
Clostridia in Insect Processed Animal Proteins—Is an Epidemiological Problem Possible?

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collections
2.2. Cultures
2.3. DNA Isolation
2.4. Characterization of Clostridia Strains by Amplification and Sequencing of 16S rDNA
2.5. Detection of BoNT-Producing Clostridia Genes
2.6. Statistical Analysis
3. Results
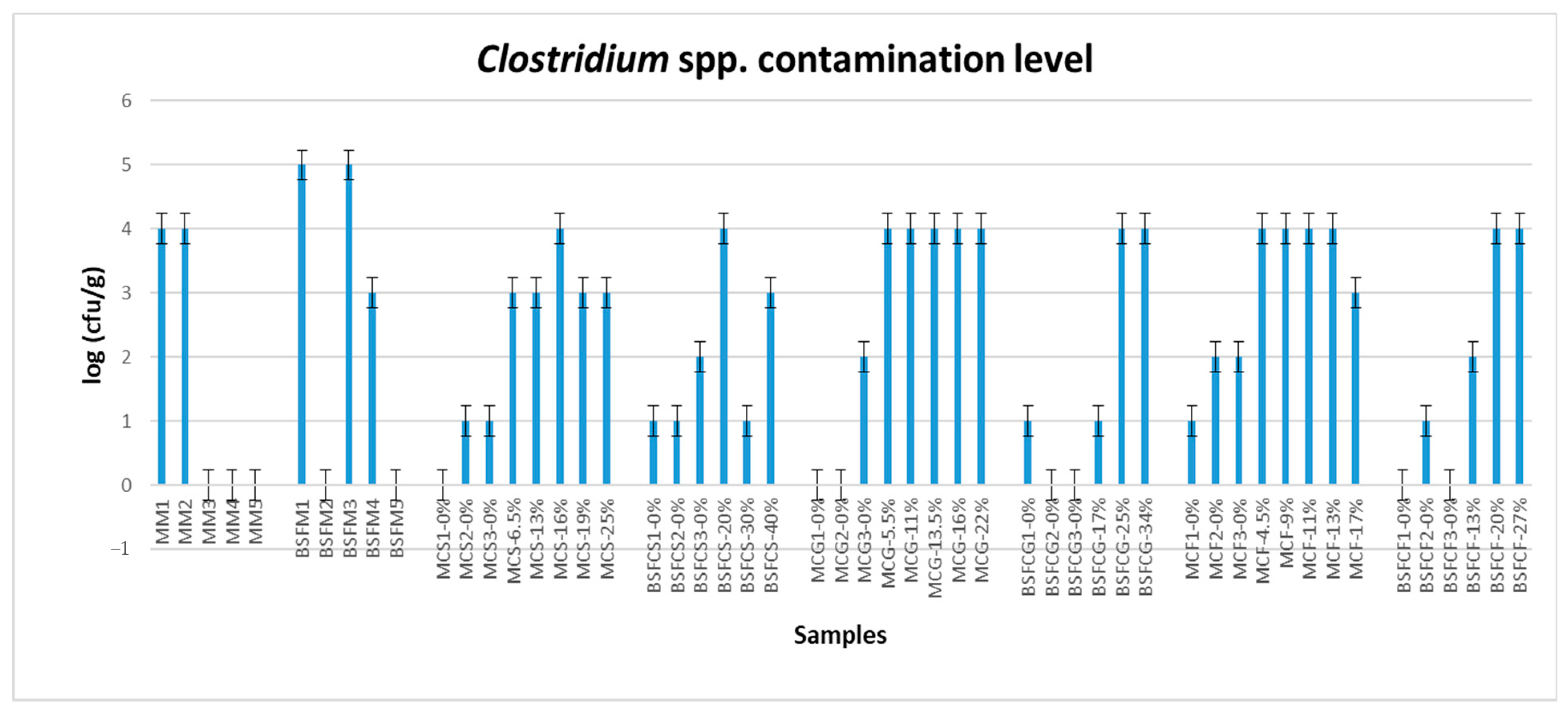
3.1. Contamination Level and Statistical Analysis Results
3.2. Results of the 16S rDNA Analysis
3.3. Occurrence of BoNT-Producing Clostridia
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Calvert, C.C. Use of Animal Excreta for Microbial and Insect Protein Synthesis. J. Anim. Sci. 1979, 48, 178–192. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Van Itterbeeck, J.; Heetkamp, M.J.W.; Van den Brand, H.; Van Loon, J.J.A.; Van Huis, A. An Exploration on Greenhouse Gas and Ammonia Production by Insect Species Suitable for Animal or Human Consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [Green Version]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The Potential Role of Insects as Feed: A Multi-Perspective Review. Animals 2019, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef] [Green Version]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Liu, Z.; Xia, L.; Wu, Y.; Xia, Q.; Chen, J.; Roux, K.H. Identification and Characterization of an Arginine Kinase as a Major Allergen from Silkworm (Bombyx mori) Larvae. Int. Arch. Allergy Immunol. 2009, 150, 8–14. [Google Scholar] [CrossRef]
- Nishimune, T.; Watanabe, Y.; Okazaki, H.; Akai, H. Thiamin is Decomposed Due to Anaphe spp. Entomophagy in Seasonal Ataxia Patients in Nigeria. J. Nutr. 2000, 130, 1625–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, H.; Kirkemo, H.-M.; Handeland, K. Wildlife as source of zoonoticinfections. Emerg. Infect. Dis. 2004, 10, 2067–2072. [Google Scholar] [CrossRef] [PubMed]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Belluco, S.; LoSasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible Insects in a Food Safety and Nutritional Perspective: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 296–313. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Jensen, A.B.; Eilenberg, J. Cannibalism as a possible entry route for opportunistic pathogenic bacteria to insect hosts, exemplified by Pseudomonas aeruginosa, a pathogen of the giant mealworm Zophobas morio. Insects 2018, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Song, F.; Peng, Q.; Brillard, J.; Lereclus, D.; Nielsen-Leroux, C. An insect gut environment reveals the induction of a new sugar-phosphate sensor system in Bacillus cereus. Gut Microbes 2013, 5, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Nayduch, D.; Cho, H.; Joyner, C. Staphylococcus aureus in the House Fly: Temporospatial fate of bacteria and expression of the antimicrobial peptide defensin. J. Med. Entomol. 2013, 50, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowski, N.T.; Klein, G. Microbiology of processed edible insect products—Results of a preliminary survey. Int. J. Food Microbiol. 2017, 243, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, Z.; Halouzka, J. Persistence of clostridium botulinum type c toxin in blow fly (calliphoridae) larvae as a possible cause of avian botulism in spring. J. Wildl. Dis. 1991, 27, 81–85. [Google Scholar] [CrossRef]
- Garofalo, C.; Osimani, A.; Milanović, V.; Taccari, M.; Cardinali, F.; Aquilanti, L.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Clementi, F. The microbiota of marketed processed edible insects as revealed by high-throughput sequencing. Food Microbiol. 2017, 62, 15–22. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as Animal Feed and Human Food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Anza, I.; Vidal, D.; Mateo, R. New insight in the epidemiology of avian botulism outbreaks: Necrophagous flies as vectors of Clostridium botulinum type C/D. Environ. Microbiol. Rep. 2014, 6, 738–743. [Google Scholar] [CrossRef]
