
- Barquero, N.; Daly, J.M.; Newton, J.R. Risk factors for influenza infection in vaccinated racehorses: Lessons from an outbreak in Newmarket, UK in 2003. Vaccine 2007, 25, 7520–7529. [Google Scholar] [CrossRef] [PubMed]
- Damiani, A.M.; Scicluna, M.T.; Ciabatti, I.; Cardeti, G.; Sala, M.; Vulcano, G.; Cordioli, P.; Martella, V.; Amaddeo, D.; Autorino, G.L. Genetic characterization of equine influenza viruses isolated in Italy between 1999 and 2005. Virus Res. 2008, 131, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Elia, G.; Decaro, N.; di Trani, L.; Lorusso, E.; Campolo, M.; Desario, C.; Parisi, A.; Cavaliere, N.; Buonavoglia, C. An outbreak of equine influenza virus in vaccinated horses in Italy is due to an H3N8 strain closely related to recent North American representatives of the Florida sub-lineage. Vet. Microbiol. 2007, 121, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I.A.; Cox, N.J. Structural basis of immune recognition of influenza virus hemagglutinin. Annu. Rev. Immunol. 1990, 8, 737–771. [Google Scholar] [CrossRef] [PubMed]
- Park, A.W.; Daly, J.M.; Lewis, N.S.; Smith, D.J.; Wood, J.L.; Grenfell, B.T. Quantifying the impact of immune escape on transmission dynamics of influenza. Science 2009, 326, 726–728. [Google Scholar] [CrossRef] [PubMed]
- Ohta, M.; Yamanaka, T.; Yoshinari, T.; Matsumura, T. Antibody responses against American and European lineage strains of equine-2 influenza virus among racehorses inoculated with the new vaccine. J. Equine Sci. 2007, 18, 117–120. [Google Scholar] [CrossRef]
- Yamanaka, T.; Bannai, H.; Nemoto, M.; Tsujimura, K.; Kondo, T.; Matsumura, T. Antibody responses induced by Japanese whole inactivated vaccines against equine influenza virus (H3N8) belonging to Florida sublineage clade2. J. Vet. Med. Sci. 2011, 73, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Yates, P.; Mumford, J.A. Equine influenza vaccine efficacy: The significance of antigenic variation. Vet. Microbiol. 2000, 74, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Yates, P.J.; Newton, J.R.; Park, A.; Henley, W.; Wood, J.L.; Davis-Poynter, N.; Mumford, J.A. Evidence supporting the inclusion of strains from each of the two co-circulating lineages of H3N8 equine influenza virus in vaccines. Vaccine 2004, 22, 4101–4109. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Yates, R.J.; Browse, G.; Swann, Z.; Newton, J.R.; Jessett, D.; Davis-Poynter, N.; Mumford, J.A. Comparison of hamster and pony challenge models for evaluation of effect of antigenic drift on cross protection afforded by equine influenza vaccines. Equine Vet. J. 2003, 35, 458–462. [Google Scholar] [PubMed]
- Paillot, R.; Prowse, L.; Donald, C.; Medcalf, E.; Montesso, F.; Bryant, N.; Watson, J.; Jeggo, M.; Elton, D.; Newton, R.; et al. Efficacy of a whole inactivated EI vaccine against a recent EIV outbreak isolate and comparative detection of virus shedding. Vet. Immunol. Immunopathol. 2010, 136, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Sindle, T.; Tearle, J.; Barquero, N.; Newton, J.R.; Corning, S. Equine influenza vaccine containing older H3N8 strains offers protection against A/eq/south africa/4/03 (H3N8) strain in a short-term vaccine efficacy study. Equine Vet. J. 2007, 39, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Prowse, L.; Montesso, F.; Huang, C.M.; Barnes, H.; Escala, J. Whole inactivated equine influenza vaccine: Efficacy against a representative clade 2 equine influenza virus, IFNgamma synthesis and duration of humoral immunity. Vet. Microbiol. 