Evaluating Genotypes and Seed Treatments to Increase Field Emergence of Low Phytic Acid Soybeans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Plot Design and Trait Measurement
2.3. Seed Treatments
2.4. Statistical Analysis
3. Results
3.1. Effects of Genetic and Environmental Factors on Field Emergence
3.2. General Effects of Seed Treatments on Field Emergence
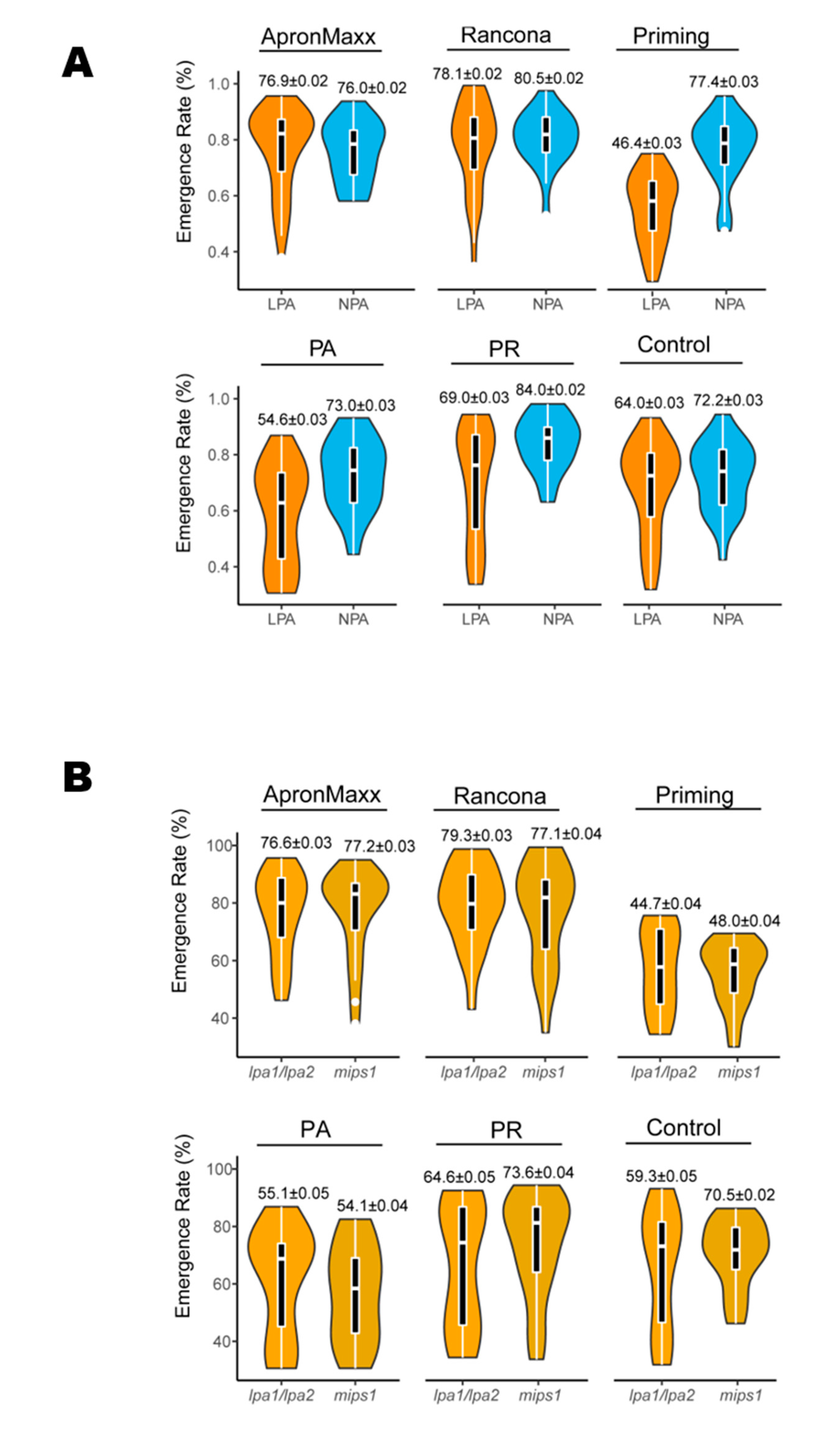
3.3. Effects of Seed Treatments on Field Emergence by PA Phenotype
3.4. Effect of Seed Treatments on Field Emergence of LPA and NPA Genotypes
3.5. Effect of Treatments, Genotypes, and Treatment × Line Interactions on Yield
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilcox, J.R.; Premachandra, G.S.; Young, K.A.; Raboy, V. Isolation of high seed inorganic P, low-phytate soybean mutants. Crop Sci. 2000, 40, 1601–1605. [Google Scholar] [CrossRef] [Green Version]
- Boehm, J.D.; Walker, F.R.; Bhandari, H.S.; Kopsell, D.; Pantalone, V.R. Seed inorganic phosphorus stability and agronomic performance of two low-phytate soybean lines evaluated across six southeastern US environments. Crop Sci. 2017, 57, 2555–2563. [Google Scholar] [CrossRef]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [Green Version]
- Sinkko, H.; Lukkari, K.; Sihvonen, L.M.; Sivonen, K.; Leivuori, M.; Rantanen, M.; Paulin, L.; Lyra, C. Bacteria contribute to sediment nutrient release and reflect progressed eutrophication-driven hypoxia in an organic-rich continental sea. PLoS ONE 2013, 8, e67061. [Google Scholar] [CrossRef] [Green Version]
- Averitt, B.; Shang, C.; Rosso, L.; Qin, J.; Zhang, M.; Rainy, K.M.; Zhang, B. Impact of mips1, lpa1, and lpa2 alleles for low phytic acid content on agronomic, seed quality, and seed composition traits of soybean. Crop Sci. 2017, 57, 2490–2499. [Google Scholar] [CrossRef]
- Gillman, J.D.; Baxte, I.; Bilyeu, K. Phosphorus partitioning of soybean lines containing different mutant alleles of two soybean seed-specific adenosine triphosphate-binding cassette phytic acid transporter paralogs. Plant Genome 2013, 6, 1–10. [Google Scholar] [CrossRef]