- Espelund, M.; Klaveness, D. Botulism outbreaks in natural environments—An update. Front. Microbiol. 2014, 5, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobritsa, A.P.; Kutumbaka, K.K.; Werner, K.; Wiedmann, M.; Asmus, A.; Samadpour, M. Clostridium tepidum sp. nov., a close relative of Clostridium sporogenes and Clostridium botulinum Group, I. Int. J. Syst. Evol. Microbiol. 2017, 67, 2317–2322. [Google Scholar] [CrossRef]
- Carter, A.T.; Peck, M.W. Genomes, neurotoxins and biology of Clostridium botulinum Group I and Group II. Res. Microbiol. 2015, 166, 303–317. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.D.; East, A.K. Phylogeny and taxonomy of the food-borne pathogen Clostridium botulinum and its neurotoxins. J. Appl. Microbiol. 1998, 84, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Scalfaro, C.; Auricchio, B.; De Medici, D.; Anniballi, F. Foodborne botulism: An evolving public health challenge. Infect. Dis. 2019, 51, 97–101. [Google Scholar] [CrossRef]
- Iyer, A.V.; Blinkova, A.L.; Yanga, S.; Harrison, M.; Tepp, W.H.; Jacobson, M.J.; Johnson, E.A.; Bennett, G.N.; Walker, J.R. Clostridium taeniosporum is a close relative of the Clostridium botulinum Group II. Anaerobe 2008, 14, 318–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrocki, E.M.; Bradshaw, M.; Johnson, E.A. Botulinum neurotoxin-encoding plasmids can be conjugatively transferred to diverse clostridial strains. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Skarin, H.; Segerman, B. Horizontal gene transfer of toxin genes in Clostridium botulinum. Mob. Genet. Elements 2011, 1, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Wentz, T.G.; Muruvanda, T.; Lomonaco, S.; Thirunavukkarasu, N.; Hoffmann, M.; Allard, M.W.; Hodge, D.R.; Pillai, S.P.; Hammack, T.S.; Brown, E.W.; et al. Closed Genome Sequence of Chryseobacterium piperi Strain CTMT/ATCC BAA-1782, a Gram-Negative Bacterium with Clostridial Neurotoxin-Like Coding Sequences. Genome Announc. 2017, 5, e01296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Lebreton, F.; Mansfield, M.J.; Miyashita, S.I.; Zhang, J.; Schwartzman, J.A.; Tao, L.; Masuyer, G.; Martinez-Carranza, M.; Stenmark, P.; et al. Emergence of a botulinum neurotoxin-like toxin in a commensalstrain of Enterococcus faecium. Cell Host Microbe 2018, 23, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Moeller, R.B., Jr.; Puschner, B.; Walker, R.L.; Rocke, T.; Galey, F.D.; Cullor, J.S.; Ardans, A.A. Determination of the median toxic dose of type C. botulinum toxin in lactating dairy cows. J. Vet. Diagn. Investig. 2003, 15, 523–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevot, A.R.; Brygoo, E.R. New investigations on botulism and its five toxictypes. Ann. Inst. Pasteur 1953, 85, 544–575. [Google Scholar]
- Rossetto, O.; Montecucco, C. Tables of Toxicity of Botulinum and Tetanus Neurotoxins. Toxins 2019, 11, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondoc, I. European Regulation in the Veterinary Sanitary and Food Safety Area, a Component of the European Policies on the Safety of Food Products and the Protection of Consumer Interests: A 2007 Retrospective. Part One: The Role of European Institutions in Laying Down and Passing Laws Specific to the Veterinary Sanitary and Food Safety Area; Universul Juridic: Bucharest, Romania, 2016; pp. 12–15. [Google Scholar]
- Bondoc, I. European Regulation in the Veterinary Sanitary and Food Safety Area, a Component of the European Policies on the Safety of Food Products and the Protection of Consumer Interests: A 2007 Retrospective. Part Two: Regulations; Universul Juridic: Bucharest, Romania, 2016; pp. 16–19. [Google Scholar]
- Commission Regulation (EU) 2017/893 of 24 May 2017 Amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R0893 (accessed on 4 August 2020).
- Commission Regulation (EU) No 142/2011 of 25 February 2011 implementing Regulation (EC) No 1069/2009 of the European Parliament and of the Council Laying Down Health Rules as Regards Animal by Products and Derived Products not Intended for Human Consumption and Implementing Council Directive 97/78/EC as Regards Certain Samples and Items Exempt from Veterinary Checks at the Border under that Directive Text with EEA relevance. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32011R0142 (accessed on 4 August 2020).
