
A New Breeding Strategy towards Introgression and Characterization of Stay-Green QTL for Drought Tolerance in Sorghum

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. SSR Markers and Stay-Green QTL
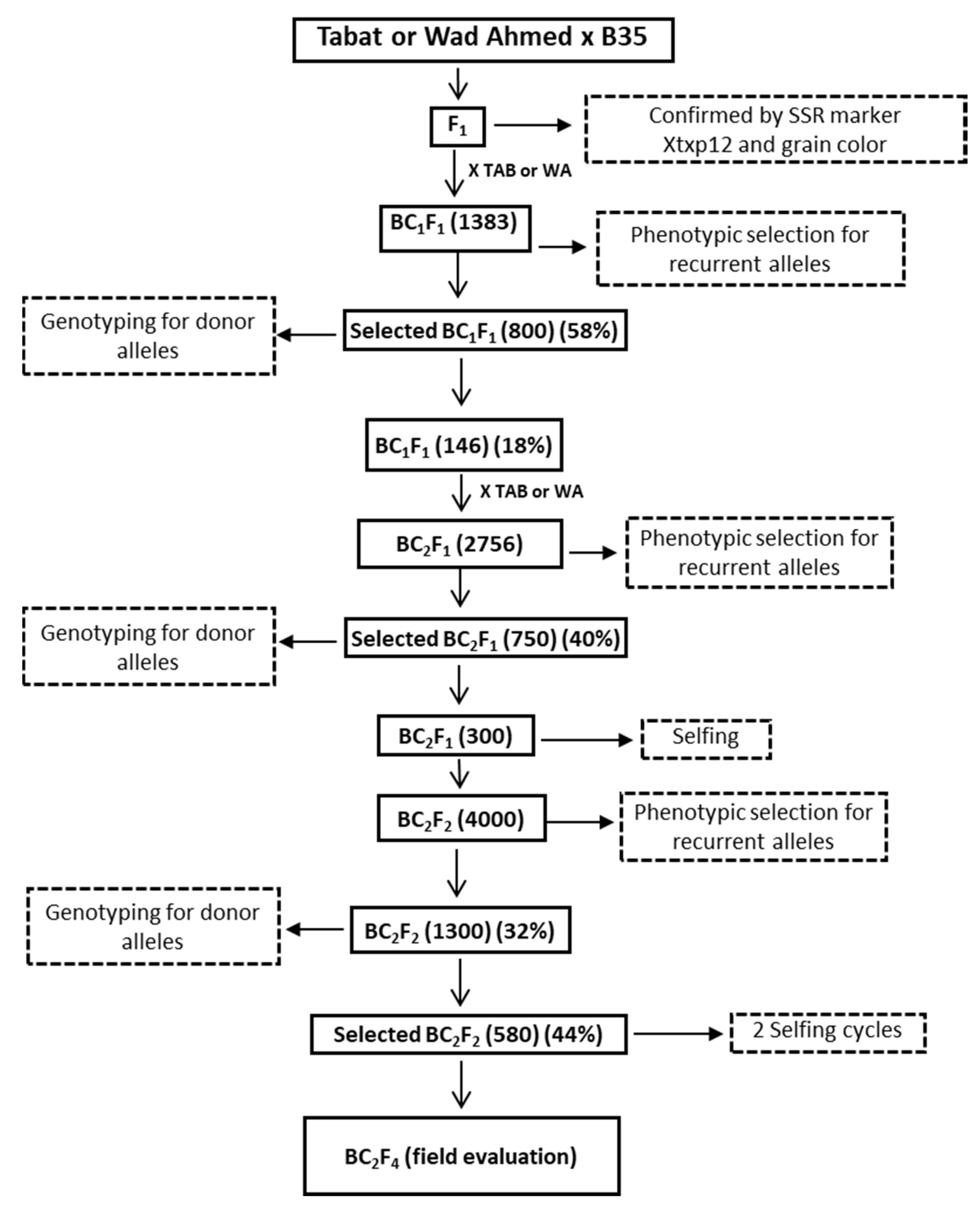
2.3. Crossing Scheme and Generations Development
2.4. DNA Pooling, Duplex, and Multiplex PCR Approach in Genotyping
2.5. Drought Adaptation Experiment for BC2F4 Stay-Green Introgression Lines
2.6. Statistical Analysis
2.7. Cost Analysis
3. Results
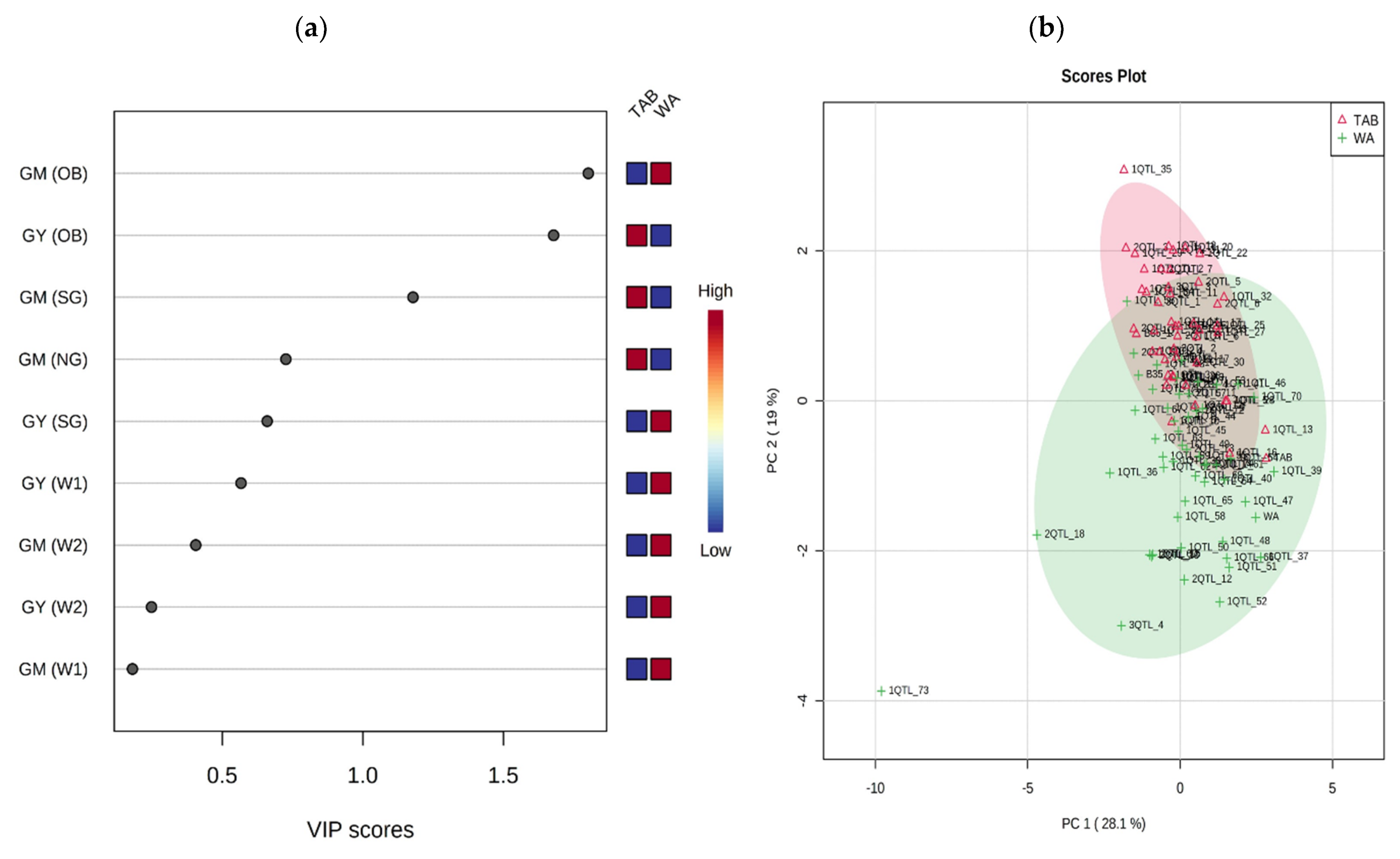
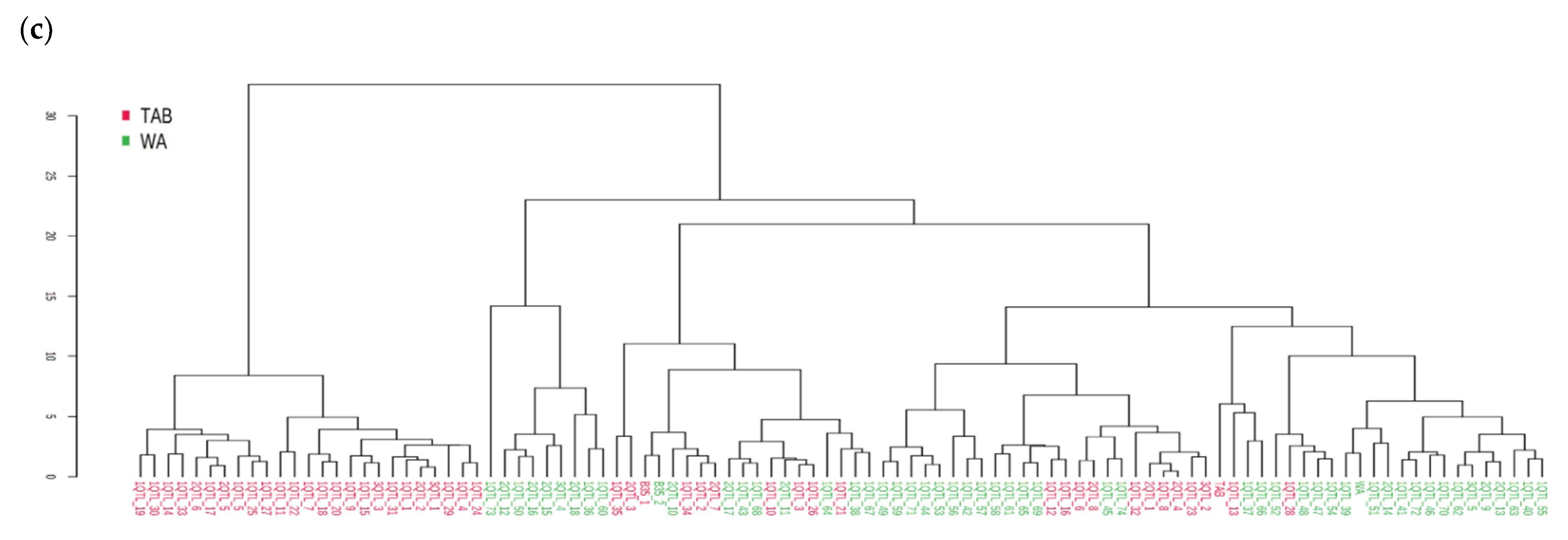
3.1. Development of Stay-Green Introgression Lines Using C-MAB
3.2. Development of BC2F1 and BC2F2 Generation
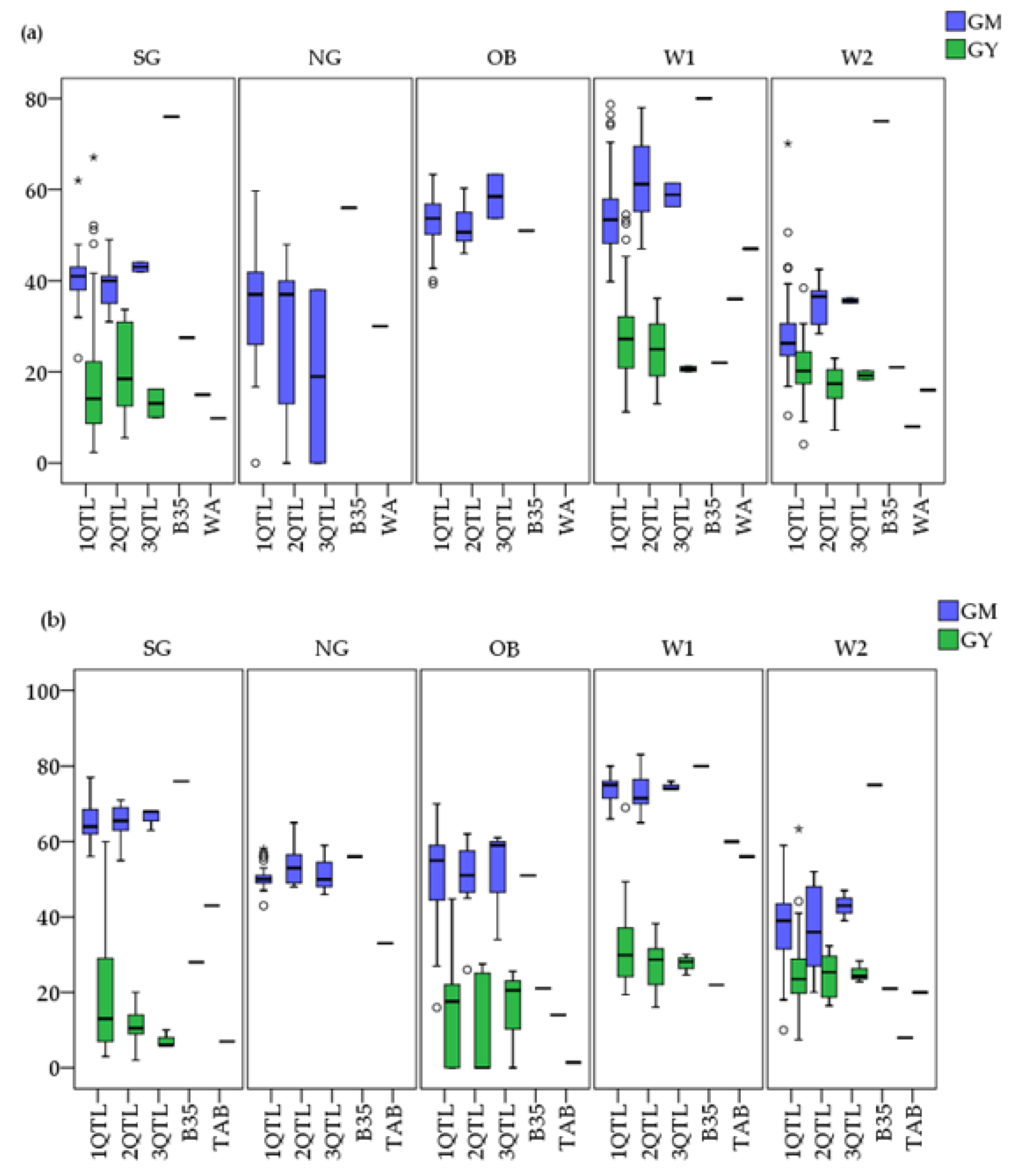
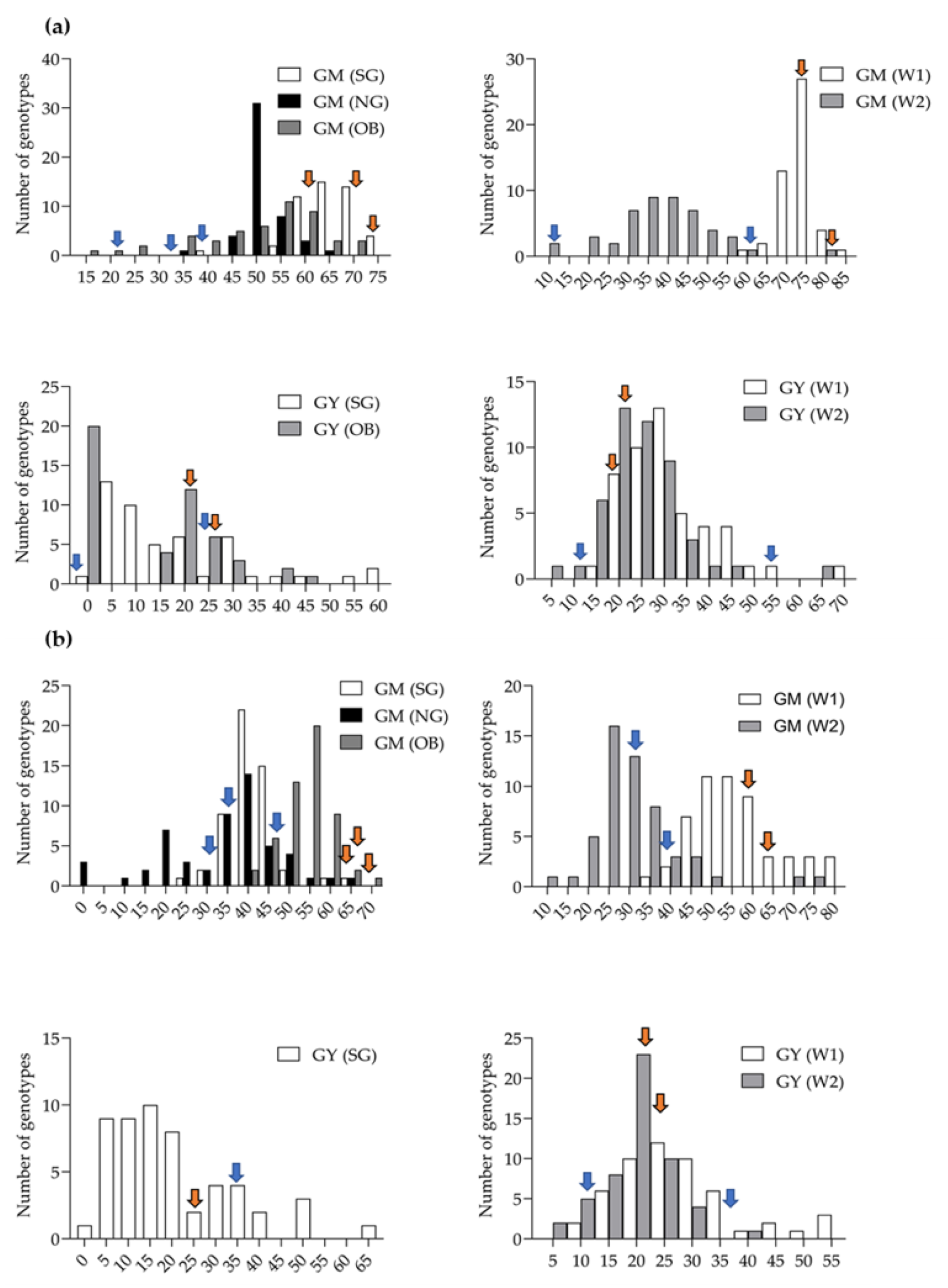
3.3. Field Performance of C-MAB-Introgression Lines
3.4. Performance of ILs under Irrigated and Rainfed Environments
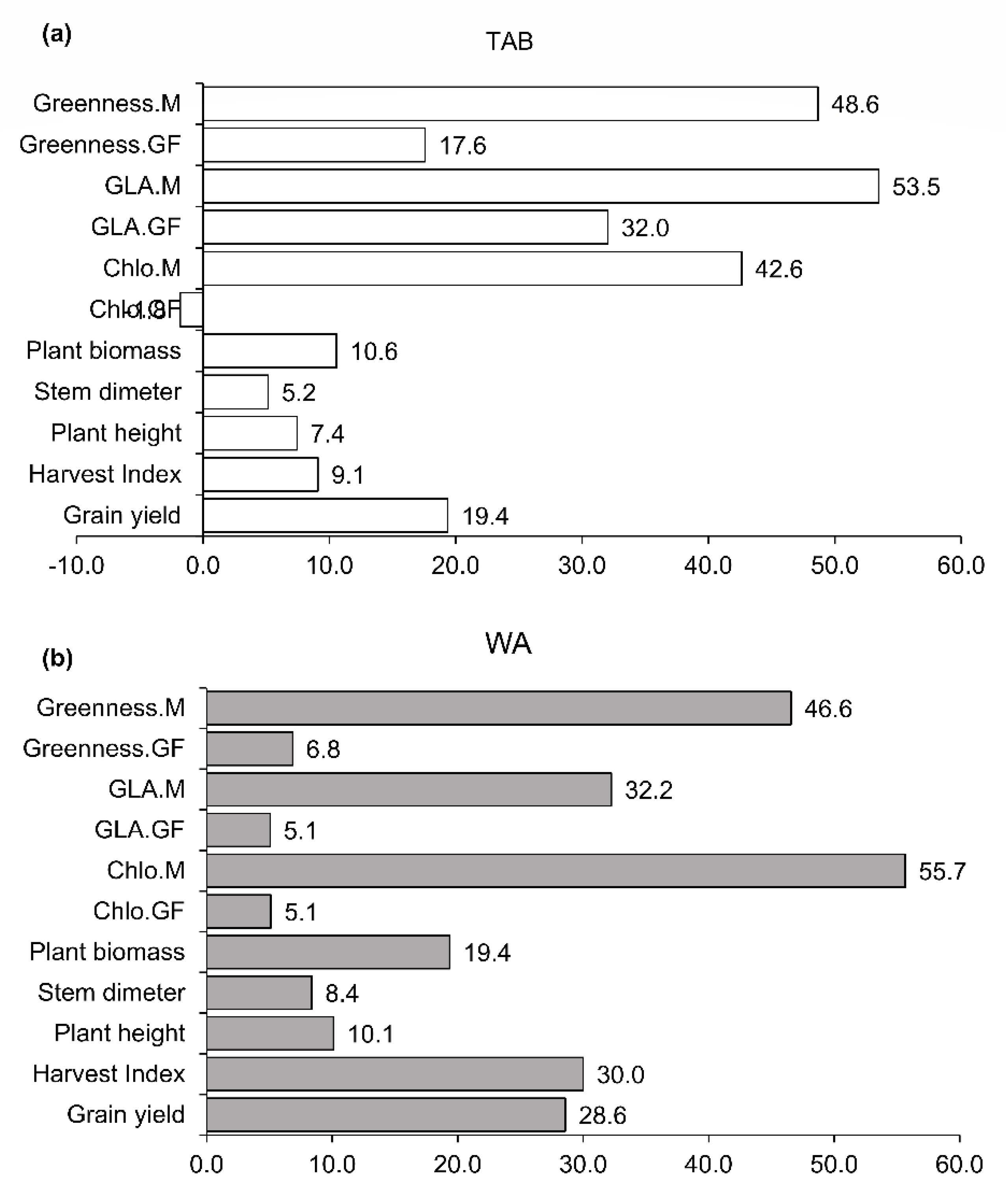
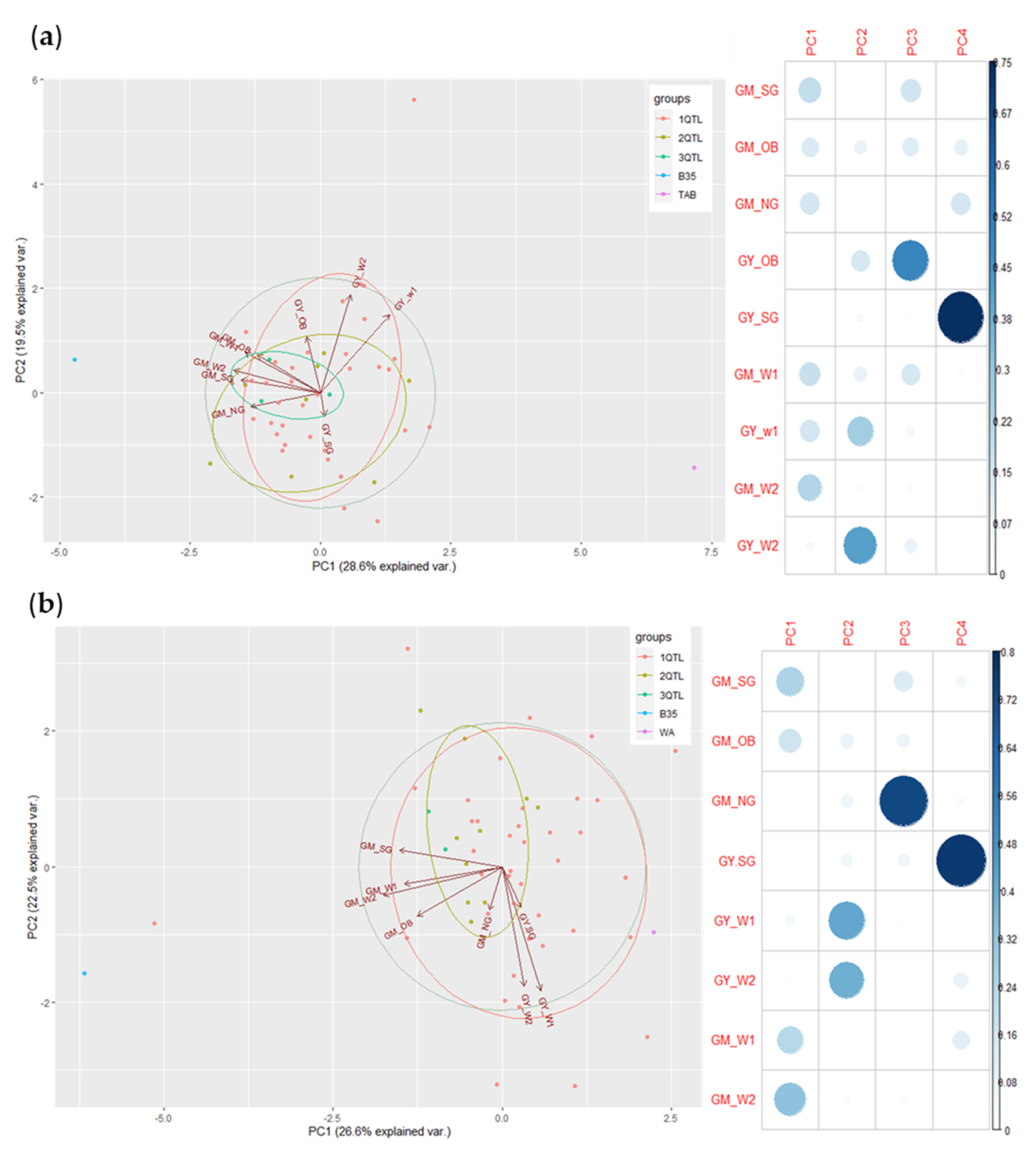
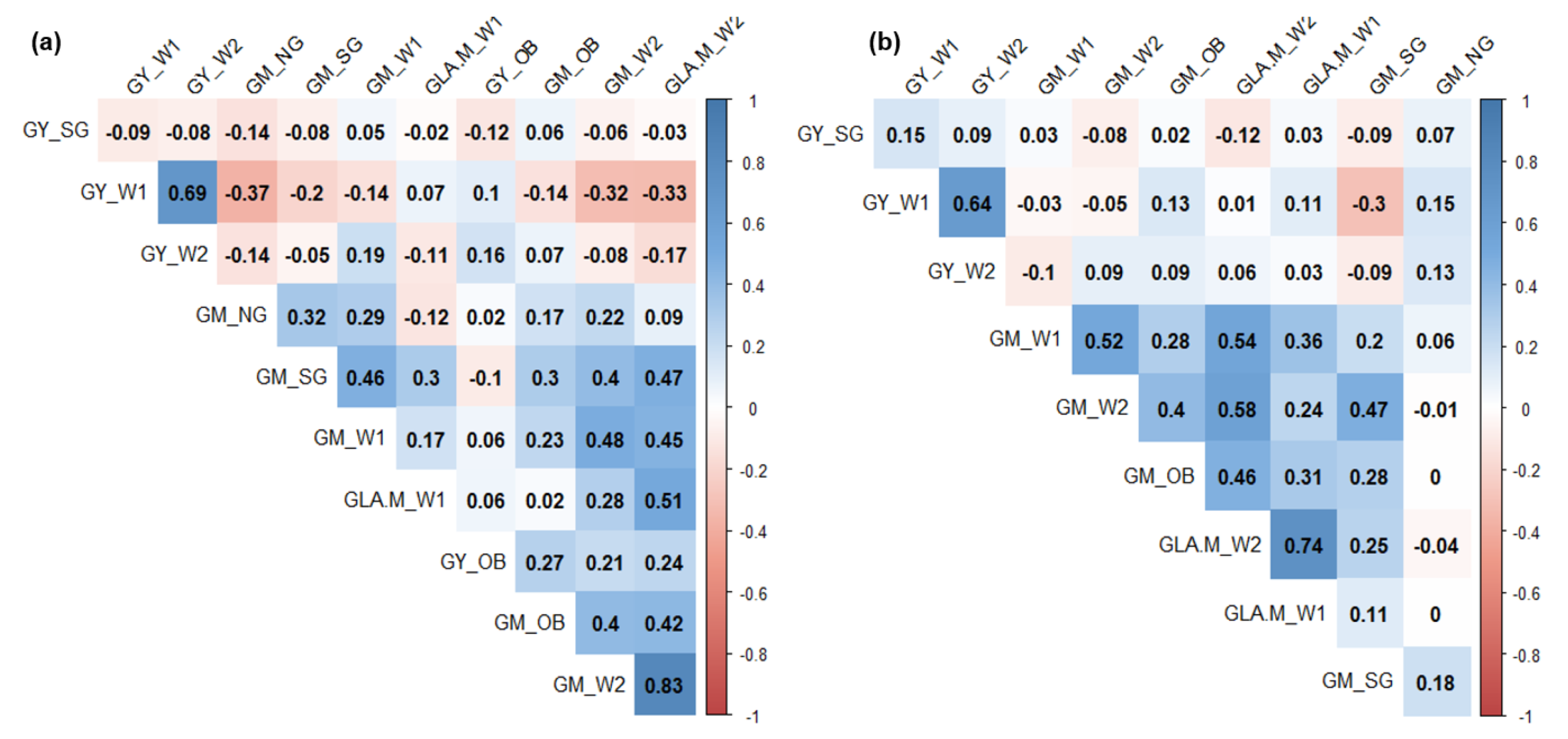
3.5. Principal Component Analysis and Pearson’s Correlation
4. Discussion
4.1. Effective C-MAB Strategy
4.2. Field Performance of Stay-Green Introgression Lines
4.3. The Cost of C-MAB
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MAS | Marker-assisted selection |
| MAB | Marker -assisted backcrossing |
| C-MAB | Combined marker-assisted backcrossing |
| QTL | Quantitative trait loci |
| NG | North Gedaref |
| SG | South Gedaref |
| OB | Al Obeid |
| W1 | Irrigated control experiment |
| W2 | Post-flowering drought |
| GY | Grain yield |
| GM | Percentage greenness at maturity |
