
Durum Wheat Mediterranean Landraces: A Valuable Source for Resistance to Tan Spot Disease

 ,
,
Abstract
:1. Introduction:
2. Materials and Methods
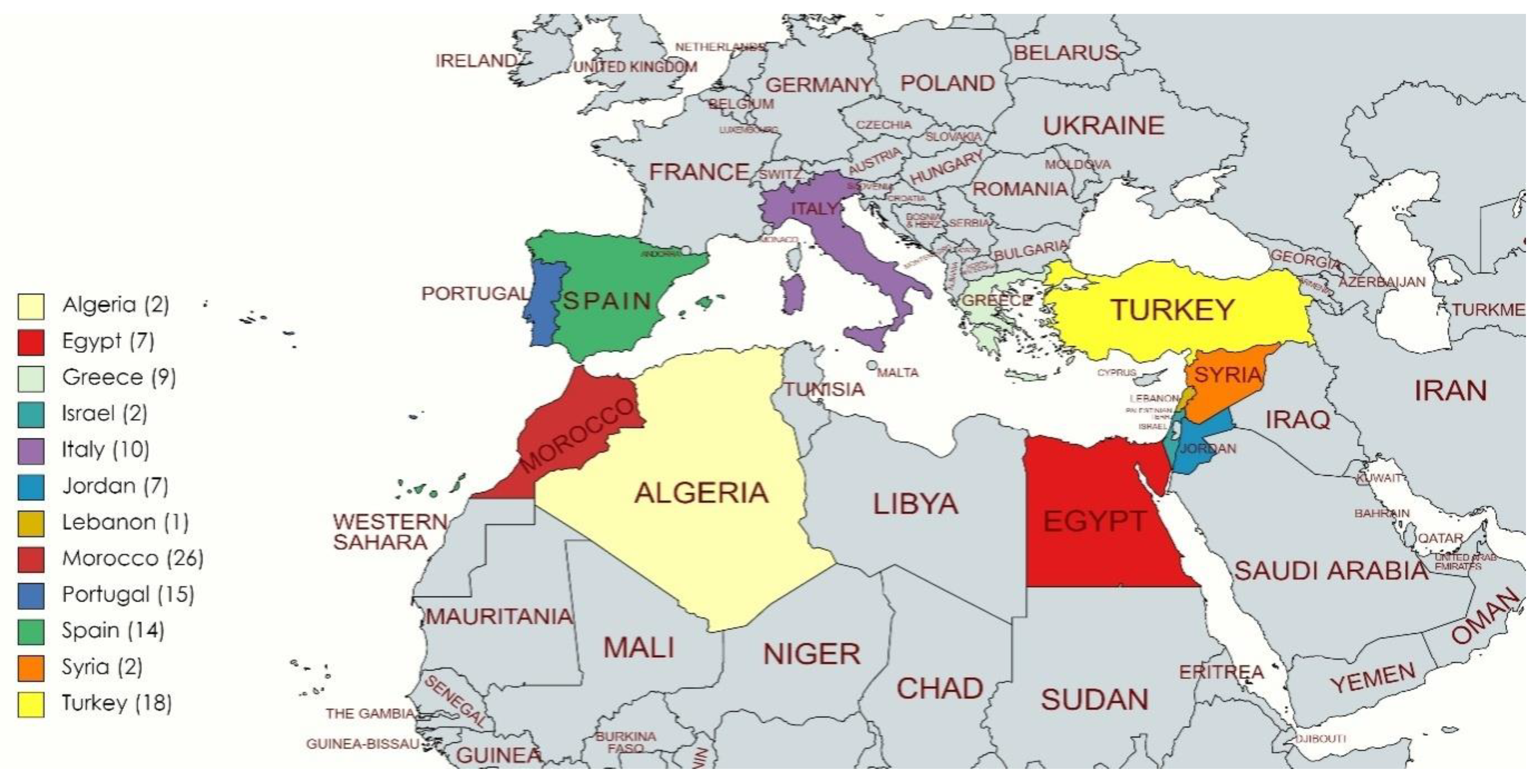
2.1. Plant Materials
2.2. Experimental Design and Inoculation Method
2.3. Agronomic Traits
2.4. Disease Severity Rating
2.5. Statistical Analysis
3. Results and Discussion
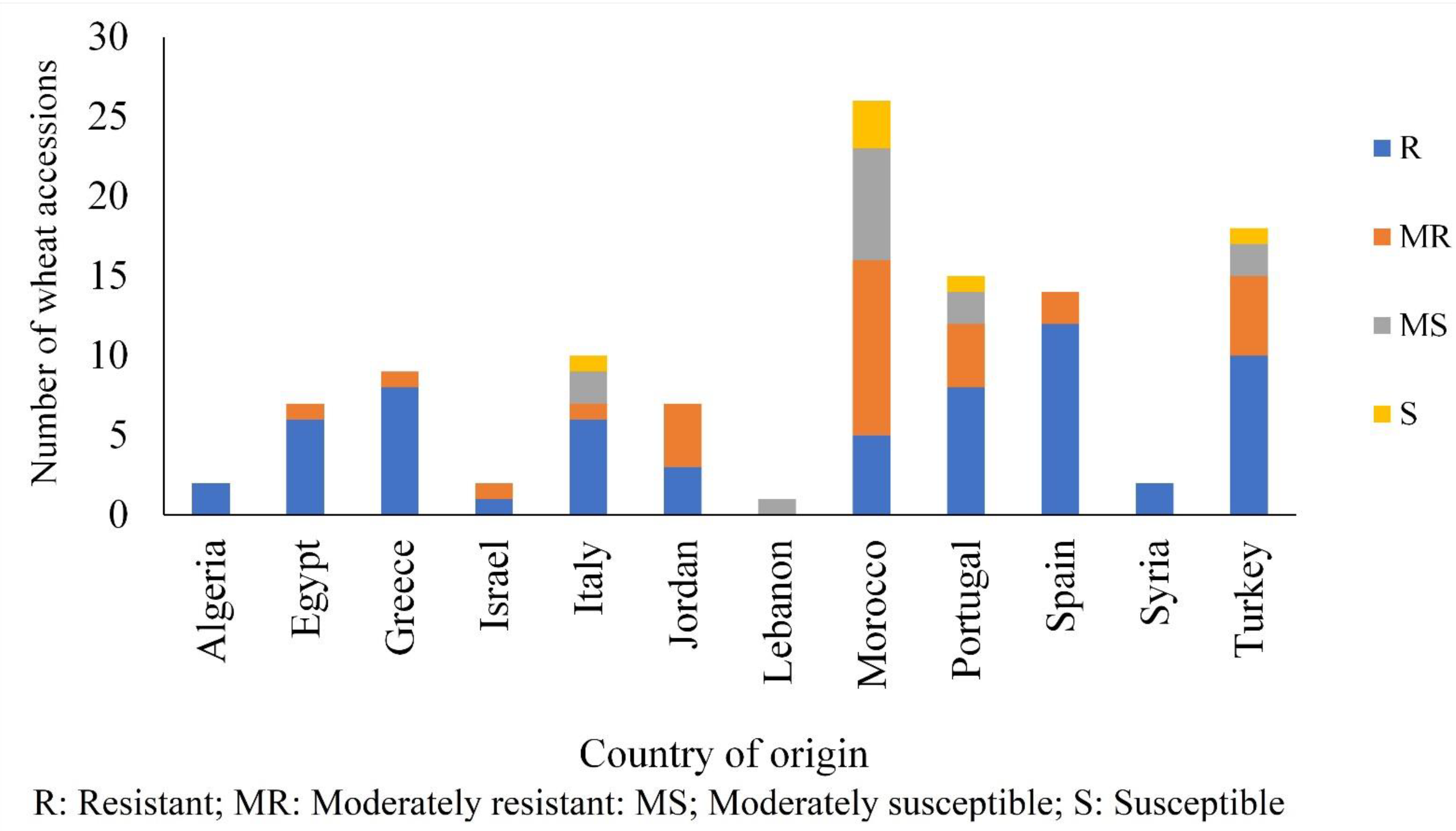
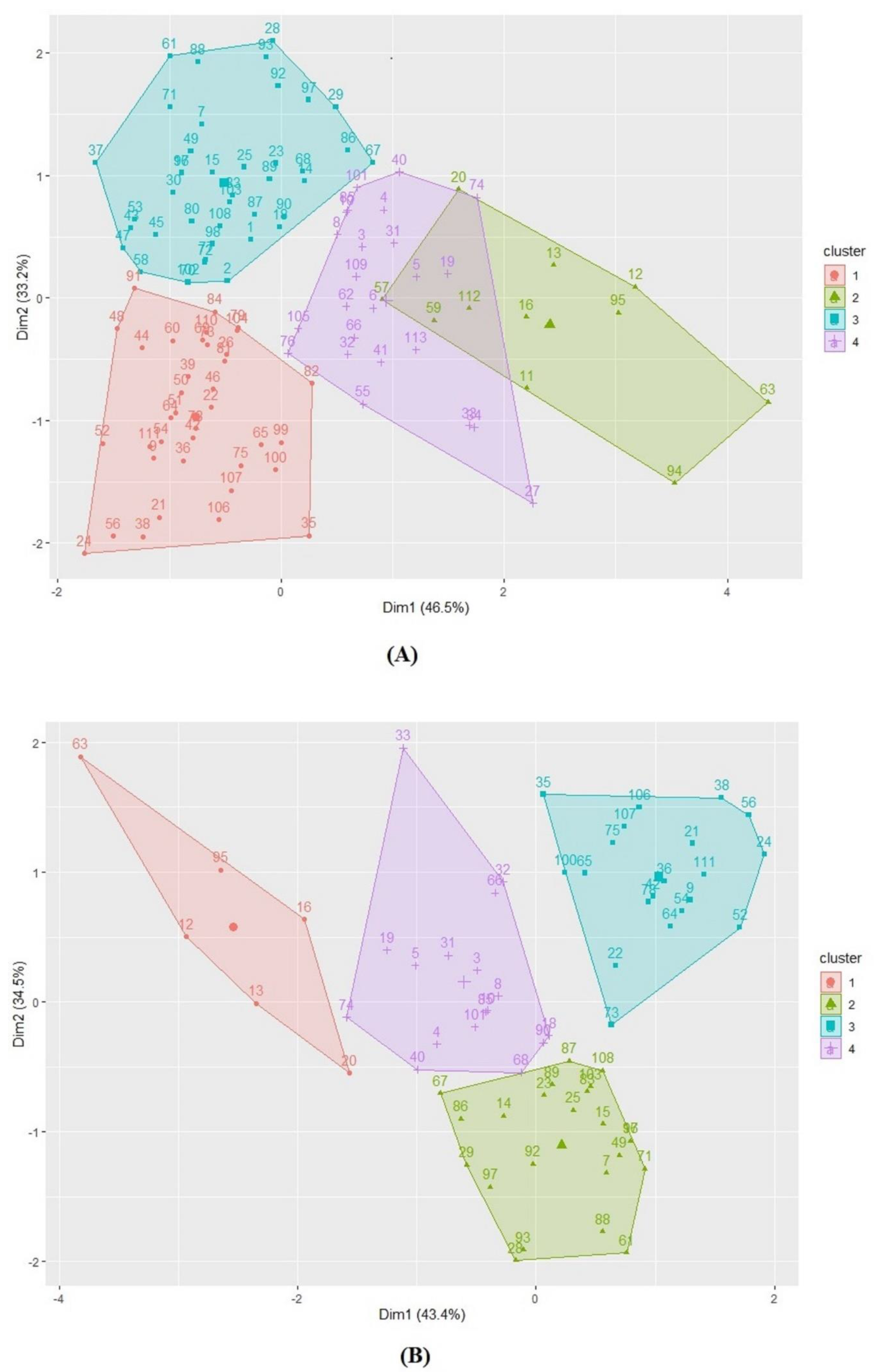
3.1. Reaction of Durum Wheat Accessions to Tan Spot
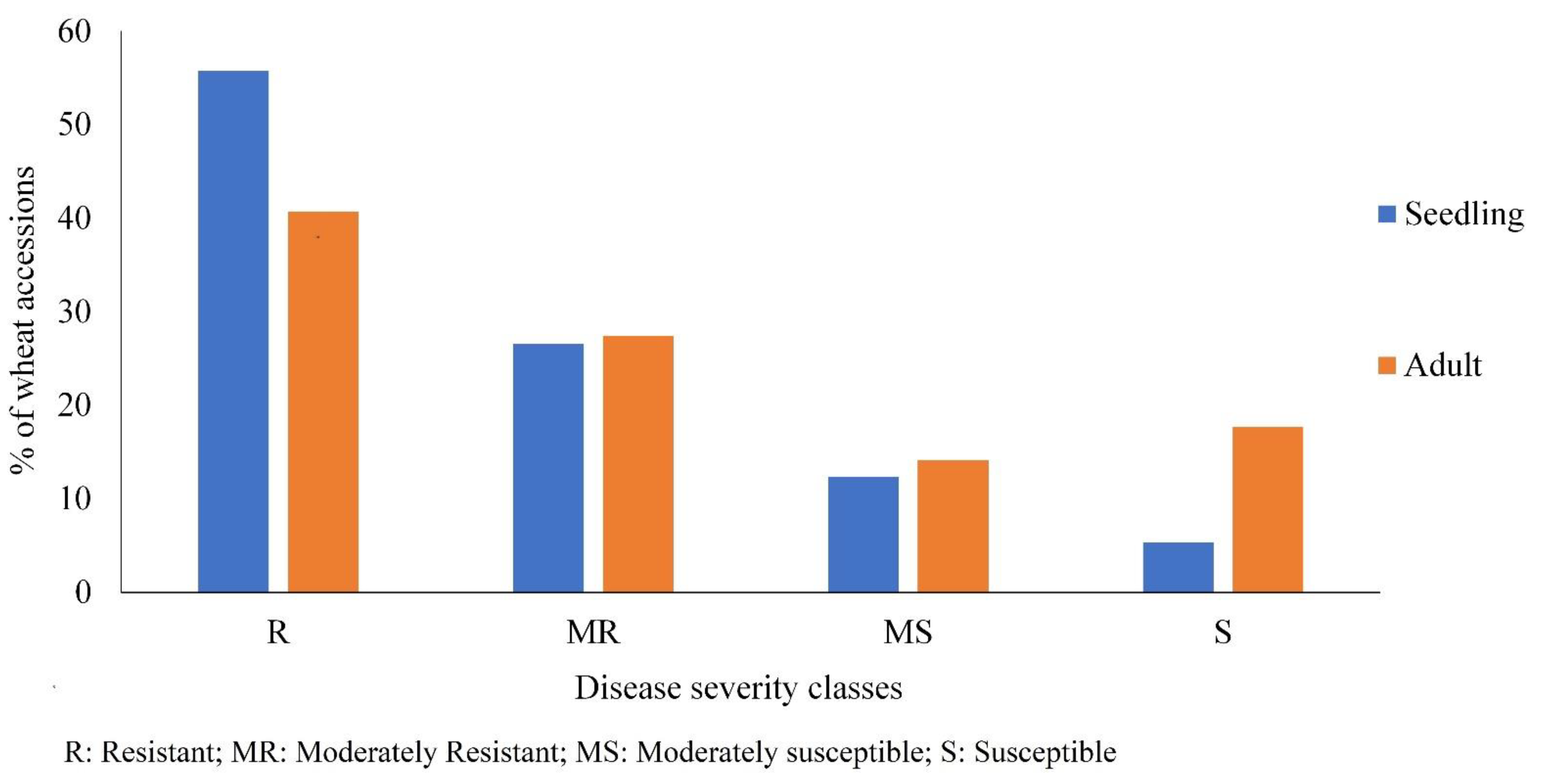