2013, 162, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Elton, D. Potential of a sequence-based antigenic distance measure to indicate equine influenza vaccine strain efficacy. Vaccine 2013, 31, 6043–6045. [Google Scholar] [CrossRef] [PubMed]
- Genzel, Y.; Fischer, M.; Reichl, U. Serum-free influenza virus production avoiding washing steps and medium exchange in large-scale microcarrier culture. Vaccine 2006, 24, 3261–3272. [Google Scholar] [CrossRef] [PubMed]
- Genzel, Y.; Olmer, R.M.; Schafer, B.; Reichl, U. Wave microcarrier cultivation of MDCK cells for influenza virus production in serum containing and serum-free media. Vaccine 2006, 24, 6074–6087. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, E.B. Aluminium adjuvants—In retrospect and prospect. Vaccine 2004, 22, 3658–3668. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Newton, J.R.; Mumford, J.A. Current perspectives on control of equine influenza. Vet. Res. 2004, 35, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Gildea, S.; Quinlivan, M.; Murphy, B.A.; Cullinane, A. Humoral response and antiviral cytokine expression following vaccination of thoroughbred weanlings—A blinded comparison of commercially available vaccines. Vaccine 2013. [Google Scholar] [CrossRef]
- Wattrang, E.; Jessett, D.M.; Yates, P.; Fuxler, L.; Hannant, D. Experimental infection of ponies with equine influenza A2 (H3N8) virus strains of different pathogenicity elicits varying interferon and interleukin-6 responses. Viral Immunol. 2003, 16, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Quinlivan, M.; Nelly, M.; Prendergast, M.; Breathnach, C.; Horohov, D.; Arkins, S.; Chiang, Y.W.; Chu, H.J.; Ng, T.; Cullinane, A. Pro-inflammatory and antiviral cytokine expression in vaccinated and unvaccinated horses exposed to equine influenza virus. Vaccine 2007, 25, 7056–7064. [Google Scholar] [CrossRef] [PubMed]
- Gildea, S.; Arkins, S.; Walsh, C.; Cullinane, A. A comparison of antibody responses to commercial equine influenza vaccines following primary vaccination of thoroughbred weanlings—A randomised blind study. Vaccine 2011, 29, 9214–9223. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.A.; Townsend, H.G.; Kohler, A.K.; Hussey, S.; Breathnach, C.; Barnett, C.; Holland, R.; Lunn, D.P. Immune responses to commercial equine vaccines against equine herpesvirus-1, equine influenza virus, eastern equine encephalomyelitis, and tetanus. Vet. Immunol. Immunopathol. 2006, 111, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.M.; Hecker, R.; Mutwiri, G.; van Drunen Littel-van den Hurk, S.; Babiuk, L.A.; Townsend, H.G. Formulation with CpG ODN enhances antibody responses to an equine influenza virus vaccine. Vet. Immunol. Immunopathol. 2006, 114, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Anonimous Equilis new prequenza. Available online: http://www.msd-animal-health.no/binaries/2013_GAH_EQ_004_Prequenza_DetailAid_v2_m_FK_tekster_Screen_tcm84–136701.pdf (accessed on 23 October 2013).