| GGF | Percentage greenness at grain filling |
| Chlo.M | Chlorophyll content score at maturity |
| SD | Stem diameter |
| PH | Plant height |
| GY | Grain yield |
| PB | Plant biomass |
| HI | Harvest index |
| STI | Stress tolerance index |
| GLA.M | Green leaf area at maturity |
| R% | Reduction percentage |
References
- Voss-Fels, K.P.; Stahl, A.; Hickey, L.T. Q&A: Modern crop breeding for future food security. BMC Biol. 2019, 17, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ganal, M.W.; Röder, M.S. Microsatellite and SNP Markers in Wheat Breeding. In Genomics-Assisted Crop Improvement; Varshney, R.K., Tuberosa, R., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 1–24. [Google Scholar] [CrossRef]
- Shu, Y.; Yu, D.; Wang, D.; Bai, X.; Zhu, Y.; Guo, C. Genomic selection of seed weight based on low-density SCAR markers in soybean. Genet. Mol. Res. 2013, 12, 2178–2188. [Google Scholar] [CrossRef]
- Lande, R.; Thompson, R. Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics 1990, 124, 743–756. [Google Scholar] [CrossRef]
- Bohn, M.; Groh, S.; Khairallah, M.M.; Hoisington, D.A.; Utz, H.F.; Melchinger, A.E. Re-evaluation of the prospects of marker-assisted selection for improving insect resistance against Diatraea spp. in tropical maize by cross validation and independent validation. Theor. Appl. Genet. 2001, 103, 1059–1067. [Google Scholar] [CrossRef]
- Zhou, W.C.; Kolb, F.L.; Bai, G.H.; Domier, L.L.; Boze, L.K.; Smith, N.J. Validation of a major QTL for scab resistance with SSR markers and use of marker-assisted selection in wheat. Plant Breed. 2003, 122, 40–46. [Google Scholar] [CrossRef]
