1. Introduction
Rice (
Oryza sativa) is the principal dietary component for more than 50% of the world’s population [
1]. Global cereal production is expected to increase by 13% by the year 2027. Rice production is forecasted to increase by 64 metric tons (Mt) to 562 Mt to satisfy the dietary calorie requirements of the population [
2]. For decades, conventional rice production has relied heavily on the use of chemical fertilizer (CF) to obtain the desired crop growth and yield output. The expected increase in rice requirement will further increase the need for CF during the cultivation phase for rice crop. This raises concern as plants do not absorb all nutrients supplied by CF. Plants absorb essential macronutrients such as phosphate ions (HPO
42− and H
2PO
4−), nitrate (NO
3−), and ammonium (NH
4+). Pollution is known to occur when these elements discharge through leaching, mineralization, or volatilization, a situation that is further amplified through rapid degradation of the soil structure during the intensive rice cropping cycle [
3,
4,
5].
Studies that have investigated the use of microorganisms as an alternative to conventional methods of supplying nutrients to plants have recently gained widespread interest [
6,
7,
8,
9]. Past experiments revealed complex interactions between these microorganisms and plants that may conceivably allow reduction in the use of CF in rice production [
10,
11].
Plants have a limited ability to genetically adapt to rapid environmental changes (heat, drought, toxins, or insufficient nutrients) due to their short life cycles. To assist them, plants have complex interactions with microorganisms present in their rhizosphere [
12,
13,
14,
15,
16]. Most microorganisms have no direct importance for plant growth and vigor. Nevertheless, there are a few microbe communities that are beneficial for plants. These microorganisms were termed plant-growth-promoting rhizobacteria (PGPR) by Kloepper and Schroth [
17]. PGPR is known to stimulate the synthesis of plant growth regulators, phytohormones, and many biologically active substances in the host plant. Moreover, previous studies showed that specific species have the potential to accrue and alter nutrients for plant uptake [
18,
19,
20,
21,
22].
Nguyen et al. [
23] demonstrated that replacement of 50% of the required nitrogen (N) fertilizer with two
Bacillus spp. and an
Azospirillus sp. in wheat (
T. aestivum) resulted in vigorous vegetative growth with an increased biomass weight. Similarly, Yang et al. [
15] applied
Sphingomonas to
Dendrobium officinale and observed increases of 8.6% in the plant culm structure and 7.5% in its fresh weight. Castanheira et al. [
12] also tested the same species of bacteria by applying three different strains along with N and phosphorous (P) supplements to ryegrass (
Lolium spp.) and observed improvement in the overall plant growth. These studies demonstrated that the bacterium infiltrated the plant roots and stem tissues, triggering rigorous plant growth and development. Furthermore, the plants’ overall physiological growth and development were more favorable when two or more PGPR strains were combined with CF. This also resulted in improvements in the crop productivity as well as a reduction in fertilizer usage. For instance, Shaharoona et al. [
11] studied wheat inoculation with two species of
Pseudomonads (
P. fluorescens and
P. fluorescens biotype F) with different levels of nitrogen, phosphorous, and potassium (NPK). The inoculated plants showed an increase in yield of up to 22% compared with crops cultivated with NPK alone. Besides the yield, there were significant increases in root development and the overall biomass production.
Preliminary studies have demonstrated that amending CF with PGPR reduced the demand of rice plants for synthetic fertilizer without compromising vegetative growth or the grain yield output [
24,
25,
26,
27]. Tarigan et al. [
26] and Tarigan et al. [
27] further reported that with the presence of PGPR, the availability of NO
3− and NH
4+ was maintained throughout the crop growth cycle to a greater extent than in fields farmed using CF alone. Such information is vital for effectively reducing the use of synthetic fertilizers. However, the application of PGPR during those studies was confined to growing conditions with ample irrigation. Besides maintaining soil moisture, irrigation aids with the even dispersal of nutrients. At present, limited studies have reported on the application and response of rice cultivated with a combination of synthetic fertilizer and PGPR under alternate wetting and drying (AWD) cultivation. Therefore, a study was undertaken to investigate the effects of replacing CF with PGPR on vegetative growth, biomass production, and grain yield in rice plants cultivated under AWD cultivation. Furthermore, the study aimed to determine an optimal reduction in CF dose when incorporated with PGPR for application in rice cultivated under AWD.