- Maikanov, B.; Mustafina, R.; Auteleyeva, L.; Wiśniewski, J.; Anusz, K.; Grenda, T.; Kwiatek, K.; Goldsztejn, M.; Grabczak, M. Clostridium botulinum and Clostridium perfringens Occurrence in Kazakh Honey Samples. Toxins 2019, 11, 472. [Google Scholar] [CrossRef] [Green Version]
- Delaquis, P.J.; Baker, R.; McCurdy, A.R. Microbiological stability of pasteurized ham subjected to asecondary treatment in retort pouches. J. Food Prot. 1986, 1, 42–46. [Google Scholar] [CrossRef]
- Stumbo, C.R.; Murphy, J.R.; Cochran, J. Nature of thermal death time curves for PA3679 and Clostridium botulinum. Food Technol. 1950, 4, 321–326. [Google Scholar]
- Grecz, N.; Wagenaar, R.O.; Dack, G.M. Storage stability of Clostridium botulinum toxin and spores in processed cheese. Appl. Microbiol. 1965, 6, 1014–1022. [Google Scholar] [CrossRef]
- Mah, J.-H.; Kang, D.-H.; Tang, J. Comparison of Viability and Heat Resistance of Clostridium sporogenesStored at Different Temperatures. J. Food Sci. 2009, 74, M23–M27. [Google Scholar] [CrossRef]
- Standard Polish. Animal Feeding Stuffs-Requirements and Microbiological Examinations; Polish Committee for Standardization: Warsaw, Poland, 1994.
- Grenda, T.; Kukier, E.; Goldsztejn, M.; Kwiatek, K. In-house Validation of PCR Based Procedure for Specific Detection of Clostridium Botulinum Types C and D. Bull. Vet. Inst. Pulawy 2014, 58, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Grenda, T.; Kukier, E.; Sieradzki, Z.; Goldsztejn, M.; Kwiatek, K. In-House Validation of Multiplex PCR Method for Detection of Clostridium Botulinum in Food and Feed. Bull. Vet. Inst. Pulawy 2012, 56, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Vaneechoutte, M.; Cartwright, C.P.; Williams, E.C.; Jäger, B.; Tichy, H.-V.; De Baere, T.; De Rouck, A.; Verschraegen, G. Evaluation of 16S rRNA Gene Restriction Analysis for the Identification of Cultured Organisms of Clinically Important Clostridium Species. Anaerobe 1996, 2, 249–256. [Google Scholar] [CrossRef]
- Raphael, B.H.; Andreadis, J.D. Real-time PCR detection of the nontoxic nonhemagglutinin gene as a rapid screening method for bacterial isolates harboring the botulinum neurotoxin (A–G) gene complex. J. Microbiol. Methods 2007, 71, 343–346. [Google Scholar] [CrossRef]
- Kirchner, S.; Krämer, K.M.; Schulze, M.; Pauly, D.; Jacob, D.; Gessler, F.; Nitsche, A.; Dorner, B.G.; Dorner, M.B. Pentaplexed Quantitative Real-Time PCR Assay for the Simultaneous Detection and Quantification of Botulinum Neurotoxin-Producing Clostridia in Food and Clinical Samples. Appl. Environ. Microbiol. 2010, 76, 4387–4395. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality. Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a Test of Whether one of two Random Variables is Stochastically Larger than the Other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Agabou, A.; Alloui, N. Importance of Alphitobius diaperinus (Panzer) as a reservoir for pathogenic bacteria in Algerian broiler houses. Vet. World 2010, 3, 71–73. [Google Scholar]
- En, A.; Db, K.-K. Nutritional Composition and Microbiology of some Edible Insects Commonly Eaten in Africa, Hurdles and Future Prospects: A Critical Review. J. Food: Microbiol. Saf. Hyg. 2016, 1, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Braide, W.; Oranusi, S.U.; Udegbunam, L.I.; Oguoma, O.I.; Akobondu, C.; Nwaoguikpe, R.N. Microbiological quality of an edible caterpillar of an emperor moth, Bunaea alcinoe. J. Ecol. Nat. Environ. 2011, 3, 176–180. [Google Scholar]
- Klunder, H.; Wolkers-Rooijackers, J.; Korpela, J.; Nout, M. Microbiological aspects of processing and storage of edible insects. Food Control 2012, 26, 628–631. [Google Scholar] [CrossRef]
- SHC. Food Safety Aspects of Insects Intended for Human Consumption (Sci Com Dossier 2014/04; SHC Dossier n° 9160); FASFC’s Scientific Committee and the Committee of the Superior Health Council: Brussels, Belgium, 2014. [Google Scholar]
- Abusnina, W.; Shehata, M.; Karem, E.; Koc, Z.; Khalil, E. Clostridium sporogenes bacteremia in an immunocompetent patient. IDCases 2019, 15, e00481. [Google Scholar] [CrossRef]
- Popoff, M.R. Botulinum neurotoxins: Still a privilege of Clostridia? Cell Host Microbe 2018, 23, 145–146. [Google Scholar] [CrossRef] [Green Version]
- Nyaoke, A.C.; Navarro, M.A.; Fresneda, K.; Diab, S.S.; Moore, J.; Lyras, D.; Awad, M.; Uzal, F.A. Paeniclostridium (Clostridium) sordellii-associated enterocolitis in 7 horses. J. Vet. Diagn. Investig. 2020, 32, 239–245. [Google Scholar] [CrossRef]
- Aldape, M.J.; Bryant, A.E.; Stevens, D.L. Clostridium sordellii Infection: Epidemiology, Clinical Findings, and Current Perspectives on Diagnosis and Treatment. Clin. Infect. Dis. 2006, 43, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Rabi, R.; Turnbull, L.; Whitchurch, C.B.; Awad, M.; Lyras, D. Structural Characterization of Clostridium sordellii Spores of Diverse Human, Animal and Environmental Origin and Comparison to Clostridium difficile Spores. Msphere 2017, 2, e00343. [Google Scholar] [CrossRef] [Green Version]