- Stam, P. Marker-assisted breeding. In Biometrics in Plant Breeding: Applications of Molecular Markers, Proceedings of the Ninth Meeting of the EUCARPIA Section Biometrics in Plant Breeding, Wageningen, The Netherlands, 6–8 July 1994; Van Ooijen, J.W., Jansen, J., Eds.; CPRO-DLO: Wageningen, The Netherlands, 1995; pp. 32–44. [Google Scholar] [CrossRef]
- Moreau, L.; Charcosset, A.; Gallais, A. Experimental evaluation of several cycles of marker-assisted selection in maize. Euphytica 2004, 137, 111–118. [Google Scholar] [CrossRef]
- Belícuas, P.R.; Aguiar, A.M.; Bento, D.A.V.; Câmara, T.M.M.; Junior, C.L.D.S. Inheritance of the stay-green trait in tropical maize. Euphytica 2014, 198, 163–173. [Google Scholar] [CrossRef]
- Sánchez, A.; Brar, D.; Huang, N.; Li, Z.; Khush, G. Sequence Tagged Site Marker-Assisted Selection for Three Bacterial Blight Resistance Genes in Rice. Crop. Sci. 2000, 40, 792–797. [Google Scholar] [CrossRef]
- Sharp, P.J.; Johnston, S.; Brown, G.; McIntosh, R.A.; Pallotta, M.; Carter, M.; Bariana, H.S.; Khatkar, S.; Lagudah, E.S.; Singh, R.P.; et al. Validation of molecular markers for wheat breeding. Aust. J. Agric. Res. 2001, 52. [Google Scholar] [CrossRef]
- Elagib, N.A.; Khalifa, M.; Rahma, A.E.; Babker, Z.; Gamaledin, S.I. Performance of major mechanized rainfed agricultural production in Sudan: Sorghum vulnerability and resilience to climate since 1970. Agric. For. Meteorol. 2019, 276-277, 107640. [Google Scholar] [CrossRef]