4. Discussion
According to the literature, the three bacteria species
B. aryabhattai, B. ambifaria, and
S. yanoikuyae are considered PGPR based on their interactions with and beneficial influences on host plants. The bacterial strain
B. aryabhattai belongs to the
Bacillus genus of the Bacillaceae family. This genus boasts some well-known PGPR species [
19,
37,
38,
39]. Park et al. [
40] found that the strain was able to trigger production of the phytohormone abscisic acid (ABA). The bacterial strain
B. ambifaria belongs to the
Proteobacteria genus. Similar to
Bacillus, it contains some known PGPR strains used in the field of agriculture [
10,
19,
41,
42,
43,
44]. Parra-Cota et al. [
10] showed that
B. ambifaria has the potential to be used as a PGPR. The study found that the bacteria strain was able to improve the vegetative growth and yield production of two species of amaranth (
Amaranthus cruentus and
A.
hypochondriacus). The bacterial strain
S. yanoikuyae belongs to the
Sphingomonas genus and has medical, industrial, and agronomical importance [
45,
46,
47]. According to Yang et al., the strain was found to promote rigorous cell wall development along the culm of
Dendrobium officinale. A study by Hoo et al. [
48] determined that the bacterial strain was able to restore magnesium ion (Mg
2+) levels along the rhizoplane for plant cell growth.
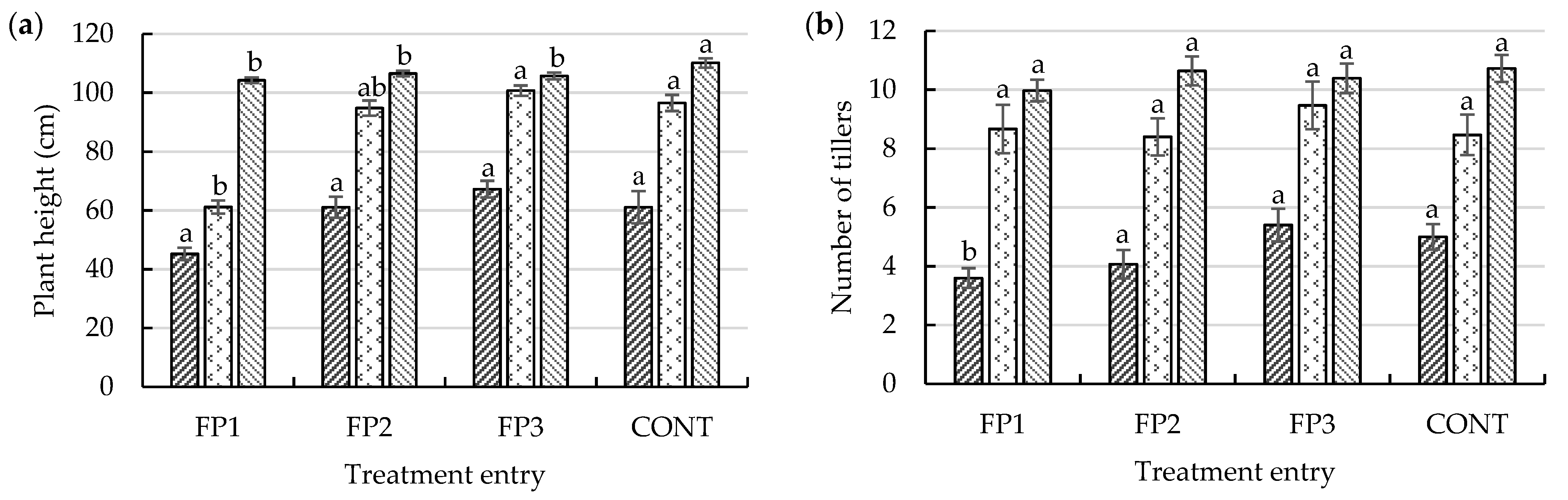
Throughout the growth cycle, the effects across treatments on vegetative parameters were minimal. A notable effect of treatment was observed in plant height, particularly at the FS and MS (
Figure 3a). The height of KH-147 at maturity ranges 90–125 cm. According to the International Rice Research Institute (IRRI) [
49], it is intermediate in height. This enables maximum exposure of the crop’s vegetative canopy to direct sunlight for photosynthetic activities. Rice varieties with tall plant heights are disadvantaged in some situations as they are vulnerable to lodging. An increased plant height can also result from excess N fertilizer application [
50]. N is fundamental for plant growth and development; however, it also encourages rapid elongation of the cell walls at the culm structure. It may weaken the resilience of crops to other forms of biotic and abiotic stress [
51]. As such, lodging can lead to severe crop loss and poor grain quality [
52,
53]. Regardless, plant height is still an important trait that affects the potential rice yield [
54].