- Hale, A.; Kirby, J.E.; Albrecht, M. Fatal Spontaneous Clostridium bifermentans Necrotizing Endometritis: A Case Report and Literature Review of the Pathogen. Open Forum Infect. Dis. 2016, 3, ofw095. [Google Scholar] [CrossRef] [Green Version]
- Scanlan, D.R.; Smith, M.A.; Isenberg, H.D.; Engrassia, S.; Hilton, E. Clostridium bifermentans bacteremia with metastatic osteomyelitis. J. Clin. Microbiol. 1994, 32, 2867–2868. [Google Scholar] [CrossRef] [Green Version]
- Misra, D.P.; Hurst, D.J. Necrotising pneumonia and empyema caused by Clostridium bifermentans. Thorax 1980, 35, 310–311. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.T.; Paul, C.J.; Mason, D.R.; Twine, S.M.; Alston, M.J.; Logan, S.M.; Austin, J.W.; Peck, M.W. Independent evolution of neurotoxin and flagellar genetic loci in proteolytic Clostridium botulinum. BMC Genom. 2009, 10, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobritsa, A.P.; Kutumbaka, K.K.; Samadpour, M. Reclassification of Eubacterium combesii and discrepancies in the nomenclature of botulinum neurotoxin-producing clostridia: Challenging Opinion 69. Request for an Opinion. Int. J. Syst. Evol. Microbiol. 2018, 68, 3068–3075. [Google Scholar] [CrossRef]
- Brunt, J.; Van Vliet, A.H.M.; Carter, A.T.; Stringer, S.C.; Amar, C.; Grant, K.A.; Godbole, G.; Peck, M.W. Diversity of the Genomes and Neurotoxins of Strains of Clostridium botulinum Group I and Clostridium sporogenes Associated with Foodborne, Infant and Wound Botulism. Toxins 2020, 12, 586. [Google Scholar] [CrossRef] [PubMed]
- Peck, M.W. Biology and Genomic Analysis of Clostridium botulinum. Adv. Microb. Physiol. 2009, 55, 183–320. [Google Scholar] [CrossRef]
- Kutsuna, R.; Tomida, J.; Morita, Y.; Kawamura, Y. Paraclostridium bifermentans exacerbates pathosis in a mouse model of ulcerative colitis. PLoS ONE 2018, 13, e0197668. [Google Scholar] [CrossRef]
- Doxey, A.C.; Lynch, M.D.; Müller, K.M.; Meiering, E.M.; McConkey, B.J. Insights into the evolutionary origins of clostridial neurotoxins from analysis of the Clostridium botulinum strain A neurotoxin gene cluster. BMC Evol. Biol. 2008, 8, 316. [Google Scholar] [CrossRef] [Green Version]
- Marshall, K.M.; Bradshaw, M.; Pellett, S.; Johnson, E.A. Plasmid encoded neurotoxin genes in Clostridium botulinum serotype A subtypes. Biochem. Biophys. Res. Commun. 2007, 361, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Dineen, S.S.; Bradshaw, M.; Johnson, E.A. Neurotoxin Gene Clusters in Clostridium botulinum Type A Strains: Sequence Comparison and Evolutionary Implications. Curr. Microbiol. 2003, 46, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Franciosa, G.; Ferreira, J.L.; Hatheway, C.L. Detection of type A, B, and E botulism neurotoxin genes in Clostridium botulinum and other Clostridium species by PCR: Evidence of unexpressed type B toxin genes in type A toxigenic organisms. J. Clin. Microbiol. 1994, 32, 1911–1917. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.J.; Hill, K.K.; Raphael, B.H. Historical and current perspectives on Clostridium botulinum diversity. Res. Microbiol. 2015, 166, 290–302. [Google Scholar] [CrossRef]
- Lindberg, A.; Skarin, H.; Knutsson, R.; Blomqvist, G.; Båverud, V. Real-time PCR for Clostridium botulinum type C neurotoxin (BoNTC) gene, also covering a chimeric C/D sequence—Application on outbreaks of botulism in poultry. Vet. Microbiol. 2010, 146, 118–123. [Google Scholar] [CrossRef]
- Souillard, R.; Woudstra, C.; Le Maréchal, C.; Dia, M.; Bayon-Auboyer, M.H.; Chemaly, M.; Fach, P.; Le Bouquin, S. Investigation of Clostridium botulinum in commercial poultry farms in France between 2011 and 2013. Avian Pathol. 2014, 43, 458–464. [Google Scholar] [CrossRef] [Green Version]
- Circella, E.; Camarda, A.; Bano, L.; Marzano, G.; Lombardi, R.; D’Onghia, F.; Greco, G. Botulism in Wild Birds and Changes in Environmental Habitat: A Relationship to be Considered. Animals 2019, 9, 1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhaus, J.; Schrödl, W.; Shehata, A.A.; Krüger, M. Detection of Clostridium botulinum in liquid manure and biogas plant wastes. Folia Microbiol. 2015, 60, 451–456. [Google Scholar] [CrossRef]
- Rasetti-Escargueil, C.; Lemichez, E.; Popoff, M.R. Public Health Risk Associated with Botulism as Foodborne Zoonoses. Toxins 2019, 12, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stringer, S.C.; Haque, N.; Peck, M.W. Growth from Spores of Nonproteolytic Clostridium botulinum in Heat-Treated Vegetable Juice. Appl. Environ. Microbiol. 1999, 65, 2136–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Regulation (EC) No 1069/2009 of the European Parliament and of the Council of 21 October 2009 Laying down Health Rules as Regards Animal by-Products and Derived Products not Intended for Human Consumption and Repealing Regulation (EC) No 1774/2002 (Animal by-Products Regulation); European Commission: Brussels, Belgium, 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32009R1069 (accessed on 9 March 2020).