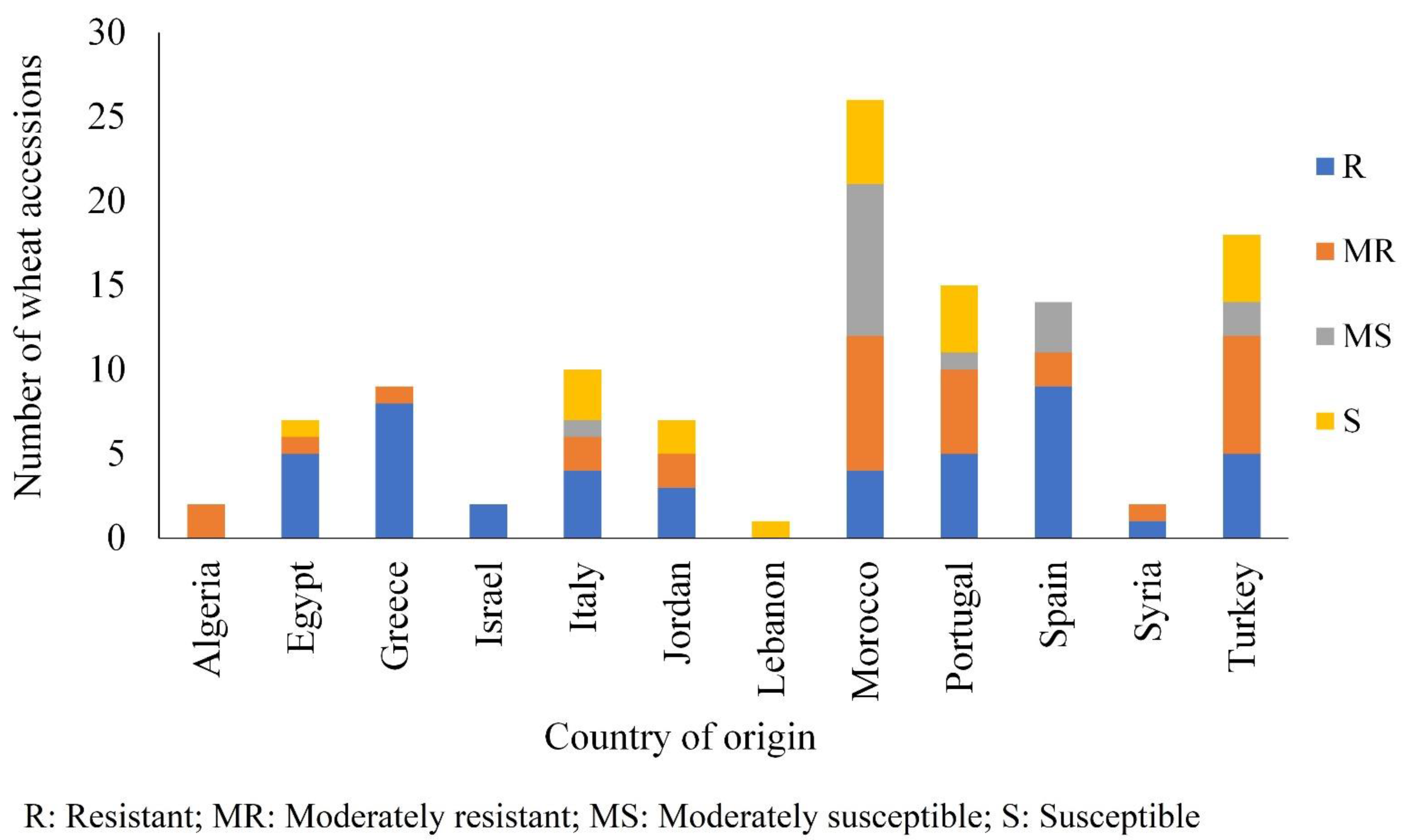
3.2. Comparison of Seedling and Adult Plant Resistance
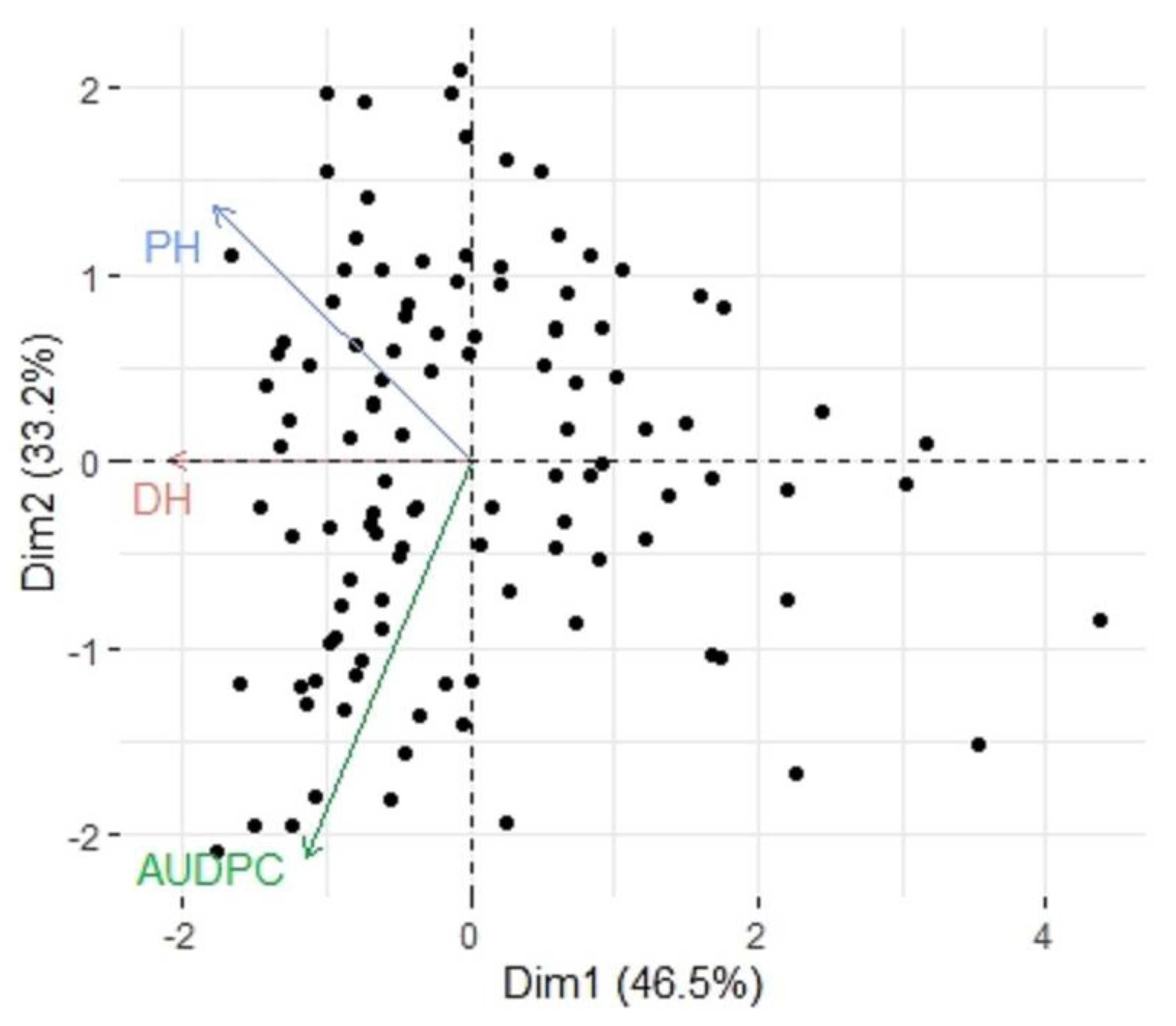
3.3. Correlation of Tan Spot Infection with Plant Height and Days to Heading
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fayaz, F.; Aghaee Sarbarzeh, M.; Talebi, R.; Azadi, A. Genetic Diversity and Molecular Characterization of Iranian Durum Wheat Landraces (Triticum turgidum durum (Desf.) Husn.) Using DArT Markers. Biochem Genet. 2019, 57, 98–116. [Google Scholar] [CrossRef]
- International Grains Council [IGC]. World Grain Statistics 2016. 2020. Available online: https://www.igc.int/en/subscriptions/subscription.aspx (accessed on 21 May 2020).
- Feldman, M. Origin of cultivated wheat. In The World Wheat Book: A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., Eds.; Lavoisier: Paris, France, 2001; pp. 3–56. [Google Scholar]
- Harlan, J.R. Agricultural Origins: Centers and Non centers. Science 1971, 174, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Bonjean, A.P.; Angus, W.J.; van Ginkel, M. The World Wheat Book: A History of Wheat Breeding; Lavoisier: Paris, France, 2016; Volume 3. [Google Scholar]
- Kabbaj, H.; Sall, A.T.; Al-Abdallat, A.; Geleta, M.; Amri, A.; Filali-Maltouf, A.; Belkadi, B.; Ortiz, R.; Bassi, F.M. Genetic Diversity within a Global Panel of Durum Wheat (Triticum durum) Landraces and Modern Germplasm Reveals the History of Alleles Exchange. Front. Plant. Sci. 2017, 8, 1277. [Google Scholar] [CrossRef] [Green Version]
- USDA. Tunisia: Grain and Feed Annual; USDA Foreign Agricultural Service: Washington, DC, USA, 2019.
