
Search for Candidate Genes Causing the Excessive Ca Accumulation in Roots of Tipburn-Damaged Lisianthus (Eustoma grandiflorum) Cultivars

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Extraction and RNA-Seq
2.3. Bioinformatics Analysis
3. Results
3.1. De Novo Transcriptome Assembly
3.2. Functional Annotation
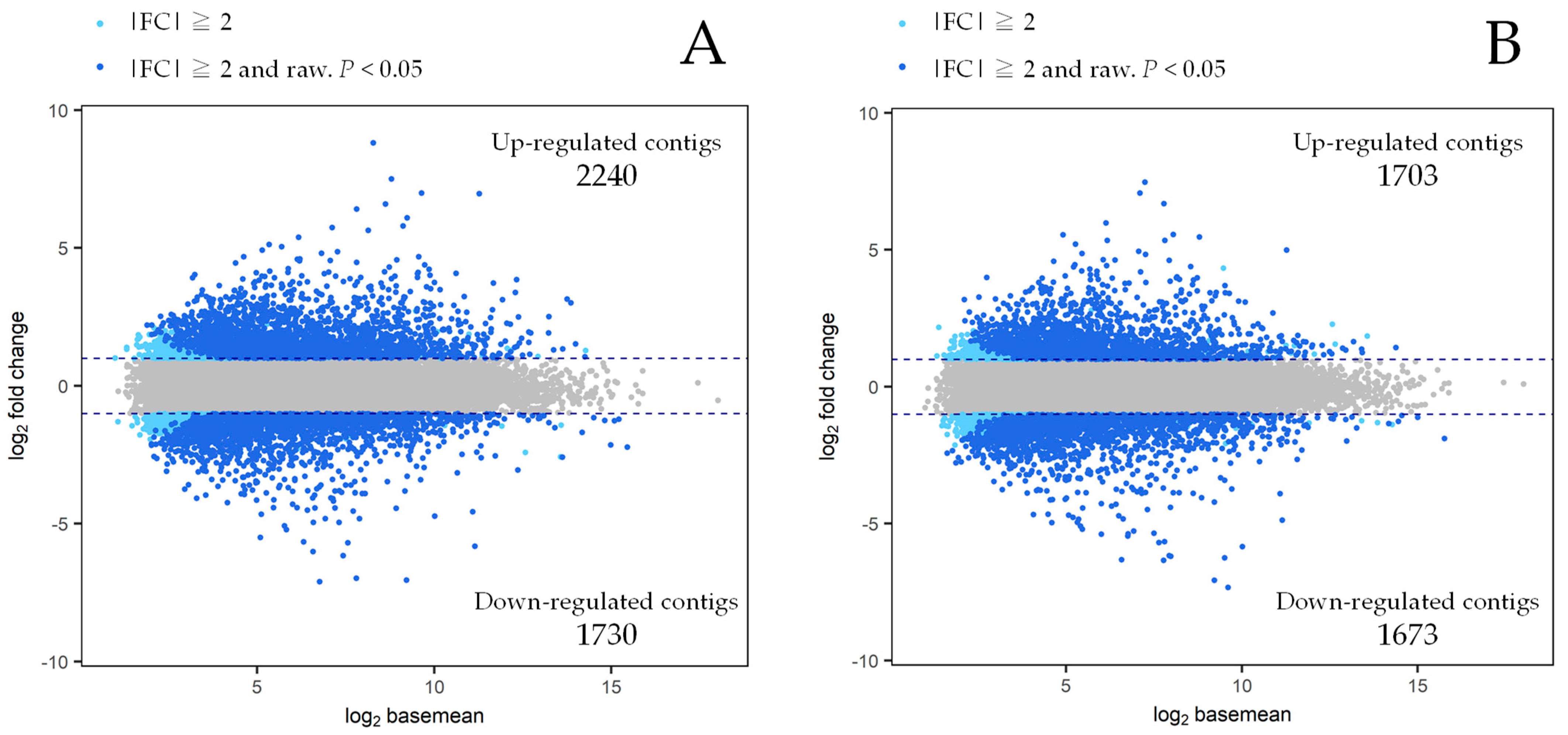
3.3. Differentially Expressed Genes (DEGs)
4. Discussion
4.1. Candidate Genes Causing the Difference of Ca Accumulation in Cultivars
4.2. Candidate Genes Causing Excessive Ca Accumulation in Roots of VP
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling: An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Navazio, L.; Szabo, I. The contribution of organelles to plant intracellular calcium signalling. J. Exp. Bot. 2018, 69, 4175–4193. [Google Scholar] [CrossRef] [Green Version]
- Hagassou, D.; Francia, E.; Ronga, D.; Buti, M. Blossom end-rot in tomato (Solanum lycopersicum L.): A multi-disciplinary overview of inducing factors and control strategies. Sci. Hortic. 2019, 249, 49–58. [Google Scholar] [CrossRef]
- Sago, Y. Effects of light intensity and growth rate on tipburn development and leaf calcium concentration in butterhead lettuce. HortScience 2016, 51, 1087–1091. [Google Scholar] [CrossRef]
- Hernandez, J.; Soriano, T.; Morales, M.I.; Castilla, N. Row covers for quality improvement of Chinese cabbage (Brassica rapa subsp. pekinensis). N. Z. J. Crop Hortic. Sci. 2004, 32, 379–388. [Google Scholar] [CrossRef]
- Barta, D.J.; Tibbitts, T.W. Calcium localization and tipburn development in lettuce leaves during early enlargement. J. Am. Soc. Hortic. Sci. 2000, 125, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.G.; Tsay, J.S.; Tsai, C.L.; Chen, R.J. Tipburn of Chinese cabbage in relation to calcium nutrition and distribution. Sci. Hortic. 1981, 14, 131–138. [Google Scholar] [CrossRef]
- Olle, M.; Bender, I. Causes and control of calcium deficiency disorders in vegetables: A review. J. Hortic. Sci. Biotrch. 2009, 84, 577–584. [Google Scholar] [CrossRef]
- Kuronuma, T.; Watanabe, Y.; Ando, M.; Watanabe, H. Relevance of tipburn incidence to the competence for Ca acquirement and Ca distributivity in lisianthus [Eustoma grandiflorum (Raf.) Shinn.] cultivars. Sci. Hortic. 2019, 246, 805–811. [Google Scholar] [CrossRef]
- Kuronuma, T.; Kinoshita, N.; Ando, M.; Watanabe, H. Difference of Ca distribution before and after the onset of tipburn in lisianthus [Eustoma grandiflorum (Raf.) Shinn.] cultivars. Sci. Hortic. 2020, 261. [Google Scholar] [CrossRef]
- Kuronuma, T.; Ando, M.; Watanabe, H. Tipburn Incidence and Ca acquisition and distribution in lisianthus (Eustoma grandiflorum (Raf.) Shinn.) cultivars under different Ca concentrations in nutrient solution. Agronomy 2020, 10, 216. [Google Scholar] [CrossRef] [Green Version]