{kind=link}
| Number of Samples | Mealworm IPAP | Black Soldier IPAP | Compound Feed Samples with and without Addition of Mealworm IPAP | Compound Feed Samples with and without Addition of Black Soldier Fly IPAP | ||
|---|---|---|---|---|---|---|
| Control Group | Experimental Group | Control Group | Experimental Group | |||
| 1 | MM1 | BSFM1 | MCS1-0% * | MCS-6.5% * | BSFCS1-0% * | BSFCS-20% * |
| 2 | MM2 | BSFM2 | MCS2-0% | MCS-13% | BSFCS2-0% | BSFCS-30% |
| 3 | MM3 | BSFM3 | MCS3-0% | MCS-16% | BSFCS3-0% | BSFCS-40% |
| 4 | MM4 | BSFM4 | MCG1-0% | MCS-19% | BSFCG1-0% | BSFCG-17% |
| 5 | MM5 | BSFM5 | MCG2-0% | MCS-25% | BSFCG2-0% | BSFCG-25% |
| 6 | MCG3-0% | MCG-5.5% | BSFCG3-0% | BSFCG-34% | ||
| 7 | MCF1-0% | MCG-11% | BSFCF1-0% | BSFCF-13% | ||
| 8 | MCF2-0% | MCG-13.5% | BSFCF2-0% | BSFCF-20% | ||
| 9 | MCF3-0% | MCG-16% | BSFCF3-0% | BSFCF-27% | ||
| 10 | MCG-22% | |||||
| 11 | MCF-4.5% | |||||
| 12 | MCF-9% | |||||
| 13 | MCF-11% | |||||
| 14 | MCF-13% | |||||
| 15 | MCF-17% | |||||
| Total number of samples | 52 | |||||
| IPAP Samples | Producer Location (Country) |
|---|---|
| MM1 | Poland |
| MM2 | Poland |
| MM3 | Denmark |
| MM4 | France |
| MM5 | Spain |
| BSFM1 | Poland |
| BSFM2 | Germany |
| BSFM3 | Ireland |
| BSFM4 | France |
| BSFM5 | France |
| MCS-0% | MCS-6.5% | MCS-13% | MCS-16% | MCS-19% | MCS-25% | MCG-0% | MCG-5.5% | MCG-11% | MCG-13.5% | MCG-16% | MCG-22% | MCF-0% | MCF-4.5% | MCF-9% | MCF-11% | MCF-13% | MM-17% | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Substitution of post-extraction soybean meal (%) | 0 | 25 | 50 | 62.5 | 75 | 100 | 0 | 25 | 50 | 62.5 | 75 | 100 | 0 | 25 | 50 | 62.5 | 75 | 100 |
| Feed Ingredients | ||||||||||||||||||
| Maize (%) | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 20 | 20 | 20 | 20 | 20 | 20 |
| Wheat (%) | 29.39 | 33.05 | 35.32 | 37.58 | 39.28 | 42.83 | 31.40 | 34.13 | 37.47 | 39.02 | 40.83 | 43.35 | 46.85 | 49.15 | 51.75 | 53.19 | 54.67 | 57.59 |
| Post-extraction soybean meal (%) | 34 | 25 | 17 | 12 | 8.5 | 0 | 30 | 22.5 | 15 | 11.25 | 7.5 | 0 | 24 | 18 | 12 | 9 | 6 | - |
| Soybean oil (%) | 2.5 | 2 | 1.5 | 0.9 | 0.6 | - | 4.5 | 4 | 3.5 | 3.2 | 3 | 2.5 | 5.5 | 5 | 4.5 | 4.2 | 4 | 3.5 |
| Mealworm IPAP (%) | 0 | 6.5 | 13 | 16 | 19 | 25 | 0 | 5.5 | 11 | 13.5 | 16 | 22 | 0 | 4.5 | 9 | 11 | 13 | 17 |
| Monocalcium phosphate (%) | 1.5 | 1.1 | 0.9 | 0.8 | 0.6 | 0.3 | 1.3 | 1.2 | 0.7 | 0.8 | 0.5 | 0.3 | 1.2 | 0.9 | 0.7 | 0.60 | 0.4 | 0.2 |
| Fodder chalk (%) | 1.25 | 0.8 | 0.7 | 0.6 | 0.4 | 0.2 | 1.25 | 1.2 | 0.8 | 0.7 | 0.6 | 0.2 | 1.25 | 1.1 | 0.80 | 0.7 | 0.5 | 0.3 |
| Sodium bicarbonate (%) | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Fodder salt (%) | 0.2 | 0.15 | 0.1 | 0.1 | 0.05 | 0.05 | 0.2 | 0.12 | 0.1 | 0.05 | - | - | 0.2 | 0.15 | 0.1 | 0.05 | 0.05 | 0.05 |
| Mineral and vitamin premix (%) | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Lysine 78% | 0.25 | 0.4 | 0.45 | 0.5 | 0.55 | 0.6 | 0.25 | 0.4 | 0.48 | 0.5 | 0.57 | 0.65 | 0.18 | 0.3 | 0.3 | 0.35 | 0.37 | 0.45 |
| Methionine 99% | 0.24 | 0.3 | 0.3 | 0.32 | 0.32 | 0.32 | 0.16 | 0.2 | 0.2 | 0.23 | 0.25 | 0.25 | 0.12 | 0.15 | 0.15 | 0.2 | 0.2 | 0.2 |
| Threonine 98% | 0.02 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.04 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.05 | 0.10 | 0.05 | 0.06 | 0.06 | 0.06 |
| Choline chloride | 0.09 | 0.09 | 0.09 | 0.90 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 |
| Total (%) | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| BSFCS-0% | BSFCS-20% | BSFCS-30% | BSFCS-40% | BSFCG-0% | BSFCG-17% | BSFCG-25% | BSFCG-34% | BSFCF-0% | BSFCF-13% | BSFCF-20% | BSFCF-27% | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Substitution of Post-Extraction Soybean Meal (%) | 0 | 50 | 75 | 100 | 0 | 50 | 75 | 100 | 0 | 50 | 75 | 100 |