The average plant height at the MS was 105.46 cm (
Figure 3a). Application of a high concentration of CF during the early growth stages may have led to increased heights in control plants (110.11 cm). Plants treated with amended doses had slower growth paces. The results indicate that plants exposed to PGPR could have accrued sturdy cells, resulting in slow growth. Previously, studies showed that plants inoculated with
Burkholderia spp. and
Sphingomonas spp. have thick culms and robust cell structures [
10,
15]. The presence of bacterial strains in plant tissues also improved the plants metabolic processes, which eventually improves their physiological growth [
12].
The replacement of CF with PGPR showed no effects on tiller growth during the HS and MS (
Figure 3b). The only notable effect of treatment on tiller production occurred during the ATS period. Both CONT (5.33) and FP3 (5.39) produced an average of five tillers at ATS, while the effect of a high CF input at the early active growth stage was visible. This may be advantageous but could also result in the plants being vulnerable to pest infestation and lodging [
51]. Sample plants exposed to CF responded swiftly to the nutrients supplied during the basal and ATS phases, but the number of tillers then subsided once treatment with CF ceased. Plants treated with CONT produced the highest number of tillers at the FS (9.56) and MS (10.72). However, no statistical differences in the tiller number were observed between treatments during the respective growth stages. According to Nguyen et al. [
23] and Akram et al. [
55], the tiller number of monocot plants such as rice is less (significantly) affected by most treatments than the crop height and other vegetative parameters. Furthermore, it is also possible that plants treated with some amount of PGPR may form resilient cell structures and have better metabolic processes along the culm area [
10,
15]. As such, the plants have a better chance of withstanding external stresses, thus ensuring their survival and enhancing plant productivity.
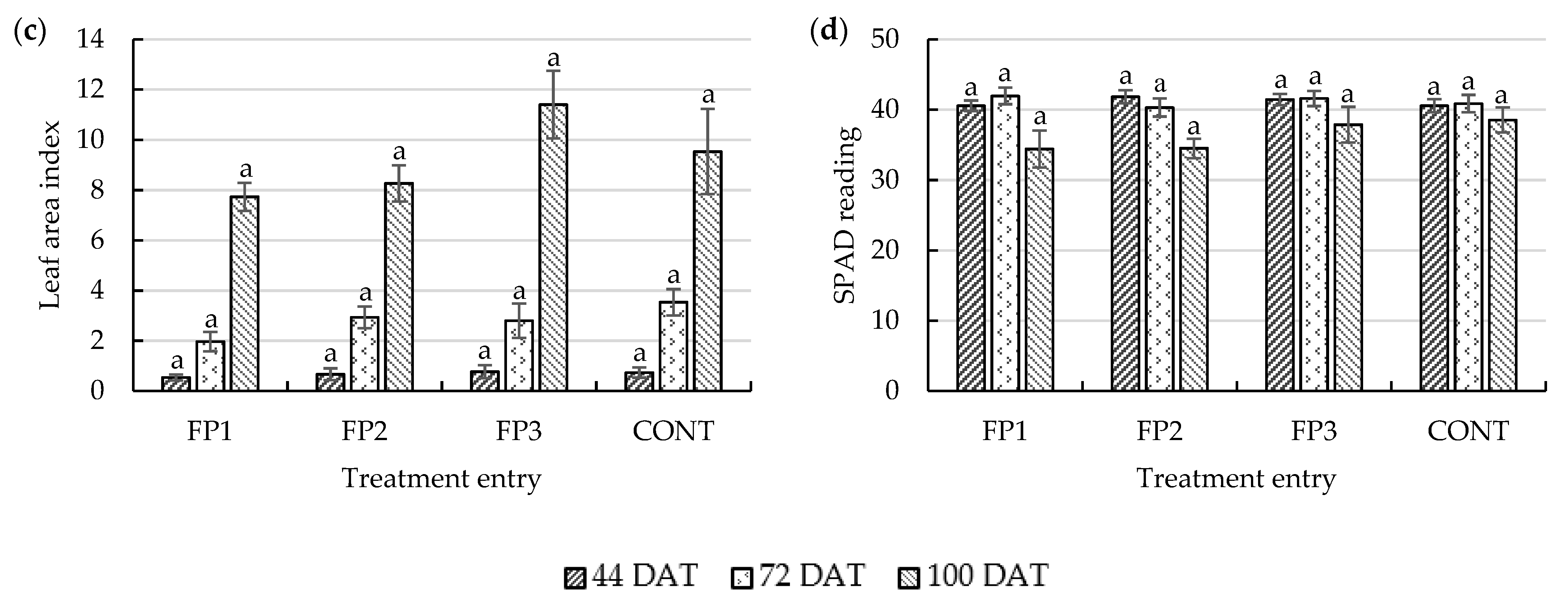
Assessment of the LAI at different growth stages is paramount, as it can be used to predict the potential yield [
56,
57]. An increase in the LAI of rice plants can result in an increase in their ability to intercept light [
34]. The results showed no significant difference (
p ≤ 0.05) in the LAI of treated plants in this study. Throughout the initial growth stage, rice plants treated with a combination of CF and PGPR attained higher SPAD readings than those of CONT rice plants (40.56). The average leaf greenness of plants treated with FP2 (41.83) was highest at the ATS and then matched by FP1 (41.94) at the FS. From ripening to maturity, CONT (38.53) had the highest SPAD reading. The chlorophyll content fluctuated during the ATS and FS. Plants with high levels of chlorophyll during these two periods had greater photosynthetic potential, a critical factor in rice grain development. This theory was proposed based on the biomass production and dry matter partitioning observed in this study (see
Table 3 and
Table 4). According to Tian et al. [