- Borrell, A.K.; Bidinger, F.R.; Sunitha, K. Stay-green associated with yield in recombinant inbred sorghum lines varying in rate of leaf senescence. Int. Sorghum Millets Newsl. 1999, 40, 31–34. [Google Scholar]
- George-Jaeggli, B.; Mortlock, M.Y.; Borrell, A.K. Bigger is not always better: Reducing leaf area helps stay-green sorghum use soil water more slowly. Environ. Exp. Bot. 2017, 138, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Kassahun, B.; Bidinger, F.R.; Hash, C.T.; Kuruvinashetti, M.S. Stay-green expression in early generation sorghum [Sorghum bicolor (L.) Moench] QTL introgression lines. Euphytica 2009, 172, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.K.; Kumari, M.; Singh, V.P.; Reddy, C.M.; Kumar, S.; Rane, J.; Chand, R. Stay green trait: Variation, inheritance and its association with spot blotch resistance in spring wheat (Triticum aestivum L.). Euphytica 2006, 153, 59–71. [Google Scholar] [CrossRef]
- Haussmann, B.I.G.; Mahalakshmi, V.; Reddy, B.V.S.; Seetharama, N.; Hash, C.T.; Geiger, H.H. QTL mapping of stay-green in two sorghum recombinant inbred populations. Theor. Appl. Genet. 2002, 106, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, K.; Subudhi, P.K.; Borrell, A.; Jordan, D.; Rosenow, D.; Nguyen, H.; Klein, P.; Klein, R.; Mullet, J. Sorghum stay-green QTL individually reduce post-flowering drought-induced leaf senescence. J. Exp. Bot. 2006, 58, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.M.; Kamal, N.M.; Noureldin, I.Y.; Hiraoaka, Y.; Yamauchi, Y.; Sujimoto, Y. Marker-assisted breeding of stay-green trait of sorghum to enhance terminal drought tolerance: Candidate donor and recipient genotypes. Sudan J. Agric. Res. 2007, 10, 133–141. [Google Scholar]