| Feed Ingredients | ||||||||||||
| Maize (%) | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 20 | 20 | 20 | 20 |
| Wheat (8%) | 28.75 | 30.36 | 29.16 | 27.78 | 31.46 | 33.36 | 40.68 | 41.21 | 46.51 | 50.51 | 49.66 | 48.61 |
| Post-extraction soybean meal (%) | 34 | 17 | 8.5 | 0 | 30 | 15 | 7.5 | 0 | 24 | 12 | 6 | 0 |
| Soybean oil (%) | 3 | - | - | - | 4.5 | 2.5 | 3 | 2.5 | 5.5 | 3.5 | 3 | 2.5 |
| Black soldier fly IPAP (%) | 0 | 20 | 30 | 40 | 0 | 17 | 25 | 34 | 0 | 13 | 20 | 27 |
| Monocalcium phosphate (%) | 1.5 | 0.4 | 0.3 | 0.2 | 1.3 | 0.4 | 0.5 | 0.3 | 1.2 | 0.3 | 0.2 | 0.2 |
| Fodder chalk (%) | 1.25 | 0.5 | 0.3 | 0.2 | 1.25 | 0.5 | 0.6 | 0.2 | 1.25 | 0.4 | 0.3 | 0.3 |
| Sodium bicarbonate (%) | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Fodder salt (%) | 0.2 | 0.05 | 0.05 | 0.05 | 0.2 | 0.1 | - | - | 0.2 | 0.05 | - | - |
| Mineral and vitamin premix (%) | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Lysine (%) | 0.25 | 0.42 | 0.45 | 0.60 | 0.25 | 0.35 | 0.57 | 0.65 | 0.18 | 0.30 | 0.35 | 0.4 |
| Methionine (%) | 0.24 | 0.28 | 0.3 | 0.3 | 0.16 | 0.25 | 0.19 | 0.19 | 0.12 | 0.15 | 0.15 | 0.15 |
| Threonine (%) | 0.02 | 0.15 | 0.15 | 0.15 | 0.04 | 0.15 | 0.1 | 0.1 | 0.05 | 0.1 | 0.1 | 0.1 |
| Choline chloride (%) | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 | 0.09 |
| Total (%) | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| MCS-0% | MCS-6.5% | MCS-13% | MCS-16% | MCS-19% | MCS-25% | MCG-0% | MCG-5.5% | MCG-11% | MCG-13.5% | MCG-16% | MCG-22% | MCF-0% | MCF-4.5% | MCF-9% | MCF-11% | MCF-13% | MM-17% | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM, kcal/kg | 2990 | 2992 | 2990 | 3000 | 3000 | 3013 | 3120 | 3120 | 3128 | 3120 | 3138 | 3120 | 3180 | 3180 | 3180 | 3180 | 3180 | 3180 |
| EM, MJ/kg | 12.35 | 12.53 | 12.51 | 12.56 | 12.56 | 12.62 | 13.12 | 13.12 | 13.14 | 13.12 | 13.17 | 13.12 | 13.28 | 13.28 | 13.28 | 13.28 | 13.23 | 13.23 |
| Total proteins (%) | 22.00 | 21.96 | 22.11 | 22.02 | 22.08 | 22.00 | 20.00 | 20.17 | 20.00 | 20.13 | 20.00 | 20.00 | 18.30 | 18.45 | 18.21 | 18.39 | 18.26 | 18.29 |
| Lysine (%) | 1.35 | 1.38 | 1.35 | 1.36 | 1.35 | 1.35 | 1.25 | 1.29 | 1.26 | 1.27 | 1.27 | 1.26 | 1.03 | 1.08 | 1.00 | 1.04 | 1.00 | 1.00 |
| Methionine (%) | 0.58 | 0.60 | 0.59 | 0.60 | 0.62 | 0.62 | 0.47 | 0.48 | 0.54 | 0.49 | 0.51 | 0.53 | 0.45 | 0.46 | 0.47 | 0.46 | 0.50 | 0.52 |
| Met+ Cys (%) | 1.04 | 1.07 | 1.02 | 1.03 | 1.08 | 1.03 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.82 | 0.84 | 0.81 | 0.82 | 0.83 | 0.83 |
| Threonine (%) | 0.80 | 0.82 | 0.80 | 0.82 | 0.84 | 0.88 | 0.80 | 0.81 | 0.80 | 0.80 | 0.80 | 0.80 | 0.67 | 0.73 | 0.66 | 0.67 | 0.66 | 0.66 |
| Tryptophan (%) | 0.25 | 0.25 | 0.23 | 0.26 | 0.23 | 0.23 | 0.22 | 0.23 | 0.21 | 0.24 | 0.22 | 0.21 | 0.21 | 0.22 | 0.20 | 0.20 | 0.20 | 0.20 |
| Crude fiber (%) | 2.88 | 3.88 | 3.69 | 3.94 | 3.71 | 3.98 | 2.78 | 3.67 | 3.22 | 3.70 | 3.50 | 3.65 | 2.78 | 3.51 | 3.19 | 3.56 | 3.37 | 3.55 |
| Raw fat (%) | 5.96 | 5.16 | 5.44 | 5.29 | 5.37 | 5.54 | 6.31 | 6.93 | 7.83 | 7.15 | 7.38 | 8.60 | 7.67 | 7.47 | 7.55 | 7.53 | 7.57 | 7.18 |
| Linoleic acid (%) | 0.83 | 2.63 | 4.14 | 5.24 | 6.07 | 7.73 | 0.86 | 2.37 | 3.89 | 4.57 | 5.26 | 6.89 | 0.73 | 1.96 | 3.20 | 3.76 | 4.31 | 5.41 |
| Crude ash (%) | 5.86 | 5.06 | 4.65 | 4.45 | 3.99 | 3.44 | 5.52 | 4.91 | 4.38 | 3.88 | 3.98 | 3.30 | 5.26 | 4.77 | 4.17 | 3.50 | 3.51 | 3.10 |
| Total Ca (%) | 1.10 | 1.08 | 1.18 | 1.31 | 1.25 | 1.30 | 1.00 | 1.08 | 1.00 | 1.08 | 1.00 | 1.00 | 0.86 | 0.90 | 0.90 | 0.91 | 0.87 | 0.86 |
| Total phosphorus (%) | 1.52 | 1.58 | 1.65 | 1.68 | 1.66 | 1.79 | 1.51 | 1.57 | 1.57 | 1.64 | 1.63 | 1.70 | 1.17 | 1.19 | 1.25 | 1.27 | 1.26 | 1.30 |