- Pearse, M.J.; Drane, D. Iscomatrix adjuvant for antigen delivery. Adv. Drug Deliv. Rev. 2005, 57, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Xie, Y.; Ye, Y.P. Iscoms and iscomatrix. Vaccine 2009, 27, 4388–4401. [Google Scholar] [CrossRef] [PubMed]
- Sjolander, A.; Cox, J.C.; Barr, I.G. Iscoms: An adjuvant with multiple functions. J. Leukoc. Biol. 1998, 64, 713–723. [Google Scholar] [PubMed]
- Watson, D.L.; Watson, N.A.; Fossum, C.; Lovgren, K.; Morein, B. Interactions between immune-stimulating complexes (ISCOMS) and peritoneal mononuclear leucocytes. Microbiol. Immunol. 1992, 36, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Bungener, L.; Huckriede, A.; Wilschut, J.; Daemen, T. Delivery of protein antigens to the immune system by fusion-active virosomes: A comparison with liposomes and ISCOMS. Biosci. Rep. 2002, 22, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Morein, B.; Bengtsson, K.L. Immunomodulation by ISCOMS, immune stimulating complexes. Methods 1999, 19, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Robson, N.C.; McAlpine, T.; Knights, A.J.; Schnurr, M.; Shin, A.; Chen, W.; Maraskovsky, E.; Cebon, J. Processing and cross-presentation of individual HLA-A, -B, or -C epitopes from NY-ESO-1 or an HLA-A epitope for Melan-A differ according to the mode of antigen delivery. Blood 2010, 116, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Villacres, M.C.; Behboudi, S.; Nikkila, T.; Lovgren-Bengtsson, K.; Morein, B. Internalization of ISCOM-borne antigens and presentation under MHC class I or class II restriction. Cell. Immunol. 1998, 185, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Sjolander, A.; van’t Land, B.; Lovgren-Bengtsson, K. Iscoms containing purified Quillaja saponins upregulate both Th1-like and Th2-like immune responses. Cell. Immunol. 1997, 177, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Sambhara, S.; Kurichh, A.; Miranda, R.; Tamane, A.; Arpino, R.; James, O.; McGuinness, U.; Kandil, A.; Underdown, B.; Klein, M.; et al. Enhanced immune responses and resistance against infection in aged mice conferred by Flu-ISCOMs vaccine correlate with up-regulation of costimulatory molecule CD86. Vaccine 1998, 16, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.A.; Petersen, H.H.; Ersboll, A.K.; Falk-Ronne, J.; Jacobsen, S. Vaccination elicits a prominent acute phase response in horses. Vet. J. 2012, 191, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Grimmett, H.; Elton, D.; Daly, J.M. Protection, systemic IFN and antibody responses induced by an ISCOM-based vaccine against a recent equine influenza virus in its natural host. Vet. Res. 2008. [Google Scholar] [CrossRef]
- Bryant, N.; Rash, A.; Lewis, N.; Elton, D.; Montesso, F.; Ross, J.; Newton, R.; Paillot, R.; Watson, J.; Jeggo, M. Australian equine influenza: Vaccine protection in the UK. Vet. Rec. 2008, 162, 491–492. [Google Scholar] [CrossRef] [PubMed]
- Van de Zande, S.; Ragni-Alunni, R. Efficacy of Equilis Prequenza in Horses Individually Challenged with a/equi2/newmarket/5/03. In Proceedings of the 10th International Congress of World Equine Veterinary Association, Moscow, Russia, 28 January–1 February 2008; IVIS: Moscow, Russia; pp. 428–430.
- Ragni-Alunni, R.; van de Zande, S. Animal Studies with Individual Challenge Method. In Efficacy of Equilis Prequenza te against Recent Equine Influenza Isolates; Equitana: Essen, Germany, 2009. [Google Scholar]
- Cullinane, A.; Weld, J.; Osborne, M.; Nelly, M.; McBride, C.; Walsh, C. Field studies on equine influenza vaccination regimes in thoroughbred foals and yearlings. Vet. J. 2001, 161, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Heldens, J.G.; Pouwels, H.G.; Derks, C.G.; van de Zande, S.M.; Hoeijmakers, M.J. The first safe inactivated equine influenza vaccine formulation adjuvanted with ISCOM-matrix that closes the immunity gap. Vaccine 2009, 27, 5530–5537. [Google Scholar] [CrossRef] [PubMed]
- Crouch, C.F.; Daly, J.; Hannant, D.; Wilkins, J.; Francis, M.J. Immune responses and protective efficacy in ponies immunised with an equine influenza ISCOM vaccine containing an “American lineage” H3N8 virus. Vaccine 2004, 23, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Crouch, C.F.; Daly, J.; Henley, W.; Hannant, D.; Wilkins, J.; Francis, M.J. The use of a systemic prime/mucosal boost strategy with an equine influenza ISCOM vaccine to induce protective immunity in horses. Vet. Immunol. Immunopathol. 2005, 108, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Prowse, L. Iscom-matrix-based equine influenza (EIV) vaccine stimulates cell-mediated immunity in the horse. Vet. Immunol. Immunopathol. 2012, 145, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Prowse, L.; Montesso, F.; Stewart, B.; Jordon, L.; Newton, J.R.; Gilkerson, J.R. Duration of equine influenza virus shedding and infectivity in immunised horses after experimental infection with EIV A/eq2/richmond/1/07. Vet. Microbiol. 2013, 166, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Heldens, J.G.; Pouwels, H.G.; Derks, C.G.; van de Zande, S.M.; Hoeijmakers, M.J. Duration of immunity induced by an equine influenza and tetanus combination vaccine formulation adjuvanted with ISCOM-matrix. Vaccine 2010, 28, 6989–6996. [Google Scholar] [CrossRef] [PubMed]