- Salih, A.A.; Ali, I.A.; Lux, A.; Luxová, M.; Cohen, Y.; Sugimoto, Y.; Inanaga, S. Rooting, Water Uptake, and Xylem Structure Adaptation to Drought of Two Sorghum Cultivars. Crop. Sci. 1999, 39, 168–173. [Google Scholar] [CrossRef]
- Tsuji, W.; Ali, M.; Inanaga, S.; Sugimoto, Y. Growth and Gas Exchange of Three Sorghum Cultivars Under Drought Stress. Biol. Plant. 2003, 46, 583–587. [Google Scholar] [CrossRef]
- Kamal, N.M.; Gorafi, Y.S.A.; Ghanim, A.M.A. Performance of sorghum stay-green introgression lines under post-flowering drought. Int. J. Plant Res. 2017, 7, 65–74. [Google Scholar] [CrossRef]
- Rosenow, D.; Quisenberry, J.; Wendt, C.; Clark, L. Drought tolerant sorghum and cotton germplasm. Agric. Water Manag. 1983, 7, 207–222. [Google Scholar] [CrossRef]
- Bhattramakki, D.; Dong, J.; Chhabra, A.K.; Hart, G.E. An integrated SSR and RFLP linkage map of Sorghum bicolor (L.) Moench. Genome 2000, 43, 988–1002. [Google Scholar] [CrossRef] [PubMed]
- Rosenow, D.T.; Clark, L.E.; Dahlberg, J.A.; Frederiksen, R.A.; Odvody, G.N.; Peterson, G.C.; Miller, F.R.; Woodfin, C.A.; Schaefer, K.S.; Collins, D.; et al. Release of Four A/B Sorghum Parental Lines ATx642 through ATx645. 2002. Available online: https://lubbock.tamu.edu/programs/crops/sorghum/release-proposal-for-four-ab-sorghum-parental-lines/ (accessed on 10 August 2020).
- Duvick, D.N.; Smith, J.S.C.; Cooper, M.E. Long-Term Selection in a Commercial Hybrid Maize Breeding Program. In Plant Breeding Reviews: Long-term Selection: Crops, Animals, and Bacteria; Wiley: Hoboken, NJ, USA, 2010; Volume 24, Pt 2, pp. 109–151. [Google Scholar] [CrossRef]