- Kuronuma, T.; Saotome, M.; Ando, M.; Watanabe, H. Excessive Calcium Accumulation in the Roots Is a Key Factor in Tipburn Incidence under High Ca Supply in Lisianthus (Eustoma grandiflorum) Cultivars. Agronomy 2020, 10, 1123. [Google Scholar] [CrossRef]
- Lee, J.; Kim, J.; Choi, J.P.; Lee, M.; Kim, M.K.; Lee, Y.H.; Hur, Y.; Nou, S.; Park, S.U.; Min, S.R.; et al. Intracellular Ca 2+ and K+ concentration in Brassica oleracea leaf induces differential expression of transporter and stress-related genes. BMC Genom. 2016, 17, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.; Li, P.; Wang, H.; Wang, W.; Zhao, X.; Yu, Y.; Zhang, D.; Yu, S.; Zhang, F. Natural variation in a calreticulin gene causes reduced resistance to Ca2+ deficiency-induced tipburn in Chinese cabbage (Brassica rapa ssp. pekinensis). Plant Cell Environ. 2019, 42, 3044–3060. [Google Scholar] [CrossRef]
- Seki, K.; Komatsu, K.; Tanaka, K.; Hiraga, M.; Kajiya-Kanegae, H.; Matsumura, H.; Uno, Y. A CIN-like TCP transcription factor (LsTCP4) having retrotransposon insertion associates with a shift from Salinas type to Empire type in crisphead lettuce (Lactuca sativa L.). Hortic. Res. 2020, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conn, S.J.; Gilliham, M.; Athman, A.; Schreiber, A.W.; Baumann, U.; Moller, I.; Cheng, N.H.; Stancombe, M.A.; Hirschi, K.D.; Webb, A.A.R.; et al. Cell-specific vacuolar calcium storage mediated by CAX1 regulates apoplastic calcium concentration, gas exchange, and plant productivity in Arabidopsis. Plant Cell 2011, 23, 240–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Cheng, N.H.; Pittman, J.K.; Yoo, K.S.; Park, J.; Smith, R.H.; Hirschi, K.D. Increased calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol. 2005, 139, 1194–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, S.T.; Padda, M.; Wu, Q.; Park, S.; Mitcham, E.J. Dynamic alternations in cellular and molecular components during blossom-end rot development in tomatoes expressing sCAX1, a constitutively active Ca2+/H+ antiporter from Arabidopsis. Plant Physiol. 2011, 156, 844–855. [Google Scholar] [CrossRef] [Green Version]
- Zorrilla, C.; Schabow, J.E.; Chernov, V.; Palta, J.P. CAX1 vacuolar antiporter overexpression in potato results in calcium deficiency in leaves and tubers by sequestering calcium as calcium oxalate. Crop Sci. 2019, 59, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Wu, X.; Zorrilla, C.; Vega, S.E.; Palta, J.P. Fractionating of calcium in tuber and leaf tissues explains the calcium deficiency symptoms in potato plant overexpressing CAX1. Front. Plant Sci. 2020, 10, 1793. [Google Scholar] [CrossRef]
- Wormit, A.; Usadel, B. The multifaceted role of pectin methylesterase inhibitors (PMEIs). Int. J. Mol. Sci. 2018, 19, 2878. [Google Scholar] [CrossRef] [Green Version]