- Chakraborty, S.; Tiedemann, A.V.; Teng, P.S. Climate Change: Potential Impact on Plant Diseases. Environ. Pollut. 2000, 108, 317–326. [Google Scholar] [CrossRef]
- Marzario, S.; Logozzo, G.; David, J.; Zeuli, P.; Gioia, T. Molecular Genotyping (SSR) and Agronomic Phenotyping for Utilization of Durum Wheat (Triticum durum Desf.) Ex Situ Collection from Southern Italy: A Combined Approach Including Pedigreed Varieties. Genes 2018, 9, 465. [Google Scholar] [CrossRef] [Green Version]
- Newton, A.C.; Johnson, S.N.; Gregory, P.J. Implications of Climate Change for Diseases, Crop Yields and Food Security. Euphytica 2011, 179, 3–18. [Google Scholar] [CrossRef]
- Ortiz, R.; Sayre, K.D.; Govaerts, B.; Gupta, R.; Subbarao, G.V.; Ban, T.; Hodson, D.; Dixon, J.M.; Iván Ortiz-Monasterio, J.; Reynolds, M. Climate Change: Can Wheat Beat the Heat? Agric. Ecosys. Environ. 2008, 126, 46–58. [Google Scholar] [CrossRef]
- Soriano, J.M.; Villegas, D.; Sorrells, M.E.; Royo, C. Durum Wheat Landraces from East and West Regions of the Mediterranean Basin Are Genetically Distinct for Yield Components and Phenology. Front. Plant. Sci. 2018, 9, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faris, J.D.; Liu, Z.; Xu, S.S. Genetics of Tan Spot Resistance in Wheat. Theor. Appl. Genet. 2013, 126, 2197–2217. [Google Scholar] [PubMed]
- Hosford, R.M. Tan spot: Developing knowledge 1902–1981, virulent races and wheat differentials, methodology, rating systems, other leaf diseases, literature. In Proceedings of the Tan Spot of Wheat and Related Disease Workshop, Fargo, North Dakota, 14–15 July 1981; pp. 1–20. [Google Scholar]
- Rees, R.G.; Platz, G.J. Tan spot and its control: Some Australian experiences. In Advances Tan Spot Research; ND Agricultural Experiment Station: Fargo, North Dakota, 1992. [Google Scholar]
- Kamel, S.; Cherif, M. Tan spot of wheat in Northern Tunisia: Distribution, prevalence, incidence and severity. Cereal Res. Commun. 2021, 49, 1–12. [Google Scholar] [CrossRef]
- De Wolf, E.D.; Effertz, R.J.; Ali, S.; Francl, L.J. Vistas of tan spot research. Can. J. Plant. Pathol. 1998, 20, 349–370. [Google Scholar]
- Singh, P.K.; Duveiller, E.; Singh, R.P. Resistance breeding for tan spot (Pyrenophora tritici-repentis) of wheat. In Disease Resistance in Wheat; Sharma, I., Ed.; CABI: Wallingford, UK, 2012; pp. 136–150. [Google Scholar]
- Lamari, L.; Bernier, C.C. Evaluation of Wheat Lines and Cultivars to Tan Spot [Pyrenophora tritici-repentis] Based on Lesion Type. Can. J. Plant. Pathol. 1989, 11, 49–56. [Google Scholar] [CrossRef]
- Eyal, Z. Integrated control of Septoria diseases of wheat. Plant. Dis. 1981, 65, 763–768. [Google Scholar] [CrossRef]
- Scott, P.R.; Benedikz, P.W.; Cox, C.J. A Genetic Study of the Relationship between Height, Time of Ear Emergence and Resistance to Septoria Nodorum in Wheat. Plant. Pathol. 1982, 31, 45–60. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Clarke, J.M.; DePauw, R.M. The Effect of Plant Height on Tan Spot on Durum Wheat in Southern Saskatchewan. Crop. Sci. 2002, 42, 159–164. [Google Scholar] [CrossRef]
- Elias, E.; Cantrell, R.G.; Hosford, R.M. Heritability of Resistance to Tan Spot in Durum Wheat and Its Association with Other Agronomic Traits. Crop. Sci. 1989, 29, 299. [Google Scholar] [CrossRef]
- Joshi, A.K.; Chand, R.; Arun, B. Relationship of plant height and days to maturity with resistance to spot blotch in wheat. Euphytica 2002, 123, 221–228. [Google Scholar]
- Simon, M.R.; Perelló, A.E.; Cordo, C.A.; Larrán, S.; van der Putten, P.E.; Struik, P.C. Association between Septoria tritici blotch, plant height, and heading date in wheat. Agron. J. 2005, 97, 1072–1081. [Google Scholar] [CrossRef]
- Fernandez, M.R.; DePauw, R.M.; Clarke, J.M.; Fox, S.L. Discoloration of Wheat Kernels by Pyrenophora tritici-repentis. Can. J. Plant. Pathol. 1998, 20, 380–383. [Google Scholar] [CrossRef]
- Schilder, A.M.C.; Bergstrom, G.C. Infection of Wheat Seed by Pyrenophora tritici-repentis. Can. J. Bot. 1994, 72, 510–519. [Google Scholar] [CrossRef]
- See, P.T.; Schultz, N.; Moffat, C.S. Evaluation of Pyrenophora tritici-repentis Infection of Wheat Heads. Agriculture 2020, 10, 417. [Google Scholar] [CrossRef]
- Aboukhaddour, R.; Cloutier, S.; Ballance, G.M.; Lamari, L. Genome Characterization of Pyrenophora tritici-repentis Isolates Reveals High Plasticity and Independent Chromosomal Location of ToxA and ToxB. Mol. Plant. Pathol. 2009, 10, 201–212. [Google Scholar] [CrossRef]
- Manning, V.A.; Pandelova, I.; Dhillon, B.; Wilhelm, L.J.; Goodwin, S.B.; Berlin, A.M.; Figueroa, M.; Freitag, M.; Hane, J.K.; Henrissat, B.; et al. Comparative Genomics of a Plant-Pathogenic Fungus, Pyrenophora tritici-repentis, Reveals Transduplication and the Impact of Repeat Elements on Pathogenicity and Population Divergence. G3 Genes Genomes Genet. 2013, 3, 41–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moolhuijzen, P.; See, P.T.; Hane, J.K.; Shi, G.; Liu, Z.; Oliver, R.P.; Moffat, C.S. Comparative Genomics of the Wheat Fungal Pathogen Pyrenophora tritici-repentis Reveals Chromosomal Variations and Genome Plasticity. BMC Genomics 2018, 19, 279. [Google Scholar] [CrossRef]