58], a sufficient LAI is necessary for dry matter production. However, with a reduction in the amount of CF, the insignificant difference between the LAI and crop greenness demonstrates the potential of PGPR to adequately maintain typical crop growth throughout the crop’s life cycle. Rice grain yield is a result of the combination of various processes, such as photosynthetic activity from the crop canopy and the portioning and conversion of assimilates to biomass [
59,
60,
61]. Besides growth maintenance, the presence of PGPR may have contributed to the improvement of yield components (see below).
As observed in
Table 3, the crops responded to the presence of PGPR with a significant increase (
p ≤ 0.05) during the early growth stages, as reflected by the SDW and LDW. The increased dry weights of these respective plant organs indicate the effects of
B. ambifaria and
S. yanoikuyae in the PGPR treatment. Plants inoculated with
Sphingomonas spp. and
Burkholderia spp. showed increased growth. The improvement in the vascular system by these PGPR species aids in the transport of water and nutrients from the plant roots to the aboveground plant biomass [
10,
12,
15,
62]. Another contributing strain is
B. aryabhattai. Previous studies reported that it could positively influence both plant DMW and grain yield production [
23,
63]. According to Radhakrishnan et al. [
64], the
Bacillus spp. is a unique and economically useful PGPR strain. It is known to vary in strength and effectiveness and may trigger multiple effects when in contact with the plant. It could break down soil carbon and mineralize insoluble soil nutrients for plant uptake. The availability of N and P ions for uptake from soil is then increased along with traces of minor elements, such as magnesium (Mg
2+), iron (Fe
2+), zinc (Zn
2+), and chelates, when inoculated with
Bacillus spp. The activities of these microorganisms may have improved the crop biomass production, as indicated by the increase in the crops dry weight.
The PFG for FP2 plants (83.99%) was significantly greater than that of control plants (79.07%). According to Castanheira et al. [
12] and Elekhtyar [
65], the microbes present in the plant tissue may assist the crop in the mobilization of nutrients and carbohydrates accumulated during the grain-filling period: the higher the PFG, the greater the rice grain yield at the maturity stage. The 1000-GW displayed a similar outcome. The results show that plants treated with a combination of fertilizer and PGPR had a significantly increased (
p ≤ 0.05) 1000-GW. The results correspond to those of Alam et al. [
66], who found similar outcomes when testing different combinations of bacterial strains (
Azotobacter spp.,
Bacillus spp.,
Enterobacter spp., and
Xanthobacter spp.). Plants exposed to FP3 (27.18 g), FP2 (26.93 g), and FP1 (26.50 g) gained statistically higher 1000-GW values than plants under the CONT (25.74 g) treatment. The results indicate that rice plants inoculated with PGPR have ample plant heights, low numbers of tillers, and consistent chlorophyll contents (SPAD reading) during the critical growth stages, contributing to statistically higher PFG and 1000-GW values.