- Câmara, T.M.M.; Bento, D.A.V.; Alves, G.F.; Santos, M.F.; Moreira, J.U.V.; Souza, C.L.J. Genetic parameters of drought tolerance related traits in tropical maize. Bragantia 2007, 66, 595–603. [Google Scholar] [CrossRef]
- Costa, E.F.N.; Santos, M.F.; Moro, G.V.; Alves, G.F.; Souza, C.L.J. Inheritance of the delayed senescence in maize. Pesq. Agropec. Bras. 2008, 43, 207–213. [Google Scholar] [CrossRef]
- Ragheb, S.M.; Yassin, A.S.; Amin, M.A. The Application of Uniplex, Duplex, and Multiplex PCR for the Absence of Specified Microorganism Testing of Pharmaceutical Excipients and Drug Products. PDA J. Pharm. Sci. Technol. 2012, 66, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Nikiforova, M.N.; LaFramboise, W.A.; Nikiforov, Y.E. Amplification-Based Methods. Clin. Genom. 2015, 57–67. [Google Scholar] [CrossRef]
- Mahalakshmi, V.; Bidinger, F.R. Evaluation of stay-green sorghum germplasm lines at ICRISAT. Crop Sci. 2002, 42, 965–974. [Google Scholar]
- Xu, W.; Rosenow, D.T.; Nguyen, H.T. Stay green trait in grain sorghum: Relationship between visual rating and leaf chlorophyll concentration. Plant Breed. 2000, 119, 365–367. [Google Scholar] [CrossRef]
- Lema, M. Marker Assisted Selection in Comparison to Conventional Plant Breeding: Review Article. Agric. Res. Technol. Open Access J. 2018, 14. [Google Scholar] [CrossRef] [Green Version]
- Boughton, D.; Crawford, E.; Krause, M.; Henry, D.B. Economic Analysis of On-Farm Trials: A Review of Approaches and Implications for Research Program Design. Staff Paper No. 90-78; Department of Agricultural, Food and Resource Economics, Michigan State University: East Lansing, MI, USA, 1990. [Google Scholar]