- Mundt, C.C. Use of multiline cultivars and cultivar mixtures for disease management. Annu. Rev. Phytopathol. 2002, 40, 381–410. [Google Scholar] [CrossRef] [Green Version]
- Mundt, C.C. Durable Resistance: A Key to Sustainable Management of Pathogens and Pests. Infect. Genet. Evol. 2014, 27, 446–455. [Google Scholar] [CrossRef]
- Strelkov, S.E.; Lamari, L. Host-Parasite Interactions in Tan Spot [Pyrenophora tritici-repentis] of Wheat. Can. J. Plant. Pathol. 2003, 25, 339–349. [Google Scholar] [CrossRef]
- Baloch, F.S.; Alsaleh, A.; Shahid, M.Q.; Çiftçi, V.; de Miera, L.E.S.; Aasim, M.; Nadeem, M.A.; Aktaş, H.; Özkan, H.; Hatipoğlu, R. A Whole Genome DArTseq and SNP Analysis for Genetic Diversity Assessment in Durum Wheat from Central Fertile Crescent. PLoS ONE 2017, 12, e0167821. [Google Scholar] [CrossRef] [Green Version]
- Galagedara, N.; Liu, Y.; Fiedler, J.; Shi, G.; Chiao, S.; Xu, S.S.; Faris, J.D.; Li, X.; Liu, Z. Genome-Wide Association Mapping of Tan Spot Resistance in a Worldwide Collection of Durum Wheat. Theor. Appl. Genet. 2020, 133, 2227–2237. [Google Scholar] [CrossRef] [PubMed]
- Gurung, S.; Mamidi, S.; Bonman, J.M.; Jackson, E.W.; del Río, L.E.; Acevedo, M.; Mergoum, M.; Adhikari, T.B. Identification of Novel Genomic Regions Associated with Resistance to Pyrenophora tritici-repentis Races 1 and 5 in Spring Wheat Landraces Using Association Analysis. Theor. Appl. Genet. 2011, 123, 1029. [Google Scholar] [CrossRef] [PubMed]
- Halder, J.; Zhang, J.; Ali, S.; Sidhu, J.S.; Gill, H.S.; Talukder, S.K.; Kleinjan, J.; Turnipseed, B.; Sehgal, S.K. Mining and Genomic Characterization of Resistance to Tan Spot, Stagonospora nodorum Blotch (SNB), and Fusarium Head Blight in Watkins Core Collection of Wheat Landraces. BMC Plant. Biol. 2019, 19, 480. [Google Scholar] [CrossRef]
- Singh, P.K.; Crossa, J.; Duveiller, E.; Singh, R.P.; Djurle, A. Association Mapping for Resistance to Tan Spot Induced by Pyrenophora tritici-repentis race 1 in CIMMYTs Historical Bread Wheat Set. Euphytica 2016, 207, 515–525. [Google Scholar] [CrossRef]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting Genetic Diversity from Landraces in Wheat Breeding for Adaptation to Climate Change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef] [PubMed]
- Mangini, G.; Margiotta, B.; Marcotuli, I.; Signorile, M.A.; Gadaleta, A.; Blanco, A. Genetic Diversity and Phenetic Analysis in Wheat (Triticum Turgidum Subsp. Durum and Triticum Aestivum Subsp. Aestivum) Landraces Based on SNP Markers. Genet. Resour. Crop. Evol. 2017, 64, 1269–1280. [Google Scholar] [CrossRef]
- Pignone, D.; de Paola, D.; Rapanà, N.; Janni, M. Single Seed Descent: A Tool to Exploit Durum Wheat (Triticum durum Desf.) Genetic Resources. Genet. Resour. Crop. Evol. 2015, 62, 1029–1035. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar]
- Saari, E.E.; Prescott, J.M. Scale for appraising the foliar intensity of wheat diseases. Plant. Dis. Report. 1975, 59, 377–380. [Google Scholar]
- Simko, I.; Piepho, H.-P. The Area under the Disease Progress Stairs: Calculation, Advantage, and Application. Phytopathology 2012, 102, 381–389. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 30 June 2021).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics; Fourth, S., Ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses; R Package Version 1.0.7; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://CRAN.R-project.org/package=factoextra (accessed on 1 April 2020).
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions; R Package Version 2.1.1; Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Evans, C.K.; Hunger, R.M.; Siegerist, W.C. Comparison of Greenhouse and Field Testing to Identify Wheat Resistant to Tan Spot. Plant. Dis. 1999, 83, 269–273. [Google Scholar] [CrossRef]
- Cox, D.J.; Hosford, R.M. Resistant winter wheats compared at differing growth stages and leaf positions for tan spot severity. Plant. Dis. 1987, 71, 883–886. [Google Scholar] [CrossRef]
- Tadesse, W.; Reents, H.J.; Hsam, S.L.K.; Zeller, F.J. Relationship of Seedling and Adult Plant Resistance and Evaluation of Wheat Germplasm against Tan Spot (Pyrenophora Tritici-Repentis). Genet. Resour. Crop. Evol. 2011, 58, 339–346. [Google Scholar] [CrossRef]
- Perello, A.; Moreno, V.; Simón, M.R.; Sisterna, M. Tan Spot of Wheat (Triticum aestivum L.) Infection at Different Stages of Crop Development and Inoculum Type. Crop. Prot. 2003, 22, 157–169. [Google Scholar] [CrossRef]
- Rees, R.G.; Platz, G.J. Effectiveness of incomplete resistance to Pyrenophora tritici-repentis in wheat. Aust. J. Agric. Res. 1989, 40, 43–48. [Google Scholar] [CrossRef]
- Gamba, F.M.; Lamari, L.; Brülé-Babel, A.L. Inheritance of Race-Specific Necrotic and Chlorotic Reactions Induced by Pyrenophora tritici-repentis in Hexaploid Wheats. Can. J. Plant. Pathol. 1998, 20, 401–407. [Google Scholar] [CrossRef]