- Lunn, D.P.; Soboll, G.; Schram, B.R.; Quass, J.; McGregor, M.W.; Drape, R.J.; Macklin, M.D.; McCabe, D.E.; Swain, W.F.; Olsen, C.W. Antibody responses to DNA vaccination of horses using the influenza virus hemagglutinin gene. Vaccine 1999, 17, 2245–2258. [Google Scholar] [CrossRef] [PubMed]
- Soboll, G.; Horohov, D.W.; Aldridge, B.M.; Olsen, C.W.; McGregor, M.W.; Drape, R.J.; Macklin, M.D.; Swain, W.F.; Lunn, D.P. Regional antibody and cellular immune responses to equine influenza virus infection, and particle mediated DNA vaccination. Vet. Immunol. Immunopathol. 2003, 94, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Soboll, G.; Nelson, K.M.; Leuthner, E.S.; Clark, R.J.; Drape, R.; Macklin, M.D.; Swain, W.F.; Olsen, C.W.; Lunn, D.P. Mucosal co-administration of cholera toxin and influenza virus hemagglutinin-DNA in ponies generates a local IgA response. Vaccine 2003, 21, 3081–3092. [Google Scholar] [CrossRef] [PubMed]
- Landolt, G.A.; Hussey, S.B.; Kreutzer, K.; Quintana, A.; Lunn, D.P. Low-dose DNA vaccination into the submandibular lymph nodes in ponies. Vet. Rec. 2010, 167, 302–303. [Google Scholar] [CrossRef] [PubMed]
- Ault, A.; Zajac, A.M.; Kong, W.P.; Gorres, J.P.; Royals, M.; Wei, C.J.; Bao, S.; Yang, Z.Y.; Reedy, S.E.; Sturgill, T.L.; et al. Immunogenicity and clinical protection against equine influenza by DNA vaccination of ponies. Vaccine 2012, 30, 3965–3974. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.M.; Holland, R.E.; Tudor, L.R.; Townsend, H.G.; Cook, A.; Bogdan, J.; Lunn, D.P.; Hussey, S.; Whitaker-Dowling, P.; Youngner, J.S.; et al. A new modified live equine influenza virus vaccine: Phenotypic stability, restricted spread and efficacy against heterologous virus challenge. Equine Vet. J. 2001, 33, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Lunn, D.P.; Hussey, S.; Sebing, R.; Rushlow, K.E.; Radecki, S.V.; Whitaker-Dowling, P.; Youngner, J.S.; Chambers, T.M.; Holland, R.E., Jr.; Horohov, D.W. Safety, efficacy, and immunogenicity of a modified-live equine influenza virus vaccine in ponies after induction of exercise-induced immunosuppression. J. Am. Vet. Med. Assoc. 2001, 218, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Schickli, J.H.; Flandorfer, A.; Nakaya, T.; Martinez-Sobrido, L.; Garcia-Sastre, A.; Palese, P. Plasmid-only rescue of influenza a virus vaccine candidates. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2001, 356, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Quinlivan, M.; Dempsey, E.; Ryan, F.; Arkins, S.; Cullinane, A. Real-time reverse transcription pcr for detection and quantitative analysis of equine influenza virus. J. Clin. Microbiol. 2005, 43, 5055–5057. [Google Scholar] [CrossRef] [PubMed]
- Quinlivan, M.; Zamarin, D.; Garcia-Sastre, A.; Cullinane, A.; Chambers, T.; Palese, P. Attenuation of equine influenza viruses through truncations of the NS1 protein. J. Virol. 2005, 79, 8431–8439. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.M.; Quinlivan, M.; Sturgill, T.; Cullinane, A.; Horohov, D.W.; Zamarin, D.; Arkins, S.; Garcia-Sastre, A.; Palese, P. Influenza a viruses with truncated NS1 as modified live virus vaccines: Pilot studies of safety and efficacy in horses. Equine Vet. J. 2009, 41, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Toulemonde, C.E.; Daly, J.; Sindle, T.; Guigal, P.M.; Audonnet, J.C.; Minke, J.M. Efficacy of a recombinant equine influenza vaccine against challenge with an American lineage H3N8 influenza virus responsible for the 2003 outbreak in the United Kingdom. Vet. Rec. 2005, 156, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Kydd, J.H.; Sindle, T.; Hannant, D.; Edlund Toulemonde, C.; Audonnet, J.C.; Minke, J.M.; Daly, J.M. Antibody and IFN-gamma responses induced by a recombinant canarypox vaccine and challenge infection with equine influenza virus. Vet. Immunol. Immunopathol. 2006, 112, 225–233. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health (OIE). Conclusion and recommendations from the expert surveillance panel on equine influenza vaccines. Available online: http://www.oie.int/fileadmin/Home/eng/Publications_%26_Documentation/docs/pdf/bulletin/Bull_2006-2-ENG.pdf (accessed on 1 Aprile 2014).