- Hegeman, C.E.; Good, L.L.; Grabau, E.A. Expression of D-myo-inositol-3-phosphate synthase in soybean. Implications for phytic acid biosynthesis. Plant Physiol. 2001, 125, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Redekar, N.; Pilot, G.; Raboy, V.; Li, S.; Saghai Maroof, M.A. Inference of transcription regulatory network in low phytic acid soybean seeds. Front. Plant Sci. 2017, 8, 2029. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Yu, X.; Dong, D.; Yang, Q.; Fu, X.; Zhu, S.; Zhu, D. Whole genome-wide transcript profiling to identify differentially expressed genes associated with seed field emergence in two soybean low phytate mutants. BMC Plant Biol. 2017, 17, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Jin, H.; Fu, X.; Yang, Q.; Yuan, F. Quantitative proteomic analyses of two soybean low phytic acid mutants to identify the genes associated with seed field emergence. BMC Plant Biol. 2019, 19. [Google Scholar] [CrossRef]
- Anderson, B.P.; Fehr, W.R. Seed source affects field emergence of low-phytate soybean lines. Crop Sci. 2008, 48, 929–932. [Google Scholar] [CrossRef]
- Maupin, L.M.; Rainey, K.M. Improving emergence of modified phosphorus composition soybeans: Genotypes, germplasm, environments, and selection. Crop Sci. 2011, 51, 1946–1955. [Google Scholar] [CrossRef]
- Meis, S.J.; Fehr, W.R.; Schnebly, S.R. Seed source effect on field emergence of soybean lines with reduced phytate and raffinose saccharides. Crop Sci. 2003, 43, 1336–1339. [Google Scholar] [CrossRef]
- Punjabi, M.; Bharadvaja, N.; Jolly, M.; Dahuja, A.; Sachdev, A. Development and evaluation of low phytic acid soybean by siRNA triggered seed specific silencing of inositol polyphosphate 6-/3-/5-kinase gene. Front. Plant Sci. 2018, 9, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akamatsu, H.; Kato, M.; Ochi, S.; Mimuro, G.; Matsuoka, J.I.; Takahashi, M. Variation in the resistance of japanese soybean cultivars to phytophthora root and stem rot during the early plant growth stages and the effects of a fungicide seed treatment. Plant Pathol. J. 2019, 35, 219–233. [Google Scholar] [CrossRef]
- Costa, E.M.; Nunes, B.M.; Ventura, M.V.A.; Mortate, R.K.; Vilarinho, M.S.; Da Silva, R.M.; Chagas, J.F.R.; Nogueira, L.C.A.; Arantes, B.H.T.; Lima, A.P.A.; et al. Physiological effects of insecticides and fungicide, applied in the treatment of seeds, on the germination and vigor of soybean seeds. J. Agric. Sci. 2019, 11. [Google Scholar] [CrossRef]
- Rouhi, H.R.; Surki, A.A.; Sharif-Zadeh, F.; Afshari, R.T.; Aboutalebian, M.A.; Ahmadvand, G. Study of different priming treatments on germination traits of soybean seed lots. Not. Sci. Biol. 2011, 3, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Welbaum, G.E.; Shen, Z.; Oluoch, M.O.; Jett, L.W. The evolution and effects of priming vegetable seeds. Seed Technol. 1998, 20, 209–235. Available online: http://0-www-jstor-org.brum.beds.ac.uk/stable/23433024 (accessed on 24 May 2017).