- Freitas, S.T.; Handa, A.K.; Wu, Q.; Park, S.; Mitcham, E.J. Role of pectin methylesterases in cellular calcium distribution and blossom-end rot development in tomato fruit. Plant J. 2012, 71, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.M.; Mäser, P.; Schroeder, J.I. Plant ion channels: Gene families, physiology, and functional genomics analyses. Ann. Rev. Physiol. 2009, 71, 59–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarbreck, S.M.; Colaço, R.; Davies, J.M. Plant calcium-permeable channels. Plant Physiol. 2013, 163, 514–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García Bossi, J.; Kumar, K.; Barberini, M.L.; Domínguez, G.D.; Rondón Guerrero, Y.D.C.; Marino-Buslje, C.; Obertello, M.; Muschietti, J.P.; Estevez, J.M. The role of P-type IIA and P-type IIB Ca2+-ATPases in plant development and growth. J. Exp. Bot. 2020, 71, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Taneja, M.; Tyagi, S.; Sharma, S.; Upadhyay, S.K. Ca2+/cation antiporters (CaCA): Identification, characterization and expression profiling in bread wheat (Triticum aestivum L.). Front. Plant Sci. 2016, 7, 1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amagaya, K.; Shibuya, T.; Nishiyama, M.; Kato, K.; Kanayama, Y. Characterization and Expression Analysis of the Ca2+/Cation Antiporter Gene Family in Tomatoes. Plants 2020, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimberley, J.; Cahill, J. De novo Sequencing and Analysis of Salvia hispanica Tissue-Specific Transcriptome and Identification of Genes Involved in Terpenoid Biosynthesis. Plants 2020, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Cechin, J.; Piasecki, C.; Benemann, D.P.; Kremer, F.S.; Galli, V.; Maia, L.C.; Agostinetto, D.; Vargas, L. Transcriptome Analysis Identifies Candidate Target Genes Involved in Glyphosate-Resistance Mechanism in Lolium multiflorum. Plants 2020, 9, 685. [Google Scholar] [CrossRef]
- Qi, Z.; Stephens, N.R.; Spalding, E.P. Calcium entry mediated by GLR3.3, an Arabidopsis glutamate receptor with a broad agonist profile. Plant Physiol. 2006, 142, 963–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.A.; Kwak, J.; Jae, S.K.; Wang, M.H.; Nam, H. Overexpression of the AtGluR2 Gene Encoding an Arabidopsis Homolog of Mammalian Glutamate Receptors Impairs Calcium Utilization and Sensitivity to Ionic Stress in Transgenic Plants. Plant Cell Physiol. 2001, 42, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.; Tian, H.; Park, S.; Sreevidya, C.S.; Ward, J.M.; Hirschi, K.D. AtCCX3 is an Arabidopsis endomembrane H+-dependent K+ transporter. Plant Physiol. 2008, 148, 1474–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corso, M.; Doccula, F.G.; de Melo, J.R.F.; Costa, A.; Verbruggen, N. Endoplasmic reticulum-localized CCX2 is required for osmotolerance by regulating ER and cytosolic Ca2+ dynamics in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 3966–3971. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Li, Z.; Wei, J.; Zhao, Z.; Sun, D.; Cui, S. A Na+/Ca2+ exchanger-like protein (AtNCL) involved in salt stress in Arabidopsis. J. Biol. Chem. 2012, 287, 44062–44070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Zhang, G.; Gonzales, N.; Guo, Y.; Hu, H.; Park, S.; Zhao, J. Ca2+-regulated and diurnal rhythm-regulated Na+/Ca2+ exchanger AtNCL affects flowering time and auxin signalling in Arabidopsis. Plant Cell Environ. 2016, 39, 377–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.Q.; Yang, Y.; Zhang, A.; Fei, C.F.; Gu, L.L.; Sun, S.J.; Xu, W.; Wang, L.; Liu, H.; Wang, Y.F. Three CNGC family members, CNGC5, CNGC6, and CNGC9, are required for constitutive growth of Arabidopsis root hairs as Ca2+-permeable channels. Plant Commun. 2020, 1, 100001. [Google Scholar] [CrossRef] [PubMed]
- Edmond, C.; Shigaki, T.; Ewert, S.; Nelson, M.D.; Connorton, J.M.; Chalova, V.; Noordally, Z.; Pittman, J.K. Comparative analysis of CAX2-like cation transporters indicates functional and regulatory diversity. Biochem. J. 2009, 418, 145–154. [Google Scholar] [CrossRef]
- Choe, M.; Choe, W.; Cha, S.; Lee, I. Changes of cationic transport in AtCAX5 transformant yeast by electromagnetic field environments. J. Biol. Phys. 2018, 44, 433–448. [Google Scholar] [CrossRef]
- Huang, L.; Berkelman, T.; Franklin, A.E.; Hoffman, N.E. Characterization of a gene encoding a Ca2+-ATPase-like protein in the plastid envelope. Proc. Natl. Acad. Sci. USA 1993, 90, 10066–10070. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Yan, J.; Du, X.; Hua, J. Overlapping and differential roles of plasma membrane calcium ATPases in Arabidopsis growth and environmental responses. J. Exp. Bot. 2018, 69, 2693–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limonta, M.; Romanowsky, S.; Olivari, C.; Bonza, M.C.; Luoni, L.; Rosenberg, A.; Harper, J.F.; De Michelis, M.I. ACA12 is a deregulated isoform of plasma membrane Ca2+-ATPase of Arabidopsis thaliana. Plant Mol. Biol. 2014, 84, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Cultivars | Ca Treatments | Ca Concentrations of Each Organ (mg-Ca/kg-DW) | Tipburn (%; Values at 8 w) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 w (before the Onset of Tipburn) | 8 w (after the Onset of Tipburn) | ||||||||||||||
| Leaves | Stems | Roots | Leaves | Stems | Roots | Incidence | Severity | ||||||||
| UH | 40 ppm | 2.2 | n.s. | 1.1 | n.s. | 3.9 | n.s. | 3.3 | a | 1.2 | a | 4.0 | n.s. | 0 | 0 |
| 80 ppm | 2.2 | 1.0 | 3.9 | 4.7 | b | 1.6 | b | 4.5 | 0 | 0 | |||||
| 120 ppm | 2.3 | 1.2 | 3.7 | 5.1 | c | 2.0 | c | 4.5 | 0 | 0 | |||||
| VP | 40 ppm | 3.1 | n.s. | 2.2 | n.s. | 6.0 | n.s. | 2.4 | a | 1.1 | a | 9.1 | a | 100 | 25 |
| 80 ppm | 3.3 | 2.7 | 6.1 | 3.3 | b | 1.6 | b | 11.6 | a | 100 | 16 | ||||
| 120 ppm | 3.3 | 2.7 | 6.9 | 4.5 | c | 2.0 | c | 14.9 | b | 100 | 12 | ||||
| Description | Assembly Statics * |
|---|---|
| Total assembled bases | 176,216,995 |