- Soboll, G.; Hussey, S.B.; Minke, J.M.; Landolt, G.A.; Hunter, J.S.; Jagannatha, S.; Lunn, D.P. Onset and duration of immunity to equine influenza virus resulting from canarypox-vectored (ALVAC®) vaccination. Vet. Immunol. Immunopathol. 2010, 135, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Kannegieter, N.J.; Frogley, A.; Crispe, E.; Kirkland, P.D. Clinical outcomes and virology of equine influenza in a naive population and in horses infected soon after receiving one dose of vaccine. Aust. Vet. J. 2011, 89, S139–S142. [Google Scholar] [CrossRef] [PubMed]
- Minke, J.M.; El-Hage, C.M.; Tazawa, P.; Homer, D.; Lemaitre, L.; Cozette, V.; Gilkerson, J.R.; Kirkland, P.D. Evaluation of the response to an accelerated immunisation schedule using a canarypox-vectored equine influenza vaccine, shortened interdose intervals and vaccination of young foals. Aust. Vet. J. 2011, 89, S137–S139. [Google Scholar] [CrossRef] [PubMed]
- El-Hage, C.M.; Savage, C.J.; Minke, J.M.; Ficorilli, N.P.; Watson, J.; Gilkerson, J.R. Accelerated vaccination schedule provides protective levels of antibody and complete herd immunity to equine influenza. Equine Vet. J. 2013, 45, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Arthur, R.J.; Suann, C.J. Biosecurity and vaccination strategies to minimise the effect of an equine influenza outbreak on racing and breeding. Aust. Vet. J. 2011, 89, S109–S113. [Google Scholar]
- Gildea, S.; Arkins, S.; Walsh, C.; Cullinane, A. A comparison of antibody responses to commercial equine influenza vaccines following annual booster vaccination of national hunt horses—A randomised blind study. Vaccine 2011, 29, 3917–3922. [Google Scholar] [CrossRef] [PubMed]
- Minke, J.M.; Toulemonde, C.E.; Coupier, H.; Guigal, P.M.; Dinic, S.; Sindle, T.; Jessett, D.; Black, L.; Bublot, M.; Pardo, M.C.; et al. Efficacy of a canarypox-vectored recombinant vaccine expressing the hemagglutinin gene of equine influenza H3N8 virus in the protection of ponies from viral challenge. Am. J. Vet. Res. 2007, 68, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Paillot, R.; Kydd, J.H.; Macrae, S.; Minke, J.M.; Hannant, D.; Daly, J.M. New assays to measure equine influenza virus-specific type 1 immunity in horses. Vaccine 2007, 25, 7385–7398. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.A.; Sturgill, T.L.; Breathnach, C.C.; Chambers, T.M.; Siger, L.; Minke, J.M.; Horohov, D.W. Humoral and cell-mediated immune responses of old horses following recombinant canarypox virus vaccination and subsequent challenge infection. Vet. Immunol. Immunopathol. 2011, 139, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Gotch, F.; McMichael, A.; Smith, G.; Moss, B. Identification of viral molecules recognized by influenza-specific human cytotoxic T lymphocytes. J. Exp. Med. 1987, 165, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Kanesa-Thasan, N.; Smucny, J.J.; Hoke, C.H.; Marks, D.H.; Konishi, E.; Kurane, I.; Tang, D.B.; Vaughn, D.W.; Mason, P.W.; Shope, R.E. Safety and immunogenicity of NYVAC-JEV and ALVAC-JEV attenuated recombinant japanese encephalitis virus—Poxvirus vaccines in vaccinia-nonimmune and vaccinia-immune humans. Vaccine 2000, 19, 483–491. [Google Scholar] [CrossRef] [PubMed]