- Farooq, M.; Wahid, A.; Siddique, K.H.M. Micro-nutrient application through seed treatments—A review. J. Soil Sci. Plant Nutr. 2012, 12, 125–142. [Google Scholar] [CrossRef]
- Sharma, K.K.; Singh, U.S.; Sharma, P.; Kumar, A.; Sharma, L. Seed treatments for sustainable agriculture—A review. J. Appl. Nat. Sci. 2015, 7, 521–539. [Google Scholar] [CrossRef]
- Xue, A.G.; Cober, E.; Morrison, M.J.; Voldeng, H.D.; Ma, B.L. Effect of seed treatments on emergence, yield, and root rot severity of soybean under Rhizoctonia solani inoculated field conditions in Ontario. Can. J. Plant Sci. 2007, 87, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Nyandoro, R.; Chang, K.F.; Hwang, S.F.; Ahmed, H.U.; Turnbull, G.D.; Strelkov, S.E. Management of root rot of soybean in Alberta with fungicide seed treatments and genetic resistance. Can. J. Plant Sci. 2019, 99, 499–509. [Google Scholar] [CrossRef]
- Pantalone, V.R.; Allen, F.; Landau-Ellis, D. Registration of ‘5002T’ soybean. Crop Sci. 2004, 44, 1483–1484. [Google Scholar] [CrossRef]
- Hayter Series. Available online: https://soilseries.sc.egov.usda.gov/OSD_Docs/H/HAYTER.html (accessed on 19 October 2020).
- Fehr, W.R.; Caviness, C.E. Stages of Soybean Development; Special Report 80; Iowa State University: Ames, IA, USA, 1977. [Google Scholar]
- Burleson, S.A.; Shang, C.; Luciana Rosso, M.; Maupin, L.M.; Rainey, K.M. A modified colorimetric method for selection of soybean phytate concentration. Crop Sci. 2012, 52, 122–127. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “corrplot”: Visualization of a Correlation Matrix (Version 0.84). 2017. Available online: https://github.com/taiyun/corrplot (accessed on 18 June 2019).
- Sarkar, D. Lattice: Multivariate Data Visualization with R; Springer: New York, NY, USA, 2008. [Google Scholar]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 2nd ed.; Sage Publishing: Thousand Oaks, CA, USA, 2011; Available online: http://socserv.socsci.mcmaster.ca/jfox/Books/Companion (accessed on 18 June 2019).
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R package Version 1.3-0. 2019. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 18 June 2019).