| Absorbable phosphorus (%) | 0.47 | 0.52 | 0.60 | 0.63 | 0.67 | 0.68 | 0.50 | 0.50 | 0.51 | 0.58 | 0.57 | 0.60 | 0.40 | 0.41 | 0.47 | 0.48 | 0.48 | 0.52 |
| Na (%) | 0.17 | 0.22 | 0.24 | 0.27 | 0.28 | 0.30 | 0.17 | 0.18 | 0.20 | 0.21 | 0.23 | 0.25 | 0.17 | 0.18 | 0.19 | 0.19 | 0.20 | 0.23 |
| BSFCS-0% | BSFCS-20% | BSFCS-30% | BSFCS-40% | BSFCG-0% | BSFCG-17% | BSFCG-25% | BSFCG-34% | BSFCF-0% | BSFCF-13% | BSFCF-20% | BSFCF-27% | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM, kcal/kg | 2990 | 2990 | 3000 | 3013 | 3100 | 3100 | 3100 | 3100 | 3160 | 3160 | 3160 | 3160 |
| EM, MJ/kg | 12.35 | 12.56 | 12.56 | 12.62 | 12.98 | 12.98 | 12.98 | 12.98 | 13.22 | 13.22 | 13.22 | 13.22 |
| Total proteins (%) | 22.00 | 22.11 | 22.08 | 22.00 | 20.07 | 20.00 | 20.00 | 20.00 | 18.00 | 18.11 | 18.16 | 18.19 |
| Lysine (%) | 1.35 | 1.35 | 1.35 | 1.35 | 1.17 | 1.18 | 1.18 | 1.18 | 1.03 | 1.00 | 1.00 | 1.00 |
| Methionine (%) | 0.58 | 0.64 | 0.67 | 0.67 | 0.47 | 0.62 | 0.60 | 0.60 | 0.45 | 0.49 | 0.50 | 0.52 |
| Met + Cys (%) | 1.04 | 1.02 | 1.02 | 1.03 | 0.95 | 0.89 | 0.88 | 0.88 | 0.82 | 0.87 | 0.87 | 0.88 |
| Threonine (%) | 0.80 | 0.80 | 0.84 | 0.88 | 0.80 | 0.84 | 0.84 | 0.84 | 0.67 | 0.70 | 0.70 | 0.71 |
| Tryptophan (%) | 0.25 | 0.23 | 0.23 | 0.23 | 0.22 | 0.21 | 0.22 | 0.21 | 0.21 | 0.20 | 0.20 | 0.20 |
| Crude fiber (%) | 2.88 | 3.81 | 4.23 | 4.78 | 2.78 | 3.62 | 4.00 | 4.44 | 2.78 | 3.45 | 3.78 | 4.12 |
| Raw fat (%) | 6.96 | 8.15 | 10.99 | 13.84 | 6.31 | 9.66 | 10.99 | 13.07 | 7.67 | 9.20 | 10.70 | 12.23 |
| Linoleic acid (%) | 0.83 | 6.29 | 9.00 | 11.27 | 0.86 | 3.89 | 5.26 | 6.89 | 0.73 | 3.20 | 4.31 | 5.41 |
| Crude ash (%) | 5.86 | 4.87 | 4.98 | 5.23 | 5.52 | 4.38 | 3.98 | 3.30 | 5.26 | 4.17 | 3.51 | 3.10 |
| Total Ca (%) | 1.10 | 1.30 | 1.57 | 1.84 | 1.00 | 1.00 | 1.00 | 1.00 | 0.86 | 0.90 | 1.00 | 1.25 |
| Total phosphorus (%) | 1.52 | 1.65 | 1.66 | 1.79 | 1.51 | 1.65 | 1.78 | 1.83 | 1.17 | 1.25 | 1.26 | 1.30 |
| Absorbable phosphorus (%) | 0.47 | 0.66 | 0.87 | 0.88 | 0.50 | 0.58 | 0.70 | 0.80 | 0.40 | 0.47 | 0.59 | 0.64 |
| Na (%) | 0.17 | 0.31 | 0.32 | 0.33 | 0.17 | 0.24 | 0.29 | 0.30 | 0.17 | 0.20 | 0.25 | 0.26 |
| Sample Types | Groups | Starter | Grower | Finisher | Mann-Whitney U Test Result (p-Value) | |||
|---|---|---|---|---|---|---|---|---|
| Sample Name | Results (log cfu/g) | Sample Name | Results (log cfu/g) | Sample Name | (log cfu/g) | |||
| Feed compounds with and without addition of mealworm and black soldier fly IPAP | Control group | MCS1-0% | 0 | MCG1-0% | 0 | MCF1-0% | 1 | p < 0.001 |
| MCS2-0% | 1 | MCG2-0% | 0 | MCF2-0% | 2 | |||
| MCS3-0% | 1 | MCG3-0% | 2 | MCF3-0% | 2 | |||
| BSFCS1-0% | 0 | BSFCG1-0% | 0 | BSFCF1-0% | 1 | |||
| BSFCS2-0% | 1 | BSFCG2-0% | 0 | BSFCF2-0% | 2 | |||
| BSFCS3-0% | 2 | BSFCG3-0% | 2 | BSFCF3-0% | 2 | |||
| Experimental group | MCS-6.5% | 3 | MCG-5.5% | 4 | MCF-4.5% | 4 | ||
| MCS-13% | 3 | MCG-11% | 4 | MCF-9% | 4 | |||
| MCS-16% | 4 | MCG-13.5% | 4 | MCF-11% | 4 | |||
| MCS-19% | 3 | MCG-16% | 4 | MCF-13% | 4 | |||
| MCS-25% | 3 | MCG-22% | 4 | MCF-17% | 3 | |||
| BSFCS-20% | 4 | BSFCG-17% | 1 | BSFCF-13% | 2 | |||
| BSFCS-30% | 1 | BSFCG-25% | 4 | BSFCF-20% | 4 | |||
| BSFCS-40% | 3 | BSFCG-34% | 4 | BSFCF-27% | 4 | |||
| Feed compounds with and without addition of mealworm IPAP | Control group | MCS1-0% | 0 | MCG1-0% | 0 | MCF1-0% | 1 | p = 0.001 |
| MCS2-0% | 1 | MCG2-0% | 0 | MCF2-0% | 2 | |||
| MCS3-0% | 1 | MCG3-0% | 2 | MCF3-0% | 2 | |||
| Experimental group | MCS-6.5% | 3 | MCG-5.5% | 4 | MCF-4.5% | 4 | ||
| MCS-13% | 3 | MCG-11% | 4 | MCF-9% | 4 | |||
| MCS-16% | 4 | MCG-13.5% | 4 | MCF-11% | 4 | |||
| MCS-19% | 3 | MCG-16% | 4 | MCF-13% | 4 | |||
| MCS-25% | 3 | MCG-22% | 4 | MCF-17% | 3 | |||