- El Garch, H.; Minke, J.M.; Rehder, J.; Richard, S.; Edlund Toulemonde, C.; Dinic, S.; Andreoni, C.; Audonnet, J.C.; Nordgren, R.; Juillard, V. A west nile virus (WNV) recombinant canarypox virus vaccine elicits WNV-specific neutralizing antibodies and cell-mediated immune responses in the horse. Vet. Immunol. Immunopathol. 2008, 123, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, A.J.; Quan, M.; Lourens, C.W.; Audonnet, J.C.; Minke, J.M.; Yao, J.; He, L.; Nordgren, R.; Gardner, I.A.; Maclachlan, N.J. Protective immunization of horses with a recombinant canarypox virus vectored vaccine co-expressing genes encoding the outer capsid proteins of African horse sickness virus. Vaccine 2009, 27, 4434–4438. [Google Scholar] [CrossRef] [PubMed]
- Karaca, K.; Dubovi, E.J.; Siger, L.; Robles, A.; Audonnet, J.C.; Jiansheng, Y.; Nordgren, R.; Minke, J.M. Evaluation of the ability of canarypox-vectored equine influenza virus vaccines to induce humoral immune responses against canine influenza viruses in dogs. Am. J. Vet. Res. 2007, 68, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P.C.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.P.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J.; et al. Transmission of equine influenza virus to dogs. Science 2005, 310, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Rosas, C.; van de Walle, G.R.; Metzger, S.M.; Hoelzer, K.; Dubovi, E.J.; Kim, S.G.; Parrish, C.R.; Osterrieder, N. Evaluation of a vectored equine herpesvirus type 1 (EHV-1) vaccine expressing H3 haemagglutinin in the protection of dogs against canine influenza. Vaccine 2008, 26, 2335–2343. [Google Scholar] [CrossRef] [PubMed]
- Van de Walle, G.R.; May, M.A.; Peters, S.T.; Metzger, S.M.; Rosas, C.T.; Osterrieder, N. A vectored equine herpesvirus type 1 (EHV-1) vaccine elicits protective immune responses against EHV-1 and H3N8 equine influenza virus. Vaccine 2010, 28, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Reeve-Johnson, L. Pharmaco-vigilance observations on a recombinant vector vaccine used as part of the eradication campaign for equine influenza in Australia. J. Vet. Pharmacol. Ther. 2009, 32, 117–118. [Google Scholar]
- Gershwin, L.J.; Netherwood, K.A.; Norris, M.S.; Behrens, N.E.; Shao, M.X. Equine IgE responses to non-viral vaccine components. Vaccine 2012, 30, 7615–7620. [Google Scholar] [CrossRef] [PubMed]
- Kannegieter, N.J.; Schaaf, K.L.; Lovell, D.K.; Simon, C.D.; Stone, B.M. Myofibroblastic fibrosarcoma with multifocal osseous metaplasia at the site of equine influenza vaccination. Aust. Vet. J. 2010, 88, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Baguelin, M.; Newton, J.R.; Demiris, N.; Daly, J.; Mumford, J.A.; Wood, J.L. Control of equine influenza: Scenario testing using a realistic metapopulation model of spread. J. R. Soc. Interface 2010, 7, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.; Cooper, K.; Williams, J.; Eastwood, S.; Peake, C. Equine influenza immunity in the special restricted area (purple zone) of New South Wales, Australia. Aust. Vet. J. 2011, 89, S116–S120. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.R.; Webster, W.R.; Wright, T.; Denney, I.; Links, I. Vaccination program in the response to the 2007 equine influenza outbreak in Australia. Aust. Vet. J. 2011, 89, S126–S134. [Google Scholar]
- Kirkland, P.D.; Delbridge, G. Use of a blocking ELISA for antibodies to equine influenza virus as a test to distinguish between naturally infected and vaccinated horses: Proof of concept studies. Aust. Vet. J. 2011, 89, S45–S46. [Google Scholar] [CrossRef] [PubMed]