- Raboy, V. Seeds for a better future:’low phytate’grains help to overcome malnutrition and reduce pollution. Trends Plant Sci. 2001, 6, 458–462. [Google Scholar] [CrossRef]
- Bregitzer, P.; Raboy, V. Effects of four independent low-phytate mutations on barley agronomic performance. Crop Sci. 2006, 46, 1318–1322. [Google Scholar] [CrossRef]
- Bilyeu, K.D.; Zeng, P.; Coello, P.; Zhang, Z.J.; Krishnan, H.B.; Bailey, A.; Beuselinck, P.R.; Polacco, J.C.; Department, B. Quantitative conversion of phytate to inorganic phosphorus in soybean seeds expressing a bacterial phytase. Am. Soc. Plant Biol. 2008, 146, 468–477. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Wang, H.; Schellin, K.; Li, B.; Faller, M.; Stoop, J.M.; Meeley, R.B.; Ertl, D.S.; Ranch, J.P.; Glassman, K. Embryo-specific silencing of a transporter reduces phytic acid content of maize and soybean seeds. Nat. Biotechnol. 2007, 25, 930–937. [Google Scholar] [CrossRef]
- Lueschen, W.E.; Hicks, D.R. Influence of plant population on field performance of three soybean cultivars. Agron. J. 1977, 69, 390–393. [Google Scholar] [CrossRef]
- Franca Neto, J.B.; Krzyzanowski, F.C.; Henning, A.A.; West, S.H.; Miranda, L.C. Soybean seed quality as aff ected by shriveling due to heat and drought stress during seed filling. Seed Sci. Technol. 1993, 21, 107–116. [Google Scholar]
- Spears, J.; TeKrony, D.; Egli, D.B. Temperature during seed filling and soybean seed germination and vigor. Seed Sci. Technol. 1997, 25, 233–244. [Google Scholar]
- Vieira, R.D.; TeKrony, D.M.; Egli, D.B. Effect of drought and defoliation stress in the field on soybean seed germination and vigor. Crop Sci. 1992, 32, 471–475. [Google Scholar] [CrossRef]
- Vieira, R.; TeKrony, D.; Egli, D.B. Effect of drought stress on soybean seed germination and vigor. Seed Sci. Technol. 1991, 15, 12–21. [Google Scholar]
- McDonald, M.B. Physical seed quality of soybean. Seed Sci. Technol. 1985, 13, 601–628. [Google Scholar]
- Mugnisjah, W.; Nakamura, S. Vigour of soybean seed produced from different harvest date and phosphorus fertiliser application. Seed Sci. Technol. 1984, 12, 483–491. [Google Scholar]
- Miles, D.F.; TeKrony, D.M.; Egli, D.B. Changes in viability, germination, and respiration of freshly harvested soybean seed during development. Crop Sci. 1988, 28, 700–704. [Google Scholar] [CrossRef]
- Chamma, H.; Filho, J.M.; Crocomo, O.J. Maturation of seeds of “Aroana” beans (Phaseolus vulgaris L.) and its influence on the storage potential. Seed Sci. Technol. 1990, 18, 371–382. [Google Scholar]
- Rodriguez, A.; McDonald, M.B. Seed quality influence on plant growth and dinitrogen fixation of red field bean. Crop Sci. 1989, 29, 1309–1314. [Google Scholar] [CrossRef]
- Chauhan, K. The incidence of deterioration and its localisation in aged seeds of soybean and barley. Seed Sci. Technol. 1985, 13, 769–773. [Google Scholar]