| Total number of assembled transcripts by Trinity | 234,725 |
| Total number of assembled genes by Trinity | 160,301 |
| Contig N50 | 1282 |
| Average contig length (bp) | 750.74 |
| GC (%) | 40.64 |
| Contig ID | Uniprot ID | Functional Annotation-Gene | Putative Localization | Log2 FC VP vs. UH |
|---|---|---|---|---|
| c133995_g1_i7 | Q9SHV1 | GLR2.2 | plasma membrane | 6.03 |
| c133636_g2_i4 | Q8LGN0 | GLR2.7 | membrane | 4.21 |
| c130483_g1_i5 | Q9C8E7 | GLR3.3 | plasma membrane | 5.92 |
| c123516_g1_i2 | Q9SW97 | GLR3.5 | plasma membrane | −2.15 |
| c132090_g1_i2 | Q9M0A4 | CNGC9 | plasma membrane | −4.35 |
| c133601_g3_i1 | Q37145 | ACA1 | chloroplast inner membrane | 2.29 |
| c133341_g1_i1 | Q9LY77 | ACA12 | membrane | 4.28 |
| c134561_g2_i1 | Q9SYG9 | CCX4 | membrane | 3.19 |
| c132676_g3_i2 | Q8L636 | NCL | vacuole membrane, plasma membrane | 5.67 |
| c131169_g3_i1 | Q43867 | PME1 | cell wall, golgi apparatus membrane | 2.07 |
| c126734_g1_i1 | O48711 | PME12 | cell wall | 2.75 |
| c130878_g3_i1 | Q9M9W7 | PME22 | cell wall | −2.35 |
| c134669_g5_i2 | Q9M3B0 | PME34 | membrane | 16.36 |
| c116073_g1_i1 | Q8VYZ3 | PME53 | cell wall | 3.09 |
| c132368_g4_i1 | Q43062 | N/A (protein name: PE PPE8B) | cell wall | 2.73 |
| Contig ID | Uniprot ID | Functional Annotation-Gene | Putative Localization | Log2 FC VP vs. UH |
|---|---|---|---|---|
| c130483_g1_i5 | Q9C8E7 | GLR3.3 | plasma membrane | 10.85 |
| c132090_g1_i2 | Q9M0A4 | CNGC9 | plasma membrane | −8.36 |
| c133341_g1_i1 | Q9LY77 | ACA12 | membrane | −2.68 |
| c131257_g1_i2 | Q8L783 | CAX5 | vacuole membrane | 2.83 |
| c134561_g2_i1 | Q9SYG9 | CCX4 | membrane | 3.13 |
| c132676_g3_i2 | Q8L636 | NCL | vacuole membrane, plasma membrane | 4.70 |
| c113153_g1_i1 | Q9SIJ9 | PME11 | membrane | 3.63 |
| c130878_g3_i1 | Q9M9W7 | PME22 | cell wall | 4.38 |
| c123408_g2_i1 | O81301 | PME40 | cell wall | 2.40 |
| c133846_g1_i1 | Q9FHN5 | PME59 | cell wall | 2.88 |
| c132368_g4_i1 | Q43062 | N/A (protein name: PE PPE8B) | cell wall | 4.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuronuma, T.; Watanabe, H. Search for Candidate Genes Causing the Excessive Ca Accumulation in Roots of Tipburn-Damaged Lisianthus (Eustoma grandiflorum) Cultivars. Agriculture 2021, 11, 254. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030254
Kuronuma T, Watanabe H. Search for Candidate Genes Causing the Excessive Ca Accumulation in Roots of Tipburn-Damaged Lisianthus (Eustoma grandiflorum) Cultivars. Agriculture. 2021; 11(3):254. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030254
Chicago/Turabian StyleKuronuma, Takanori, and Hitoshi Watanabe. 2021. "Search for Candidate Genes Causing the Excessive Ca Accumulation in Roots of Tipburn-Damaged Lisianthus (Eustoma grandiflorum) Cultivars" Agriculture 11, no. 3: 254. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030254