| Feed compounds with and without addition of black soldier fly IPAP | Control group | BSFCS1-0% | 0 | BSFCG1-0% | 0 | BSFCF1-0% | 1 | p = 0.003 |
| BSFCS2-0% | 1 | BSFCG2-0% | 0 | BSFCF2-0% | 2 | |||
| BSFCS3-0% | 2 | BSFCG3-0% | 2 | BSFCF3-0% | 2 | |||
| Experimental group | BSFCS-20% | 4 | BSFCG-17% | 1 | BSFCF-13% | 2 | ||
| BSFCS-30% | 1 | BSFCG-25% | 4 | BSFCF-20% | 4 | |||
| BSFCS-40% | 3 | BSFCG-34% | 4 | BSFCF-27% | 4 | |||
| MM * | BSFM * | ||
|---|---|---|---|
| Sequencing Results According to BLAST Analysis | Sequence ID; % Similarity | Sequencing Results According to BLAST Analysis | Sequence ID; % Similarity |
| Clostridium tepidum | NR_157639.1; 97% | Peaniclostridium sordelli | LC15549.1; 97% |
| Clostridium botulinum, group I | CP046450.1; 95% | Paraclostridium bifermentans | MF510818; 97% |
| Peaniclostridium sordelli | LC15549.1; 97% | ||
| Paraclostridium bifermentans | MF510818; 97% | ||
| Clostridium tepidum | NR_157639.1; 98% | ||
| Clostridium botulinum group I | CP046450.1; 97% | ||
| Feed Type | Control Group, Mealworm Feed Compounds: MCS, MCG, and MCF-0% | Experimental Group, Mealworm Feed Compounds: MCS, MCG, and MCF | ||
|---|---|---|---|---|
| Sequencing Results According to BLAST Analysis | Sequence ID; % Similarity | Sequencing Results According to BLAST Analysis | Sequence ID; % Similarity | |
| Starter | Paraclostridium bifermentans | MN75886.1; 97% | Clostridium tepidum | NR_157639.1; 98% |
| Paraclostridium bifermentans | MN758863.1; 97% | |||
| Paraclostridium benzoelyticum | MN999977.1; 98% | |||
| Clostridium sporogenes | KY962939.1; 90% | |||
| Grower | Paraclostridium bifermentans | MN75886.1; 97% | Paraclostridium bifermentans | MK894870.1; 98% |
| Clostridium tepidum | NR_157639.1; 98% | |||
| Finisher | Paraclostridium bifermentans | MN75886.1; 97% | Paraclostridium sulfidigenes | MF967245.1; 96% |
| Paraclostridium benzoelyticum | LC515632.1; 97% | |||
| Paraclostridium bifermentans | MF510818.1; 98% | |||
| Feed Type | Control Group, Black Soldier Fly Feed Compound: HCS, HCG, and HCF-0% | Experimental Group, Black Soldier Fly Feed Compound: BSFCS, BSFCG, and BSFCF | ||
|---|---|---|---|---|
| Sequencing Results According to BLAST Analysis | Sequence ID; % Similarity | Sequencing Results According to Blast Analysis | Sequence ID, % Similarity | |
| Starter | Clostridium sporogenes | MT356160.1; 95% | Clostridium botulinum group I | (CP031097.1; 93%), |
| Clostridium sartagoforme | MN646980.1; 90% | Paraclostridium benzoelyticum | (MT510437.1; 100%) | |
| Grower | Clostridium sporogenes | MT356160.1; 95% | Paraclostridium benzoelyticum | (MT510437.1, 97%) |
| Clostridium sporogenes | (MT356160.1; 95%) | |||
| Finisher | Clostridium sporogenes | MT356160.1; 95% | Clostridium bifermentans | (DQ680018.1; 97%), |
| Paraclostridium benzoelyticum | (MT510437.1; 97%) | |||
| Clostridium tepidum | (NR_157639.1, 97%) | |||
| Clostridium sporogenes | (MT356160.1; 100%) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grenda, T.; Kwiatek, K.; Goldsztejn, M.; Sapała, M.; Kozieł, N.; Domaradzki, P. Clostridia in Insect Processed Animal Proteins—Is an Epidemiological Problem Possible? Agriculture 2021, 11, 270. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030270
Grenda T, Kwiatek K, Goldsztejn M, Sapała M, Kozieł N, Domaradzki P. Clostridia in Insect Processed Animal Proteins—Is an Epidemiological Problem Possible? Agriculture. 2021; 11(3):270. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030270
Chicago/Turabian StyleGrenda, Tomasz, Krzysztof Kwiatek, Magdalena Goldsztejn, Magdalena Sapała, Nina Kozieł, and Piotr Domaradzki. 2021. "Clostridia in Insect Processed Animal Proteins—Is an Epidemiological Problem Possible?" Agriculture 11, no. 3: 270. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030270