- Hoy, D.J.; Gamble, E.E. Field performance in soybean with seeds of differing size and density. Crop Sci. 1987, 27, 121–126. [Google Scholar] [CrossRef]
- Hoy, D.J.; Gamble, E.E. The effects of seed size and seed density on germination and vigor in soybean (Glycine max (L.) Merr.). Can. J. Plant Sci. 1985, 65, 1–8. [Google Scholar] [CrossRef]
- Egli, D.B.; TeKrony, D.M.; Wiralaga, R.A. Effect of soybean seed vigor and seed size on soybean seedling growth. Seed Sci. Technol. 1990, 14, 1–12. [Google Scholar]
- Elias, S.G.; Copeland, L.O.; McDonald, M.B.; Baalbaki, R.Z. Seed Testing: Principles and Practices; Michigan State University Press: East Lansing, MI, USA, 2012. [Google Scholar]
- Veresoglou, S.D.; Barto, E.K.; Menexes, G.; Rillig, M.C. Fertilization affects severity of disease caused by fungal plant pathogens. Plant Pathol. 2013, 62, 961–969. [Google Scholar] [CrossRef]
- Douglass, S.K.; Juvik, J.A.; Splittstoesser, W.E. Sweet corn seedling emergence and variation in kernel carbohydrate reserves. Seed Sci. Technol. 1993, 21, 433–445. [Google Scholar]
- Mushtaq, S.; Hafiz, A.I.; Hasan, S.Z.; Arif, M.; Shehzad, M.A.; Rafique, R.; Rasheed, M.; Ali, M.; Iqbal, M.S. Evaluation of seed priming on germination of Gladiolus alatus. Afr. J. Biotechnol. 2012, 11, 11520–11523. [Google Scholar] [CrossRef]
- Kering, M.; Zhang, B. Effect of priming and seed size on germination and emergence of six food-type soybean varieties. Int. J. Agron. 2015, 1, 1–6. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Genotype | LPA Gene | Years Planted | PA Content (ppm) |
|---|---|---|---|
| 5002T | N/A | 2014, 2015 | 6116.10 |
| AG 5632 | N/A | 2014, 2015 | 5886.72 |
| 56CX-1283 | lpa1/lpa2 | 2014, 2015 | 2486.03 |
| MD 03-5453 | lpa1/lpa2 | 2015 | 2131.68 |
| V12-4557 | mips1 | 2015 | 4060.80 |
| V12-BB144 | mips1 | 2014, 2015 | 4420.50 |
| Treatment | Years Used | Use |
|---|---|---|
| Control | 2014, 2015 | Untreated control |
| ApronMaxx | 2014, 2015 | Broad spectrum fungicide |
| MicroCel-E | 2014 | Weak fertilizer |
| Priming | 2014, 2015 | Post-harvest preplant controlled hydration treatment followed by redrying prior to planting |
| Rancona Summit | 2014, 2015 | Broad spectrum fungicide |
| Priming + Rancona | 2014, 2015 | |
| Priming + ApronMaxx | 2014, 2015 | |
| Priming + MicroCel-E | 2014 | |
| Priming + MicroCel-E + Rancona | 2014 | |
| Priming + MicroCel-E + ApronMaxx | 2014 | |
| MicroCel-E + Rancona | 2014 | |
| MicroCel-E + ApronMaxx | 2014 |
| Line | Irrigation | Phytate | Emergence % | |||||
|---|---|---|---|---|---|---|---|---|
| 2014 Only | 2015 Only | 2014 BB 1 | 2014 O | 2015 BB | 2015 O | |||
| 5002T | I 2 | Normal | 64.6 * | 71.7 * | 65.4d | 71.4c | 76.5bc | 74.7cd |
| N | 72.1 | 79.5 | ||||||
| Mean | 68.4c 3 | 75.6ab | ||||||