- Galvin, P.; Gildea, S.; Arkins, S.; Walsh, C.; Cullinane, A. The evaluation of a nucleoprotein ELISA for the detection of equine influenza antibodies and the differentiation of infected from vaccinated horses (DIVA). Influenza Other Respir. Viruses 2013, 7, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.G.; Cowled, B.; East, I.J.; Moloney, B.J.; Kung, N.Y. Evaluating the effectiveness of early vaccination in the control and eradication of equine influenza—A modelling approach. Prev. Vet. Med. 2011, 99, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, P.; Sergeant, E.S. Risk of an equine influenza virus reservoir establishing in wild horses in new south wales during the Australian epidemic. Aust. Vet. J. 2011, 89, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Happold, J.; Rubira, R. Equine influenza: Patterns of disease and seroprevalence in thoroughbred studs and implications for vaccination. Aust. Vet. J. 2011, 89, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.G.; Scanlan, W.A.; Cowled, B.D.; Carroll, A. Regaining Australia’s equine influenza-free status: A national perspective. Aust. Vet. J. 2011, 89, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Cullinane, A.; Gildea, S.; Weldon, E. Comparison of primary vaccination regimes for equine influenza—Working towards an evidence based regime. Equine Vet. J. 2013. [Google Scholar] [CrossRef]
- May, T. Vaccines as community-focused therapy. Expert Rev. Vaccines 2003, 2, 341–343. [Google Scholar] [CrossRef] [PubMed]
- Satou, K.; Nishiura, H. Basic reproduction number for equine-2 influenza virus a (H3N8) epidemic in racehorse facilities in Japan, 1971. J. Equine Vet. Sci. 2006, 26, 310–316. [Google Scholar] [CrossRef]
- Ireland, J.L.; Wylie, C.E.; Collins, S.N.; Verheyen, K.L.; Newton, J.R. Preventive health care and owner-reported disease prevalence of horses and ponies in great Britain. Res. Vet. Sci. 2013, 95, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Slater, J. National equine health survey. Available online: http://www.bluecross.org.uk/80135/national-equine-helath-survey.html (accessed on 22 January 2014).
- Boni, M.F. Vaccination and antigenic drift in influenza. Vaccine 2008, 26, C8–C14. [Google Scholar]
- Daly, J.M.; Newton, J.R.; Wood, J.L.; Park, A.W. What can mathematical models bring to the control of equine influenza? Equine Vet. J. 2013, 45, 784–788. [Google Scholar]
- Folsom, R.W.; Littlefield-Chabaud, M.A.; French, D.D.; Pourciau, S.S.; Mistric, L.; Horohov, D.W. Exercise alters the immune response to equine influenza virus and increases susceptibility to infection. Equine Vet. J. 2001, 33, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.L.; Mumford, J.A.; Mair, T.S.; Slater, J. Boosting in equine influenza vaccination schedules: Timing and time for a re-evaluation of requirements of national and international authorities. Vet. J. 2007, 174, 449–450. [Google Scholar] [CrossRef]
- Murcia, P.R.; Baillie, G.J.; Stack, J.C.; Jervis, C.; Elton, D.; Mumford, J.A.; Daly, J.; Kellam, P.; Grenfell, B.T.; Holmes, E.C.; et al. Evolution of equine influenza virus in vaccinated horses. J. Virol. 2013, 87, 4768–4771. [Google Scholar] [CrossRef] [PubMed]
- Heldens, J.G.; van Loon, A.A.; van de Zande, S. Is there a benefit from an early booster vaccination in the control of equine influenza? Vet. J. 2007, 174, 592–598. [Google Scholar]
- Van de Zande, S. Intervet International Prime-boost Vaccine for the Protection of Equine Viral Infection. Patent WO/2007/051763. 10 May 2007. Available online: http://patentscope.wipo.int/search/en/detail.jsf?docId=WO2007051763&recNum=1&maxRec=&office=&prevFilter=&sortOption=&queryString=&tab=PCT+Biblio (accessed on 1 October 2014).