- Ribaut, J.-M.; Hoisington, D. Marker-assisted selection: New tools and strategies. Trends Plant Sci. 1998, 3, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Christopher, J.T.; Christopher, M.J.; Borrell, A.K.; Fletcher, S.; Chenu, K. Stay-green traits to improve wheat adaptation in well-watered and water-limited environments. J. Exp. Bot. 2016, 67, 5159–5172. [Google Scholar] [CrossRef] [Green Version]
- Cerrudo, D.; Cao, S.; Yuan, Y.; Martinez, C.; Suarez, E.A.; Babu, R.; Zhang, X.; Trachsel, S. Genomic Selection Outperforms Marker Assisted Selection for Grain Yield and Physiological Traits in a Maize Doubled Haploid Population Across Water Treatments. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Moreau, L.; Charcosset, A.; Hospital, F.; Gallais, A. Marker-Assisted Selection Efficiency in Populations of Finite Size. Genetics 1998, 148, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Ribaut, J.-M.; Ragot, M. Marker-assisted selection to improve drought adaptation in maize: The backcross approach, perspectives, limitations, and alternatives. J. Exp. Bot. 2006, 58, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Valencia, R.C.; Rooney, W.L. Genetic Control of Sorghum Grain Color. INTSORMIL Presentations. Part of the Agronomy and Crop Sciences Commons, and the Plant Breeding and Genetics Commons. 2009. Available online: https://digitalcommons.unl.edu/intsormilpresent/10?utm (accessed on 15 August 2020).