| AG-5632 | I | Normal | 79.2 * | 77.4 | 76.8b | 88.4a | 85.5a | 72.2cd |
| N | 86.0 | 79.9 | ||||||
| Mean | 82.6a | 78.7a | ||||||
| 56CX-1283 | I | lpa1/lpa2 | 69.7 * | 79.2 | 67.6d | 76.5b | 82.4ab | 76.7bc |
| N | 74.4 | 79.9 | ||||||
| Mean | 72.1b | 79.5a | ||||||
| V12-BB144 | I | mips1 | 66.0 * | 56.5 * | 65.2d | 75.3b | 77.1bc | 46.8f |
| N | 74.6 | 66.9 | ||||||
| Mean | 70.3b | 61.8c | ||||||
| MD 03-5453 | I | lpa1/lpa2 | - | 44.8 | - | - | 54.0e | 39.6g |
| N | - | 48.7 | ||||||
| Mean | - | 46.7d | ||||||
| V12-4557 | I | mips1 | - | 68.1 * | - | - | 75.8bc | 68.2d |
| N | - | 76.0 | ||||||
| Mean | - | 71.9b | ||||||
| Mean | 73.3 | 69.1 | 68.8 | 77.9 | 75.2 | 63.0 | ||
| Treatment | Irrigation | Emergence % | |||||
|---|---|---|---|---|---|---|---|
| 2014 Only | 2015 Only | 2014 BB 1 | 2014 O | 2015 BB | 2015 O | ||
| Control (untreated) | I 2 | 77.8 | 64.4 | 74.0fghij | 86.3a | 71.0b | 63.8cd |
| N | 82.6 | 70.3 | |||||
| Mean | 80.2ab 4 | 67.4c | |||||
| ApronMaxx | I | 77.9 * | 73.7 | 78.3cdefg | 85.5ab | 83.4a | 69.8bc |
| N | 85.8 | 79.5 | |||||
| Mean | 81.9a | 76.6ab | |||||
| Rancona | I | 78.6 * | 76.2 | 76.8defgh | 87.4a | 86.3a | 72.0b |
| N | 85.6 | 81.7 | |||||
| Mean | 82.1a | 79.0a | |||||
| Microcel | I | 72.2 | - | 69.7ijk | 79.2bcdef | - | - |
| N | 76.7 | - | |||||
| Mean | 74.5c | - | |||||
| Priming | I | 63.7 * | 55.7 | 63.4lmn | 73.3fghijk | 62.6d | 52.0e |
| N | 73.0 | 58.8 | |||||
| Mean | 68.4de | 57.3d | |||||
| MxA 3 | I | 76.6 | - | 74.9efghi | 81.8abcd | - | - |
| N | 80.2 | - | |||||
| Mean | 78.4abc | - | |||||
| MxR | I | 73.0 * | - | 70.2ijk | 83.5abc | - | - |
| N | 80.7 | - | |||||
| Mean | 76.8bc | - | |||||
| PA | I | 61.0 | 58.0 | 56.9o | 68.0jklm | 68.9bcd | 52.6e |
| N | 63.9 | 63.5 | |||||
| Mean | 62.4f | 60.7d | |||||
| PR | I | 71.5 * | 70.3 * | 71.1hijk | 81.2abcde | 79.5a | 68.5bcd |
| N | 81.3 | 77.4 | |||||
| Mean | 76.3bc | 73.8b | |||||
| PM | I | 60.2 * | - | 62.2mno | 68.9ijkl | - | - |
| N | 70.9 | - | |||||
| Mean | 65.5ef | - | |||||
| PxMxA | I | 59.3 * | - | 59.8no | 67.6klm | - | - |
| N | 68.1 | - | |||||
| Mean | 63.7f | - | |||||
| PxMxR | I | 66.8 | - | 67.9jklm | 71.9ghijk | - | - |
| N | 73.0 | - | |||||
| Mean | 69.9d | - | |||||
| Mean | 73.3 | 69.1 | 68.8 | 77.9 | 75.3 | 63.1 | |
| 2014 | ||||||||||||||
| Line | Phytate | Emergence % | ||||||||||||
| C 1 | A | R | M | P | MxA | MxR | PA | PR | PM | PxMxA | PxMxR | Mean | ||
| 5002T | NPA 2 | 77.6 | 84.4 | 81.3 | 70.4 | 66.0 * | 72.1 | 75 | 53.3 * | 70.7 | 51.5 * | 50.8 * | 67.1 * | 68.4c 3 |
| AG-5632 | NPA | 81.8 | 87.3 | 87.3 | 82 | 79.4 | 87.7 | 82.5 | 80.3 | 84.5 | 80 | 77.6 | 80.8 | 82.6a |
| 56CX-1283 | LPA | 85.3 | 79.9 | 83.6 | 73.9 * | 69.4 * | 76.4 | 77.9 | 48.4 * | 79.1 | 71.0 * | 58.4 * | 61.3 * | 72.1b |
| V12-BB144 | LPA | 75.9 | 75.9 | 76.1 | 71.6 | 58.5 * | 77.3 | 72 | 67.7 | 70.2 | 59.6 * | 68 | 70.5 | 70.3b |