The grain yield is the final indicator of crop performance for the different treatments. The combined effects of PGPR and CF in FP2 produced a 14.41% increase in performance compared with plants treated with the conventional method (CONT). The results from this study are similar to those obtained by Parra-Cota et al. [
10], Shaharoona et al. [
11], and Islam et al. [
67]. The results for the yield component, PFG, and 1000-GW showed evidence that the presence of PGPR improved the crop performance by 4–5% compared with the control treatment (
Table 5). According to Xie et al. [
68] and Huang et al. [
69], there is a close relationship between the two yield components. Both the grain size and grain filling rate were used to determine the 1000-GW.
The rice yield is known to be regulated by both genetic and external environmental factors [
54,
70,
71]. The number of tillers produced by a rice plant is closely related to the number of panicles per hill [
35]. However, the negative correlation indicates that high tiller production may significantly reduce the grain yield, which is a trait limited to the specific rice variety. Presumably, the conventional CF dosage may cause plants to prioritize tiller production well beyond the FS while failing to shift their efforts towards grain development. As per the results shown in
Figure 3b, plants treated with all CF doses showed an increase in their respective tiller numbers, except for those treated with FP1. However, plants treated with PGPR had significant increases (
p ≤ 0.05) in their PFG and 1000-GW (
Table 3). As explained by Castanheira et al. [
12], the microbial strains present inside a plant’s tissues aid in efficiently mobilizing nutrients and carbohydrates, which may have contributed to the improvements in the two yield components.
The LAI indicates a crop’s ability to intercept radiation and precipitation, convert energy, and maintain the water balance [
72,
73]. As such, it is an agronomic parameter commonly assessed for predicting grain production [
57]. The correlation results indicate accession in the LAI significantly improves the grain yield (r = 0.65 ***). An increase in the LAI will improve grain yield production [
74,
75,
76]. The canopy of leaves over a plant allows much-needed sunlight to be intercepted to drive the plant growth and rice grain development. However, the extent to which these processes occur also depends on the height of the plant. The heights of rice plants in this study demonstrated intermediate characteristics that may have contributed towards the positive correlation between the grain yield and LAI (
Figure 3a).
The correlation between the grain yield and the number of panicles per hill indicates a reduction in the grain yield output. This may be based on the genetic traits of the rice variety rather than on the treatment effect. The No. Panicle/hill is a vital grain yield component, as it reflects the effects of both the environment and cultivation practice used [
77]. A literature review suggested that improvements in crop management are generally associated with an increase in the No. Panicle/hill [
78,
79].
The correlation between grain yield and the 1000-GW (r = 0.23 *) indicated a positive relationship. However, according to Yang and Zhang [
80], the yield component 1000-GW remains constant across different environments for some rice cultivars. In our study, the application of PGPR influenced the 1000-GW (
Table 5). This is an essential contribution toward an improvement in grain yield. These improvements can be attributed to the combination of CF and PGPR, thus further confirming an advantage of incorporating them both into the rice farming systems.
This study observed a significant treatment effect (
p ≤ 0.05) on biomass production across all treatment groups during the FS (
Table 3) and particularly for plants exposed to PGPR treatment. According to Wu et al. [
59] and Katsura et al. [
81], the grain yield increases as the biomass weight increases. Katsura et al. [
81] further mentioned that dry matter accumulation before the FS greatly influences the grain yield output. Although our analysis indicates no statistical difference at the MS, an increase in the total DMW at FS contributed considerably to the respective PFG (
p ≤ 0.05) and 1000-GW (
p ≤ 0.01) values (refer to
Table 3 and
Table 5), which ultimately resulted in higher grain yields (
Table 6). The positive interaction between the grain yield and DMW (r = 0.22 *) further justifies this hypothesis (
Table 8).
The HI values in this study correspond to those reported by Parra-Cota et al. [
10]. A combination of both N fertilizer and PGPR was able to increase the crop HI. The assessment of plant HI indicated a highly significant difference (
p ≤ 0.01) between treatments (
Table 6). However, its interaction with grain yield production showed no significant effect (
p ≤ 0.05) (
Table 8). Regardless, rice grain yield production is the outcome of the biomass yield and HI, and if either or both are affected by the replacement of CF with PGPR, an increase in grain yield will occur [
30,
61,
82]. This study only observed an increase in biomass weight in plants treated with PGPR. These improvements in biomass production may have influenced the HI. The results of this study show that PGPR aid rice plant growth as it is being reflected the by increased in biomass production thereby improving grain production.








{kind=link}
{kind=link}
{kind=link}
{kind=link}