- Moose, S.P.; Mumm, R.H. Molecular Plant Breeding as the Foundation for 21st Century Crop Improvement. Plant Physiol. 2008, 147, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R. Marker assisted selection. In Plant breeding reviews: Part 1: Long-Term Selection: Maize; Janick, J., Ed.; Wiley: Hoboken, NJ, USA, 2004; Volume 24, pp. 293–310. [Google Scholar]
- Reddy, N.R.R.; Ragimasalawada, M.; Sabbavarapu, M.M.; Nadoor, S.; Patil, J.V. Detection and validation of stay-green QTL in post-rainy sorghum involving widely adapted cultivar, M35-1 and a popular stay-green genotype B35. BMC Genom. 2014, 15, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subudhi, P.K.; Rosenow, D.T.; Nguyen, H.T. Quantitative trait loci for the stay green trait in sorghum (Sorghum bicolor L. Moench): Consistency across genetic backgrounds and environments. Theor. Appl. Genet. 2000, 101, 733–741. [Google Scholar] [CrossRef]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Reyes, B.G.D.L. Genomic and epigenomic bases of transgressive segregation–New breeding paradigm for novel plant phenotypes. Plant Sci. 2019, 288. [Google Scholar] [CrossRef]
- Kamal, N.M.; Gorafi, Y.S.A.; Tsujimoto, H.; Ghanim, A.M.A. Stay-Green QTLs Response in Adaptation to Post-Flowering Drought Depends on the Drought Severity. BioMed Res. Int. 2018, 2018, 1–15. [Google Scholar] [CrossRef]
- Kamal, N.M.; Gorafi, Y.S.A.; Abdelrahman, M.; Abdellatef, E.; Tsujimoto, H. Stay-Green Trait: A Prospective Approach for Yield Potential, and Drought and Heat Stress Adaptation in Globally Important Cereals. Int. J. Mol. Sci. 2019, 20, 5837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Subudhi, P.K.; Crasta, O.R.; Rosenow, D.T.; Mullet, J.E.; Nguyen, H.T. Molecular mapping of QTLs conferring stay-green in grain sorghum (Sorghum bicolor L. Moench). Genome 2000, 43, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Drought adaptation in cereal crops a prologue. In Drought Tolerance in Cereals; Ribaut, J.-M., Ed.; The Haworth Press Inc.: Binghamtown, NY, USA, 2006; pp. 10–18. [Google Scholar]
- Van Berloo, R.; Stam, P. Simultaneous marker-assisted selection for multiple traits in autogamous crops. Theor. Appl. Genet. 2001, 102, 1107–1112. [Google Scholar] [CrossRef]
- Abalo, G.; Tongoona, P.; Derera, J.; Edema, R. A Comparative Analysis of Conventional and Marker-Assisted Selection Methods in Breeding Maize Streak Virus Resistance in Maize. Crop. Sci. 2009, 49, 509–520. [Google Scholar] [CrossRef]
- Dreher, K.; Khairallah, M.; Ribaut, J.-M.; Morris, M. Money matters (I): Costs of field and laboratory procedures associated with conventional and marker-assisted maize breeding at CIMMYT. Mol. Breed. 2003, 11, 221–234. [Google Scholar] [CrossRef]
- Morris, M.; Dreher, K.; Ribaut, J.-M.; Khairallah, M. Money matters (II): Costs of maize inbred line conversion schemes at CIMMYT using conventional and marker-assisted selection. Mol. Breed. 2003, 11, 235–247. [Google Scholar] [CrossRef]
- Jorasch, P. Intellectual Property Rights in the Field of Molecular Marker Analysis. In Molecular Marker Systems in Plant Breeding and Crop Improvement. Biotechnology in Agriculture and Forestry; Lorz, H., Wenzel, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 55, pp. 433–471. [Google Scholar]
- Brennan, J.; Rehman, A.; Raman, H.; Milgate, A.W.; Pleming, D.; Martin, P.J. An economic assessment of the value of molecular markers in plant breeding programs. In Proceedings of the 49th Annual Conference of the Australian Agricultural and Resource Economics Society, Coffs Harbour, Australia, 9–11 February 2005; pp. 1–14. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Expenses/Season | Amounts/Units | Cost per Unit (US$) | Total Cost (US$) |
|---|---|---|---|
| Field fertilizer | 1 bag | 70 | 70 |
| Field supplies | 10 irrigations | 5 | 50 |
| Labor | 2 persons/day for 4 months | 10 | 1200 |
| Total | 1320 | ||
| Total Field cost for 3 cycles | 3960 | ||
| Laboratory Expenses | Cost per Sample (US$) | No. of Samples | Total Cost (US$) |
| DNA extraction | 0.71 | 2850 | 2023.5 |
| PCR analysis | 0.08 | 20,800 | 1664 |
| Electrophoresis and gel visualization | 0.038 | 20,800 | 790.4 |
| Labor | 0.05 | 20,800 | 1040 |
| Total laboratory Cost | 5517.9 | ||
| Total Field cost | 3960 | ||
| Field and Laboratory expenses | 9477.9 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamal, N.M.; Gorafi, Y.S.A.; Abdeltwab, H.; Abdalla, I.; Tsujimoto, H.; Ghanim, A.M.A. A New Breeding Strategy towards Introgression and Characterization of Stay-Green QTL for Drought Tolerance in Sorghum. Agriculture 2021, 11, 598. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070598
Kamal NM, Gorafi YSA, Abdeltwab H, Abdalla I, Tsujimoto H, Ghanim AMA. A New Breeding Strategy towards Introgression and Characterization of Stay-Green QTL for Drought Tolerance in Sorghum. Agriculture. 2021; 11(7):598. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070598
Chicago/Turabian StyleKamal, Nasrein Mohamed, Yasir Serag Alnor Gorafi, Hanan Abdeltwab, Ishtiag Abdalla, Hisashi Tsujimoto, and Abdelbagi Mukhtar Ali Ghanim. 2021. "A New Breeding Strategy towards Introgression and Characterization of Stay-Green QTL for Drought Tolerance in Sorghum" Agriculture 11, no. 7: 598. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11070598