- Haaheim, L.R.; Schild, G.C. Antibodies to the strain-specific and cross-reactive determinants of the haemagglutinin of influenza H3N2 viruses. Antiviral activities of the antibodies in biological systems. Acta Pathol. Microbiol. Scand. 1980, 88, 335–340. [Google Scholar]
- Burrows, R.; Denyer, M.; Goodridge, D.; Hamilton, F. Field and laboratory studies of equine influenza viruses isolated in 1979. Vet. Rec. 1981, 109, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Burrows, R.; Denyer, M. Antigenic properties of some equine influenza viruses. Arch. Virol. 1982, 73, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Mumford, J. Progress in the control of equine influenza. In Equine Infectious Diseases VI; Plowright, W., Rossdale, P.D., Wade, J.F., Eds.; R & W Publications (Newmarket) Limited: Cambridge, UK, 1992; pp. 207–218. [Google Scholar]
- Davenport, F.M.; Hennessy, A.V.; Francis, T., Jr. Epidemiologic and immunologic significance of age distribution of antibody to antigenic variants of influenza virus. J. Exp. Med. 1953, 98, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, M.; Zheng, G.S.; Li, W.K.; Kawaoka, Y.; Webster, R.G. Seroepidemiological and molecular evidence for the presence of two H3N8 equine influenza viruses in China in 1993–94. J. Gen. Virol. 1995, 76, 2009–2014. [Google Scholar] [CrossRef] [PubMed]
- Goto, H.; Shimizu, K.; Taya, Y.; Noda, H.; Tokunaga, T. Antibody responses of horses to equine influenza viruses during a postepizootic period in Japan. Can. J. Comp. Med. 1982, 46, 27–32. [Google Scholar] [PubMed]
- Eman, E.M.; Zeinab, S.T.; A-I.S., H. Immune response of pregnant mares and their foals for inactitatved equine influenza vaccine. Int. J. Virol. 2011, 7, 210–214. [Google Scholar] [CrossRef]
- Minke, J.M.; Toulemonde, C.E.; Dinic, S.; Cozette, V.; Cullinane, A.; Audonnet, J.C. Effective priming of foals born to immune dams against influenza by a canarypox-vectored recombinant influenza H3N8 vaccine. J. Comp. Pathol. 2007, 137, S76–S80. [Google Scholar]
- Horohov, D.W.; Adams, A.A.; Chambers, T.M. Immunosenescence of the equine immune system. J. Comp. Pathol. 2010, 142, S78–S84. [Google Scholar]
- Muirhead, T.L.; McClure, J.T.; Wichtel, J.J.; Stryhn, H.; Frederick Markham, R.J.; McFarlane, D.; Lunn, D.P. The effect of age on serum antibody titers after rabies and influenza vaccination in healthy horses. J. Vet. Intern. Med. 2008, 22, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Horohov, D.W.; Dimock, A.; Guirnalda, P.; Folsom, R.W.; McKeever, K.H.; Malinowski, K. Effect of exercise on the immune response of young and old horses. Am. J. Vet. Res. 1999, 60, 643–647. [Google Scholar] [PubMed]
- McFarlane, D.; Sellon, D.C.; Gibbs, S.A. Age-related quantitative alterations in lymphocyte subsets and immunoglobulin isotypes in healthy horses. Am. J. Vet. Res. 2001, 62, 1413–1417. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paillot, R. A Systematic Review of Recent Advances in Equine Influenza Vaccination. Vaccines 2014, 2, 797-831. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines2040797
Paillot R. A Systematic Review of Recent Advances in Equine Influenza Vaccination. Vaccines. 2014; 2(4):797-831. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines2040797
Chicago/Turabian StylePaillot, Romain. 2014. "A Systematic Review of Recent Advances in Equine Influenza Vaccination" Vaccines 2, no. 4: 797-831. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines2040797