| Mean | - | 80.2ab | 81.9a | 82.1a | 74.5c | 68.4de | 78.4abc | 76.8bc | 62.4f | 76.3bc | 65.5ef | 63.7f | 69.9d | - |
| 2015 | ||||||||||||||
| 5002T | NPA | 65.1 | 77.8 | 78.8 * | 76.3 | 71.6 | 84.0 * | 75.6ab | ||||||
| AG-5632 | NPA | 79.3 | 74.3 | 82.3 | 78.6 | 74.3 | 84.1 | 78.7a | ||||||
| 56CX-1283 | LPA | 80.4 | 85.4 | 88.6 | 63.2 * | 74.6 | 85.1 | 79.5a | ||||||
| V12-BB144 | LPA | 67.9 | 72.5 | 71.6 | 41.3 * | 47.9 * | 69 | 61.8c | ||||||
| MD 03-5453 | LPA | 36.3 | 67.7 * | 69.5 * | 26.2 | 35.5 | 44.1 | 46.7d | ||||||
| V12-4557 | LPA | 72.9 | 81.8 | 82.6 | 56.7 * | 60.4 | 78.6 | 71.9b | ||||||
| Mean | - | 67.4c | 76.6ab | 79.0a | 57.3d | 60.7d | 73.8b | - | ||||||
| 2014 | ||||||||||||||
| Line | Phytate | Yield kg ha−1 | ||||||||||||
| C 1 | A | R | M | P | MxA | MxR | PA | PR | PM | PxMxA | PxMxR | Mean | ||
| 5002T | NPA 2 | 4782 | 4983 | 4909 | 4782 | 4459 | 4842 | 5057 | 4815 | 4701 | 4519 | 4573 | 4768 | 4768b 3 |
| AG-5632 | NPA | 4936 | 5151 | 5151 | 5091 | 4956 | 5077 | 5387 | 5185 | 5219 | 5185 | 5151 | 5205 | 5144a |
| 56CX-1283 | LPA | 4882 | 4882 | 4922 | 4829 | 4882 | 4936 | 5010 | 4546 | 5098 | 4808 | 4707 | 4721 | 4849b |
| V12-BB144 | LPA | 4304 | 4431 | 4566 | 4734 | 4465 | 4425 | 4492 | 4580 | 4526 | 4156 | 4539 | 4499 | 4479c |
| Mean | - | 4728ab | 4862ab | 4889ab | 4856ab | 4687ab | 4822ab | 4990a | 4782ab | 4896ab | 4667b | 4741ab | 4795ab | - |
| 2015 | ||||||||||||||
| 5002T | NPA | 3477 | 3618 | 3551 | 3558 | 3564 | 3511 | 3544b | ||||||
| AG-5632 | NPA | 4284 | 3827 | 3867 | 4270 | 3867 | 3901 | 4008a | ||||||
| 56CX-1283 | LPA | 4001 | 3921 | 4304 | 4479 | 4102 | 4055 | 4143a | ||||||
| V12-BB144 | LPA | 3685 | 3894 | 3732 | 3141 | 3268 | 3470 | 3531b | ||||||
| MD 03-5453 | LPA | 1110 | 1506 | 1076 | 659 | 1446 | 1439 | 1211c | ||||||
| V12-4557 | LPA | 3571 | 3800 | 3531 | 3477 | 3443 | 3289 | 3517b | ||||||
| Mean | - | 3416a | 3430a | 3336a | 3302a | 3309a | 3255a | - | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Averitt, B.J.; Welbaum, G.E.; Li, X.; Prenger, E.; Qin, J.; Zhang, B. Evaluating Genotypes and Seed Treatments to Increase Field Emergence of Low Phytic Acid Soybeans. Agriculture 2020, 10, 516. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10110516
Averitt BJ, Welbaum GE, Li X, Prenger E, Qin J, Zhang B. Evaluating Genotypes and Seed Treatments to Increase Field Emergence of Low Phytic Acid Soybeans. Agriculture. 2020; 10(11):516. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10110516
Chicago/Turabian StyleAveritt, Benjamin J., Gregory E. Welbaum, Xiaoying Li, Elizabeth Prenger, Jun Qin, and Bo Zhang. 2020. "Evaluating Genotypes and Seed Treatments to Increase Field Emergence of Low Phytic Acid Soybeans" Agriculture 10, no. 11: 516. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10110516