- Gamba, F.M.; Lamari, L. Mendelian Inheritance of Resistance to Tan Spot [Pyrenophora tritici-repentis] in Selected Genotypes of Durum Wheat (Triticum turgidum). Can. J. Plant. Pathol. 1998, 20, 408–414. [Google Scholar] [CrossRef]
- Lamari, L.; Bernier, C.C. Genetics of tan necrosis and extensive chlorosis in tan spot of wheat caused by Pyrenophora tritici-repentis. Phytopathology 1991, 81, 1092–1095. [Google Scholar] [CrossRef]
- Singh, P.K.; Hughes, G.R. Genetic Similarity among Isolates of Pyrenophora tritici-repentis, causal agent of tan spot of wheat. J. Phytopathol. 2006, 154, 178–184. [Google Scholar] [CrossRef]
- Singh, P.K.; Mergoum, M.; Gonzalez-Hernandez, J.L.; Ali, S.; Adhikari, T.B.; Kianian, S.F.; Elias, E.M.; Hughes, G.R. Genetics and Molecular Mapping of Resistance to Necrosis Inducing Race 5 of Pyrenophora tritici-repentis in Tetraploid Wheat. Mol. Breed. 2008, 21, 293–304. [Google Scholar] [CrossRef]
- Chu, C.-G.; Chao, S.; Friesen, T.L.; Faris, J.D.; Zhong, S.; Xu, S.S. Identification of Novel Tan Spot Resistance QTLs Using an SSR-Based Linkage Map of Tetraploid Wheat. Mol. Breed. 2010, 25, 327–338. [Google Scholar] [CrossRef]
- Chu, C.-G.; Friesen, T.L.; Xu, S.S.; Faris, J.D. Identification of Novel Tan Spot Resistance Loci beyond the Known Host-Selective Toxin Insensitivity Genes in Wheat. Theor. Appl. Genet. 2008, 117, 873–881. [Google Scholar] [CrossRef]
- Faris, J.D.; Overlander, M.E.; Kariyawasam, G.K.; Carter, A.; Xu, S.S.; Liu, Z. Identification of a Major Dominant Gene for Race-Nonspecific Tan Spot Resistance in Wild Emmer Wheat. Theor. Appl. Genet. 2020, 133, 829–841. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D. Molecular Mapping of Resistance to Pyrenophora tritici-repentis Race 5 and Sensitivity to Ptr ToxB in Wheat. Theor. Appl. Genet. 2004, 109, 464–471. [Google Scholar] [CrossRef]
- Tadesse, W.; Hsam, S.L.K.; Wenzel, G.; Zeller, F.J. Chromosome Location of a Gene Conferring Resistance to Pyrenophora tritici-repentis in Ethiopian Wheat Cultivars. Euphytica 2008, 162, 423–430. [Google Scholar] [CrossRef]
- Tadesse, W.; Schmolke, M.; Hsam, S.L.K.; Mohler, V.; Wenzel, G.; Zeller, F.J. Molecular Mapping of Resistance Genes to Tan Spot [Pyrenophora tritici-repentis race 1] in Synthetic Wheat Lines. Theor. Appl. Genet. 2007, 114, 855–862. [Google Scholar] [CrossRef]
- Tadesse, W.; Hsam, S.L.K.; Wenzel, G.; Zeller, F.J. Identification and Monosomic Analysis of Tan Spot Resistance Genes in Synthetic Wheat Lines (Triticum Turgidum L. × Aegilops Tauschii Coss.). Crop. Sci. 2006, 46, 1212–1217. [Google Scholar] [CrossRef]
- Dinglasan, E.G.; Peressini, T.; Marathamuthu, K.A.; See, P.T.; Snyman, L.; Platz, G.; Godwin, I.; Voss-Fels, K.P.; Moffat, C.S.; Hickey, L.T. Genetic Characterization of Adult-Plant Resistance to Tan Spot (Syn, Yellow Spot) in Wheat. Theor. Appl. Genet. 2021, 134, 2823–2839. [Google Scholar] [CrossRef] [PubMed]
- Dinglasan, E.G.; Godwin, I.D.; Phan, H.T.T.; Tan, K.-C.; Platz, G.J.; Hickey, L.T. Vavilov Wheat Accessions Provide Useful Sources of Resistance to Tan Spot (Syn. Yellow Spot) of Wheat. Plant. Pathol. 2018, 67, 1076–1087. [Google Scholar] [CrossRef]
- Dinglasan, E.; Godwin, I.D.; Mortlock, M.Y.; Hickey, L.T. Resistance to Yellow Spot in Wheat Grown under Accelerated Growth Conditions. Euphytica 2016, 209, 693–707. [Google Scholar] [CrossRef]
- Kokhmetova, A.; Sehgal, D.; Ali, S.; Atishova, M.; Kumarbayeva, M.; Leonova, I.; Dreisigacker, S. Genome-Wide Association Study of Tan Spot Resistance in a Hexaploid Wheat Collection from Kazakhstan. Front. Genet. 2021, 11, 581214. [Google Scholar] [CrossRef]
- Laribi, M.; Akhavan, A.; M’Barek, S.; Yahyaoui, A.; Strelkov, S.E.; Sassi, K. Characterization of Pyrenophora tritici-repentis in Tunisia and Comparison with a Global Pathogen Population. Plant. Dis. 2021, 107, 2411–2502. [Google Scholar] [CrossRef]
- Laribi, M.; Gamba, F.M.; Hassine, M.; Singh, P.K.; Yahyaoui, A.; Sassi, K. Race Structure and Distribution of Pyrenophora tritici-repentis in Tunisia. Phytopathol. Mediterr. 2019, 58, 473–483. [Google Scholar] [CrossRef]
- Kamel, S.; Cherif, M.; Hafez, M.; Despins, T.; Aboukhaddour, R. Pyrenophora tritici–repentis in Tunisia: Race Structure and Effector Genes. Front. Plant. Sci. 2019, 10, 1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; El-Basyoni, I.; Kariyawasam, G.; Zhang, G.; Fritz, A.; Hansen, J.; Marais, F.; Friskop, A.; Chao, S.; Akhunov, E.; et al. Evaluation and Association Mapping of Resistance to Tan Spot and Stagonospora Nodorum Blotch in Adapted Winter Wheat Germplasm. Plant. Dis. 2015, 99, 1333–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- German, S.E.; Kolmer, J.A. Effect of Gene Lr34 in the Enhancement of Resistance to Leaf Rust of Wheat. Theor. Appl. Genet. 1992, 84, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Foessel, S.A.; Singh, R.P.; Lillemo, M.; Huerta-Espino, J.; Bhavani, S.; Singh, S.; Lan, C.; Calvo-Salazar, V.; Lagudah, E.S. Lr67/Yr46 Confers Adult Plant Resistance to Stem Rust and Powdery Mildew in Wheat. Theor. Appl. Genet. 2014, 127, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, A.; Gilbert, B.; Boni, R.; Krattinger, S.G.; Singh, D.; Park, R.F.; Lagudah, E.; Ayliffe, M. The Lr34 Adult Plant Rust Resistance Gene Provides Seedling Resistance in Durum Wheat without Senescence. Plant. Biotechnol. J. 2017, 15, 894–905. [Google Scholar] [CrossRef] [Green Version]
- Bansal, U.K.; Kazi, A.G.; Singh, B.; Hare, R.A.; Bariana, H.S. Mapping of Durable Stripe Rust Resistance in a Durum Wheat Cultivar Wollaroi. Mol. Breed. 2014, 33, 51–59. [Google Scholar] [CrossRef]
- Nelson, R.; Wiesner-Hanks, T.; Wisser, R.; Balint-Kurti, P. Navigating Complexity to Breed Disease-Resistant Crops. Nat. Rev. Genet. 2018, 19, 21–33. [Google Scholar] [CrossRef]
- Parlevliet, J.E. What is durable resistance, a general outline. In Durability of Disease Resistance; Current Plant Science and Biotechnology in Agriculture; Jacobs, T., Parlevliet, J.E., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; Volume 18, pp. 23–39. ISBN 978-94-010-4885-9. [Google Scholar]
- Hotelling, H. Analysis of a Complex of Statistical Variables into Principal Components. J. Educ. Psychol. 1933, 24, 417–441. [Google Scholar] [CrossRef]
- Jackson, J.E. A User’s Guide to Principal Components; John Wiley and Sons: Hoboken, NJ, USA, 2005; ISBN 978-0-471-72532-9. [Google Scholar]
- Li, H.B.; Yan, W.; Liu, G.R.; Wen, S.M.; Liu, C.J. Identification and Validation of Quantitative Trait Loci Conferring Tan Spot Resistance in the Bread Wheat Variety Ernie. Theor. Appl. Genet. 2011, 122, 395–403. [Google Scholar] [CrossRef]
- Muqaddasi, Q.H.; Kamal, R.; Mirdita, V.; Rodemann, B.; Ganal, M.W.; Reif, J.C.; Röder, M.S. Genome-Wide Association Studies and Prediction of Tan Spot (Pyrenophora tritici-repentis) Infection in European Winter Wheat via Different Marker Platforms. Genes 2021, 12, 490. [Google Scholar] [CrossRef] [PubMed]
- Kollers, S.; Rodemann, B.; Ling, J.; Korzun, V.; Ebmeyer, E.; Argillier, O.; Hinze, M.; Plieske, J.; Kulosa, D.; Ganal, M.W.; et al. Genome-Wide Association Mapping of Tan Spot Resistance (Pyrenophora tritici-repentis) in European Winter Wheat. Mol. Breed. 2014, 34, 363–371. [Google Scholar] [CrossRef]
- Kollers, S.; Rodemann, B.; Ling, J.; Korzun, V.; Ebmeyer, E.; Argillier, O.; Hinze, M.; Plieske, J.; Kulosa, D.; Ganal, M.W.; et al. Whole Genome Association Mapping of Fusarium Head Blight Resistance in European Winter Wheat (Triticum aestivum L.). PLoS ONE 2013, 8, e57500. [Google Scholar] [CrossRef] [Green Version]
- Spanic, V.; Lemmens, M.; Drezner, G.; Dvojkovic, K. Interrelations between height of winter wheat genotypes and resistance to Fusarium head blight (fhb). Rom. Agric. Res. 2011, 28, 43–48. [Google Scholar]
- Mao, S.L.; Wei, Y.M.; Cao, W.; Lan, X.J.; Yu, M.; Chen, Z.M.; Chen, G.Y.; Zheng, Y.L. Confirmation of the Relationship between Plant Height and Fusarium Head Blight Resistance in Wheat (Triticum aestivum L.) by QTL Meta-Analysis. Euphytica 2010, 174, 343–356. [Google Scholar] [CrossRef]
- Wicki, W.; Winzeler, M.; Schmid, J.E.; Stamp, P.; Messmer, M. Inheritance of Resistance to Leaf and Glume Blotch Caused by Septoria Nodorum Berk in Winter Wheat. Theor. Appl. Genet. 1999, 99, 1265–1272. [Google Scholar] [CrossRef]
- Tavella, C.M. Date of Heading and Plant Height of Wheat Varieties, as Related to Septoria Leaf Blotch Damage. Euphytica 1978, 27, 577–580. [Google Scholar] [CrossRef]
- Danon, T.; Sacks, J.M.; Eyal, Z. The relationships among plant stature, maturity class, and susceptibility to Septoria leaf blotch of wheat. Phytopathology 1982, 72, 1037–1042. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance | ||
 |  |  |
| Score 0: No lesion present | Score 1: Small, dark brown to black singular spots lacking any distinctive chlorotic or necrotic halo | Score 2: Small dark brown to black spots with small chlorotic halo Some necrosis |
| Moderate resistance | Moderate susceptibility | Susceptibility |
 |  |  |
| Score 3: Small to medium oval shaped lesion surrounded by a chlorotic halo | Score 4: Lesions with distinctive necrotic or chlorotic halo. Main lesion coalescing with surrounding singular lesions | Score 5: Large oval shaped lesions with the main lesion coalescing with most surrounding singular lesions |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laribi, M.; Ben M’Barek, S.; Fakhfakh, M.; Yahyaoui, A.H.; Sassi, K. Durum Wheat Mediterranean Landraces: A Valuable Source for Resistance to Tan Spot Disease. Agriculture 2021, 11, 1148. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111148
Laribi M, Ben M’Barek S, Fakhfakh M, Yahyaoui AH, Sassi K. Durum Wheat Mediterranean Landraces: A Valuable Source for Resistance to Tan Spot Disease. Agriculture. 2021; 11(11):1148. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111148
Chicago/Turabian StyleLaribi, Marwa, Sarrah Ben M’Barek, Moez Fakhfakh, Amor Hassine Yahyaoui, and Khaled Sassi. 2021. "Durum Wheat Mediterranean Landraces: A Valuable Source for Resistance to Tan Spot Disease" Agriculture 11, no. 11: 1148. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111148